




 Биология
БиологияПохожие презентации:









")
Строение и функции центральных синапсов. Лекция 3
1.
Строение и функции центральныхсинапсов
2.
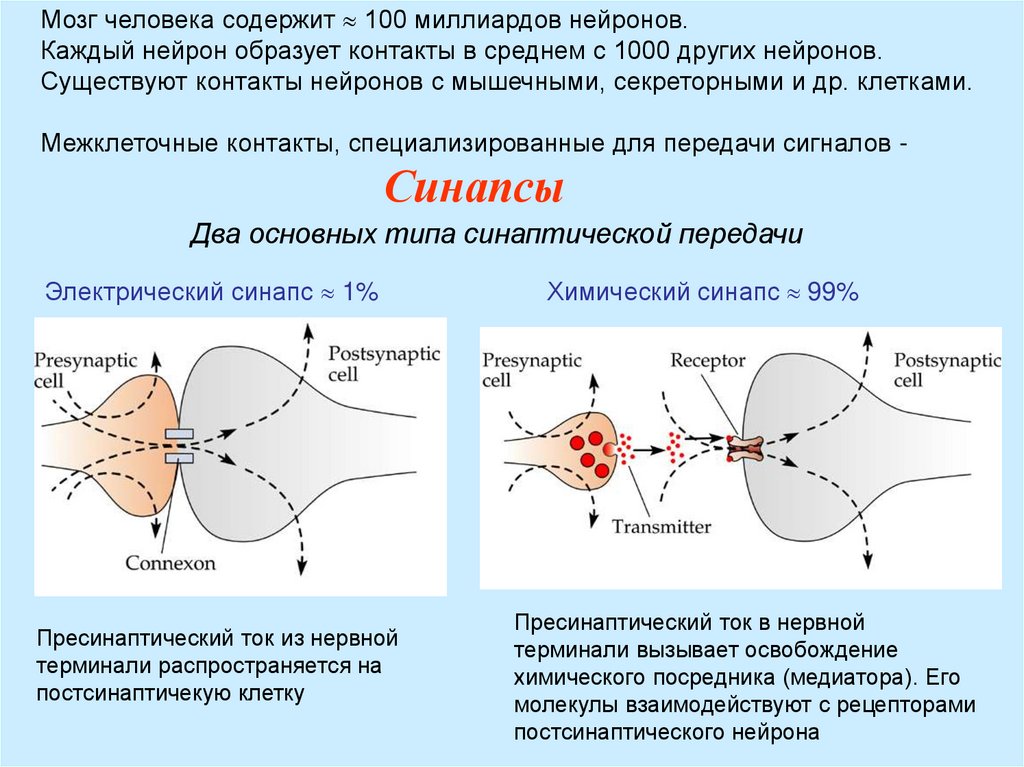
Мозг человека содержит 100 миллиардов нейронов.Каждый нейрон образует контакты в среднем с 1000 других нейронов.
Существуют контакты нейронов с мышечными, секреторными и др. клетками.
Межклеточные контакты, специализированные для передачи сигналов -
Синапсы
Два основных типа синаптической передачи
Электрический синапс 1%
Пресинаптический ток из нервной
терминали распространяется на
постсинаптичекую клетку
Химический синапс 99%
Пресинаптический ток в нервной
терминали вызывает освобождение
химического посредника (медиатора). Его
молекулы взаимодействуют с рецепторами
постсинаптического нейрона
3.
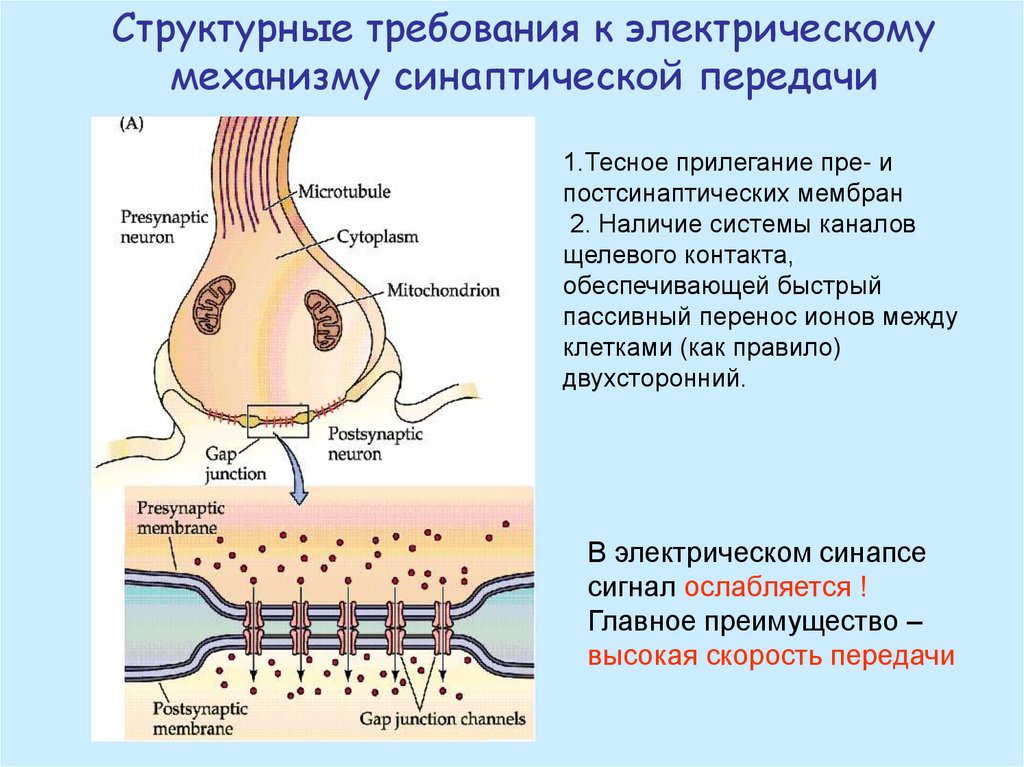
Структурные требования к электрическомумеханизму синаптической передачи
1.Тесное прилегание пре- и
постсинаптических мембран
2. Наличие системы каналов
щелевого контакта,
обеспечивающей быстрый
пассивный перенос ионов между
клетками (как правило)
двухсторонний.
В электрическом синапсе
сигнал ослабляется !
Главное преимущество –
высокая скорость передачи
4.
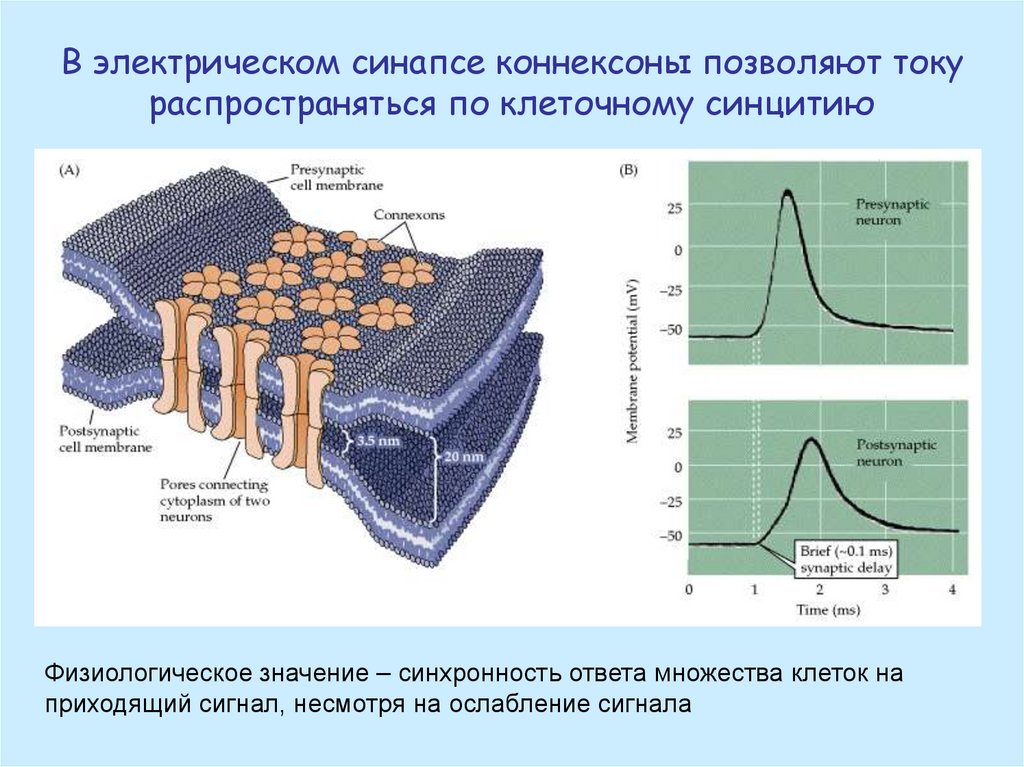
В электрическом синапсе коннексоны позволяют токураспространяться по клеточному синцитию
Физиологическое значение – синхронность ответа множества клеток на
приходящий сигнал, несмотря на ослабление сигнала
5. Структурные требования к химическому механизму синаптической передачи
Взаимодействие трех структур:(а) пресинаптической –электрически
управляемая секреция медиатора;
(б) синаптической щели –
пространство шириной 40-50 нм, где
происходит свободная диффузия
медиатора;
(в) постсинаптической мембраной,
обладающей лиганд-активируемыми
каналами.
В химическом синапсе
происходит двукратная
трансформация сигнала: из
электро в хемо и снова в электро.
Сигнал усиливается, но
возникает синаптическая
задержка
6.
МедиаторыТребования к молекулярным свойствам медиаторов
• Высокая скорость диффузии, а значит низкий молекулярный вес
• Относительная простота и скорость синтеза (небольшое число стадий)
• Доступность исходных продуктов и наличие систем поступления их в нервную клетку
• Невысокие энергетические затраты («дешевизна») на синтез или обратный захват
нейроном
• Возможность повторного использования самого медиатора или непосредственных
продуктов его метаболизма
• Почти все медиаторы способны как возбуждать, так и тормозить
• Часть медиаторов могут участвовать как в быстрых, так и медленных процессах
Основные медиаторы
Ацетилхолин
Норадреналин
Серотонин (5HT)
Гистамин
Допамин
Глутамат
ГАМК
Глицин
АТФ
Аденозин
Нейропептиды
энкефалин
вещество Р
соматостатин
нейротензин
NO (окись азота)
7.
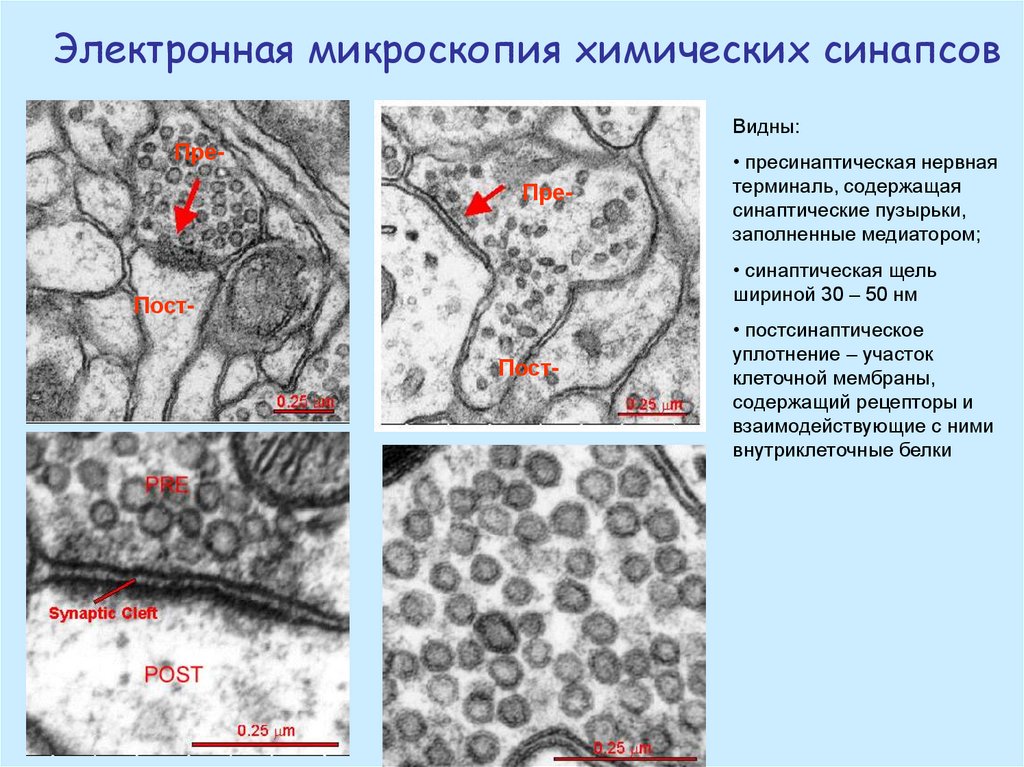
Электронная микроскопия химических синапсовВидны:
ПреПре-
• пресинаптическая нервная
терминаль, содержащая
синаптические пузырьки,
заполненные медиатором;
• синаптическая щель
шириной 30 – 50 нм
ПостПост-
• постсинаптическое
уплотнение – участок
клеточной мембраны,
содержащий рецепторы и
взаимодействующие с ними
внутриклеточные белки
8.
