



















")














 вируса")





 Биология
БиологияПохожие презентации:










Взаимодействие «Вирус-клетка»
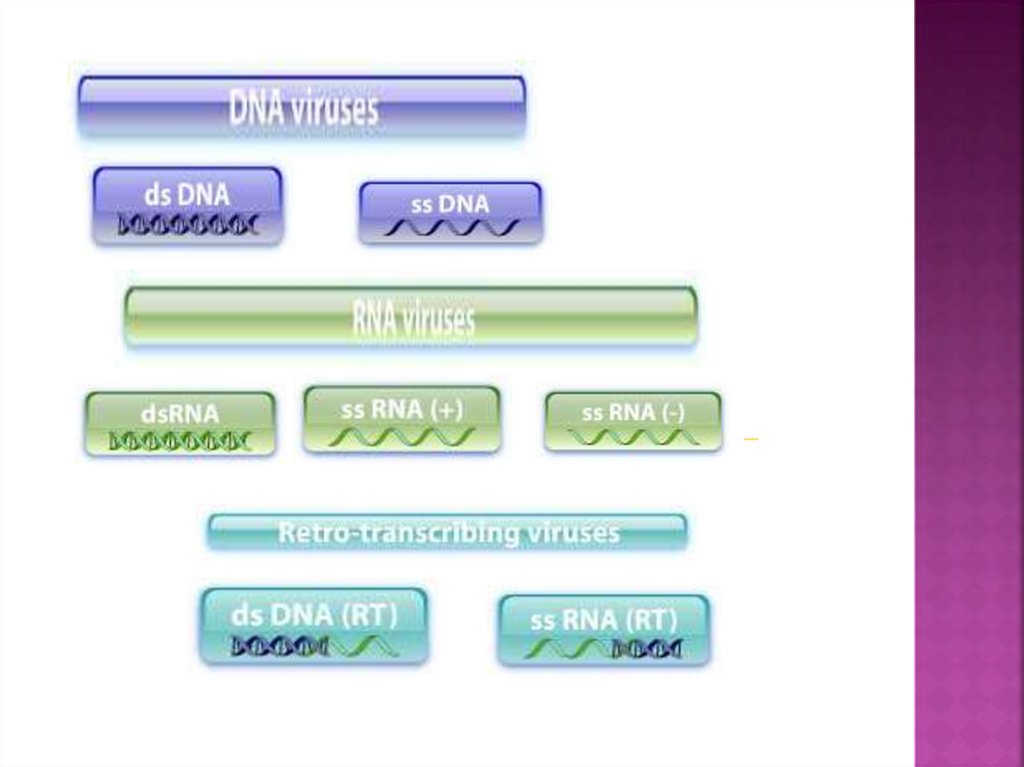
1.
Взаимодействие«Вирус-клетка»
1
2.
3.
SIB Swiss Institute of Bioinformatics | Disclaimer3
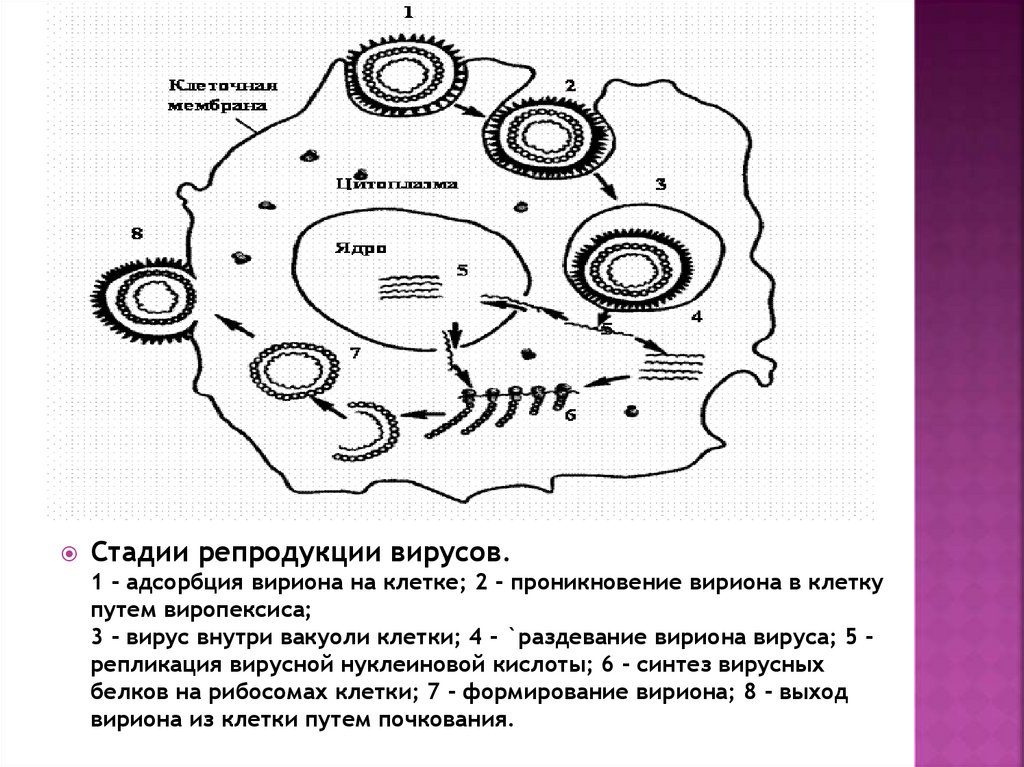
4. Стадии репродукции вирусов
Адсорбция2. Проникновение вируса в клетку
(пенетрация).
