


















, увели")

")


 Биология
БиологияПохожие презентации:

")








Регуляция и саморегуляция системы дыхания
1.
ФИЗИОЛОГИЯЛекция № 9
• РЕГУЛЯЦИЯ И САМОРЕГУЛЯЦИЯ
СИСТЕМЫ ДЫХАНИЯ
2. План лекции
Понятие о регуляции дыхательной функции. Внешний ивнутренний контуры регуляции.
2. Нервная регуляция дыхательной функции:
2.1. Дыхательный центр, уровни его локализации.
Автоматия дыхательного центра
2.2. Рецептивные поля дыхательных рефлексов:
рецепторы растяжения лёгких,
ирритантные рецепторы,
юкстаальвеолярные рецепторы (J-рецепторы).
2.3. Гуморальная регуляция дыхательной функции.
Изменение газового состава крови – основной фактор
гуморальной регуляции дыхания.
2.4. Механизм первого вдоха новорожденных.
1.
3.
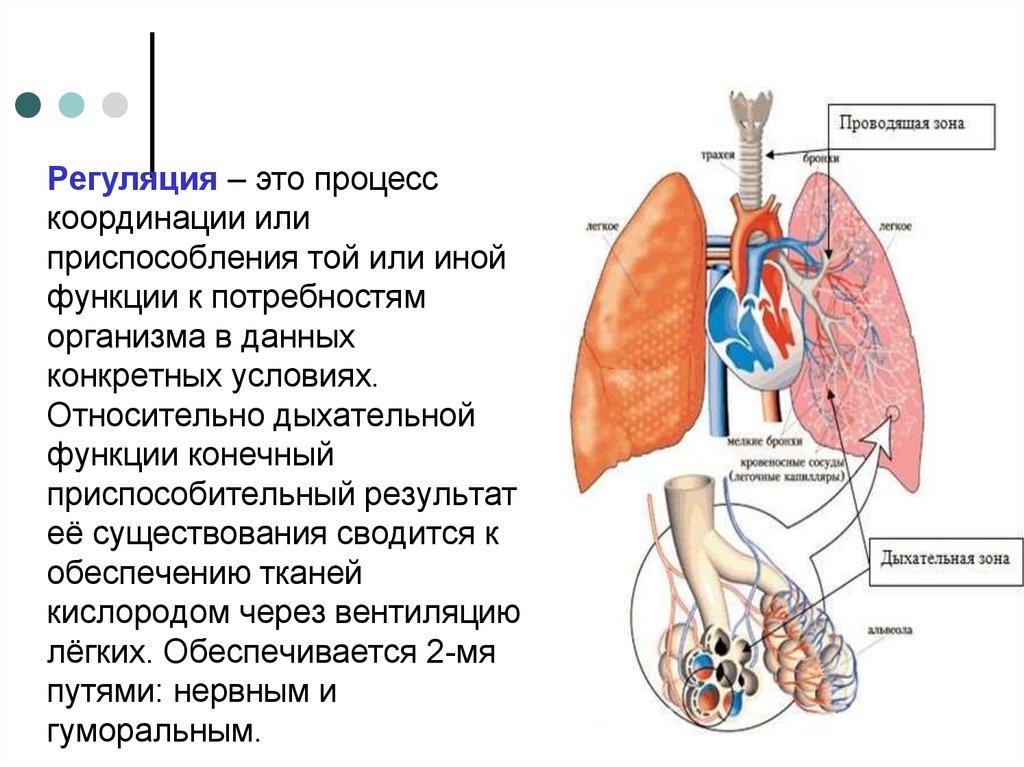
Регуляция – это процесскоординации или
приспособления той или иной
функции к потребностям
организма в данных
конкретных условиях.
Относительно дыхательной
функции конечный
приспособительный результат
её существования сводится к
обеспечению тканей
кислородом через вентиляцию
лёгких. Обеспечивается 2-мя
путями: нервным и
гуморальным.
4.
