
























 Биология
БиологияПохожие презентации:

")








Механизм действия сигнальных молекул. Гормоны (лекция)
1.
СЗГМУ им. И. И. МЕЧНИКОВАКАФЕДРА БИОЛОГИЧЕСКОЙ И ОБЩЕЙ ХИМИИ
БИОЛОГИЧЕСКАЯ ХИМИЯ-БИОХИМИЯ ПОЛОСТИ РТА
Раздел VIII
ЛЕКЦИЯ
МЕХАНИЗМ ДЕЙСТВИЯ СИГНАЛЬНЫХ МОЛЕКУЛ.
ГОРМОНЫ
Доцент Павлова Р.Н., Доцент Антонова Ж.В.
2022/2023
2.
План лекции1.Уровни регуляции обмена веществ и функций организма.
2.Метаболическая регуляция дихотомического распада глюкозы (схема)
3.Реципрокная регуляция обмена гликогена.
4. Гормональная регуляция. Сигнальные молекулы.
5. Внутриклеточный рецептор (прямой тип действия) и его характеристика.
6. Виды мембранных рецепторов
7. Мембранный (опосредованный путь передачи сигнала):
7.1. Аденилатциклазная система передачи сигнала через мембрану.
7.2. Фосфотидилинозитольный путь передачи сигнала через мембрану
7.3. Инсулиновый рецептор.
7. 4. Передача сигнала через мембранные рецепторы, ассоциированные c Янускиназами (JAK). Рецептор к СТГ
8. Гормональная регуляция уровня глюкозы в крови.
8.1. Снижение уровня глюкозы в крови (инсулин).
8.2. Повышение уровня глюкозы в крови (адреналин, глюкагон, глюкокортикоиды,
тироксин, АКТГ, ГР)
2/Ж.В.
3.
Список сокращений1. ПДК – пируватдегидрогеназный комплекс
2. α-КДК – α-кетоглутаратдегидрогеназный комплекс
3. ФИФ2 – фосфатидилинозитолбисфосфат
4. ИФ3 –инозитол-1,4,5-трифосфат
5. ДАГ –диацилглицерол
6. ФС –фосфатидилсерин
7. ЭР – эндоплазматический ретикулум
8. АЦ - аденилатциклаза
3/Ж.В.
4.
1.Уровни регуляции обмена веществ и функций организма.Системы регуляции обмена веществ и функций организма,
обеспечивающих гомеостаз, образуют 3 иерархических уровня
• Первый уровень – ЦНС. Нервные клетки получают сигналы из
внешней и внутренней среды, преобразуют их в форму нервного
импульса и передают через синапсы, используя медиаторы (химические
сигналы). Медиаторы вызывают изменения метаболизма в эффекторных
клетках.
• Второй уровень - эндокринная система. Она представлена железами
(иногда отдельными клетками), синтезирующими гормоны - химические
сигналы. Гормоны освобождаются в кровь в ответ на специфический
стимул. Гормоны транспортируются кровью, достигают клетокмишеней, модифицируют в них обмен веществ через внутриклеточные
механизмы.
4/Ж.В.
5.
• Третий уровень – внутриклеточный. Этометаболическая регуляция на клеточном уровне.
Этот уровень связан с изменениями метаболизма в
пределах клетки или отдельного метаболического
пути в результате изменения активности ферментов,
изменения количества ферментов, изменения скорости
транспорта веществ через мембраны клеток. Один из
основных механизмов регуляции метаболических
путей в клетке - это регуляция по типу отрицательной
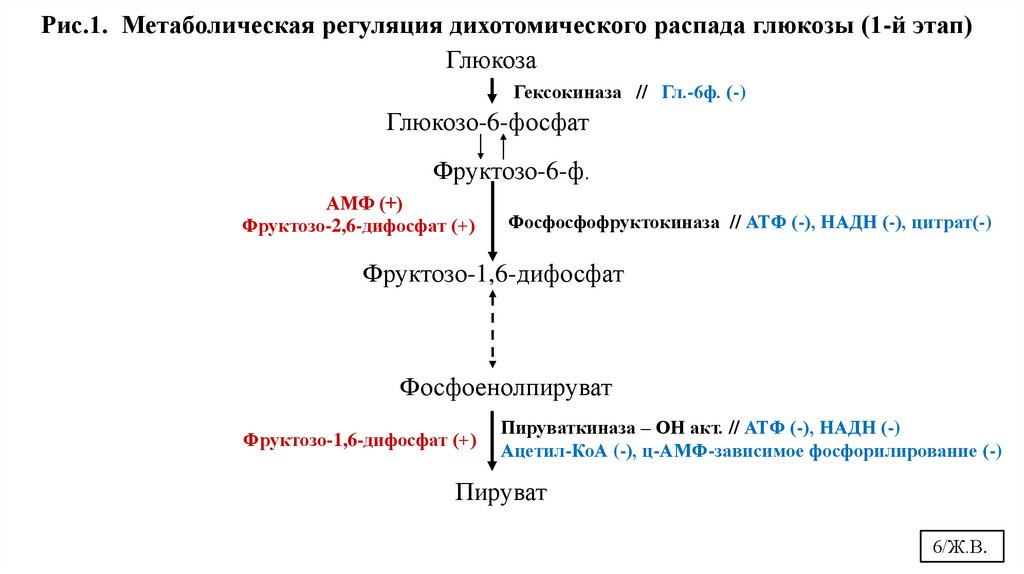
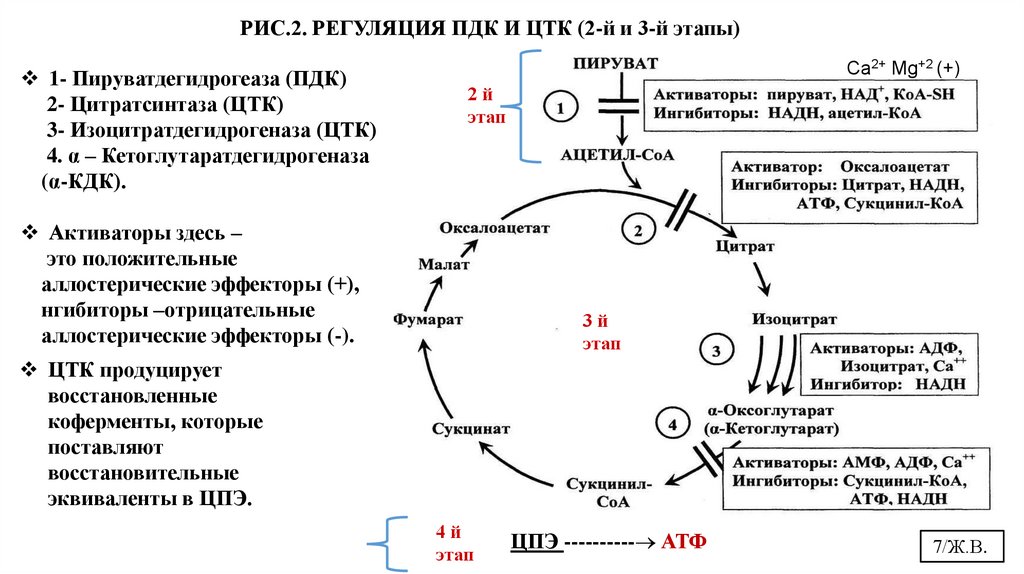
обратной связи (аллостерическая). (Рис.1, 2)