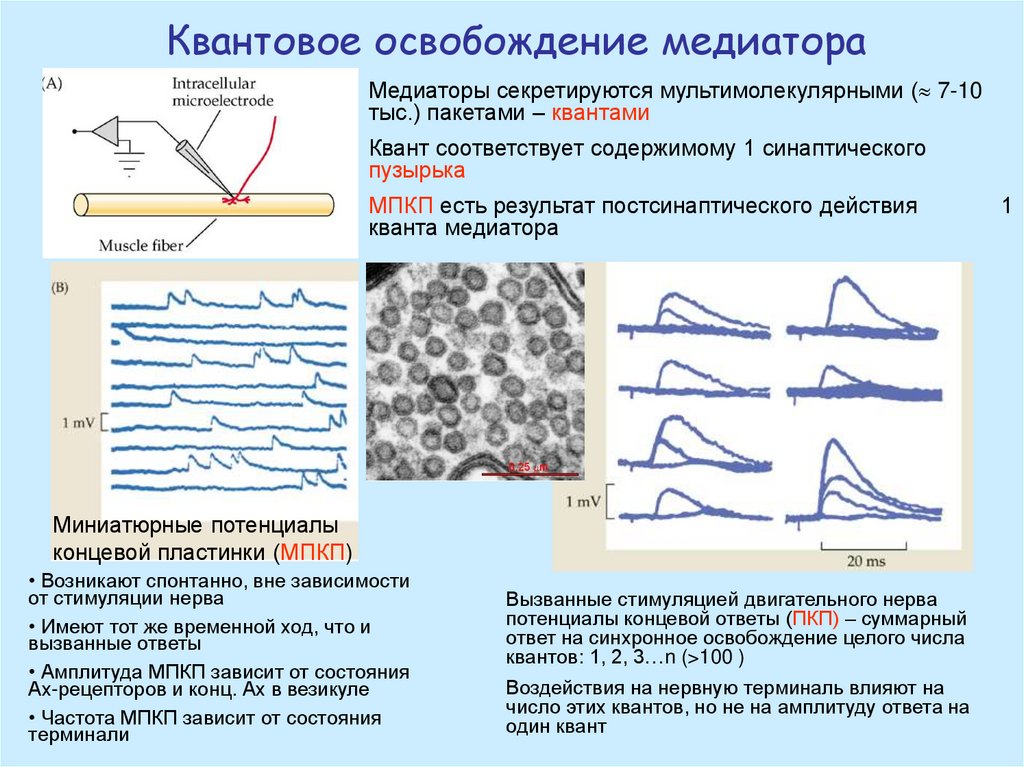
Квантовое освобождение медиатораМедиаторы секретируются мультимолекулярными ( 7-10
тыс.) пакетами – квантами
Квант соответствует содержимому 1 синаптического
пузырька
МПКП есть результат постсинаптического действия
кванта медиатора
Миниатюрные потенциалы
концевой пластинки (МПКП)
• Возникают спонтанно, вне зависимости
от стимуляции нерва
• Имеют тот же временной ход, что и
вызванные ответы
• Амплитуда МПКП зависит от состояния
Ах-рецепторов и конц. Ах в везикуле
• Частота МПКП зависит от состояния
терминали
Вызванные стимуляцией двигательного нерва
потенциалы концевой ответы (ПКП) – суммарный
ответ на синхронное освобождение целого числа
квантов: 1, 2, 3…n (>100 )
Воздействия на нервную терминаль влияют на
число этих квантов, но не на амплитуду ответа на
один квант
1
9.
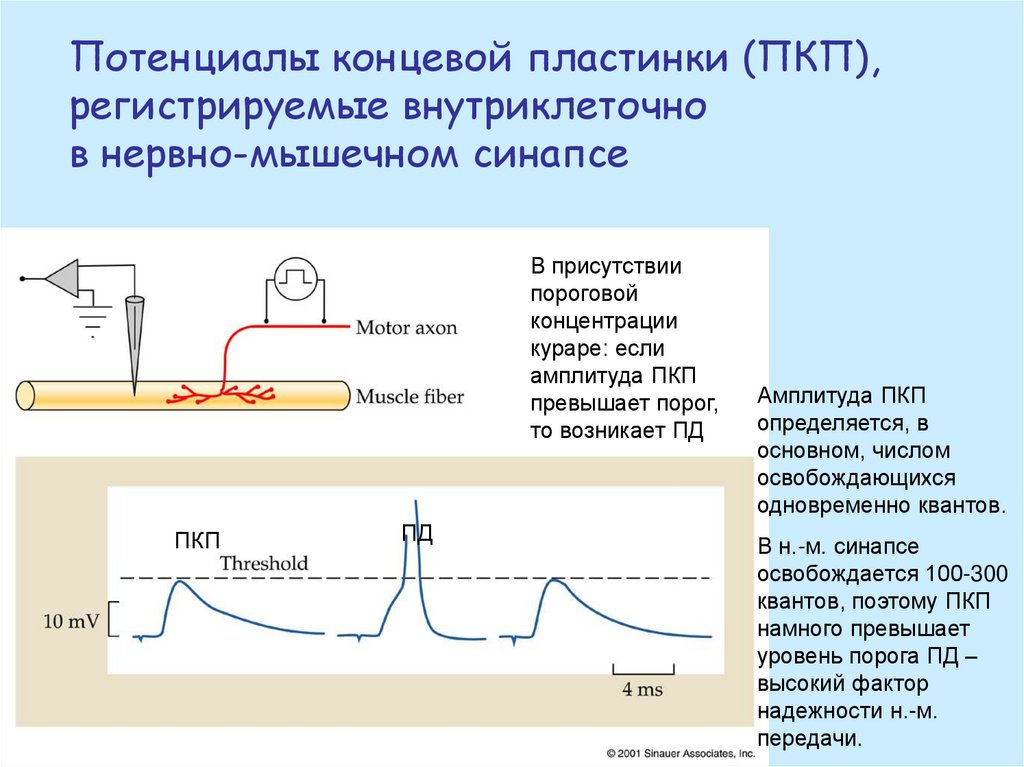
Потенциалы концевой пластинки (ПКП),регистрируемые внутриклеточно
в нервно-мышечном синапсе
В присутствии
пороговой
концентрации
кураре: если
амплитуда ПКП
превышает порог,
то возникает ПД
ПКП
ПД
Амплитуда ПКП
определяется, в
основном, числом
освобождающихся
одновременно квантов.
В н.-м. синапсе
освобождается 100-300
квантов, поэтому ПКП
намного превышает
уровень порога ПД –
высокий фактор
надежности н.-м.
передачи.
10.
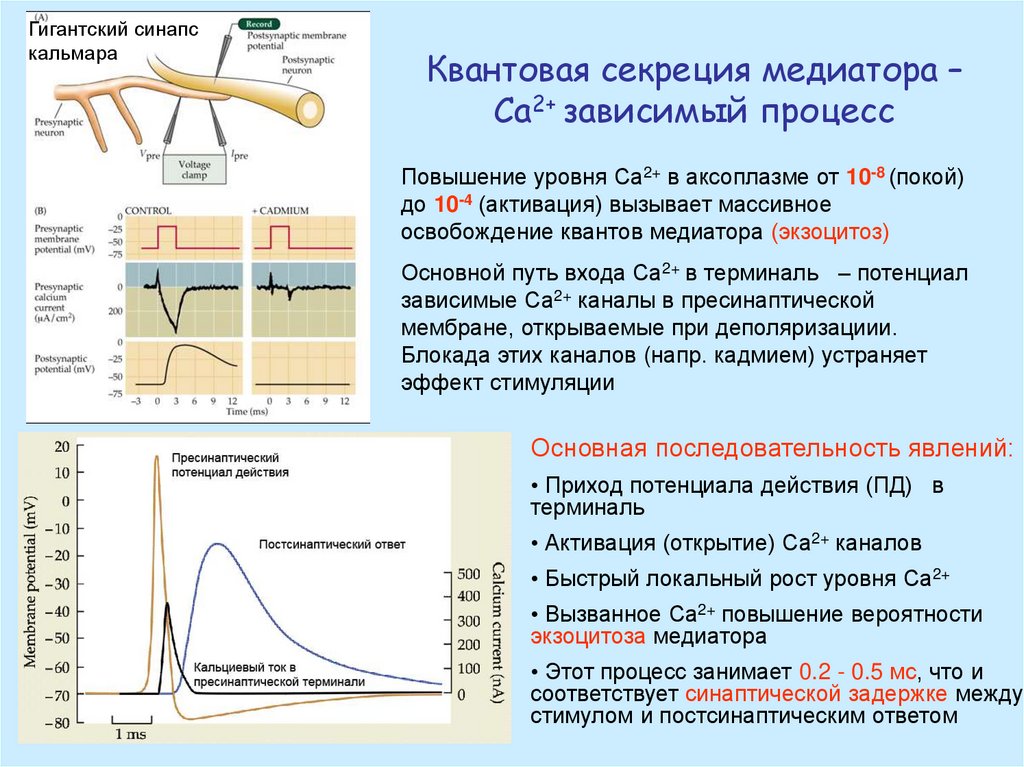
Гигантский синапскальмара
Квантовая секреция медиатора –
Са2+ зависимый процесс
Повышение уровня Са2+ в аксоплазме от 10-8 (покой)
до 10-4 (активация) вызывает массивное
освобождение квантов медиатора (экзоцитоз)
Основной путь входа Са2+ в терминаль – потенциал
зависимые Са2+ каналы в пресинаптической
мембране, открываемые при деполяризациии.
Блокада этих каналов (напр. кадмием) устраняет
эффект стимуляции
Основная последовательность явлений:
• Приход потенциала действия (ПД) в
терминаль
• Активация (открытие) Са2+ каналов
• Быстрый локальный рост уровня Са2+
• Вызванное Са2+ повышение вероятности
экзоцитоза медиатора
• Этот процесс занимает 0.2 - 0.5 мс, что и
соответствует синаптической задержке между
стимулом и постсинаптическим ответом
11.
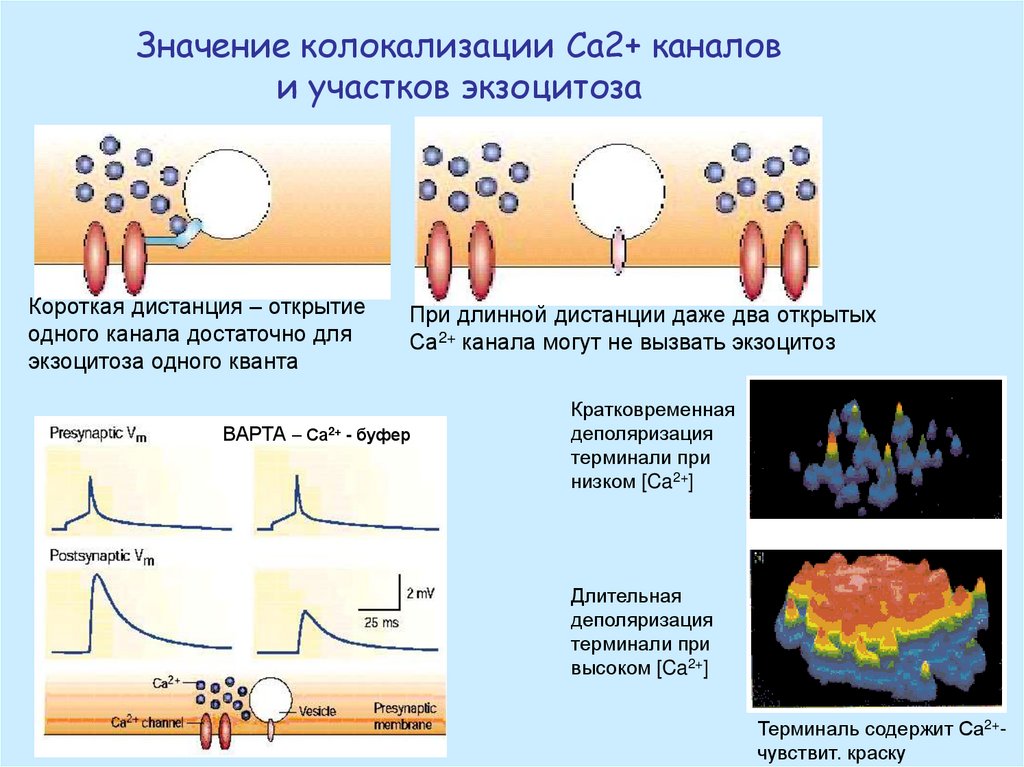
Значение колокализации Са2+ каналови участков экзоцитоза
Короткая дистанция – открытие
одного канала достаточно для
экзоцитоза одного кванта
ВАРТА
При длинной дистанции даже два открытых
Са2+ канала могут не вызвать экзоцитоз
– Са2+ - буфер
Кратковременная
деполяризация
терминали при
низком [Ca2+]
Длительная
деполяризация
терминали при
высоком [Ca2+]
Терминаль содержит Са2+чувствит. краску
12.