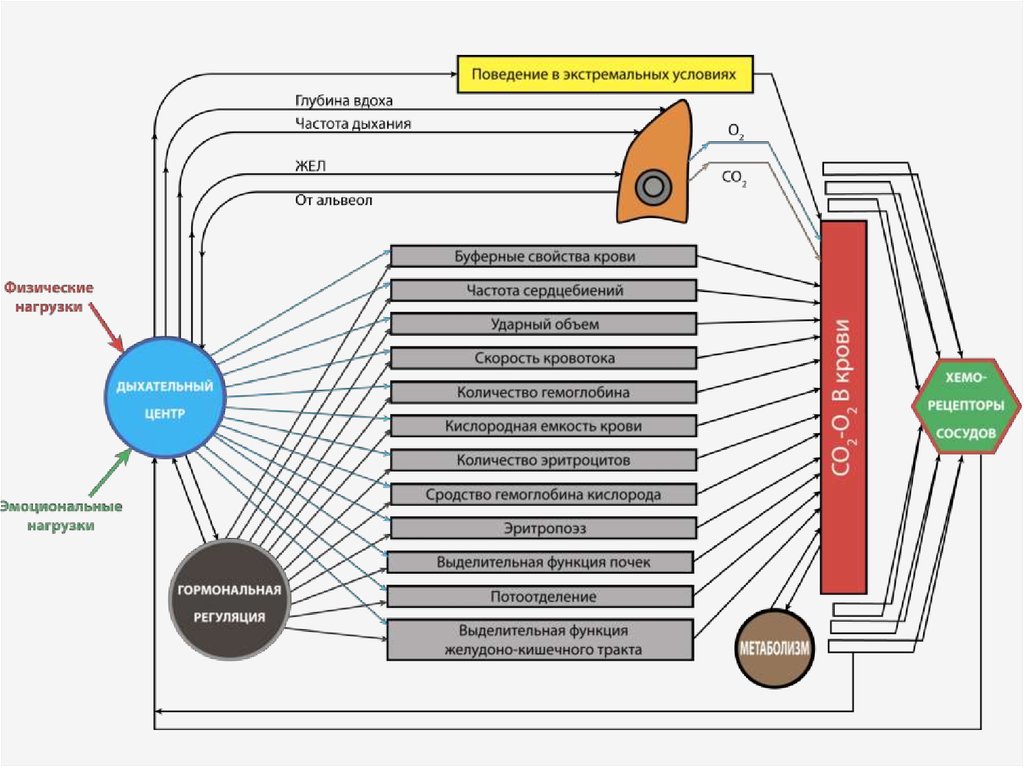
5. 1. Нервная регуляция дыхательной функции.
В основе нервной регуляции дыхательнойфункции лежит наличие структурнофункциональной организации
дыхательного центра. Под нервным
дыхательным центром понимают
совокупность нейронов, расположенных на
различных уровнях ЦНС, обеспечивающих
дыхательную функцию организма и ее
регуляцию.
6. История открытия дыхательного центра.
1. В 1812 г. Легаллуа методом перерезки головного мозга у птиц открыл впродолговатом мозге центр жизненного обеспечения дыхательной функции.
2. В 1942 г. Флуранс с помощью методики раздражения или удаления участков
головного мозга на бульбарном уровне подтвердил наличие центра жизненного
обеспечения дыхательной функции.
3. В 1885 г. Н.А. Миславский применил методику точечного раздражения или
удаления участков продолговатого мозга и открыл точную локализацию центра
регуляции дыхания и назвал его «дыхательным центром», определив отделы
центра как совокупность нейронов инспираторных (каудальные отделы
одиночного пучка, дно IV желудочка, в районе обекса, дорсальное ядро
продолговатого мозга) и экспираторных (в вентральном ядре продолговатого
мозга), и назвал их, соответственно, центр вдоха и центр выдоха.
+
−
ИЦ
ЭЦ
−
+
Дыхательный центр является парным
образованием, и каждая его половина
иннервирует аппарат внешнего дыхания
противоположной стороны тела,
обеспечивая координированную
регуляцию дыхательных движений.