5/Ж.В.
6.
Рис.1. Метаболическая регуляция дихотомического распада глюкозы (1-й этап)Глюкоза
Гексокиназа // Гл.-6ф. (-)
Глюкозо-6-фосфат
Фруктозо-6-ф.
АМФ (+)
Фруктозо-2,6-дифосфат (+)
Фосфосфофруктокиназа // АТФ (-), НАДН (-), цитрат(-)
Фруктозо-1,6-дифосфат
Фосфоенолпируват
Фруктозо-1,6-дифосфат (+)
Пируваткиназа – ОН акт. // АТФ (-), НАДН (-)
Ацетил-КоА (-), ц-АМФ-зависимое фосфорилирование (-)
Пируват
6/Ж.В.
7.
РИС.2. РЕГУЛЯЦИЯ ПДК И ЦТК (2-й и 3-й этапы)1- Пируватдегидрогеаза (ПДК)
2- Цитратсинтаза (ЦТК)
3- Изоцитратдегидрогеназа (ЦТК)
4. α – Кетоглутаратдегидрогеназа
(α-КДК).
Сa2+ Mg+2 (+)
2й
этап
Активаторы здесь –
это положительные
аллостерические эффекторы (+),
нгибиторы –отрицательные
аллостерические эффекторы (-).
3й
этап
ЦТК продуцирует
восстановленные
коферменты, которые
поставляют
восстановительные
эквиваленты в ЦПЭ.
4й
этап
ЦПЭ ---------- АТФ
7/Ж.В.
8.
Высокий уровень АТФ (конечный продукт дихотомического пути распада глюкозы)ингибирует ключевые ферменты гликолиза фосфофруктокиназу и пируваткиназу.
Поэтому если - АТФ образуется в избыточном количестве, то происходит
торможение данного метаболического пути уже на ранних его стадиях. АТФ
является отрицательным аллостерическим эффектором для фермента
фосфофруктокиназы и пируваткиназы.
Однако, первая киназная реакция, т.е. образование глюкозо-6-фосфата, не тормозится избытком АТФ, т.к. это привело бы к снижению синтеза гликогена и работы
пентозофосфатного цикла (ПФЦ), которые начинаются также с образования
глюкозо-6-фосфата.
С увеличением количества АТФ замедляется ЦТК и накапливается избыток цитрата,
который выходит в цитоплазму и аллостерически ингибирует фосфофруктокиназу
(Рис.1). Это сигнал на снижение процесса окисления глюкозы и начала синтеза
липидов из молекул ацетил-КоА, доставленного в цитоплазму в составе цитрата.
Причем, кроме АТФ тормозящее влияние оказывают и другие метаболиты,
например, сукцинил-КоА, ацетил-КоА, цитрат, НАДН+Н+
8/Ж.В.
9.
Другим распространенным примером метаболической и гормональнойрегуляции является реципрокная регуляция распада и синтеза гликогена
посредством изменения концентрации в клетке вторичного посредника
цАМФ с последующим фосфорилированием или дефосфорилированием
регуляторных ферментов.
Реципрокной регуляцией называется такой тип клеточной регуляции, при
которой одно и то же по типу воздействие (наприм. фосфорилирование или
дефосфорилирование) на ключевые ферменты двух взаимосвязанных
процессов (наприм. синтез и распад гликогена) приводят к включению
одного процесса и выключению другого.
Известно, что скорость синтеза и распада гликогена находятся в
реципрокных взаимоотношениях.
Высокая концентрация цАМФ приводит к активации фосфорилазы (путем
ее фосфорилирования), и одновременно тормозит активность
гликогенсинтетазы (путем ее фосфорилирования).
9/Ж.В.
