
")


")





. Карбоксилирование рибулозобисфосфата")





























 Биология
БиологияПохожие презентации:










Физиология растений. Лекция 3
1. Лекция 3
Физиология растений2. Фотосинтетическое фосфорилирование (ФФ)
ФФ – это синтез молекулы АТФ засчет энергии трансмембранного
потенциала (ΔμН+) тилакоидной
мембраны.
ΔμН+
АДФ + Фн
АТФ + Н2О
3. Типы фотофосфорилирования
Нециклическоефотофосфорилирование.
2АДФ + Фн + 2НАДФ+ + 2 Н2О
2АТФ +
+ О2 + 2НАДФН
Осуществляют фотосистемы ФСI и ФСII.
2. Циклическое фотофосфорилирование
АДФ + Фн
АТФ + Н2О
Осуществляет ФСI.
1.
4. Механизм фотосинтетического фосфорилирования
5. Хемио-осмотическая гипотеза (Митчелл, Ягендорф)
6. Структура АТФ-синтазы
7. Биохимия фотосинтеза
Сущность темновой фазыфотосинтеза заключается в
трансформации
макроэргической связи АТФ и
НАДФН в стабильную форму
энергии – органическую связь.
СО2 + АТФ + НАДФН
[СН2О] + АДФ + Фн + НАДФ
8. Цикл Кальвина
В 1957 году Кальвин предложилбиохимический механизм превращения
СО2 в фосфоглицериновый альдегид
(ФГА).
1 сек
*СО2
СН2ОН-СНОН-*СООН
ФГК
Это С3-путь или восстановительный
пентозофосфатный путь (ВПФП).
9. Методы Кальвина
В лаборатории Кальвина, Басхэма иБенсона впервые использовали методы:
1. Для фотосинтеза использовали *СО2 с
радиоактивным углеродом – С14.
2. Фиксация растительного образца
кипящим этиловым спиртом.
3. Бумажная хроматография.
4. Радиоавтография, необходимая
для идентификации изотопных
молекул.
5. Фотостатированная камера
(фитотрон).
10. Цикл Кальвина
I.Включение СО2 в цикл
1) Активирование первичного акцептора
(рибулозомонофосфат)
РМФ + АТФ → РБФ + АДФ
РБФ – рибулозо-1,5бисфосфат
11. 2). Карбоксилирование рибулозобисфосфата
12. Свойства рибулозобисфосфаткарбоксилазы
РБФ-карбоксилаза (Rubisco)КФ 4.1.1.39 – самый
распространенный белок биосферы:
а) 50% от общего количества белков в
хлоропласте;
б) 70% составляют растительные белки в
биосфере;
в) 20% всех белков биосферы являются
РБФ-карбоксилазой, т.е. каждый
пятый белок в природе это Rubisco.
1.
13. Свойства Rubisco
2. Фермент двойного кодирования.Rubisco имеет молекулярную массу
550 кДа. Состоит из 8 больших (L)
c Mr 51-58 кДа
и 8 малых (S) с Mr 12-18 кДа.
Формула комплекса L8-S8.
Большие субъединицы кодируются
хлоропластной ДНК (rbcL), малые
ядерные ДНК (rbcS).
14. Свойства Rubisco
Сборка белка происходит в хлоропластахпри участии белка шаперона (Hsc60),
который кодируется в ядре. Для сборки
функционально активного фермента
необходима четкая регуляция работы
двух геномов (ядерного и
хлоропластного).
Каталитический центр фермента
формируют аминокислотные остатки Cи N-концов полипептидов двух больших
субъединиц. У Rubisco высших растений
имеется 8 каталитических центров.
15. Свойства Rubisco
Димеры больших субъединицкаталитически активны и в отсутствие
малых субъединиц. Малые субъединицы
играют регуляторную роль.
3. Бифункциональность РБФкарбоксилазы. Фермент катализирует
карбоксилазную активность
(функционирование ВПФП) и
оксигеназную (функционирование
фотодыхательного метаболизма).