7. В 1956 г. Баумгартен доказал, что четкого разделения между нейронами инспираторного и экспираторного центра не существует, но есть участки с
++
ИЦ
+
−
В 1956 г. Баумгартен доказал, что четкого разделения между
нейронами инспираторного и экспираторного центра не
существует, но есть участки с их наибольшим и наименьшим
представительством.
Лумсден установил, что кроме
.
продолговатого мозга дыхательные
нейроны расположены в области
ПТЦ
варолиевого моста. Это образование было
названо им пневмотаксическим центром
(ПТЦ). ПТЦ является своеобразным
+
+
«реле», физиологическое значение
ЭЦ
которого сводится к переключению
возбуждения с инспираторного центра на
−
экспираторный центр в тот момент, когда
глубина вдоха достигает нужной величины,
и наоборот. Именно участие ПТЦ
+
варолиевого моста обеспечивает
длительность реципрокного торможения
между ИЦ и ЭЦ бульбарного отдела ДЦ. На
современном этапе развития медицины
считается, что структура ДЦ включает в
себя различные структуры ЦНС.
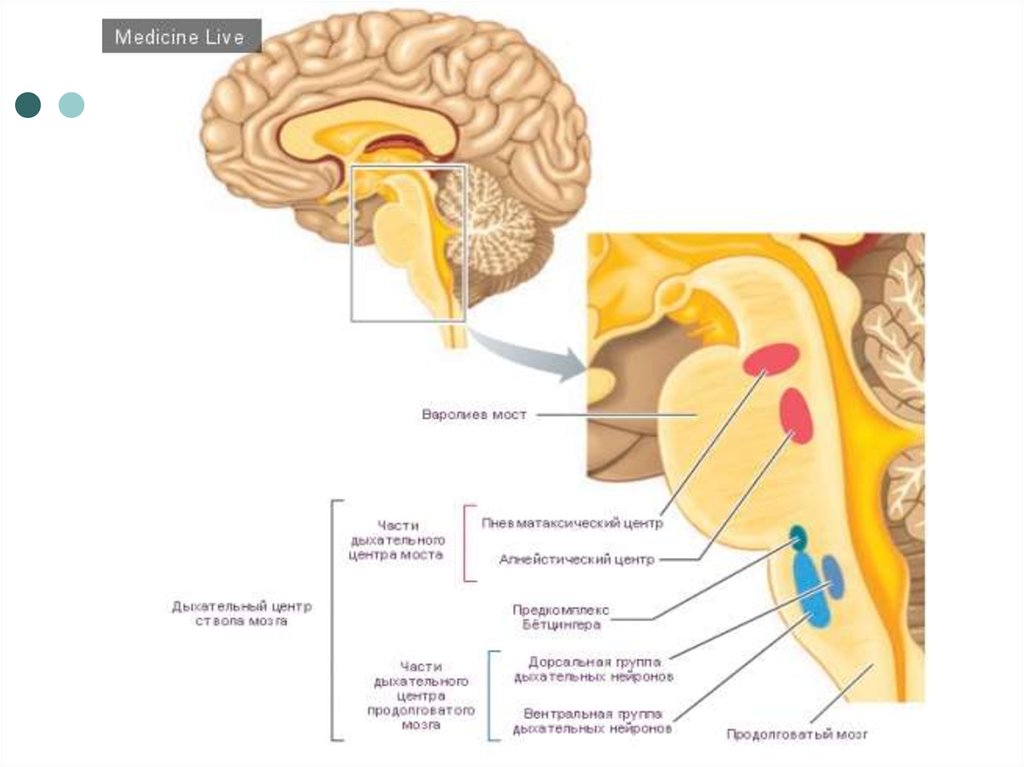
8. Уровни локализации структур ДЦ.
1. Корковое представительство (речь, пение, смех, чтение иусловно-рефлекторное изменение дыхательной функции).
2. Стриопалидарные структуры и лимбическая система мозга
(изменение дыхательной функции в условиях эмоциональных
нагрузок).
3. Гипоталамические нейроны.
4. Варолиев мост – ПТЦ.
5. Апноэстический центр.
6. Продолговатый мозг – ДЦ (центр жизненного обеспечения
дыхательной функции, состоящий из инспираторного и
экспираторного отделов).