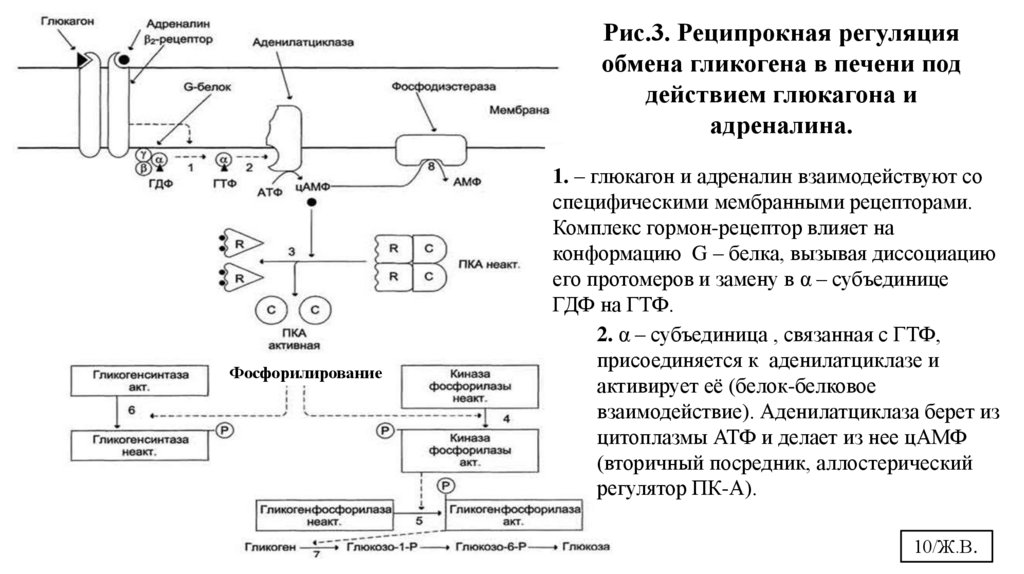
10.
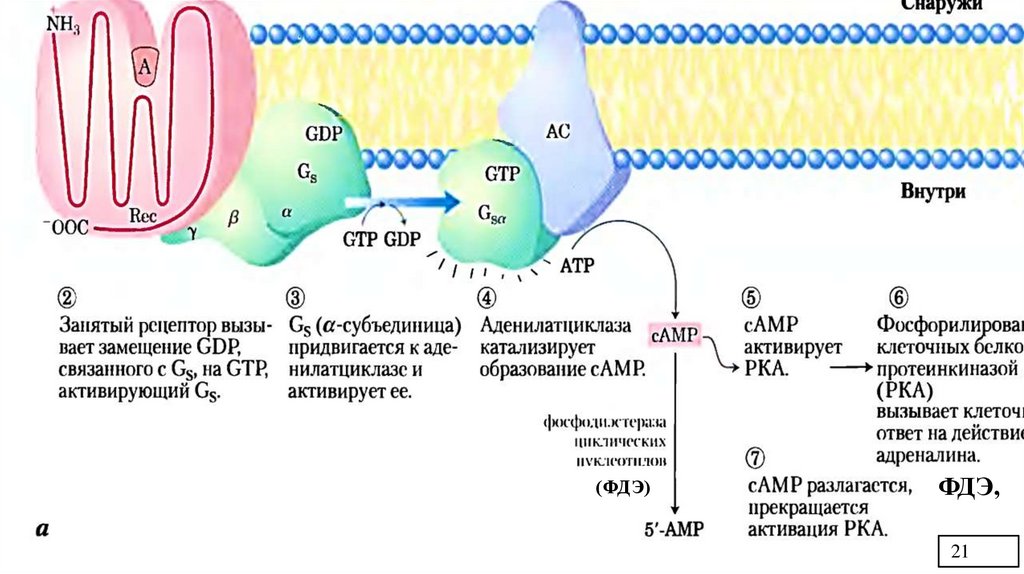
Рис.3. Реципрокная регуляцияобмена гликогена в печени под
действием глюкагона и
адреналина.
Фосфорилирование
1. – глюкагон и адреналин взаимодействуют со
специфическими мембранными рецепторами.
Комплекс гормон-рецептор влияет на
конформацию G – белка, вызывая диссоциацию
его протомеров и замену в α – субъединице
ГДФ на ГТФ.
2. α – субъединица , связанная с ГТФ,
присоединяется к аденилатциклазе и
активирует её (белок-белковое
взаимодействие). Аденилатциклаза берет из
цитоплазмы АТФ и делает из нее цАМФ
(вторичный посредник, аллостерический
регулятор ПК-А).
10/Ж.В.
11.
3. – цАМФ встраивается в аллостерические центры (по 2 цАМФ в каждый центр) регуляторныхсубъединиц ПК-А (цАМФ-зависимой), что приводит к изменению конформации регуляторных
субъединиц, связанных с каталитическими. ПК-А обратимо диссоциирует, освобождая
каталитические субъединицы (С) с каталитической активностью. ПК-А становится активной.
4. - ПК-А фосфорилирует и активирует киназу фосфорилазы.
5.- Киназа фосфорилазы активная фосфорилирует гликогенфосфорилазу, переводя её в активную
фосфорилированную форму.
6. – ПК-А фосфорилирует также гликогенсинтазу, переводя её в неактивное состояние.
7. – В результате фосфорилирования регуляторный фермент синтеза гликогена – гликогенсинтазастановится неактивной и синтез гликогена прекращается. А регуляторный фермент распада гликогена
– гликогенфосфорилаза- напротив, становится активным и начинается распад гликоген.