3. «Раздевание" (депротеинизация)
4. Транскрипция
5. Трансляция
6. Репликация
7. Сборка вириона
8. Выход вируса из клетки
1.
5.
Стадии репродукции вирусов.1 - адсорбция вириона на клетке; 2 - проникновение вириона в клетку
путем виропексиса;
3 - вирус внутри вакуоли клетки; 4 - `раздевание вириона вируса; 5 репликация вирусной нуклеиновой кислоты; 6 - синтез вирусных
белков на рибосомах клетки; 7 - формирование вириона; 8 - выход
вириона из клетки путем почкования.
6. Адсорбция
Неспецифическая – за счетэлектростатического взаимодействия
Специфическая – за счет связи между
комплементарными клеточными и
вирусными рецепторами:
Обратимая фаза
Необратимая фаза
7. Адсорбция вируса на клетке
Присоединение к клетке-хозяину опосредуетсяпротеинами вирионов с определенными молекулами
поверхности хозяина (мембранные белки, липиды
или углеводные фрагменты, присутствующие либо
на гликопротеидах, либо гликолипидах.
8. адгезия
Адгезионные рецепторы прикрепляют вирусобратимым образом к клеткам или органам.
Не является обязательной для проникновения
вируса.
Тем не менее адгезия значительно повышает
инфекционную активность, концентрируя вирус
в непосредственной близости от его входных
рецепторов.
Эти рецепторы запускают проникновение вируса
эндоцитозом / пиноцитозом или путем
индуцирования слияния / проникновения, и
последствия этого связывания необратимы.
Такие рецепторы часто называют
«корецепторами».
9.
Способыпроникновения
вируса в клетку
9
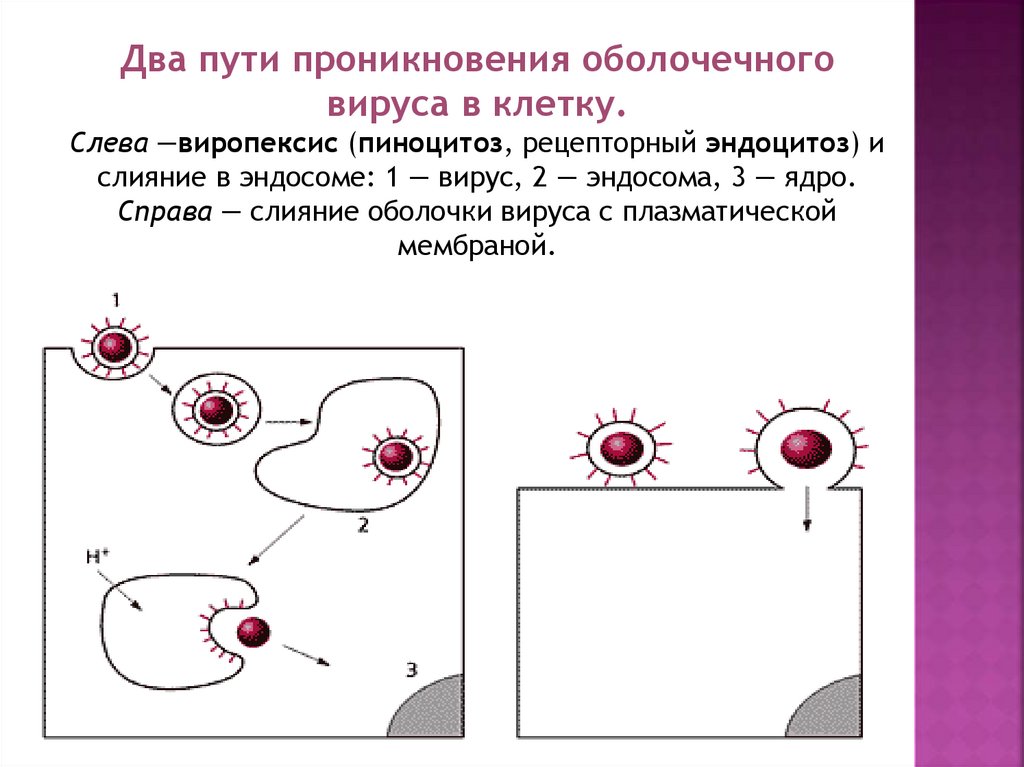
10.