7. Спинной мозг, шейные сегменты С3-С4 (диафрагмальное ядро
спинального дыхательного центра).
8. Спинной мозг, грудные сегменты Th1-Th6, аксоны которых
образуют межреберные нервы.
9.
10. АВТОМАТИЯ ДЫХАТЕЛЬНОГО ЦЕНТРА
1.2.
3.
Способность инспираторных и экспираторных нейронов
автоматически генерировать ПД описывалась еще
И.М.Сеченовым, а затем была зарегистрирована в 1931 г.
Эдрианом на ДЦ головного мозга золотой рыбки.
Автоматия ДЦ имеет ряд особенностей, отличающих ее от
автоматии сердца:
Автоматия формируется в результате взаимодействия многих
нервных клеток, некоторые из которых обладают свойствами
тормозных нейронов.
Автоматия ДЦ поддерживается постоянными тоническими
сигналами от хеморецепторов сосудистых зон, ретикулярной
формации, гипоталамуса, рецепторов растяжения легких.
Атоматия ДЦ находится под постоянным контролем
вышерасположенных уровней ДЦ – коры, поэтому человек
может изменять частоту и глубину дыхания в широких
пределах.
11. Нейроны дыхательного центра
Обнаружены две группы нейронов: инспираторные иэкспираторные. Четкой их локализации в дыхательных ядрах
продолговатого мозга нет. Однако, дорсальное ядро содержит
наибольшее количество инспираторных нейронов, а вентральное –
содержит и инспираторные, и экспираторные нейроны.
Инспираторные нейроны обнаружены с помощью
микроэлектродной техники регистрации потенциала действия
отдельных нейронов, возбуждающихся в соответствии с фазами
дыхательного цикла. Инспираторные нейроны возбуждаются за 0,10,2 с до начала дыхательного цикла, частота их ПД постепенно
возрастает и на вдохе достигает 70-100 Гц (при глубоком вдохе – до
300 Гц). При смене вдоха выдохом частота ПД инспираторных
нейронов резко уменьшается или они вовсе прекращаются.
Экспираторные нейроны генерируют ПД во время выдоха,
прекращение ПД или уменьшение их частоты происходит перед
самым началом новой инспирации.
12. ЛОКАЛИЗАЦИЯ ДЫХАТЕЛЬНЫХ НЕЙРОНОВ
В правой и левой половинах продолговатого мозга содержатся по дваскопления дыхательных нейронов – дорсальные и вентральные.
Дорсальное ядро содержит в основном инспираторные нейроны
(экспиратоных – около 5 %). Их аксоны по нисходящим путям спинного
мозга достигают диафрагмальных ядер шейного отдела спинного мозга
(СIII-CIV). Здесь мотонейроны, образующие это диафрагмальное ядро,
своими аксонами (их около 1000) формируют диафрагмальный нерв,
иннервирующий мышцы диафрагмы. Кроме этого, инспираторные
нейроны дорсального ядра посылают коллатерали к инспираторным
нейронам вентрального ядра, возбуждая их и тем самым тормозя
экспираторные нейроны вентрального ядра.
Вентральное ядро содержит инспираторные и экспираторные нейроны,
расположено в вентралатеральной области продолговатого мозга.
Аксоны нейронов вентрального ядра спускаются к спинномозговым
ядрам межреберной мускулатуры на уровень грудных сегментов ThI –
ThVI.
Парабранхиальное ядро варолиевого моста – ПТЦ – третья локализация
дыхательных нейронов в заднем мозге.
13.
14. Дыхательный цикл
Возбуждение нейронов всех уровней дыхательного центраобеспечивает характерный рисунок (паттерн) возбуждения
дыхательных мышц – формируется дыхательный цикл. У
человека его продолжительность составляет 3-5 секунд. В
определенный момент возникает возбуждение
диафрагмальных нервов, оно постепенно усиливается при
спокойном дыхании в течение 1-2,5 с. В результате сила
сокращения диафрагмальных мышц усиливается. Затем
возбуждение диафрагмальных нервов резко ослабевает,
инспирация сменяется экспирацией. Через 2-3 секунды
наступает новая инспирация (инспирация короче экспирации).