8. Фосфодиэстераза (ФДЭ) катализирует распад цАМФ и прерывает действие гормонального сигнала.
Комплекс α-субъединица-ГТФ затем распадается и α,β,γ-суюъединцы G-белка снова
реассоциируются.
ПРИМЕЧАНИЕ: данную схему можно использовать , отвечая механизм действия адреналина и
глюкагона на обмен гликогена в печени; объясняя аденилатциклазную систему, как сигнальный путь;
поясняя реципрокную регуляцию на примере обмена гликогена; приводя примеры быстрой регуляции
активности ферментов (белок-белковое взаимодействие, аллостерическая регуляция, ковалентная
модификация).
11/Ж.В.
12.
4. Гормональная регуляцияСигнальные молекулы– эндогенные соединения различной
химической природы, которые в результате взаимодействия с
рецепторами в клетках-мишенях обеспечивают регуляцию
биохимических реакций в метаболических процессах. К ним относятся:
• Гормоны
• Нейромедиаторы
• Биогенные амины
• Факторы роста
• Простагландины
• Цитокины
• Некоторые витамины (витамин Д, А)
Клетка-мишень - это клетка, имеющая рецепторы для данного
вида сигнальных молекул.
12.
13.
Эффект гормона зависит от:• Количества синтезированных гормонов
• Состояния рецептора, с которым он связывается
• Скорости распада и выведения (гормона)
• Наличия белка-транспортера (для гидрофобных гормонов)
• Модуляции гормональных систем
13
14.
По месту расположения рецептораВнутриклеточные рецепторы
(прямой тип действия)
Мембранные
(опосредованный тип действия)
14.
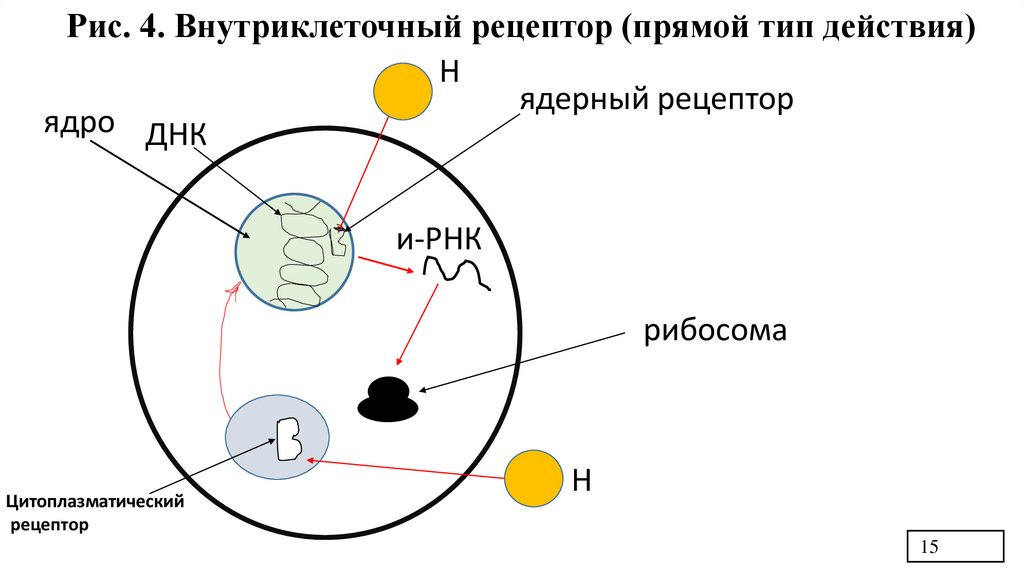
15.
Рис. 4. Внутриклеточный рецептор (прямой тип действия)Н
ядерный рецептор
ядро ДНК
и-РНК
рибосома
Цитоплазматический
рецептор
Н
15
16.
5. Характеристика внутриклеточных рецепторовПередача сигнала с помощью внутриклеточных рецепторов
1) Сигнальная молекула проникает внутрь клетки через билипидный слой мембраны,
где взаимодействует со своим рецептором, образуя гормон-рецепторный комплекс
2) Рецептор располагается в ядре или цитоплазме. Но! Цитоплазматический гормонрецепторный комплекс все равно проникает в ядро
3) В ядре гормон-рецепторный комплекс взаимодействует с определенным участком
ДНК. Димер рецептора узнает специфическую последовательность нуклеотидов в
промоторной области гена и гормон-рецепторный комплекс взаимодействует с ней в
промоторной части ДНК, что приводит к активации транскрипции.
3) Реализация ответа – процесс, затрагивающий транскрипцию и проявляющийся
индукцией или репрессией процесса синтеза белка.
4) Скрытый (латентный период) от 3 до 6 часов. Максимум ответа 8-12-24 часа.
Это медленная регуляция.
По такому типу действуют гидрофобные сигнальные молекулы
16
17.
ГИДРОФОБНЫЕ СИГНАЛЬНЫЕ МОЛЕКУЛЫСтероидные гормоны:
• Гормоны коры надпочечников
а. Глюкокортикоиды (кортизол, (гидро)кортизон, кортикостерон)
б. Минералокортикоиды (альдостерон, дезоксикортикостерон)
• Половые гормоны
а. Мужские (андрогены) андростерон, тестостерон
б. Женские (эстрогены) эстрадиол, эстриол, эстрон (фолликулин)
• Прогестерон (гормон желтого тела)
Тиреоидные гормоны
Витамины D и А
17
18.