16. Рибулозо-1,5-бисфосфаткарбоксилаза
17. II. Фаза восстановления
Осуществляется с помощью двухреакций:
А) активирование 3ФГК:
3ФГК + АТФ → 1,3ФГК + АДФ
(фермент киназа)
Б) восстановление ФГК до ФГА
1,3ФГК + НАДФН → ФГА + НАДФ+
(фермент
глицероальдегиддегидрогеназа)
18. III. Регенерация первичного акцептора
Общее уравнение образования ФГА3 РМФ + 9 АТФ + 6 НАДФН + 3 СО2 →
6 ФГА + 9 АДФ + 9 Фн + 6 НАДФ
5 ФГА
1 ФГА
на регенерацию
на синтез оргвеществ
первичного
акцептора
19. III.Регенерация первичного акцептора
С3 – фосфоглицериновыйальдегид (ФГА)
С6 – фруктозо-1,6-БФ
С4 – эритрозо-4-Ф
С7 – седогептулоза-7-Ф
С5 – рибулозомонофосфат
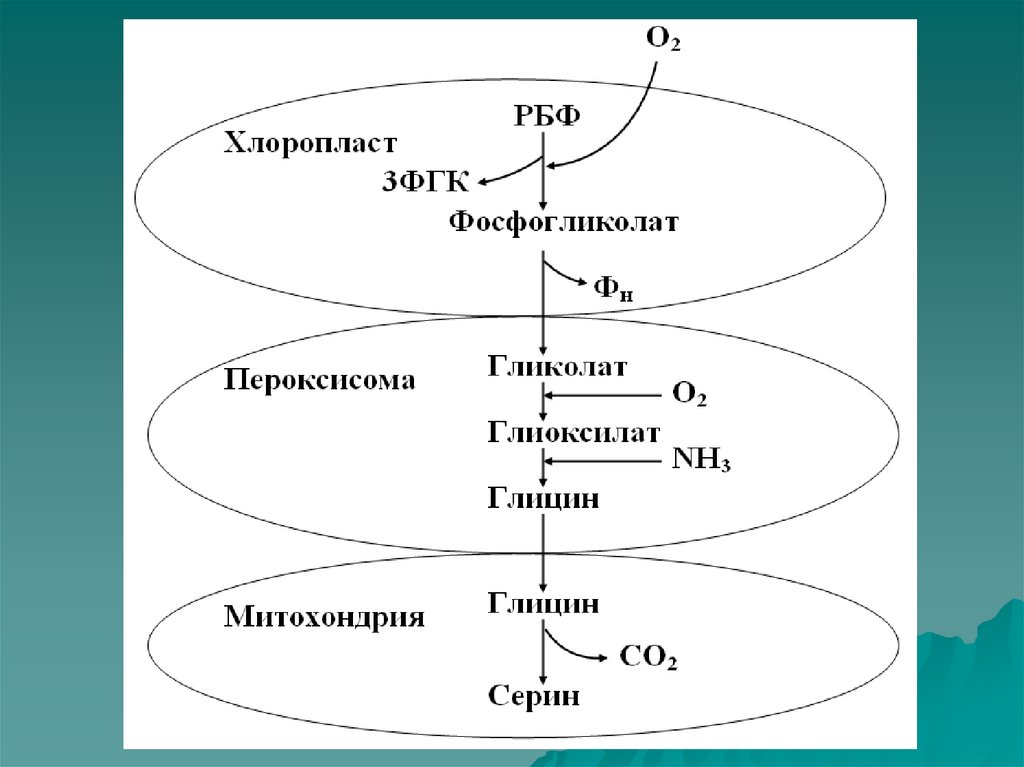
20. Фотодыхание
Фотодыхание – этосветоиндуцируемое выделение
углекислого газа зелеными
листьями растений.
Фотодыхание обнаружено в 1965
году. Его осуществляет комплекс
органоидов: хлоропласт,
пероксисома, митохондрия.
21.
22. Причины фотодыхания
1. Наличие оксигеназной функции урибулозобисфосфаткарбоксилазы.
2. Высокая концентрация кислорода
в атмосфере, составляющая 21%.
3. Значительные количества
кислорода, проникающие в ткань
растений, индуцируют
оксигеназную функцию Rubisco.
23. Значение фотодыхания
На фотодыхательный метаболизм у С3растений тратится до 55% продуктовфотосинтеза.
1. Фотодыхание выступает как футильный
цикл, обеспечивающий защиту
органических веществ (ДНК-белков) от
фотодеструкции.
2. Образование протеиногенных
аминокислот (глицина и серина).
3. Обеспечивает синтез сахарозы (с
небольшой скоростью).
24. Значение фотодыхания
Фотодыхание – это «расплата»растений за созданную ими
кислородную атмосферу.
Потеря 40-50% продуктов
фотосинтеза является необходимой
«данью» для выживания в
условиях кислородного стресса.
25. Цикл Хэтча-Слэйка
Карпилов И.С. в 1963 году обнаружилвключение 14СО2 в яблочную кислоту в
листьях кукурузы.