Возбуждение мотонейронов, аксоны которых иннервируют
наружные косые межреберные и межхрящевые мышцы вдоха,
происходит также, но наступает несколько позже. При
активном выдохе происходит возбуждение внутренних
межреберных мышц и мышц живота.
15. Влияние перерезок на разных уровнях ствола мозга на дыхание
16. Нервная регуляция дыхательной функции. Роль механорецепторов в регуляции дыхания.
1.2.
3.
Нервная регуляция дыхательной функции обеспечивает
изменение вентиляции легких в зависимости от изменения
объема легких и воздухоносных путей.
Осуществляется с рецептивных полей дыхательной
системы:
рецепторов растяжения легких (механорецепторов);
ирритантных (смешанных) рецепторов;
юкстаальвеолярных рецепторов капилляров альвеол (jрецепторов).
В лёгких специализированных рецепторов для О2 и
СО2 нет!
17. Регуляция дыхательной функции при раздражении механорецепторов легких
Рецепторы растяжения лёгких – механорецепторы. В каждом лёгкомсодержится около 1000 рецепторов растяжения. Они расположены
преимущественно в гладких мышцах стенок воздухоносных путей – от
трахеи до мелких бронхов. В альвеолах и плевре таких рецепторов нет.
Увеличение объёма лёгких косвенно стимулирует рецепторы
растяжения внутрилегочных воздухоносных путей: непосредственным
их раздражителем является внутреннее напряжение стенки
воздухоносных путей.
Частота потенциала действия в афферентных волокнах от рецепторов
растяжения увеличивается при вдохе и снижается при выдохе. Чем
глубже вдох, тем больше частота импульсов, посылаемых в ДЦ.
Рецепторы растяжения имеют различный порог возбуждения:
низкопороговые рецепторы возбуждены при выдохе, даже при
полном спадении лёгких, а при вдохе частота ПД в них нарастает.
Высокопороговые рецепторы возбуждаются только при вдохе, когда
объём лёгких увеличивается сверх функциональной остаточной
ёмкости (ФОЕ).
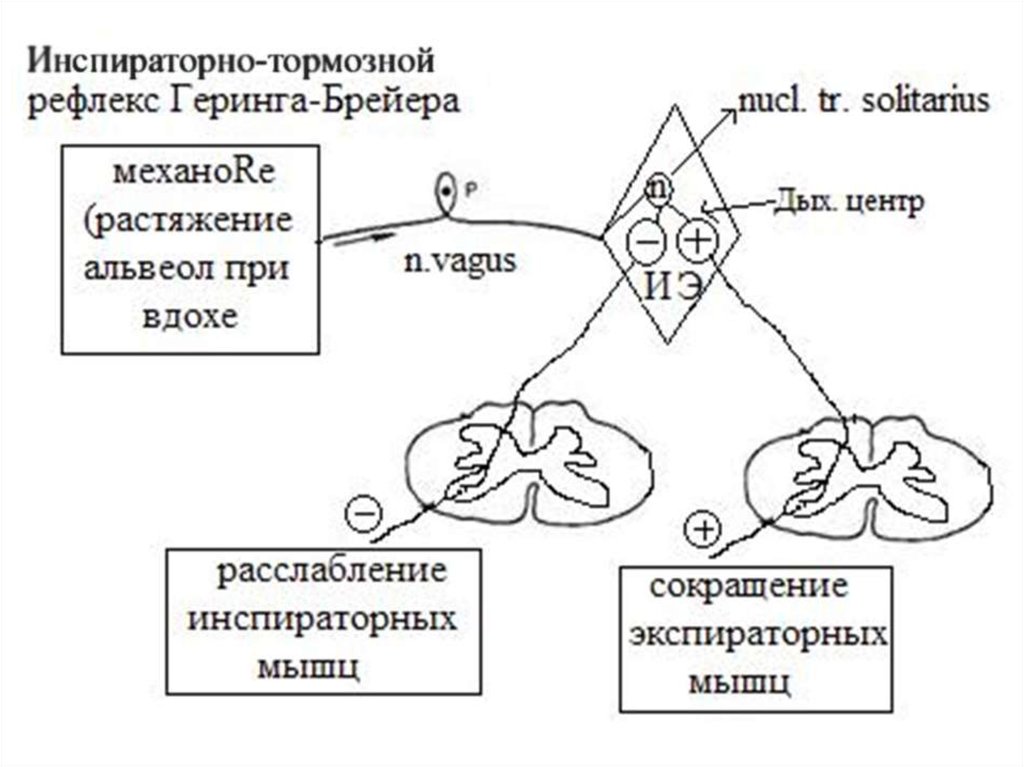
18. Регуляция дыхания при раздражении механорецепторов осуществляется рефлекторно – дыхательные рефлексы Геринга и Брейера
Их физиологическое значение состоит в регулировании глубины и частотыдыхания в зависимости от состояния лёгких. Эти рефлексы
обеспечивают объёмную обратную связь ДЦ с исполнительным
аппаратом дыхательной системы:
1.