6. Виды мембранных рецепторов1) Рецептор, для реализации ответа требующие образования вторичных
посредников –
• 7-ТМС (трансмембранный сегмент) рецептор
а) аденилатциклазный путь проведения сигнала
б) фосфатидилинозитольный путь проведения сигнала, который часто
объединяют с в) кальциевым путем проведения сигнала.
• 1-ТМС- рецептор - гуанилатциклазный путь проведения сигнала
2) Рецепторы, представленные тирозиновой протеинкиназой – передача
сигнала путем формирования каскада фосфорилирования - это
рецепторы для факторов роста, инсулина, пролактина.
3) Рецепторы, связанные с ионным каналом характерны для
нейромедиаторов . Это рецепторы для глицина, глутамата, ГАМК.
18..
19.
7. Мембранный (опосредованный путь передачи сигнала)1) Гормон в клетку не проникает, а связывается с мембранным
рецептором
2) Рецептор расположен в плазматической (наружной мембране)
3) Для реализации действия необходимо:
а) образование вторичных посредников:
- цАМФ, цГМФ, Са2+, инозитол-1,4,5 – трифосфат (ИФ3, IP3)
- диацилглицерол (ДАГ, DAG), NO
- б) каскад реакций фосфорилирования
- 4) Латентный период 15-30` максимум ответа 30-60`
19.
20.
По такому механизму действуют ГИДРОФИЛЬНЫЕсигнальные молекулы
Гормоны белковой и пептидной природы
• Все гормоны гипофиза
• Гормоны поджелудочной железы (ИНСУЛИН и ГЛЮКАГОН)
• Гормоны паращитовидной железы
• Гормон щитовидной железы (тиреокальцитонин)
Гормоны мозгового вещества надпочечников (катехоламины:
адреналин и норадреналин)
Биогенные амины
Нейромедиаторы
Факторы роста
Простагландины
Цитокины
20.
21.
(ФДЭ)ФДЭ,
21
22.
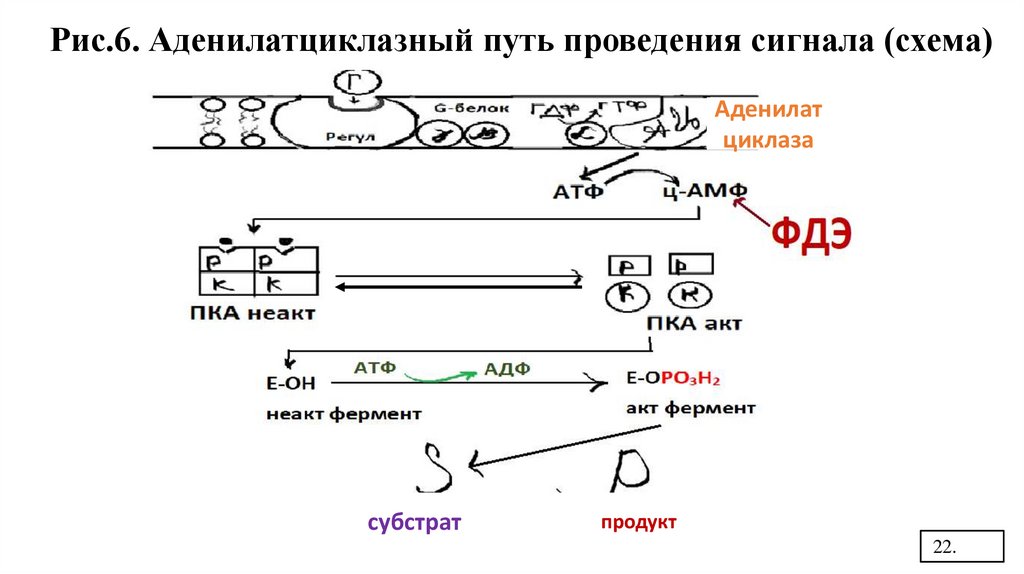
Рис.6. Аденилатциклазный путь проведения сигнала (схема)Аденилат
циклаза
субстрат
продукт
22.
23.
Примеры ферментов, активирующихся поаденилатциклазному пути.
• В жировой ткани - липаза
Липаза (Е) : ТАГ (S)
глицерол + ЖК (P)
• В печени – гликогенфосфорилаза
Гликогенфосфорилаза (E):гликоген (S)
гликоген n-1+Гл-1-Ф (P)
23.
24.
Механизмы прекращения сигнала.• За счет гидролиза ГТФ на α-субъединице
G-белка, которая теряет сродство к
АЦ (аденилатциклазе) и возвращается к
β-субъединице
• Расщепление ц-АМФ за счет ФДЭ (фосфодиэстеразы)
24.
25.
Влияние адреналина в зависимости от типа ткани и рецептора• β –адренорецепторы: повышает концентрацию ц-АМФ
• α2 –адренорецепторы: снижает концентрацию ц-АМФ (инг. АЦ)
• α1 – адренорецепторы: вызывает образование других
_____________________вторичных посредников: ДАГ, ИФ3
2/Ж.В.
26.