Два пути проникновения оболочечноговируса в клетку.
Слева —виропексис (пиноцитоз, рецепторный эндоцитоз) и
слияние в эндосоме: 1 — вирус, 2 — эндосома, 3 — ядро.
Справа — слияние оболочки вируса с плазматической
мембраной.
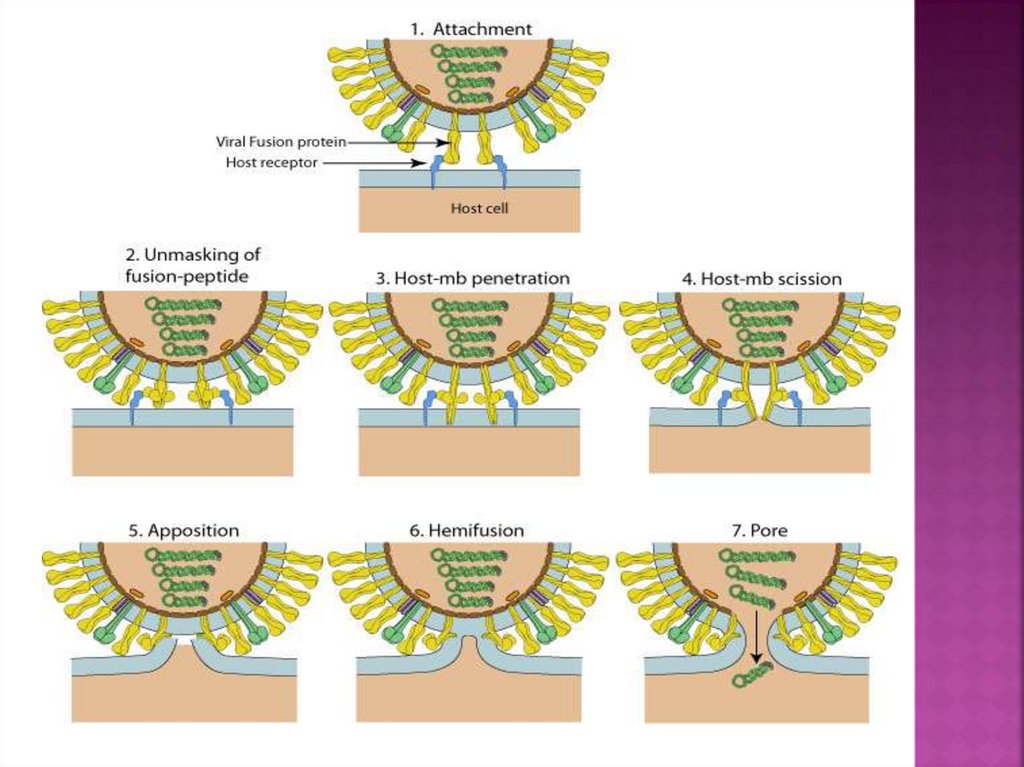
11.
12. Слияние оболочки вируса и мембраны клетки
13.
Слияние между вирусной и клеточноймембраной инициируется белками сияния
(Fusion).
Эти белки активны в тримерной
конформации.
Большинство из них являются изначально
тримерными, некоторые из них являются
димерами на поверхности вириона, но при
активации преобразуются в тримеры.
14.
15.
Эндоцито́з (англ. endocytosis) — процессзахвата (интернализации) внешнего
материала клеткой, осуществляемый путём
образования мембранных везикул. В
результате эндоцитоза клетка получает для
своей жизнедеятельности гидрофильный
материал, который иначе не проникает через
липидный бислой клеточной мембраны.
Различают фагоцитоз, пиноцитоз и рецепторопосредованный эндоцитоз. Термин был
предложен в 1963 году бельгийским
цитологом Кристианом де Дювом для
описания множества процессов
интернализации, развившихся в клетке
млекопитающих .
16.