При сохранённых блуждающих нервах (афферентные нервы, идущие
от механорецепторов лёгких) гиперпноэ, вызываемое гиперкапнией и
гипоксией, проявляется увеличением как частоты, так и глубины
дыхания, т.е. ОМВЛ=ЧД×ДО;
2.
При перерезке блуждающих нервов учащения дыхания не происходит.
Вентиляция лёгких нарастает уже не за счёт ЧД, а только за счёт
увеличения глубины дыхания, поэтому в результате максимальная
величина вентиляции лёгких оказывается уменьшенной почти вдвое.
Таким образом, сигналы от рецепторов лёгких обеспечивают
повышение ЧД и ДО при гиперкапнии или гипоксемии, что приводит к
максимальной вентиляции лёгких в этих условиях (возникает
гиперпноэ).Этот эффект достигается вследствие того, что дыхательные
рефлексы представляют собой смену дыхательных фаз, т.е. смену
вдоха выдохом и наоборот, в зависимости от изменения объёма лёгких.
19.
20.
А. Инспираторно-тормозный рефлекс Геринга и Брейера: увеличениеобъёма лёгких во время осуществляемого вдоха вызывает его выключение:
от рецепторов растяжения лёгких возбуждение по афферентным веточкам
поступает в дорсальное ядро продолговатого мозга (центр вдоха). Оно
состоит из Iα- и Iβ-нейронов. Iα-нейроны (их около 60 % всех нейронов
инспираторного центра ДЦ) под влиянием этой афферентации тормозятся,
а Iβ-нейроны – возбуждаются. Iβ-нейроны являются промежуточной и
дополнительной «станцией» торможения Iα-нейронов. В результате этого
экспираторные нейроны ДЦ освобождаются от реципрокного торможения
инспираторными нейронами, и происходит смена вдоха выдохом.
В. Экспираторно-облегчающий рефлекс Геринга-Брейера: увеличение
объема лёгких на вдохе (раздувание их дополнительным объемом воздуха)
вызывает удлинение выдоха.
С. Парадоксальный эффект Хэда: сильное дополнительное раздувание
лёгких вызывает сильное возбуждение инспираторных мышц (в течение 0,10,5 с), в результате возникает судорожный вдох – вздох.
D. Рефлекс на спадение лёгких – уменьшение объёма лёгких во время уже
осуществляемого выдоха обеспечивает усиление активности
инспираторных нейронов, следовательно – укорочение выдоха и смену его
вдохом.
21. Вывод:
Обмен воздуха в альвеолахпроисходит за счёт работы аппарата
внешнего дыхания У взрослого
человека во время спокойного
дыхания рефлексы Геринга-Брейера
большого значения не имеют.
Однако, форсированное дыхание
(гиперпноэ) сопровождается
обязательным увеличением частоты
дыхания, согласно дыхательным
рефлексам Геринга и Брейера.
Эти рефлексы хорошо выражены у
новорожденных. Они обеспечивают
укорочение дыхательных фаз –
особенно фазы выдоха.
Выраженность рефлексов постепенно
уменьшается уже в первые дни и
недели после рождения.