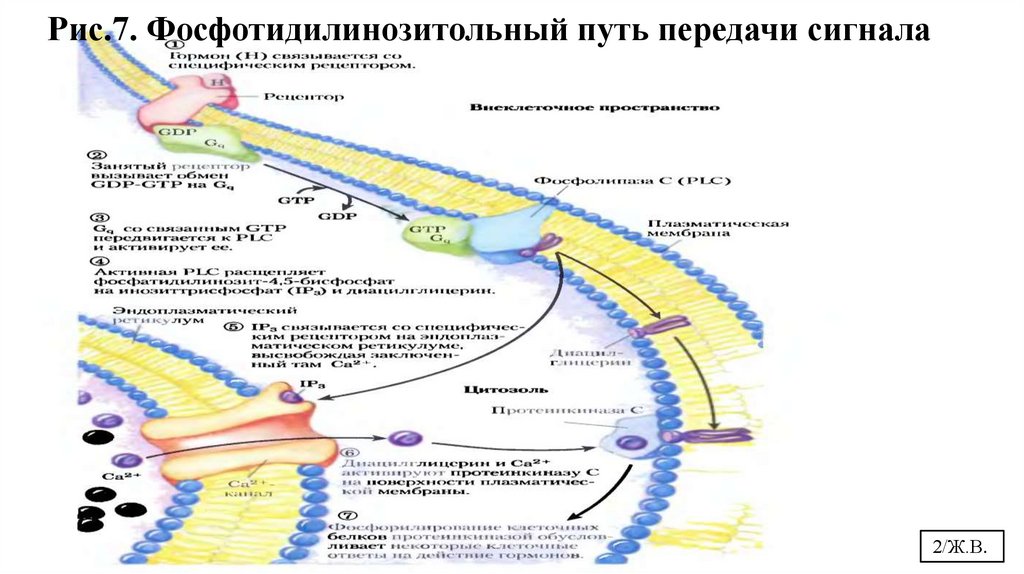
Рис.7. Фосфотидилинозитольный путь передачи сигнала2/Ж.В.
27.
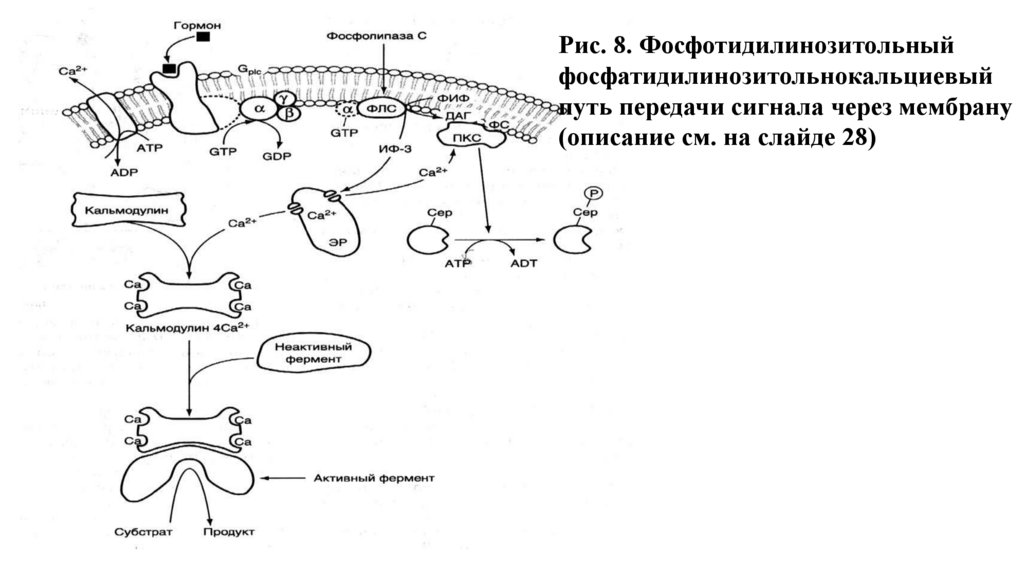
Рис. 8. Фосфотидилинозитольныйфосфатидилинозитольнокальциевый
путь передачи сигнала через мембрану
(описание см. на слайде 28)
28.
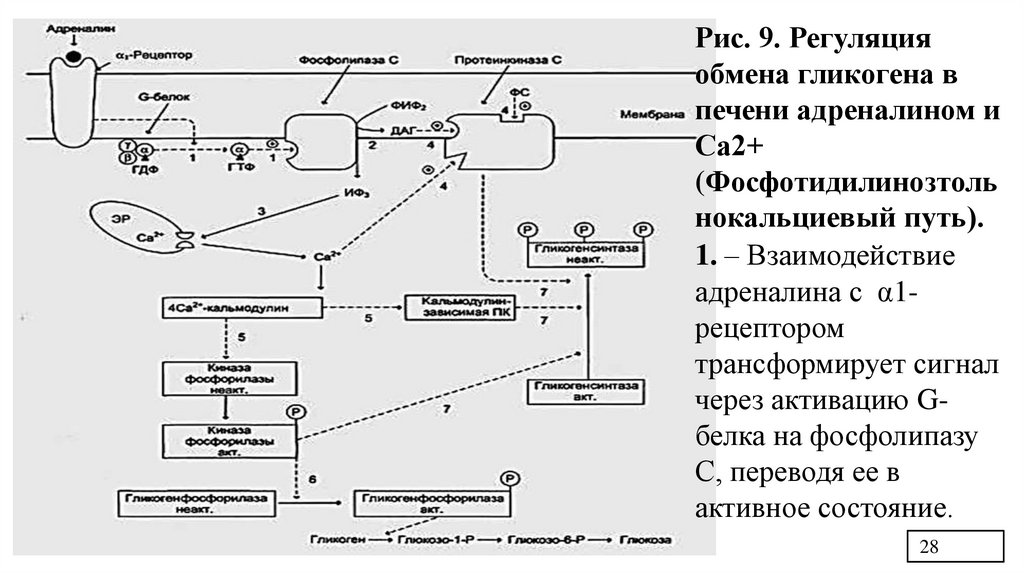
Рис. 9. Регуляцияобмена гликогена в
печени адреналином и
Са2+
(Фосфотидилинозтоль
нокальциевый путь).