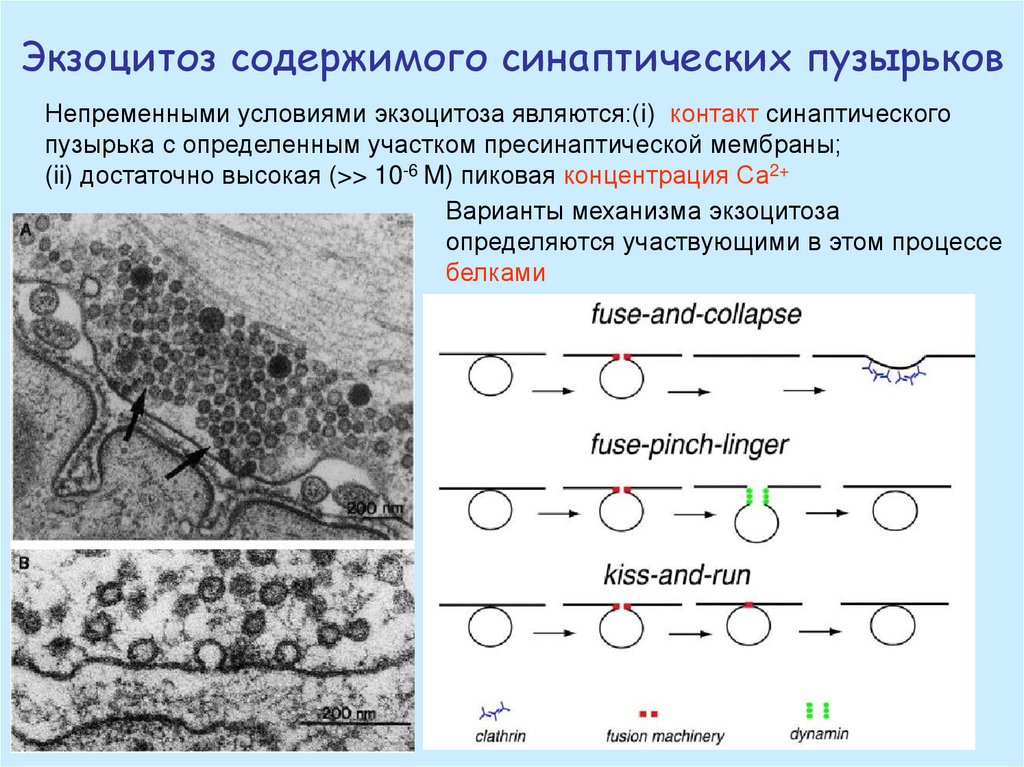
Экзоцитоз содержимого синаптических пузырьковНепременными условиями экзоцитоза являются:(i) контакт синаптического
пузырька с определенным участком пресинаптической мембраны;
(ii) достаточно высокая (>> 10-6 M) пиковая концентрация Са2+
Варианты механизма экзоцитоза
определяются участвующими в этом процессе
белками
13.
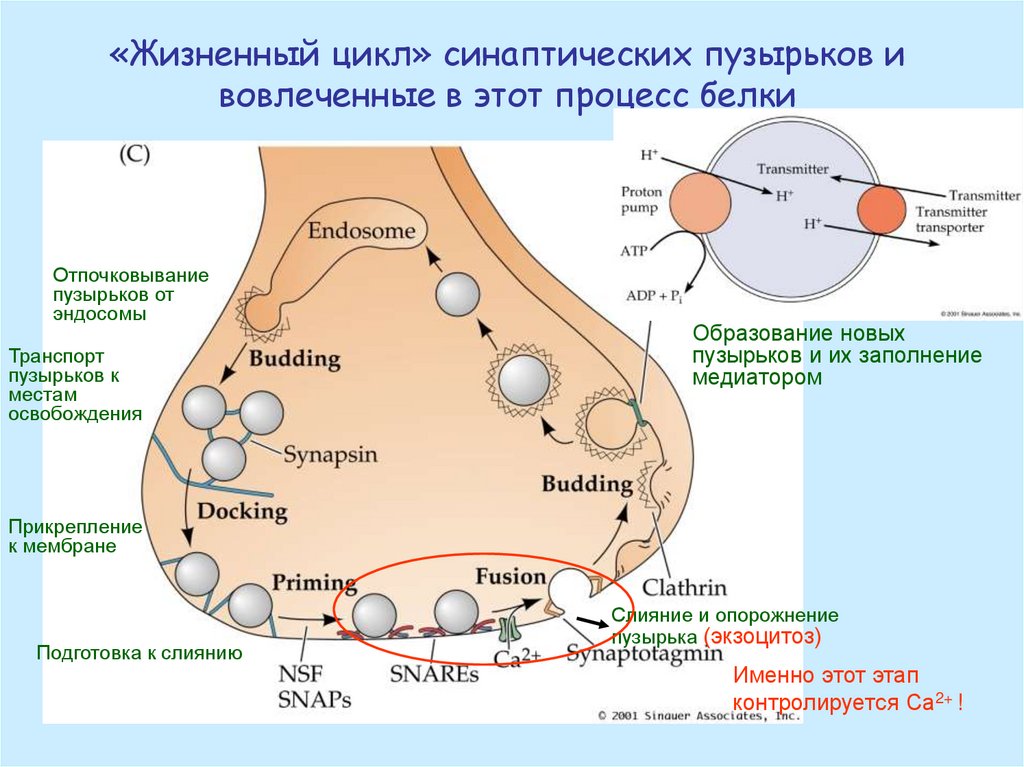
«Жизненный цикл» синаптических пузырьков ивовлеченные в этот процесс белки
Отпочковывание
пузырьков от
эндосомы
Транспорт
пузырьков к
местам
освобождения
Образование новых
пузырьков и их заполнение
медиатором
Прикрепление
к мембране
Подготовка к слиянию
Слияние и опорожнение
пузырька (экзоцитоз)
Именно этот этап
контролируется Са2+ !
14.
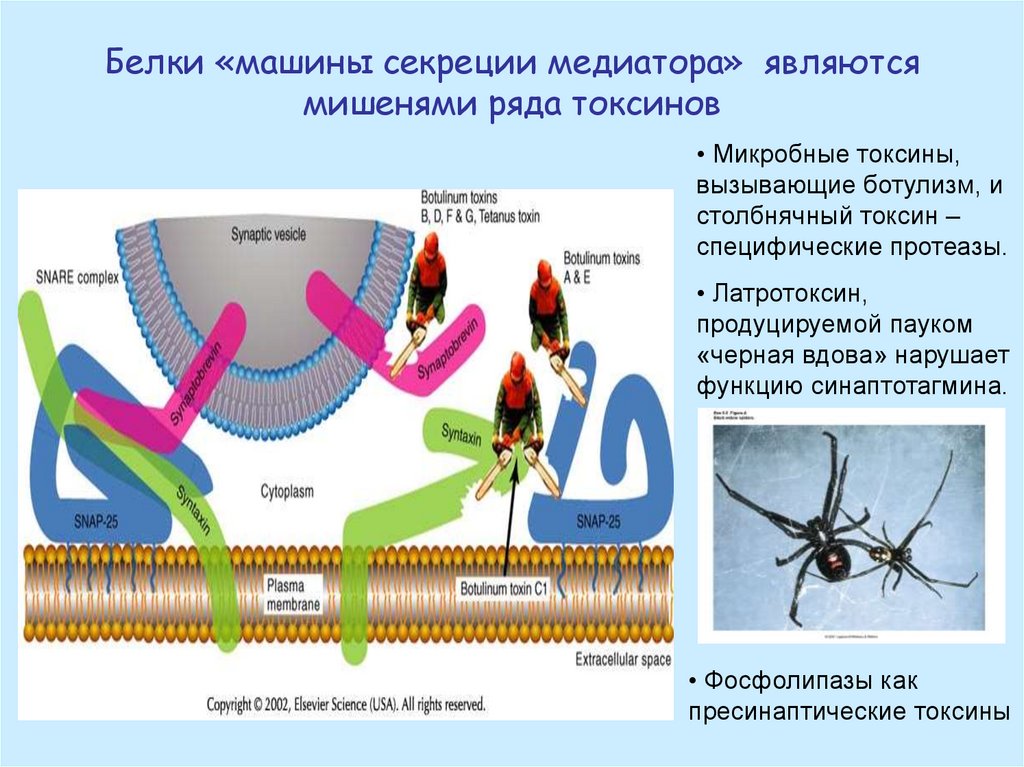
Белки «машины секреции медиатора» являютсямишенями ряда токсинов
• Микробные токсины,
вызывающие ботулизм, и
столбнячный токсин –
специфические протеазы.
• Латротоксин,
продуцируемой пауком
«черная вдова» нарушает
функцию синаптотагмина.
• Фосфолипазы как
пресинаптические токсины
15.
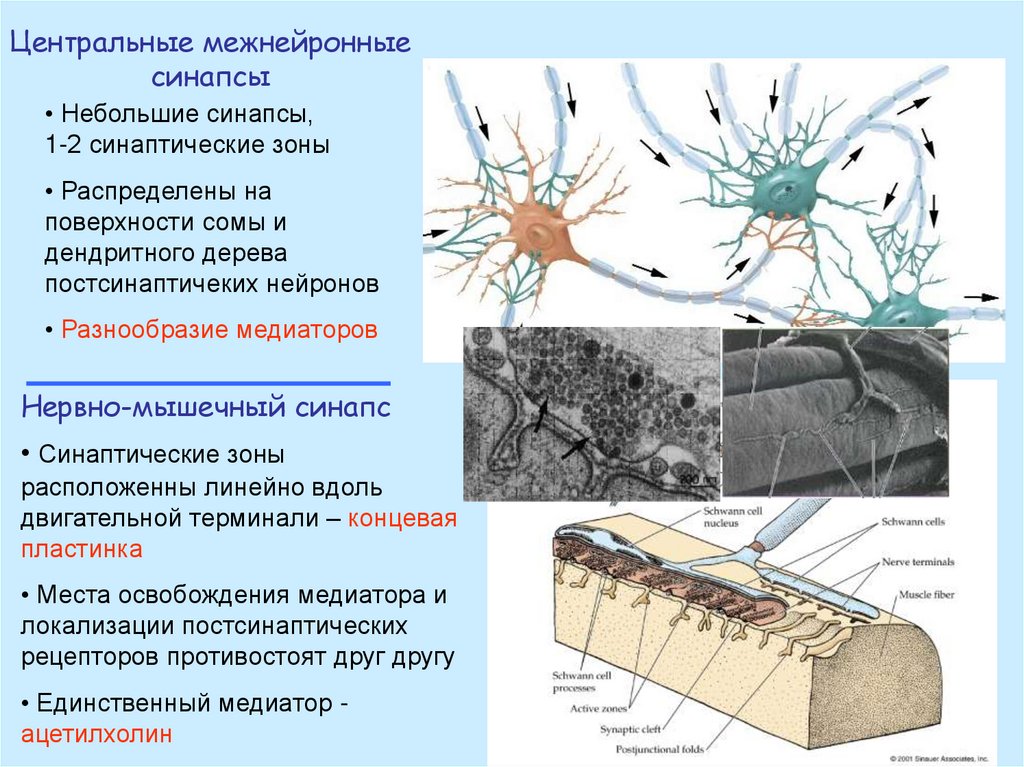
Центральные межнейронныесинапсы
• Небольшие синапсы,
1-2 синаптические зоны
• Распределены на
поверхности сомы и
дендритного дерева
постсинаптичеких нейронов
• Разнообразие медиаторов
Нервно-мышечный синапс
• Синаптические зоны
расположенны линейно вдоль
двигательной терминали – концевая
пластинка
• Места освобождения медиатора и
локализации постсинаптических
рецепторов противостоят друг другу
• Единственный медиатор ацетилхолин
16.