Коржик в 1964 году установил в сахарном
тростнике функционирование ФЕПкарбоксилазы.
Хэтч и Слэйк в 1969 году создали цикл
ассимиляции СО2 по С4-типу
фотосинтеза.
26. Анатомия листа у С4 - растений
27. Анатомия листа у С4 - растений
Для С4-растений характерна кранцанатомия листа.Внутреннее кольцо (обкладка)
изолировано клетками мезофилла.
В клетках обкладки локализован
цикл Кальвина.
В мезофильной ткани протекает
специфический С4-путь
ассимиляции углерода.
28. Биохимия цикла Хэтча-Слейка
1 – ФЕП-карбоксилаза, 2 – малатдегидрогеназа (НАД-зависимая),3 – малатдегидрогеназа (декарбоксилир.), 4 - пируваткиназа
29. Физиологическая роль цикла Хэтча-Слэйка
Физиологическое значение цикла:1) Увеличение концентрации СО2 в
клетках обкладки.
Используется новый фермент ФЕПкарбоксилаза, обладающий высоким
сродством к углекислому газу.
2) Рибулозо-бисфосфат-карбоксилаза
«спрятана» в клетках обкладки, где
концентрация кислорода минимальна.
Фотодыхание отсутствует или слабо
выражено.
30. Физиологическая роль цикла Хэтча-Слэйка
3) ФЕП-карбоксилаза являетсяуникальным ферментом,
обладающим большим сродством к
СО2, что имеет важное значение в
условиях низкой концентрации СО2
в атмосфере (0,035 %).
4) ФЕП-карбоксилаза является
термофильным ферментом (оптимум
t = 30-40оС, что обеспечивает
эффективное карбоксилирование
при высоких температурах.
31. Эволюция фотосинтеза
Причины эволюции в последние 600 млн. летобусловлены резким изменением факторов
внешней среды.
1. Концентрация кислорода выросла с 1% до
21%.
2. Количество углекислого газа уменьшилось с
10% до 0,03% (углерод использовался на
синтез органических веществ).
3. Нарушение гидробиологического режима.
Резкое уменьшение содержания воды в
биосфере.
Ксероморфогенез – основной путь эволюции.
32. Метаболизм органических кислот по типу толстянковых
Растения, осуществляющиеметаболизм по типу толстянковых,
являются суккулентами.
Большинство принадлежат к
сем.Crassulaceae, отсюда САМметаболизм. Представители:
колланхое, седум и др.
33. САМ-метаболизм
Данные растения адаптировались кдефициту воды на биохимическом
уровне. Днем устьица у этих растений
закрываются, чтобы снизить потерю
воды. Ночью у них устьица
открываются.
Следовательно, СО2 может проникать в
ткани только ночью и накапливаться в
виде органических кислот.
34. САМ-метаболизм
35. Сущность САМ-метаболизма
1. САМ-фотосинтез осуществляет разделениеассимиляции СО2 и цикла Кальвина не в
пространстве, а во времени.
2. Первичная фиксация СО2 осуществляется в
темноте, где ФЕП-карбоксилаза присоединяет
его к ФЕПу.
3. Днем устьица закрыты (предотвращение
потери воды), и проникновение СО2
невозможно. Малат служит источником
эндогенного СО2 (декарбоксилирование).
4. Образовавшийся СО2 диффундирует в
хлоропласты и включается в цикл Кальвина.
36. Сравнительная характеристика основных показателей С3-, С4- и САМ-растений
ПоказателиС3-
С4-
САМ
Транспирационный
коэффициент, мл
550-900
250-300
50-100
Оптимальная t oC
15-20
25-35
15-40
Углекислотный
компенсационный пункт, %
Интенсивность
фотосинтеза
мкМ СО2/мг
хл.час
0,0003
0,00001
100-200
200-400
30-80
37. Фотоокисление воды
До 1931 года доминировалаформальдегидная гипотеза
происхождения кислорода.
СО2
С + О2
С + Н2О
СН2О
То есть, кислород образовывался по
формальдегидной гипотезе из
углекислого газа.
38. Происхождение кислорода
Ван-Ниль в 1931 году предположил,что кислород образуется из воды.
СО2 + Н218О
СН2О + 18О2
18О – изотоп кислорода.
С помощью изотопа кислорода 18О
была доказана справедливость
этого утверждения.
39. Механизм выделения кислорода
Согласно Кутюрину (1968), молекулакислорода образуется из воды,
в окислении которой участвует
непосредственно хлорофилл.
Современная гипотеза предполагает
участие марганец-содержащего
фермента в окислении воды и
формировании молекулы
кислорода.