1. – Взаимодействие
адреналина с α1рецептором
трансформирует сигнал
через активацию Gбелка на фосфолипазу
С, переводя ее в
активное состояние.
28
29.
2.- Фосфолипаза С гидролизует ФИФ2 (мембранный фосфолипид) на двавторичных посредника: ИФ3 ( мигрирует в цитоплазму) и ДАГ (остается в
мембране).
3.- ИФ3 связывается со специфическими рецепторами на ЭР и активирует
мобилизацию Са2+ из ЭР.
4. - Са2+, ДАГ и фосфатидилсерин активируют протеинкиназу С. Она
фосфорилирует гликогенсинтазу, переводя ее в неактивное состояние.
5. – Комплекс 4Са2+ - кальмодулин активирует киназу фосфорилазы и
кальмодулинзависимые протеинкиназы.
6. – Киназа фосфорилазы фосфорилирует гликогенфосфорилазу и тем
самым ее активирует.
7. – Активные формы 3-х ферментов (кальмодулинзависимая
протеинкиназа, киназа фосфорилазы и протеинкиназа С) фосфорилируют
гликогенсинтазу в различных центрах, переводя ее в неактивное
состояние.
29
30.
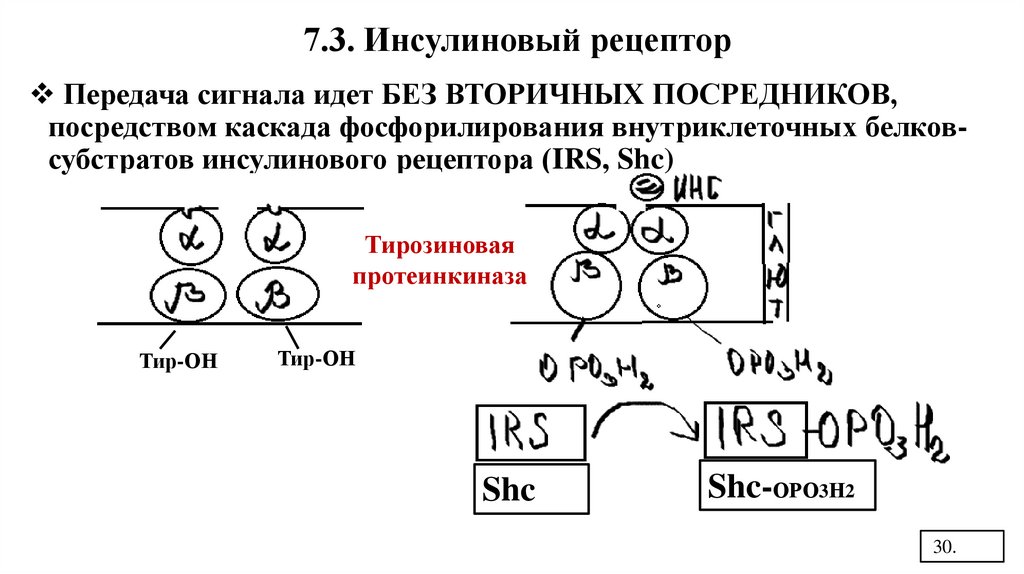
7.3. Инсулиновый рецепторПередача сигнала идет БЕЗ ВТОРИЧНЫХ ПОСРЕДНИКОВ,
посредством каскада фосфорилирования внутриклеточных белковсубстратов инсулинового рецептора (IRS, Shc)
Тирозиновая
протеинкиназа
Тир-ОН
Тир-ОН
Shc
Shc-ОРО3Н2
30.
31.
При действии инсулина рецептор димеризуется икаждая β-субъединица тирозиновой киназы делает
аутофосфорилирование по тирозину.
Активный фосфорилированный рецептор
фосфорилирует белки - субстраты инсулинового
рецептора.
Фосфорилированные субстраты инсулинового
рецептора IRS-OPO3H2 и Shc- ОРО3Н2 вызывают
активацию разных путей передачи сигнала.
31.
32.
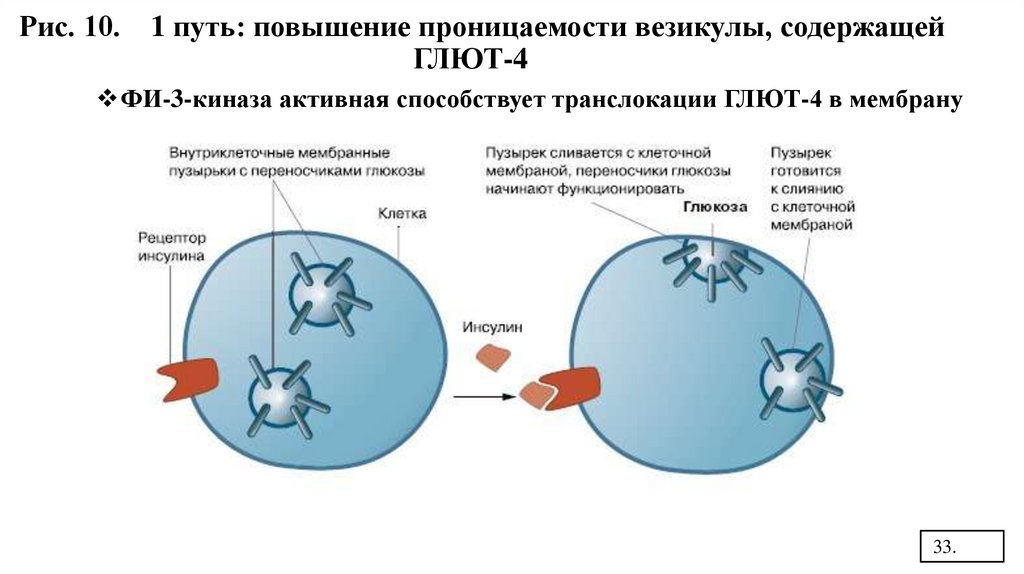
1 путь: повышение проницаемости везикулы, содержащей ГЛЮТ-4, чтоприводит к поступлению глюкозы в клетку. (Рис.10)
2 путь: активация цитоплазматических ферментов: гексокиназы, ФДЭ (Рис. 11),
протеинфосфатазы.