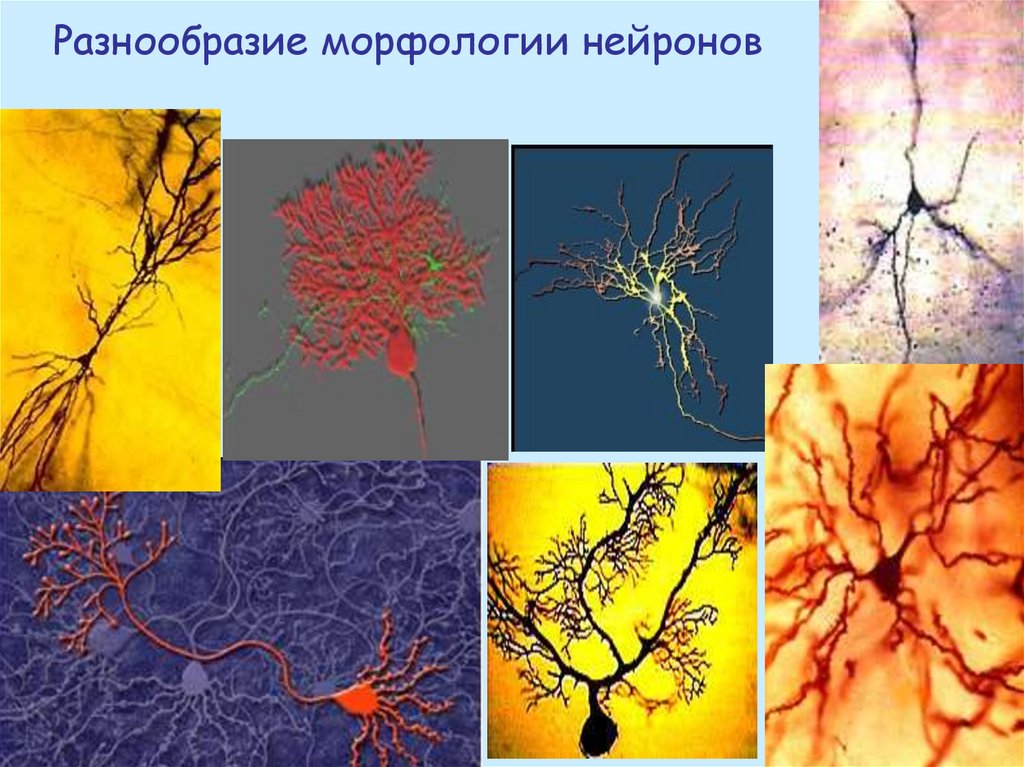
Разнообразие морфологии нейронов17.
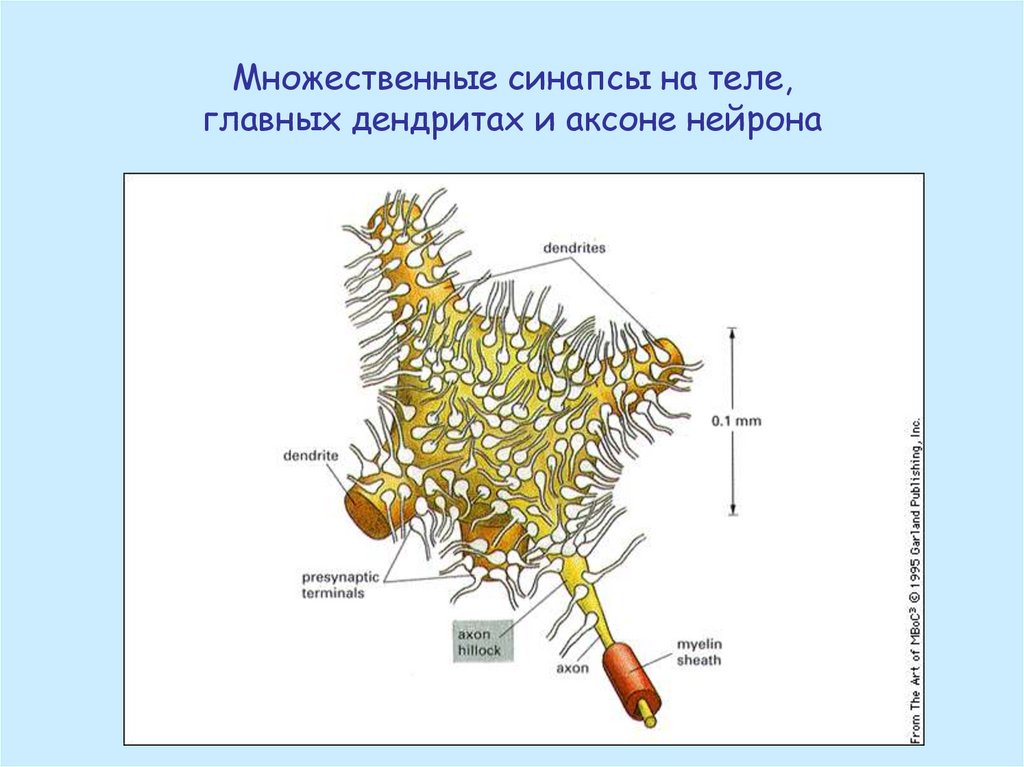
Множественные синапсы на теле,главных дендритах и аксоне нейрона
18.
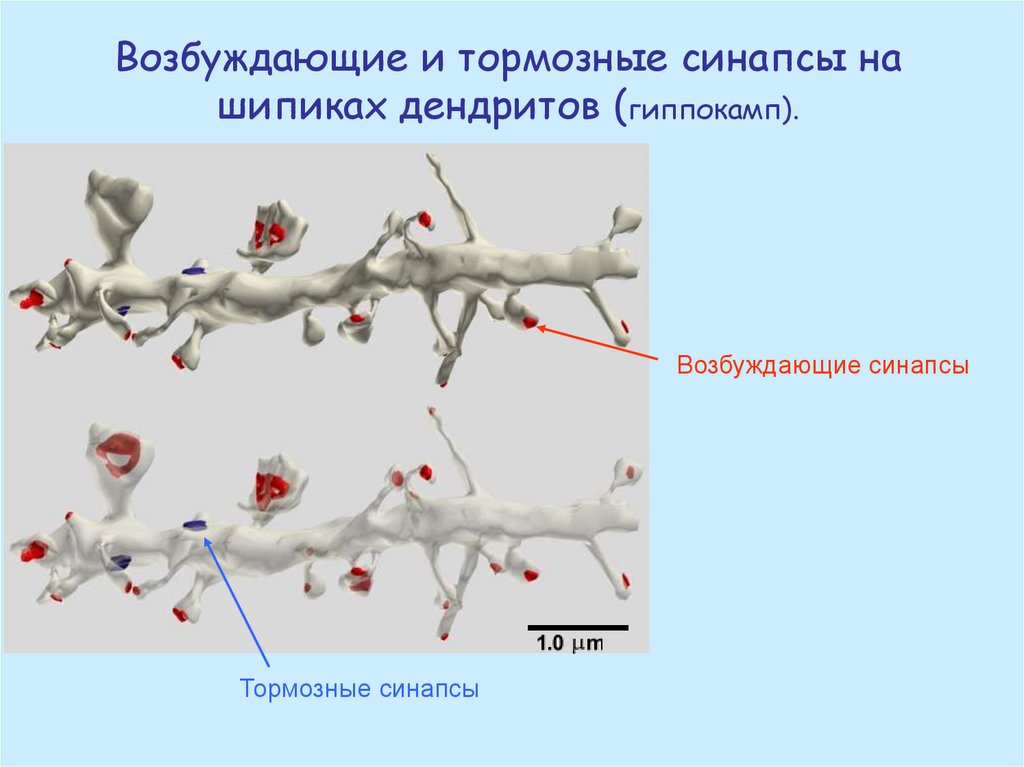
Возбуждающие и тормозные синапсы нашипиках дендритов (гиппокамп).
Возбуждающие синапсы
Тормозные синапсы
19.
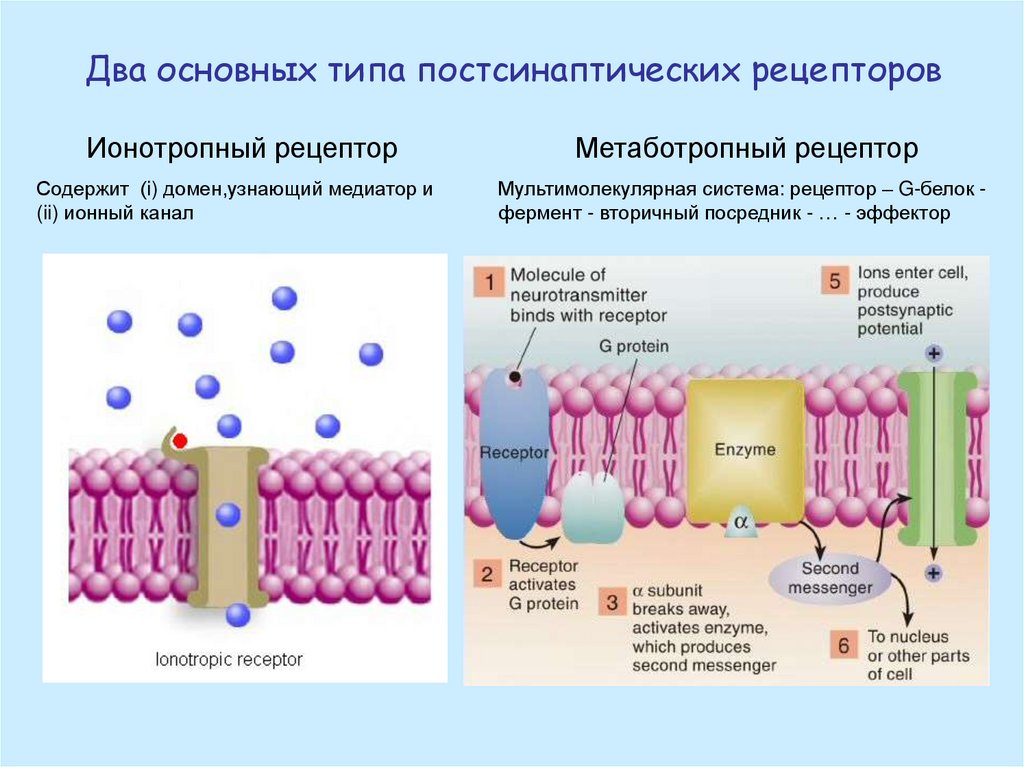
Два основных типа постсинаптических рецепторовИонотропный рецептор
Метаботропный рецептор
Содержит (i) домен,узнающий медиатор и
(ii) ионный канал
Мультимолекулярная система: рецептор – G-белок фермент - вторичный посредник - … - эффектор
20. Ионная природа постсинаптических токов
Возбуждающий постсинаптический ток = ВПСТK+
каналы
Сl –
каналы
Тормозный постсинаптический ток = ТПСТ
Physiology of behavior, Neil R. Carlson
Na+/K+
Ca2+
каналы
Cl –
каналы
21.
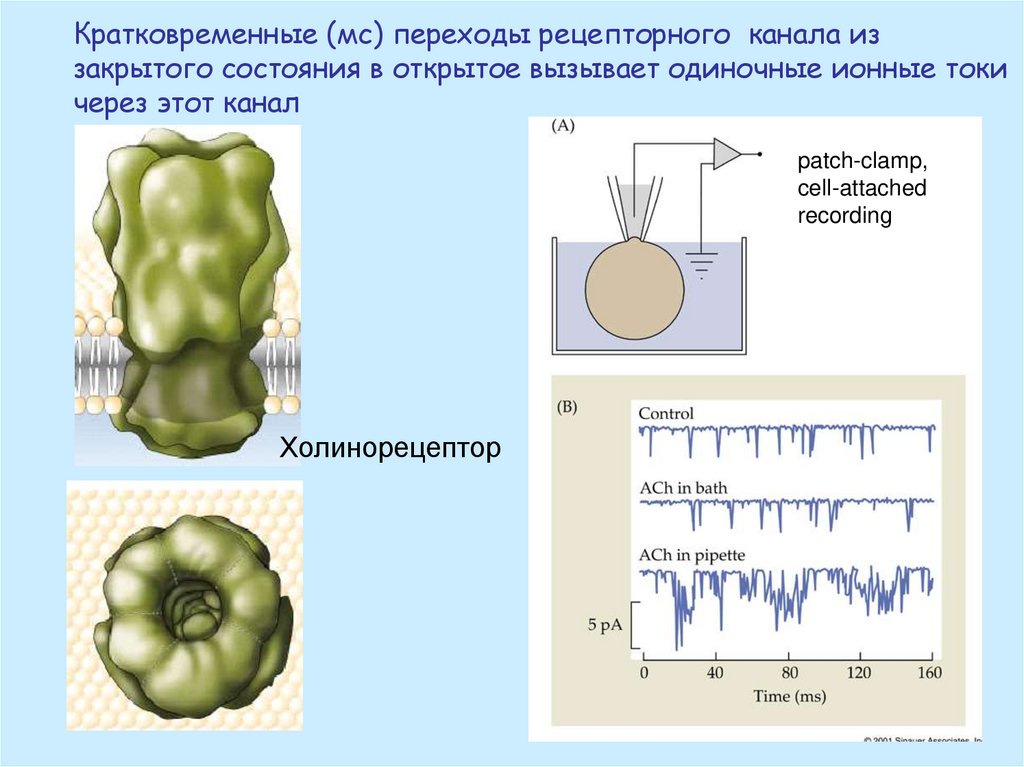
Кратковременные (мс) переходы рецепторного канала иззакрытого состояния в открытое вызывает одиночные ионные токи
через этот канал
patch-clamp,
cell-attached
recording
Холинорецептор
22.