22. Регуляция дыхательной функции при раздражении ирритантных рецепторов
Ирритантные рецепторы располагаются преимущественно в эпителии исубэпителиальном слое воздухоносных путей, их особенно много в области корней
лёгких. Они обладают одновременно свойствами механо- и хеморецепторов.
Особенности ирритантных рецепторов:
1. Порог возбуждения выше, чем у рецепторов растяжения лёгких;
2. Импульсы ПД в афферентных волокнах возникают только на короткое время в
форме вспышек во время изменения объёма лёгких (поэтому их ещё называют
быстро адаптирующимися рецепторами лёгких);
3. Раздражителями являются:
большие изменения объёма лёгких на вдохе и выдохе,
пылевые частицы и слизь, накапливающаяся в дыхательных путях,
пары едких веществ (аммиак, эфир, двуокись серы, табачный дым),
БАВ, образующиеся в стенке лёгких (гистамин),
снижение растяжимости легочной ткани,
бронхиальная астма, отёк лёгких, пневмоторакс,
застой крови в малом круге кровообращения.
Раздражение ирритантных рецепторов вызывает у человека неприятные ощущения
першения и жжения. При раздражении ирритантных рецепторов трахеи возникает
кашель. При раздражении ирритантных рецепторов бронхов изменяется паттерн
дыхательного цикла – усиливается инспираторная активность за счет укорочения
выдоха и наступления более раннего вдоха, в результате увеличивается ЧДД
23. Регуляция дыхательной функции при раздражении j-рецепторов альвеол
Эти рецепторы находятся вблизи от капилляров малогокруга кровообращения в интерстициальной ткани альвеол.
Участие j-рецепторов в регуляции дыхания у здоровых
людей не определяется. Основной раздражитель jрецепторов – увеличение объема интерстициальной
жидкости в легочной ткани. Поэтому сильное и устойчивое
возбуждение j-рецепторов наблюдается при пневмониях,
отёке лёгких, эмболии мелких сосудов лёгких, застое крови в
малом круге кровообращения, т.е. при повреждении
легочной ткани. Афферентными нервами, проводящими
возбуждение в дыхательный центр от j-рецепторов,
являются веточки вагуса, относящиеся к нервным волокнам
типа С, результатом возбуждения этих рецепторов является
учащение дыхания на фоне снижения глубины дыхательных
движений (одышка), а также рефлекторная
бронхоконстрикция.
24. Гуморальная регуляция дыхания. Зависимость деятельности дыхательного центра от газового состава крови.
Деятельность дыхательного центра определяет глубину ичастоту дыхания, тем самым регулируется объём вентиляции
лёгких, а следовательно, напряжение дыхательных газов крови.
При этом деятельность дыхательного центра зависит от газового
состава крови (обратная отрицательная связь). Ведущее место в
определении величины вентиляции лёгких имеет СО2,
напряжение которого в крови изменяет активность нейронов ДЦ.
Так, при увеличении напряжения СО2 в
артериальной крови от 40 до 60 мм
рт. ст. МОД увеличивается в 10 раз
(до 75 л/мин), тогда как снижение О2
от 90 до 30 мм рт. ст. – увеличивает
МОД всего в 2-3 раза (20-22 л/мин).
25. Напряжение СО2 в крови имеет своё определение: в норме – нормокапния (40 мм рт. ст. в артериальной крови и 46 мм рт. ст. в венозной крови), увели
Напряжение СО2 в крови имеет своё определение:в норме – нормокапния (40 мм рт. ст. в артериальной крови и
46 мм рт. ст. в венозной крови),
увеличение – гиперкапния (выше 60 мм рт. ст.),
уменьшение – гипокапния (ниже 39 мм рт.ст.).