Рецептор-опосредованный эндоцитоз— активныйспецифический процесс, при котором клеточная
мембрана выпучивается внутрь клетки, формируя
окаймлённые ямки. Внутриклеточная сторона
окаймлённой ямки содержит набор адаптивных
белков (адаптин, клатрин, обуславливающий
необходимую кривизну выпучивания, и др. белки).
Макромолекулы, связывающиеся со специфическими
рецепторами на поверхности клетки, проходят внутрь
со значительно большей скоростью, чем вещества,
поступающие в клетки за счет пиноцитоза. Внешняя
сторона мембраны при этом включает
специфические рецепторы. При связывании лиганда
из окружающей клетку среды окаймлённые ямки
формируют внутриклеточные везикулы (окаймлённые
пузырьки).
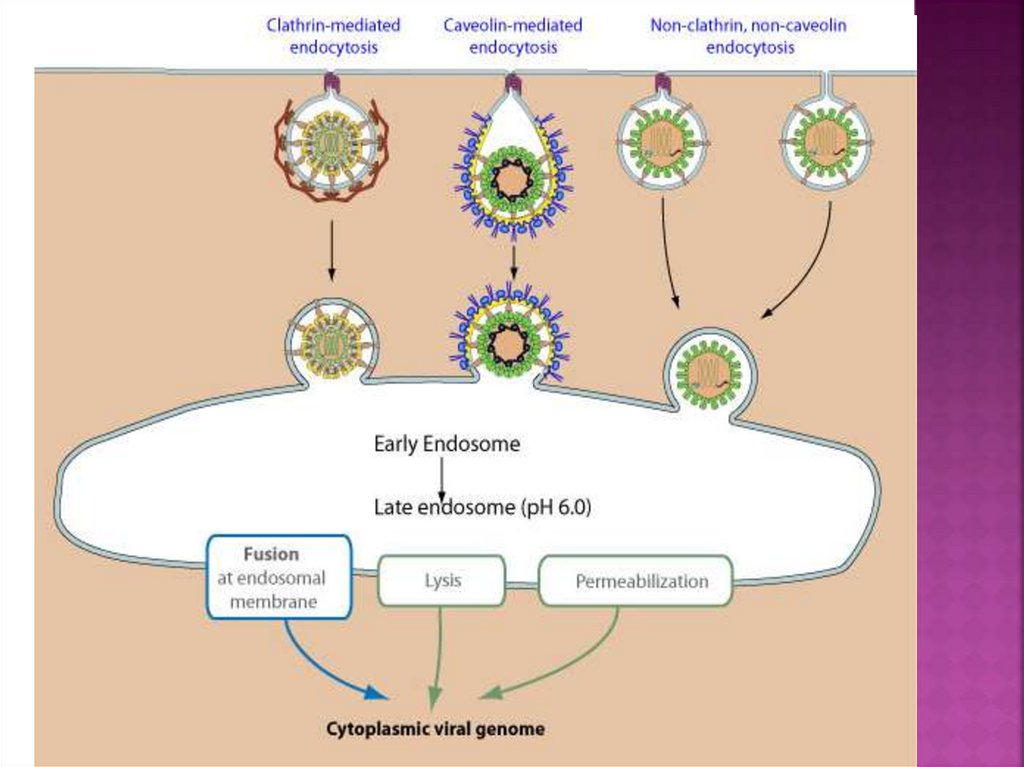
17. Клатрин-зависимый эндоцитоз
18. Клатрин-зависимый эндоцитоз
клатрин-опосредованный эндоцитоз (сlathrinmediated endoycytosis - CME) вызван связываниемрецепторов вириона с рецепторами клетки-хозяина.
Индуцируется связывание переходного белка с
рецепторным «цитоплазматическим хвостом».
Адапторные белки связываются с клатрином, а
локальная концентрация переходных белков на
внутренней поверхности плазматической мембраны
позволяет мультимеризовать клатрин с
образованием характерных инвагинаций или ямок,
покрытых клатрином (CCP). Мембранные
расщепляющие белки DNM1 / Dynamin-1 или DNM2 /
Dynamin-2 отжимают от CCP от мембраны хозяина,
тем самым высвобождая Clathrin-Coated Vesicle
(CCV).