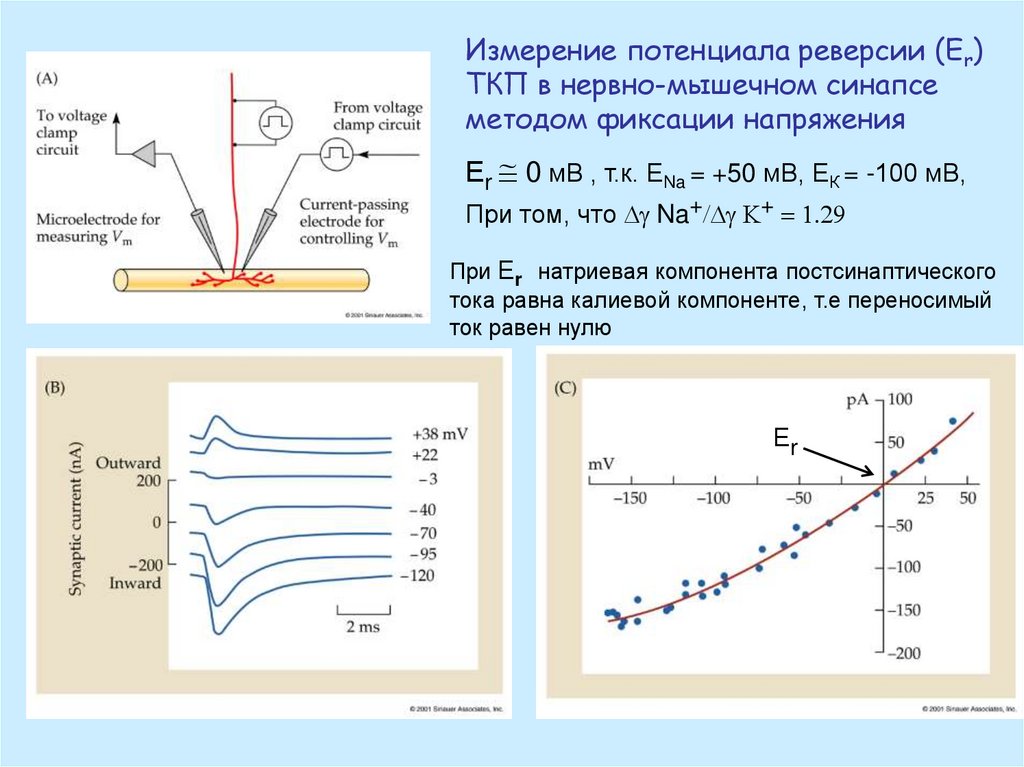
Измерение потенциала реверсии (Еr)ТКП в нервно-мышечном синапсе
методом фиксации напряжения
Еr @ 0 мВ , т.к. ЕNa = +50 мВ, ЕК = -100 мВ,
При том, что g Na+/ g K+ = 1.29
При Еr натриевая компонента постсинаптического
тока равна калиевой компоненте, т.е переносимый
ток равен нулю
Еr
23.
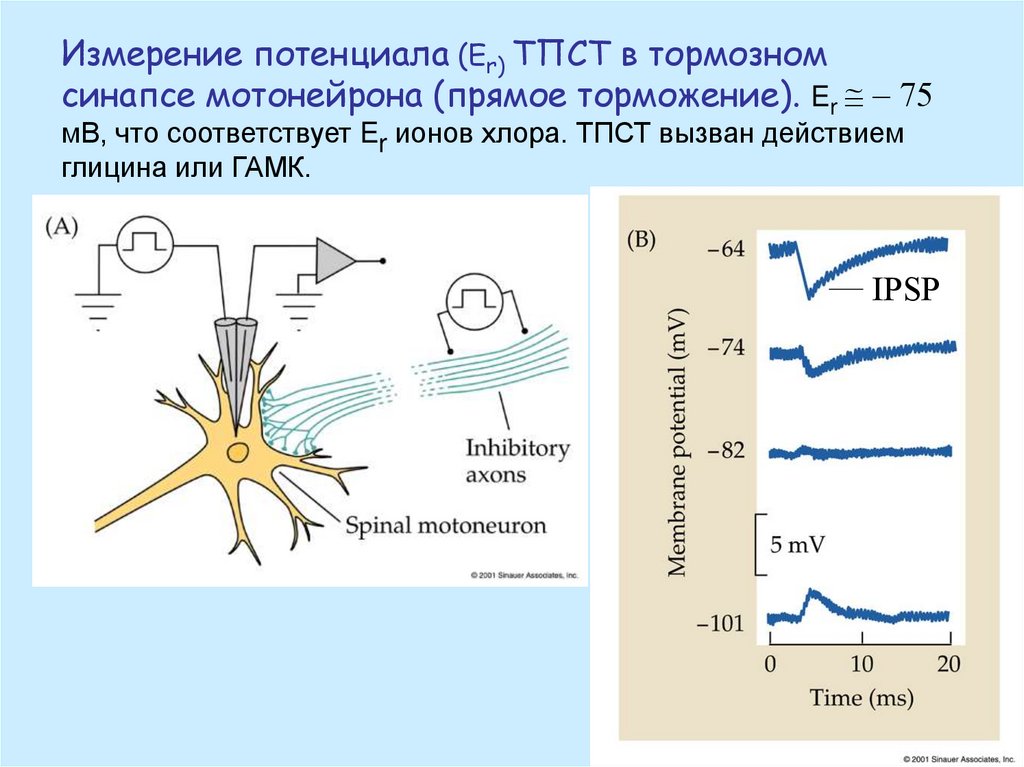
Измерение потенциала (Er) ТПСТ в тормозномсинапсе мотонейрона (прямое торможение). Er @ - 75
мВ, что соответствует Er ионов хлора. ТПСТ вызван действием
глицина или ГАМК.
IPSP
24.
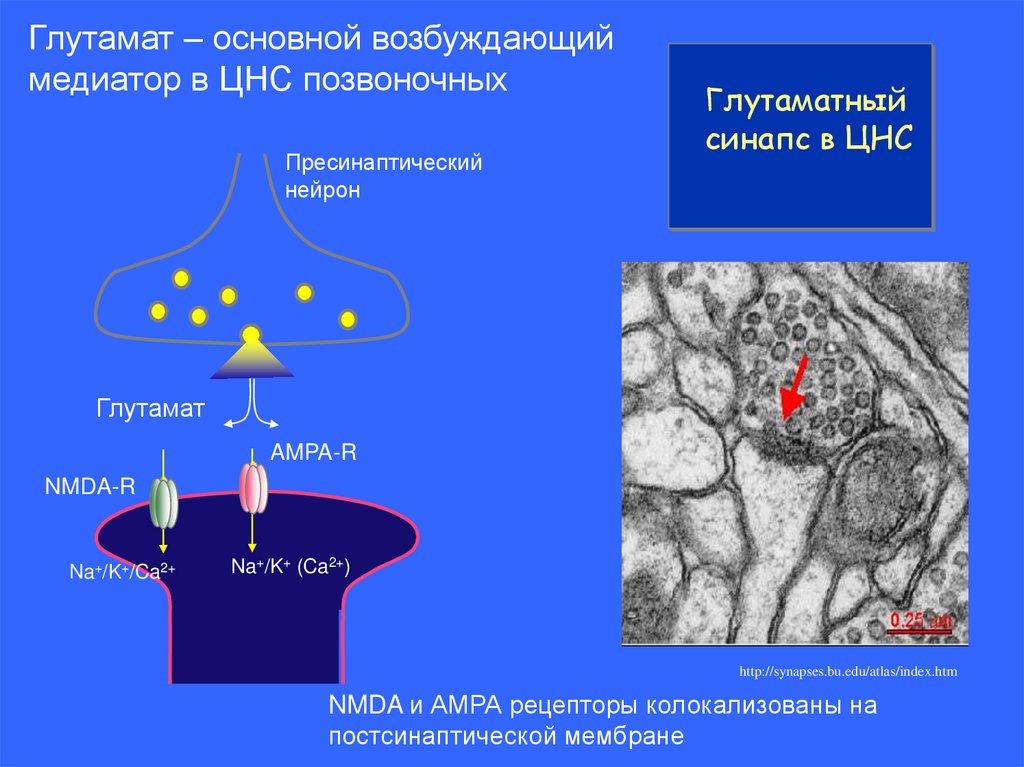
Глутамат – основной возбуждающиймедиатор в ЦНС позвоночных
Пресинаптический
нейрон
Глутаматный
синапс в ЦНС
Глутамат
AMPA-R
NMDA-R
Na+/K+/Ca2+
Na+/K+ (Ca2+)
http://synapses.bu.edu/atlas/index.htm
NMDA и АМРА рецепторы колокализованы на
постсинаптической мембране
25.
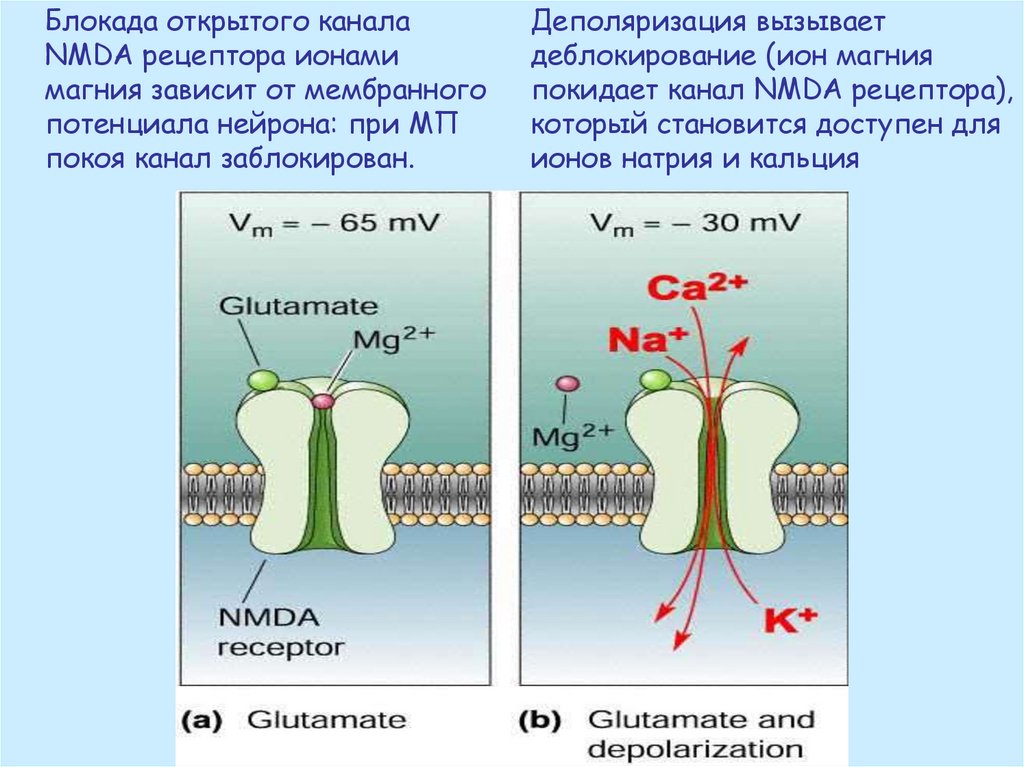
Блокада открытого каналаNMDA рецептора ионами
магния зависит от мембранного
потенциала нейрона: при МП
покоя канал заблокирован.
Деполяризация вызывает
деблокирование (ион магния
покидает канал NMDA рецептора),
который становится доступен для
ионов натрия и кальция
26.