Напряжение О2 в крови:
в норме – нормоксия (90-92 мм рт. ст. в артериальной крови)
увеличение – гипероксия
снижение – гипоксемия и гипоксия (снижение О2 в тканях)
Когда напряжение О2 и СО2 находится в пределах нормы, наблюдается
нормальная работа дыхательного центра, выражающаяся в нормальном
дыхании – эйпноэ (12-18 раз в минуту). Гиперкапния наряду с гипоксией и
увеличением концентрации водородных ионов Н+ в крови (уменьшение рН)
(ацидоз) сопровождается увеличением частоты и глубины дыхания, что носит
название гиперпноэ. При гиперкапнии и защелачивании крови (уменьшение
концентрации ионов водорода Н+, увеличении рН) (алкалоз) происходит
уменьшение вентиляции, вплоть до остановки дыхания – апноэ. Гиперкапния
наряду с гипоксией называется асфиксией. Гипоксия вначале вызывает
умеренное гиперпноэ, которое при увеличении степени гипоксии завершается
апноэ. Гипоксическое апноэ связано с ослаблением окислительных процессов
в нейронах дыхательного центра и в мозге в целом, поэтому ему
предшествует потеря сознания, что создаёт смертельную опасность развития
гипоксического апноэ.
26. Изменение газового состава крови является ведущим фактором гуморальной регуляции дыхания. Имеется две группы хеморецепторов, учавствующ
Изменение газового состава крови является ведущимфактором гуморальной регуляции дыхания.
Имеется две группы хеморецепторов, учавствующих в
регуляции дыхания: периферические и центральные.
Периферические хеморецепторы – аортальное и каротидное
тельца. Хеморецепторы этих областей особенно чувствительны к
гипоксии, т.к. информируют (афферентируют) дыхательный центр о
газовом составе крови, которая направляется к мозгу. Они
расположены в специальных маленьких тельцах, обильно снабжаемых
артериальной кровью. Хеморецепторы телец особенно чувствительны к
колебаниям газового состава крови. Степень их активации возрастает
при изменении напряжения О2 и СО2 в артериальной крови в
зависимости от фаз вдоха при глубоком и частом дыхании.
Наблюдается явление потенциации: стимулирующее действие
гипоксии и гиперкапнии на хеморецепторы каротидного тельца
усиливается. Наоборот, при гипероксии чувствительность
хеморецепторов к СО2 резко снижается.
Чувствительность хеморецепторов находится под нервным контролем:
парасимпатические эфферентные влияния снижают чувствительность
рецепторов, а раздражение симпатических волокон повышает её.
27. Центральные хеморецепторы (ЦХР)
ЦХР были обнаружены в продолговатом мозге латеральнее пирамид. Перфузияэтой области раствором со сниженным рН резко усиливает дыхание. При
защелачивании раствора дыхание ослабевает вплоть до апноэ. Такие же
изменения происходят при охлаждении или обработке местными анестетиками
этого участка продолговатого мозга.
ЦХР расположены в тонком слое мозгового вещества продолговатого мозга на
глубине не более 0,2 мм. Выделяют два поля – М-S и L. В обычных условиях
рецепторы продолговатого мозга постоянно стимулируются ионами Н+,
находящимися в спинномозговой жидкости. Концентрация водородных ионов в
ней зависит от напряжения СО2 в артериальной крови: гиперкапния (как явление
увеличения напряжения СО2) сопровождается нарастание концентрации Н+ и
снижением рН. ЦХР оказывают более сильное влияние на деятельность
дыхательного центра, чем периферические. Это проявляется через
существенное изменение вентиляции лёгких при снижении рН спинномозговой
жидкости: снижение рН на 0,01 единицу сопровождается увеличение легочной
вентиляции на 4 л/мин (т.е. на половину нормального объёма минутной
вентиляции). Вместе с тем, ЦХР реагируют на изменение напряжения СО2 в
артериальной крови позже (через 20-30 с) по сравнению с периферическими
хеморецепторами, которые возбуждаются уже через 3-5 с. эта особенность
обусловлена тем, что для диффузии СО2 из крови в спинномозговую жидкость и
далее – в ткань мозга необходимо время. Таким образом, чувствительность
периферических хеморецепторов ниже, чем центральных, но в процесс
регуляции дыхательной функции они включаются раньше!
28. Механизм первого вдоха новорожденных
Возбуждение центральныххеморецепторов происходит
у новорожденных в момент
рождения. При пережатии
пуповины возникает
гиперкапния, которая
вызывает увеличение
концентрации ионов Н+ в
ликворе, что является
возбуждающим фактором
для нейронов
инспираторного отдела
дыхательного центра.
Происходит первый вдох.