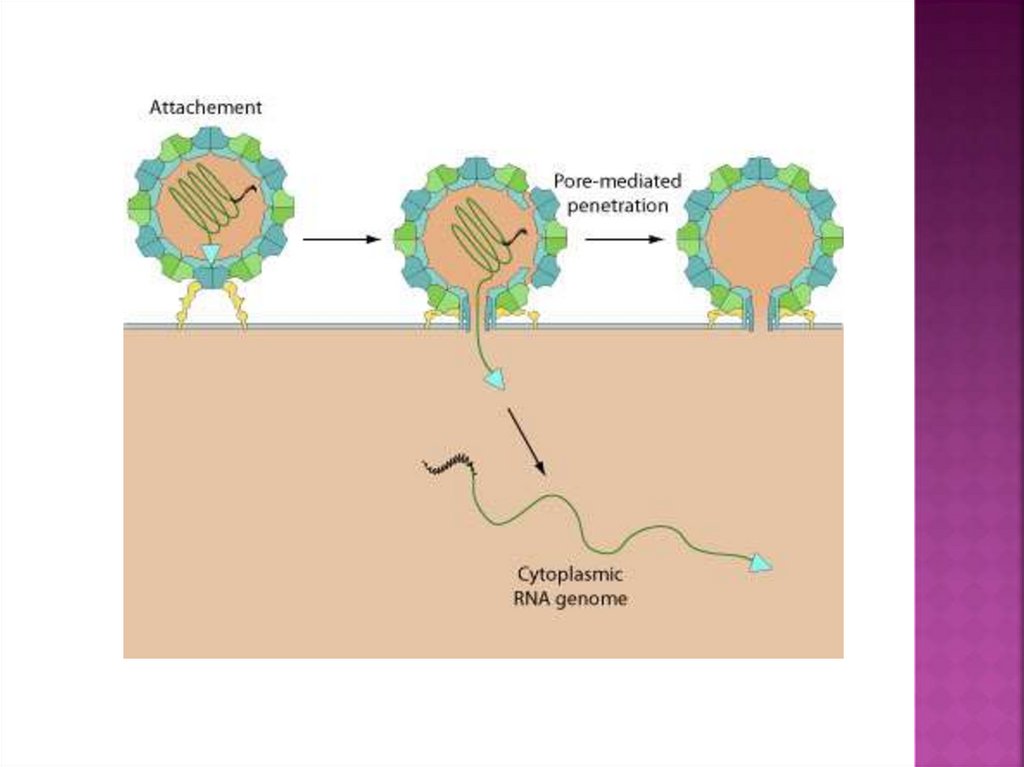
19. проникновение безоболочечных вирусов клатрин-опосредованным путём
20.
Безоболочечныевирусы переносят свой
генетический материал из эндосомного
компартмента в клеточную цитоплазму через
мембранную пермеабилизацию
(проницаемость) или лизис.
Лизис происходит, когда вирусный капсид
вызывает разрыв эндосомной мембраны.
Этот процесс запускается после
конформационного изменения белка
капсида, вызванного эндосомным кислым рН
или связыванием рецептора. Результатом
является либо выход вирусных частиц в
цитоплазму (аденовирус 2 и 5), либо
высвобождение вирусного генома в
цитоплазму (риновирус человека).
21.
Кавеолины — группа мембранныхбелков рецептор-зависимого
эндоцитоза. У позвоночных
животных существует 3 типа
кавеолина со сходной структурой:
кавеолин-1, -2 и -3. В клетке
кавеолин собирается в олигомеры,
связывает холестерин и
сфинголипиды в определённых
участках клеточной мембраны, что
приводит к формированию
кавеолы.
22. Кавеоин-зависимый эндоцитоз
23.
24. Слияние оболочки вириона и эндосомы
25.
26.
27. Липид-опосредованный эндоцитоз (Polyomaviridae)
Polyomaviridae проникают в клетку-хозяин путемсвязывания с конкретными сфинголипидами.
Интернализация обусловлена необычными
эндоцитарными механизмами, включающими
липидные «плоты» и мембранную инвагинацию.
28. макропиноцитоз
Многие вирусыиспользуют этот
неспецифический
маршрут, присоединяясь
к поверхности клетки и
вызывая
макропиноцитарный
сигнал.
Поглощение
макропиносомой
происходит в
произвольных местах на
поверхности клетки.
Любой вирус, связанный
или нет с активирующим
рецептором, имеет шанс
для интернализации.
29.
30.
Способывыхода
вируса из
клетки
30
31. Взрывной путь
Из погибающей клеткиодновременно выходит
большое количество
вирионов. По взрывному пути
выходят из клетки просто
устроенные вирусы, не
имеющие липопротеиновой
оболочки.