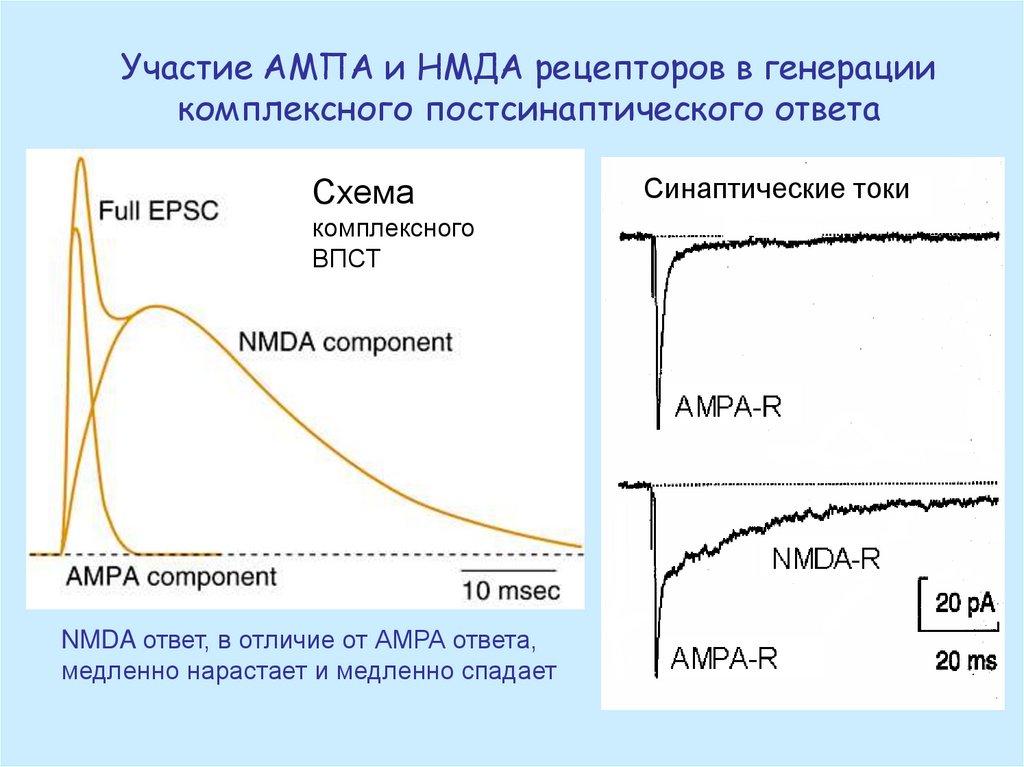
Участие АМПА и НМДА рецепторов в генерациикомплексного постсинаптического ответа
Схема
комплексного
ВПСТ
NMDA ответ, в отличие от АМРА ответа,
медленно нарастает и медленно спадает
Синаптические токи
27.
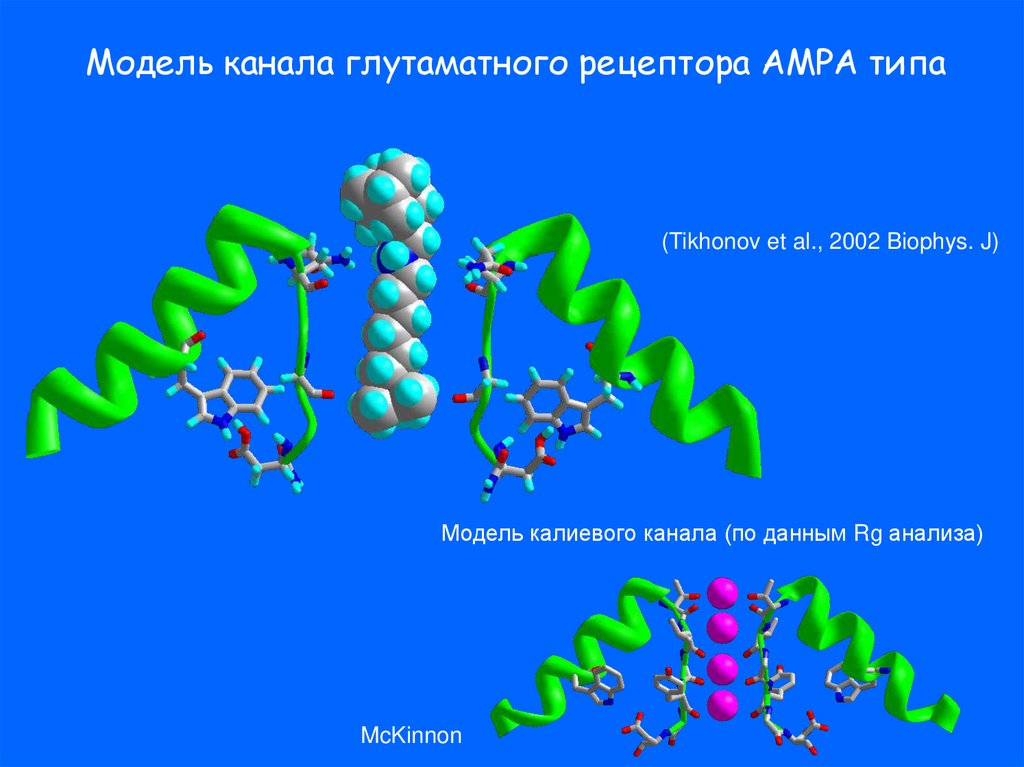
Модель канала глутаматного рецептора АМРА типа(Tikhonov et al., 2002 Biophys. J)
Модель калиевого канала (по данным Rg анализа)
McKinnon
28.
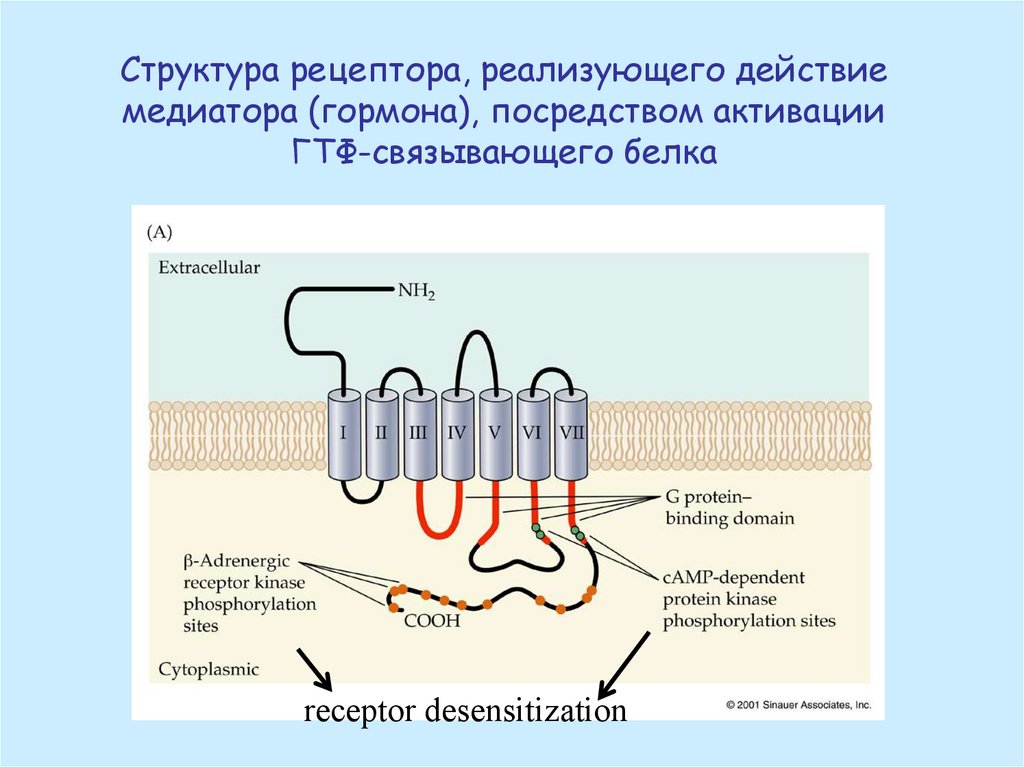
Структура рецептора, реализующего действиемедиатора (гормона), посредством активации
ГТФ-связывающего белка
receptor desensitization
29.
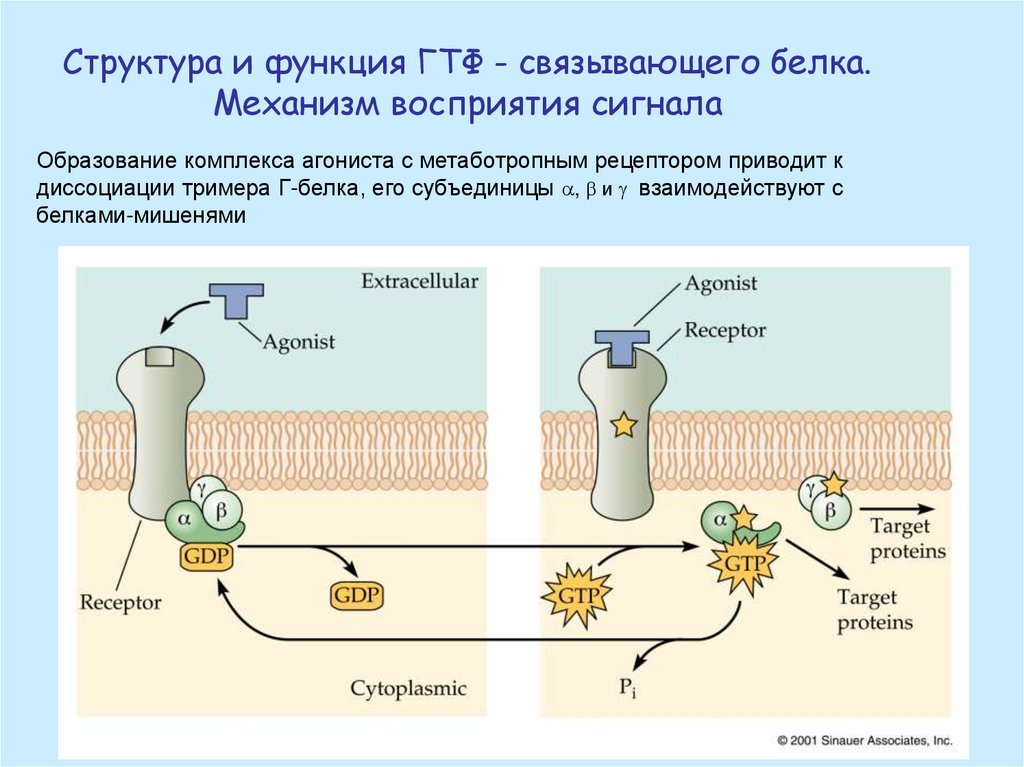
Структура и функция ГТФ - связывающего белка.Механизм восприятия сигнала
Образование комплекса агониста с метаботропным рецептором приводит к
диссоциации тримера Г-белка, его субъединицы , и g взаимодействуют с
белками-мишенями
30.
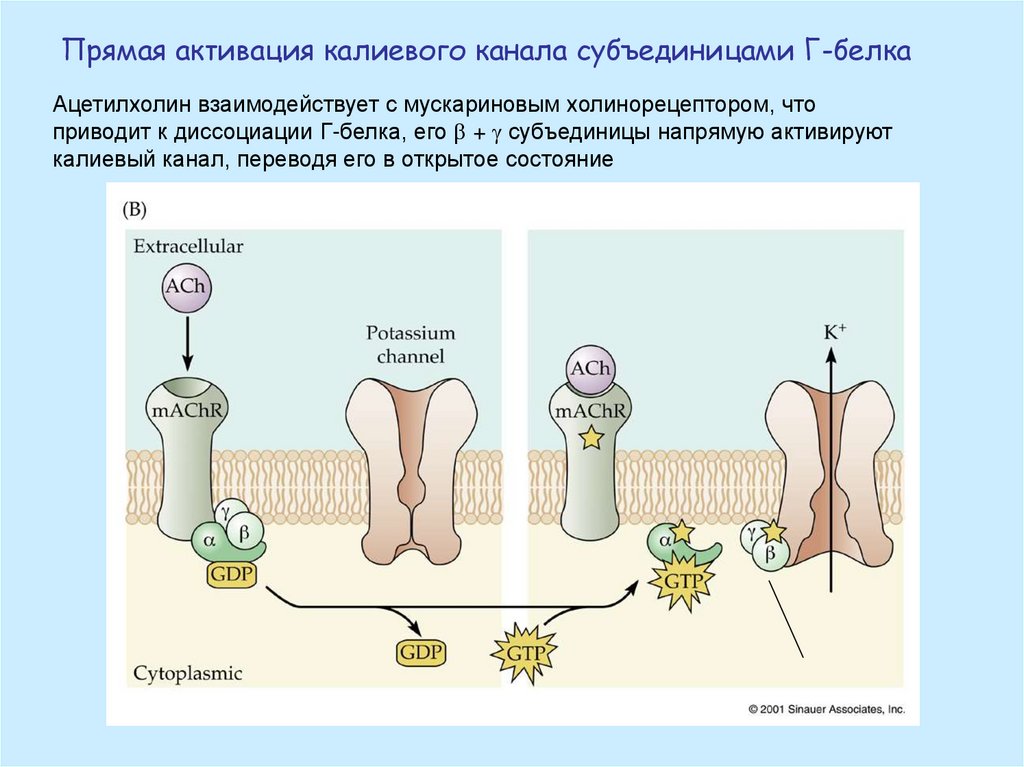
Прямая активация калиевого канала субъединицами Г-белкаАцетилхолин взаимодействует с мускариновым холинорецептором, что
приводит к диссоциации Г-белка, его + g субъединицы напрямую активируют
калиевый канал, переводя его в открытое состояние
31.
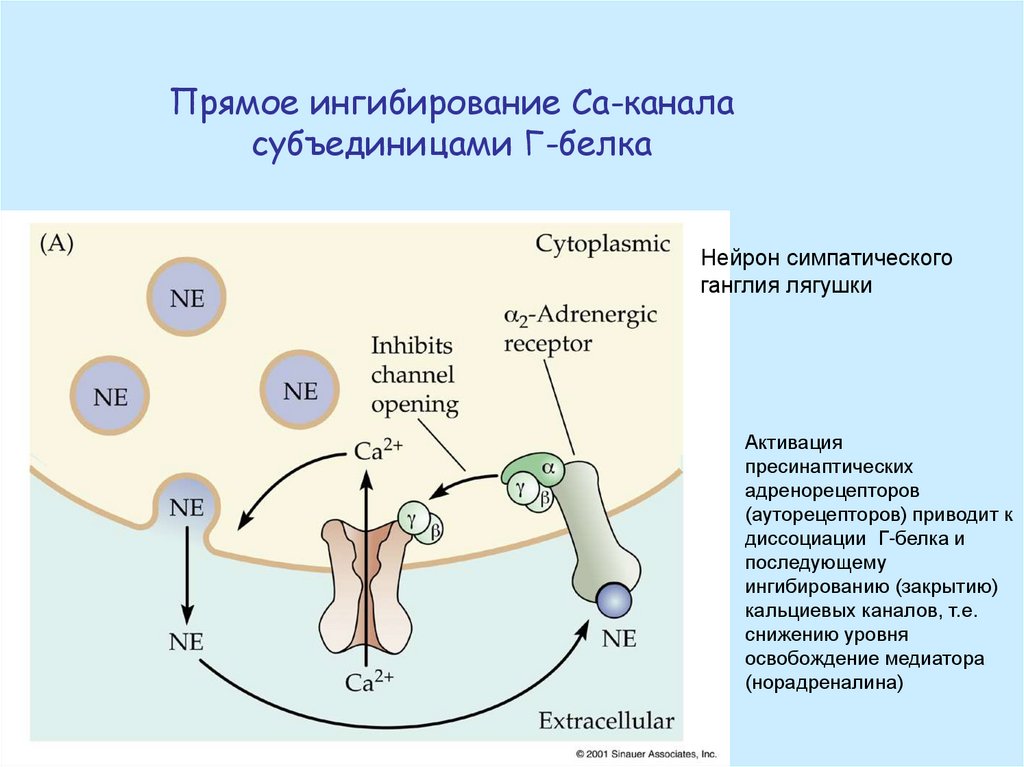
Прямое ингибирование Са-каналасубъединицами Г-белка
Нейрон симпатического
ганглия лягушки
Активация
пресинаптических
адренорецепторов
(ауторецепторов) приводит к
диссоциации Г-белка и
последующему
ингибированию (закрытию)
кальциевых каналов, т.е.
снижению уровня
освобождение медиатора
(норадреналина)
32.
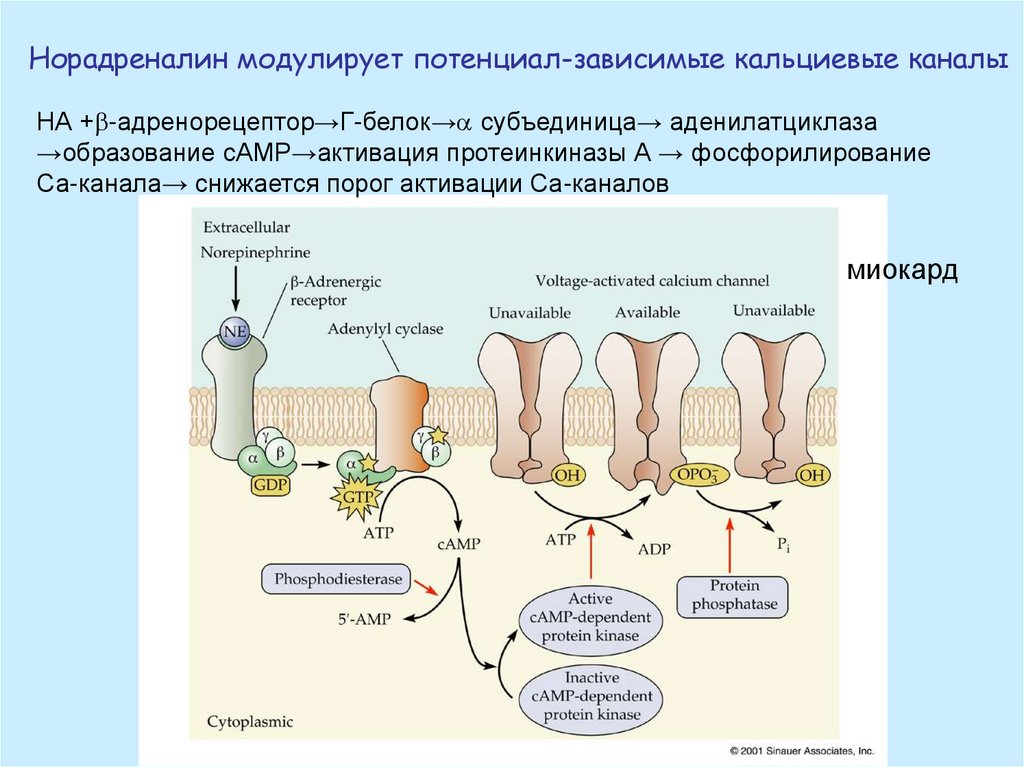
Норадреналин модулирует потенциал-зависимые кальциевые каналыНА + -адренорецептор→Г-белок→ субъединица→ аденилатциклаза
→образование сАМР→активация протеинкиназы А → фосфорилирование
Са-канала→ снижается порог активации Са-каналов
миокард
33.
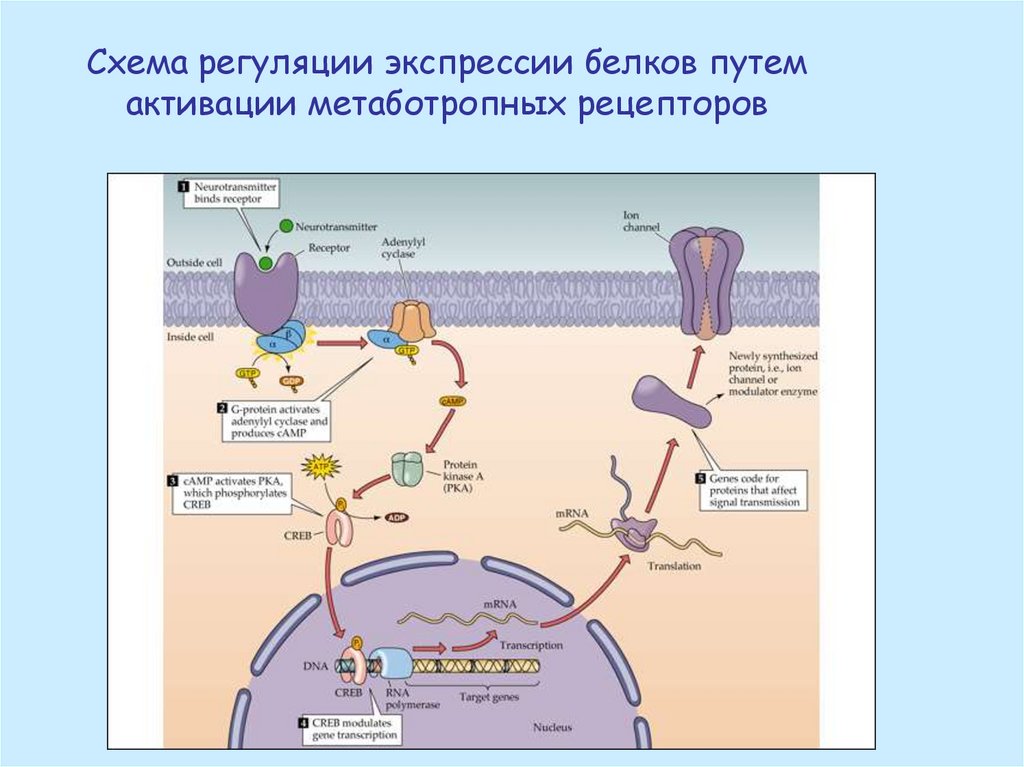
Схема регуляции экспрессии белков путемактивации метаботропных рецепторов
34.
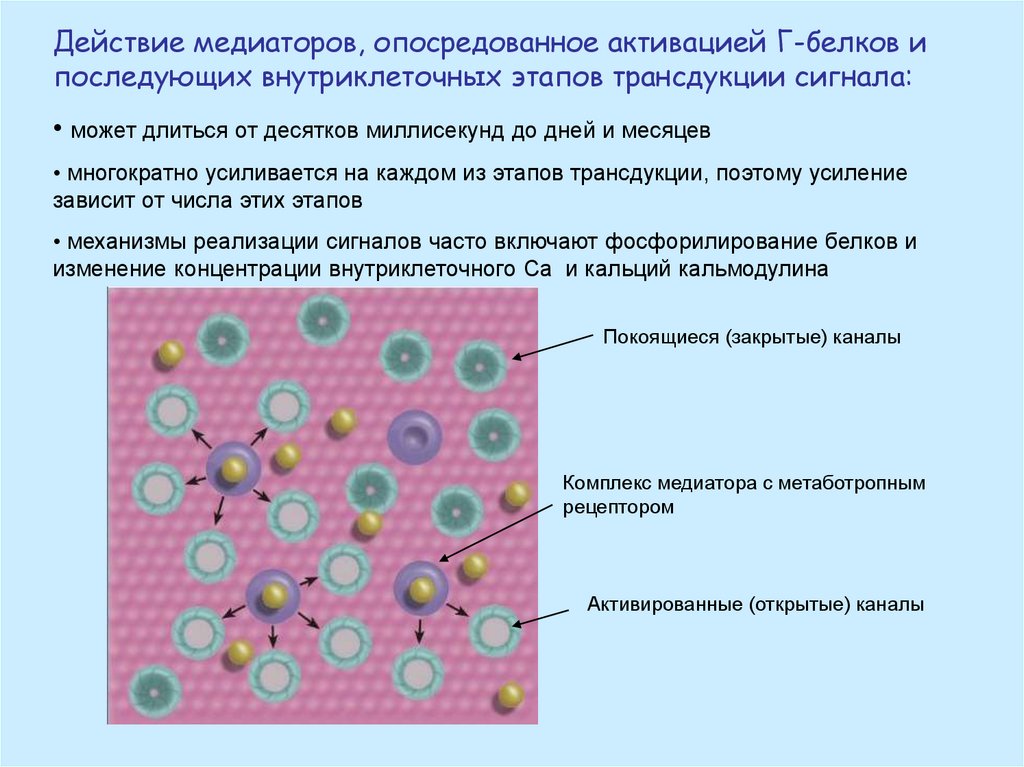
Действие медиаторов, опосредованное активацией Г-белков ипоследующих внутриклеточных этапов трансдукции сигнала:
• может длиться от десятков миллисекунд до дней и месяцев
• многократно усиливается на каждом из этапов трансдукции, поэтому усиление
зависит от числа этих этапов
• механизмы реализации сигналов часто включают фосфорилирование белков и
изменение концентрации внутриклеточного Са и кальций кальмодулина
Покоящиеся (закрытые) каналы
Комплекс медиатора с метаботропным
рецептором
Активированные (открытые) каналы
35.