32. Почкование, экзоцитоз
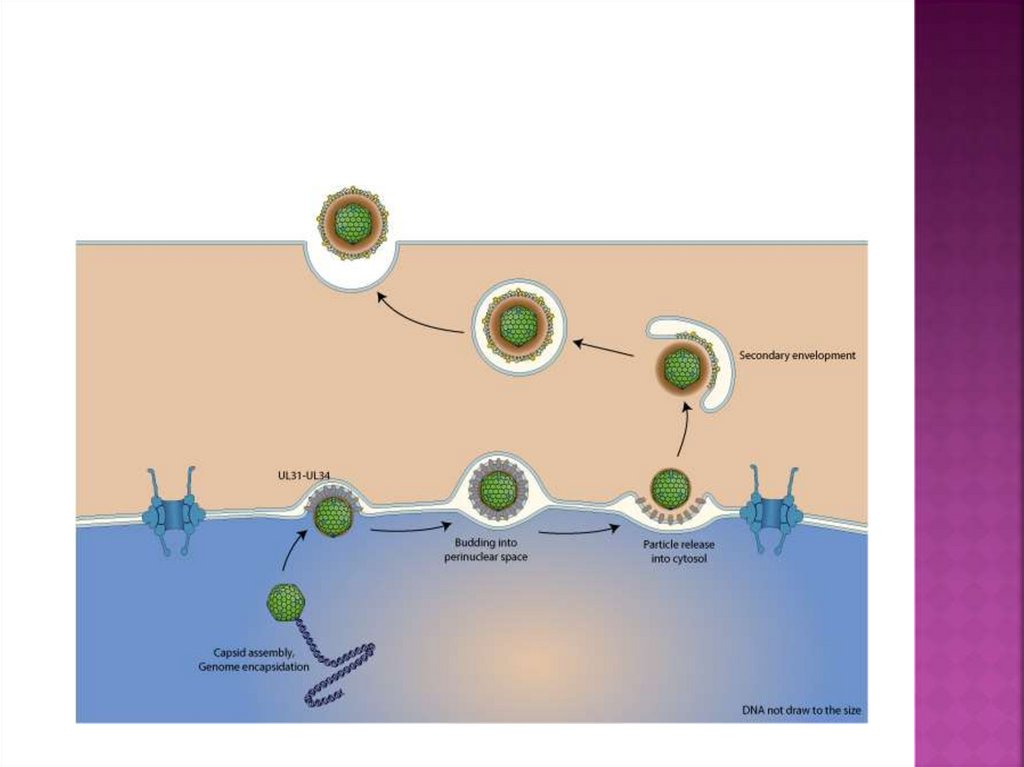
присущи вирусам, имеющим липопротеиновуюоболочку, которая является производной от
клеточных мембран. Сначала образовавшийся
нуклеокапсид или сердцевина вириона
транспортируется к клеточным мембранам, в
которые уже встроены вирусоспецифические
белки. Затем в области контакта нуклеокапсида
или сердцевины вириона с клеточной мембраной
начинается выпячивание этих участков.
Сформировавшаяся почка отделяется от клетки в
виде сложно устроенного вируса. При этом
клетка способна длительно сохранять
жизнеспособность и продуцировать вирусное
потомство.
33.
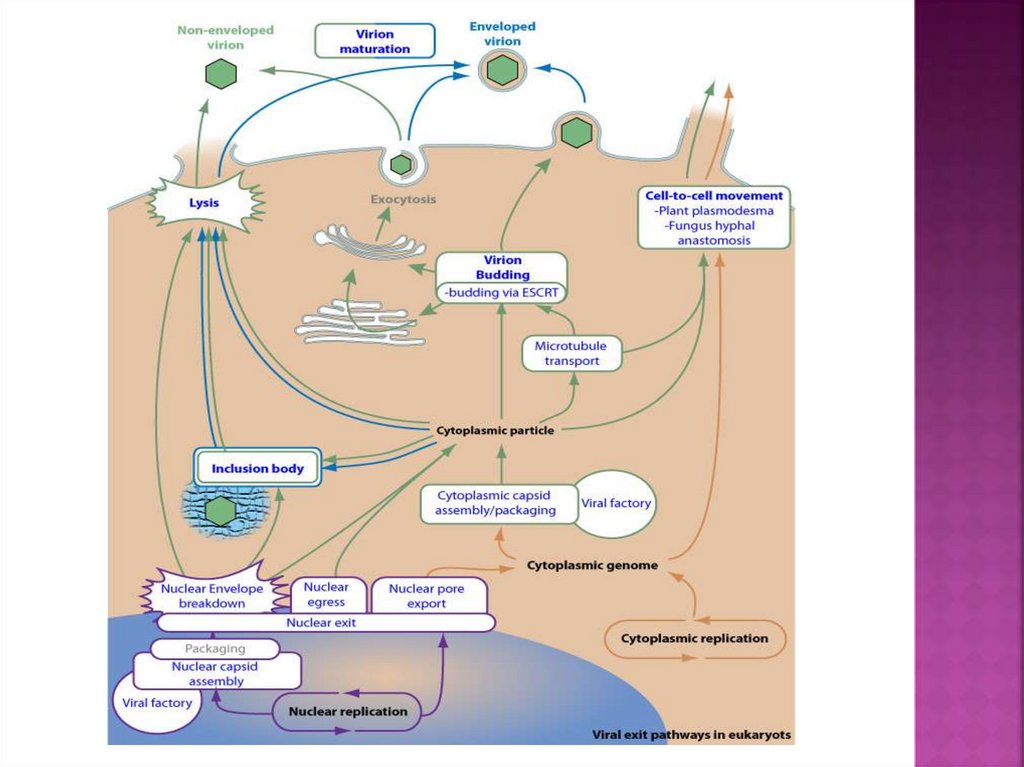
34.