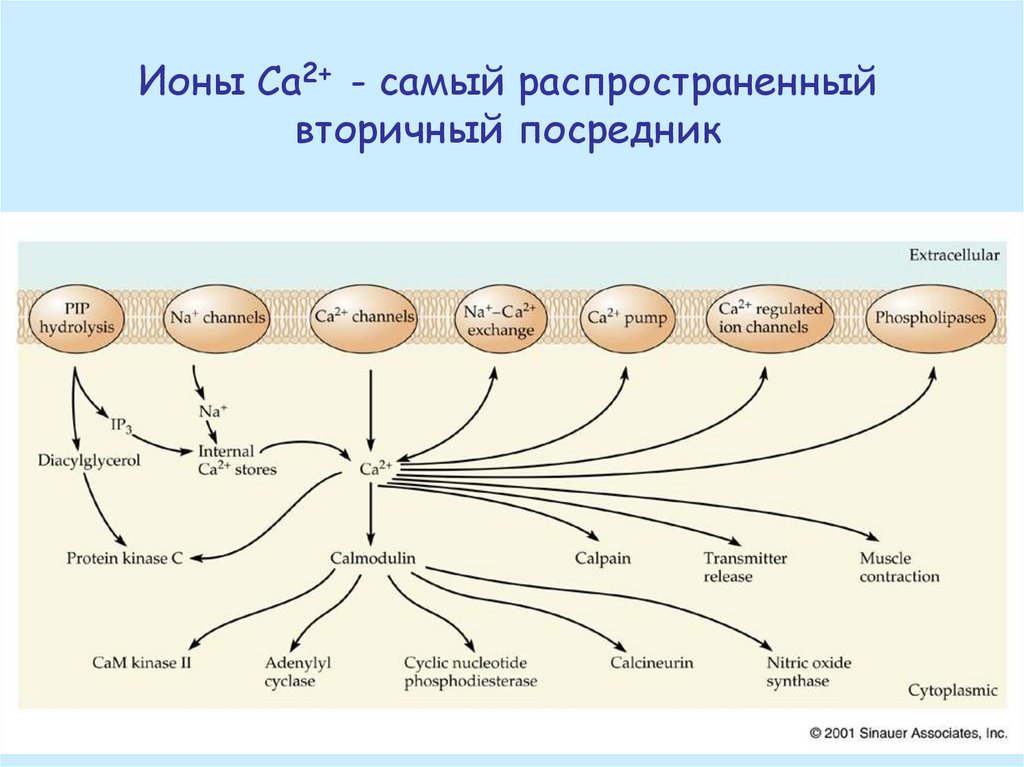
Ионы Са2+ - самый распространенныйвторичный посредник
36.
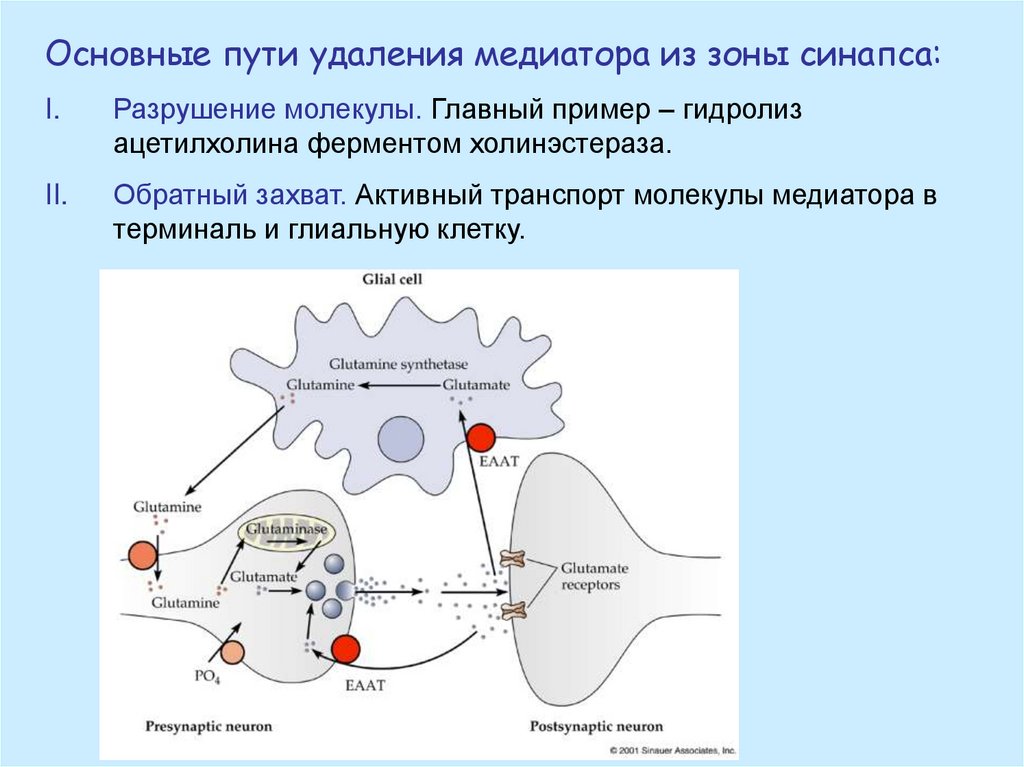
Основные пути удаления медиатора из зоны синапса:I.
Разрушение молекулы. Главный пример – гидролиз
ацетилхолина ферментом холинэстераза.
II.
Обратный захват. Активный транспорт молекулы медиатора в
терминаль и глиальную клетку.
37.
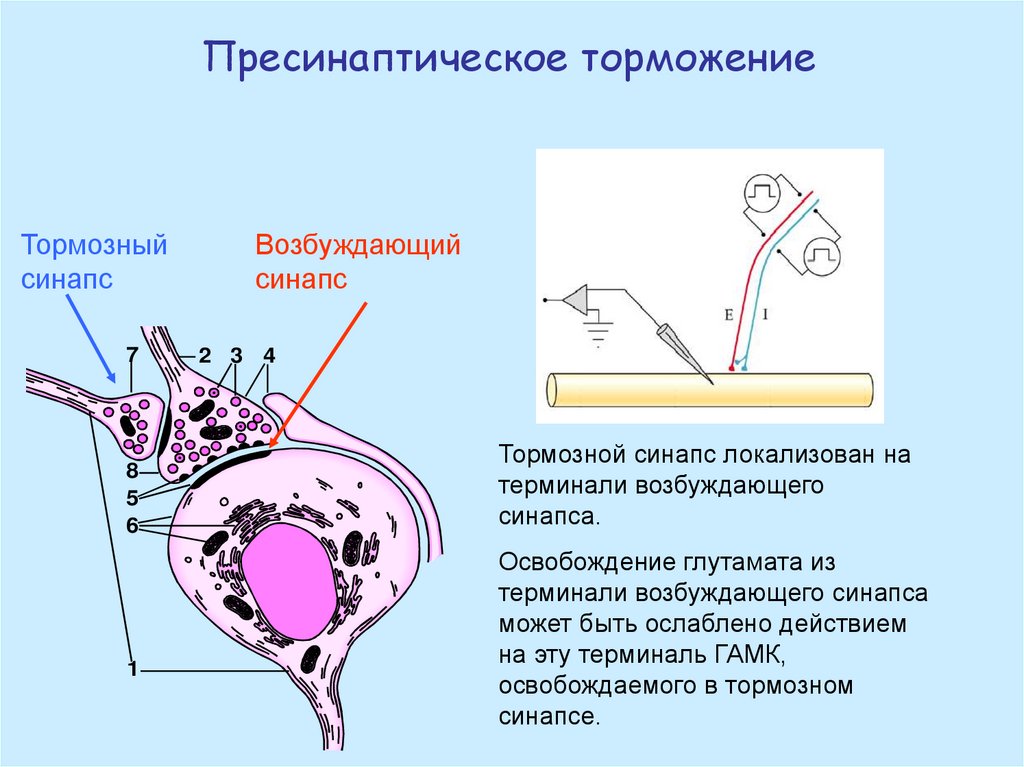
Пресинаптическое торможениеТормозный
синапс
7
8
5
6
1
Возбуждающий
синапс
2 3 4
Тормозной синапс локализован на
терминали возбуждающего
синапса.
Освобождение глутамата из
терминали возбуждающего синапса
может быть ослаблено действием
на эту терминаль ГАМК,
освобождаемого в тормозном
синапсе.
38.
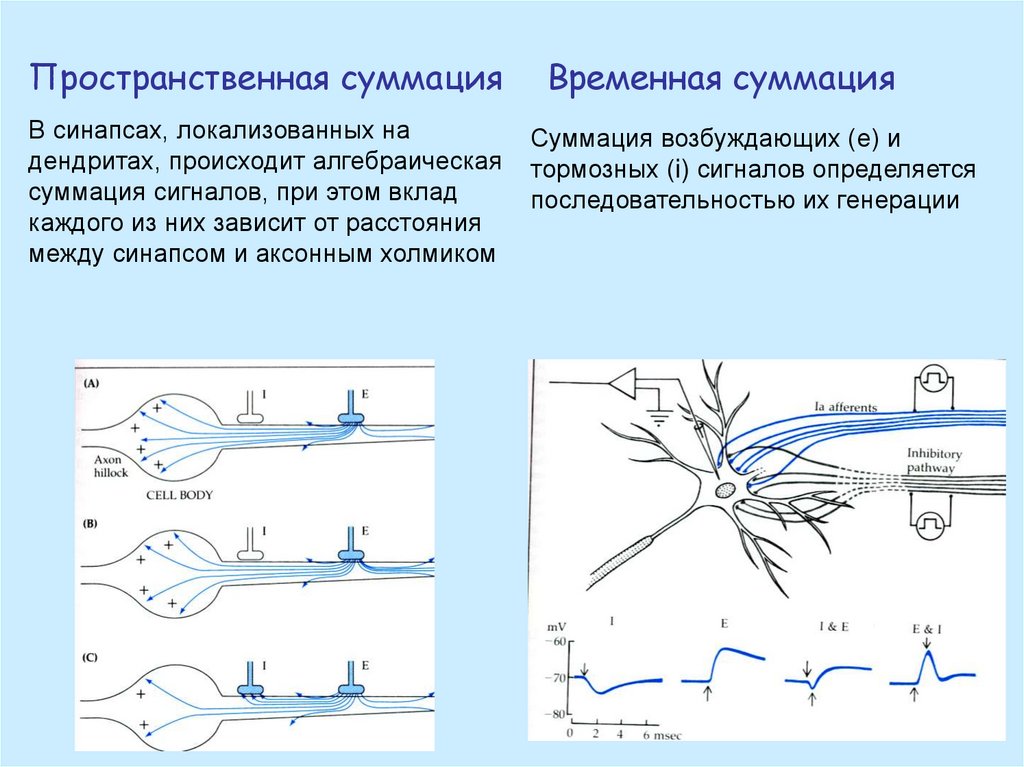
Пространственная суммацияВременная суммация
В синапсах, локализованных на
дендритах, происходит алгебраическая
суммация сигналов, при этом вклад
каждого из них зависит от расстояния
между синапсом и аксонным холмиком
Суммация возбуждающих (е) и
тормозных (i) сигналов определяется
последовательностью их генерации
39.
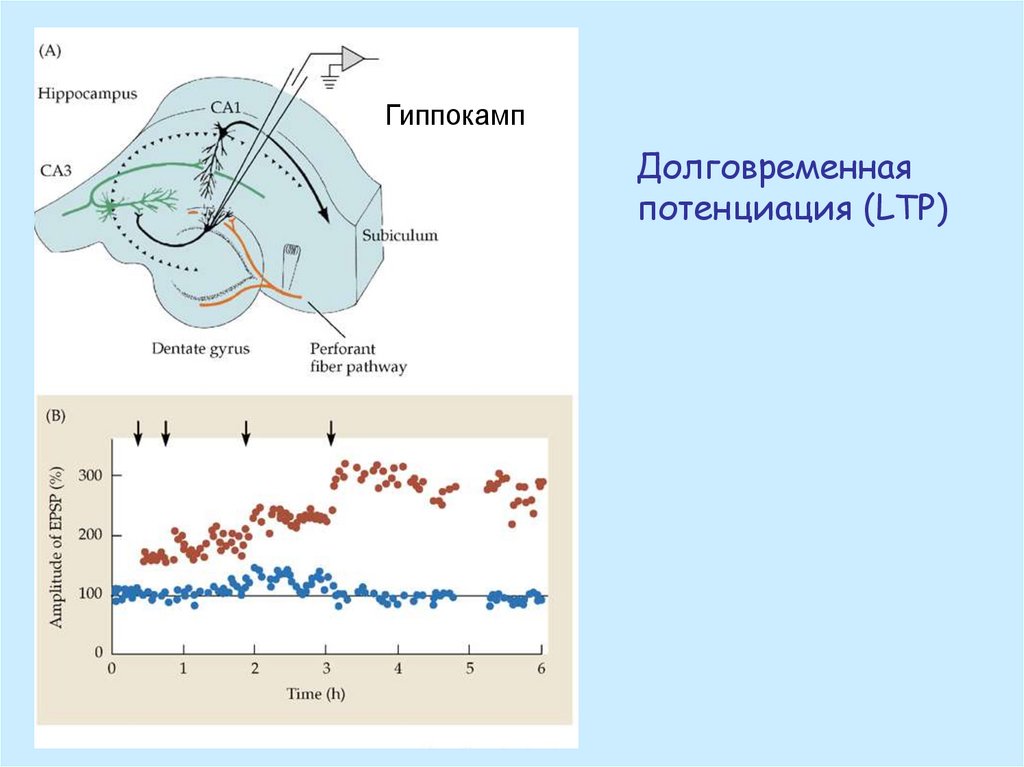
ГиппокампДолговременная
потенциация (LTP)
40.
S.Ramon y Cajal (1852 -1934)Bernard Katz (1911-2002)