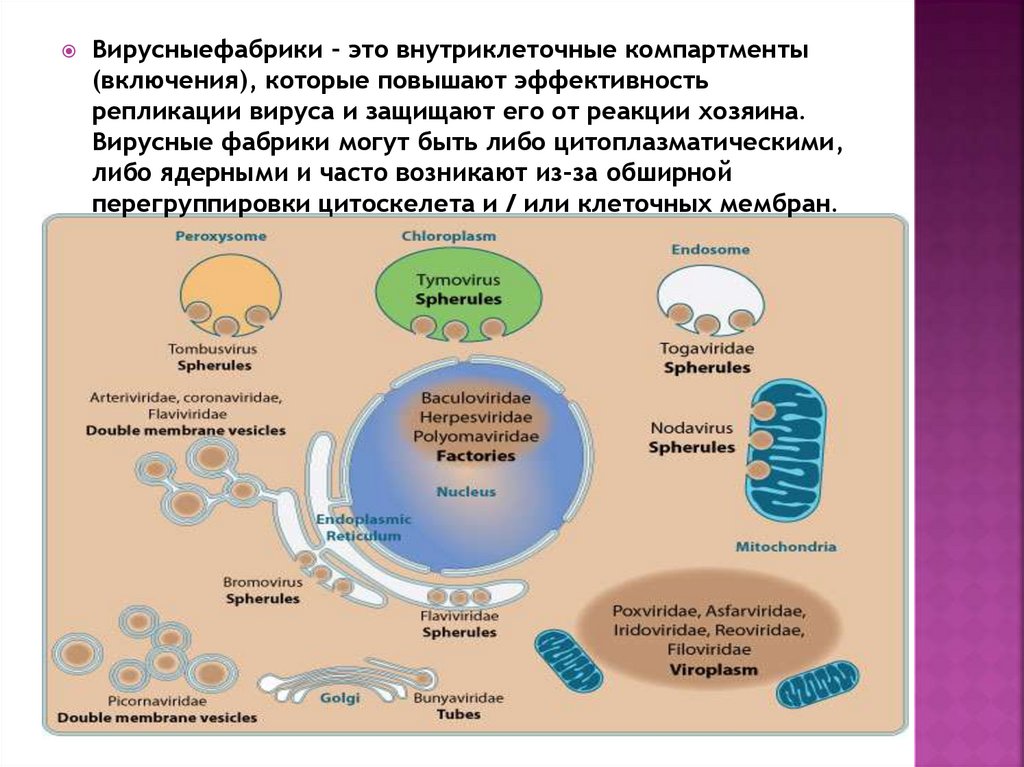
Вирусныефабрики - это внутриклеточные компартменты(включения), которые повышают эффективность
репликации вируса и защищают его от реакции хозяина.
Вирусные фабрики могут быть либо цитоплазматическими,
либо ядерными и часто возникают из-за обширной
перегруппировки цитоскелета и / или клеточных мембран.
35. Сборка цитоплазматических капсидов
36. Ядерное ассамблирование
37.
38. Экспорт генома через ядерные поры
39.
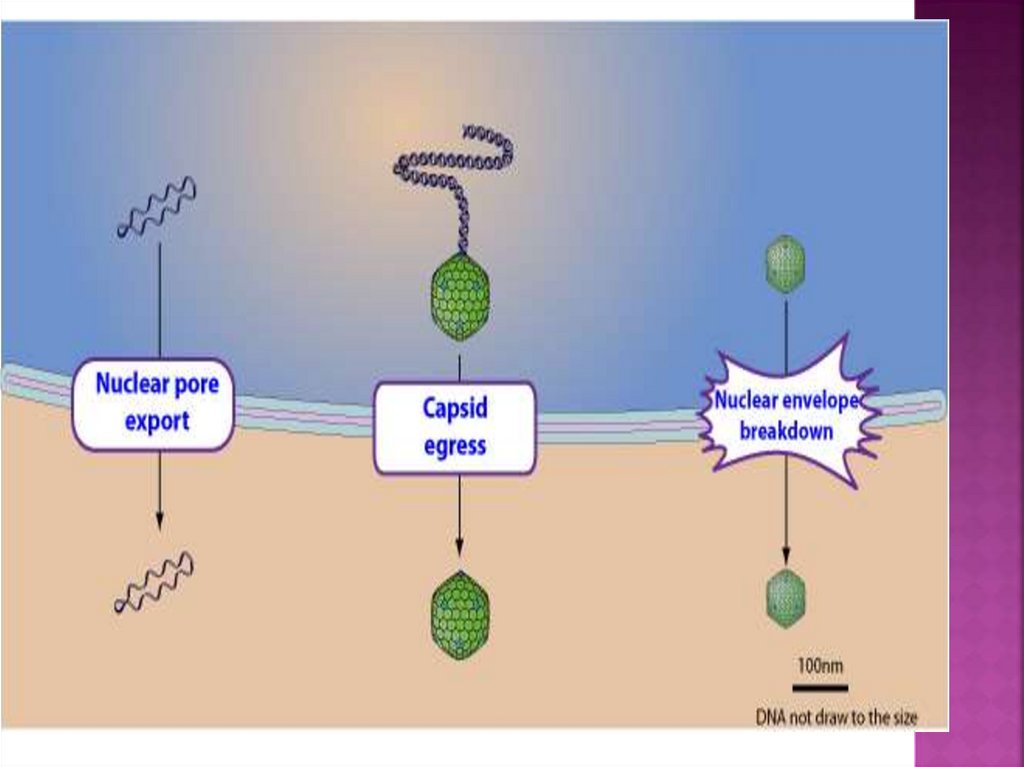
Вирусные капсиды, собранные в ядре, могутмигрировать в цитоплазму, используя
необычный путь экспорта, называемый
ядерным выходом. В этом процессе вирусный
капсид доставляется в перинуклеарное
пространство, образуя везикулярный
промежуточный продукт. После слияния с
внешней ядерной мембраной голый капсид
высвобождается в цитозоль.
Этот путь может быть эндогенным клеточным
путем, кооптированным вирусами, который
служит для транспортировки клеточных
белков, превышающих предельный размер
ядерных пор.
40.
41. почкование
42. Вирусный почкование через комплексы ESCRT хозяина
Многие оболочечные вирусы используют компонентыклеточного ESCRT (эндосомального сортировочного
комплекса, необходимого для транспорта) для
ассамблирования и почкования
43. ESCRT-независимое почкование
44. Вирус, покидающий клетку клеточным экзоцитозом
45. Вирус, почкующийся через плазматическую мембрану
46. Почкование вируса герпеса через околоядерные мембраны клетки хозяина
47. Почкование (экзоцитоз) вируса
48.
Взаимодействие вируса с клеткойхозяина – сложный и многостадийный
процесс.
В результате такого взаимодействия
могут развиваться три основные формы
клеточной инфекции:
1. Продуктивная
2. Абортивная
3. Интегративная
49. Продуктивная форма
Носит литический характер –заканчивается гибелью и лизисом
инфицированной клетки, что происходит
после полной сборки дочерней
популяции инфекционных вирусных
частиц.
A. Раннее подавление синтеза клеточных
белков
B. Накопление повреждающих клетку
вирусных компонентов
C. Повреждение лизисом и выбрасывание
их ферментов в цитоплазму
50. Абортивная форма
Не завершается образованиеминфекционных вирусных частиц или они
образуются в гораздо меньшем
количестве, чем при продуктивной
инфекции. Клетка либо погибает без
продукции инфекционного вируса, либо
инфекция прерывается на определенном
этапе.
A.
B.
Заражение чувствительных клеток
дефектным вирусом
Заражение чувствительных клеток в
неразрешающих условиях
51. интегративная форма
Не приводит к гибели клетки.Нуклеиновая кислота вируса, встроенная в
геном клетки-хозяина, функционирует как
составная часть клеточного генома.
Клетка может сохранять нормальные функции и
при ее делении вирусные последовательности
могут переходить в геном дочерних клеток.
Интеграция может привести к неопластической
трансформации клеток. Такие клетки
приобретают способность к неограниченному
делению
Ретровирусы. Аденовирусы. Герпесвирусы.
Папилломавирусы. Полиомавирусы.