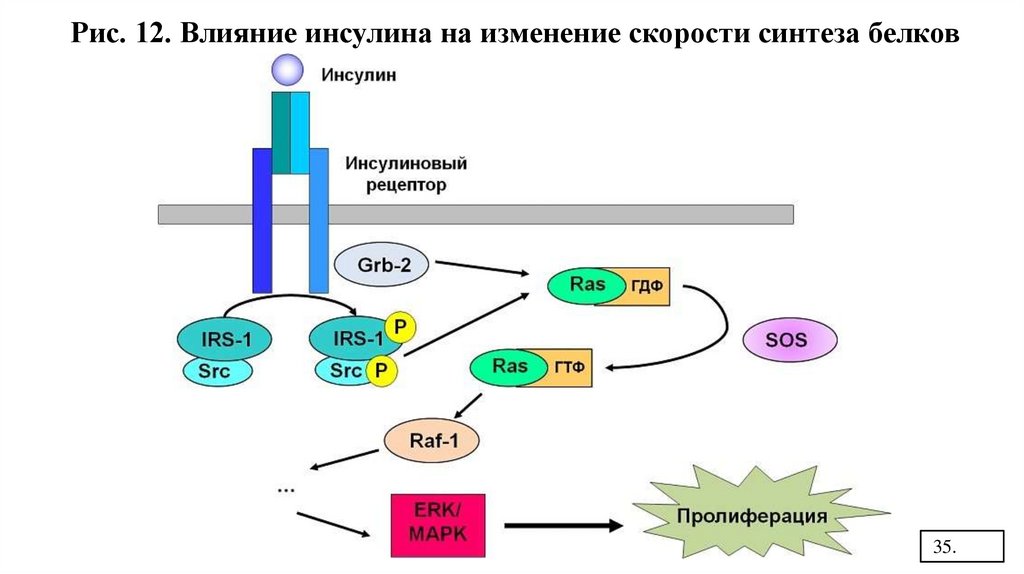
3 путь: фосфорилирование внутриклеточных белков: RAS
MAPK
сигнал в ядро
Raf
изменение скорости синтеза белка (Рис.12)
( МАПК - митогенактивируемые протеинкиназы).
32
33.
Рис. 10. 1 путь: повышение проницаемости везикулы, содержащейГЛЮТ-4
ФИ-3-киназа активная способствует транслокации ГЛЮТ-4 в мембрану
33.
34.
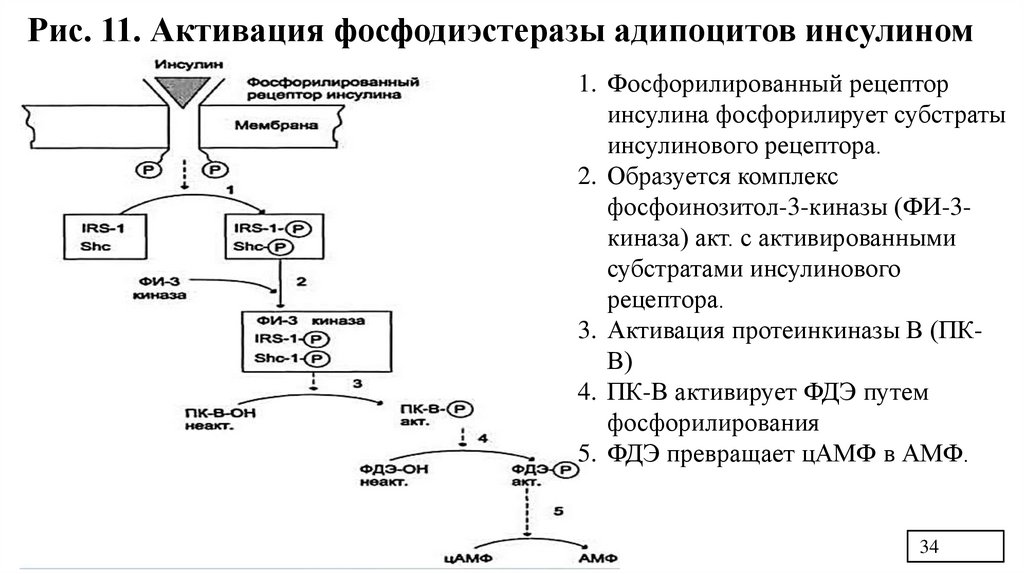
Рис. 11. Активация фосфодиэстеразы адипоцитов инсулином1. Фосфорилированный рецептор
инсулина фосфорилирует субстраты
инсулинового рецептора.
2. Образуется комплекс
фосфоинозитол-3-киназы (ФИ-3киназа) акт. с активированными
субстратами инсулинового
рецептора.
3. Активация протеинкиназы В (ПКВ)
4. ПК-В активирует ФДЭ путем
фосфорилирования
5. ФДЭ превращает цАМФ в АМФ.
34
35.
Рис. 12. Влияние инсулина на изменение скорости синтеза белков35.
36.
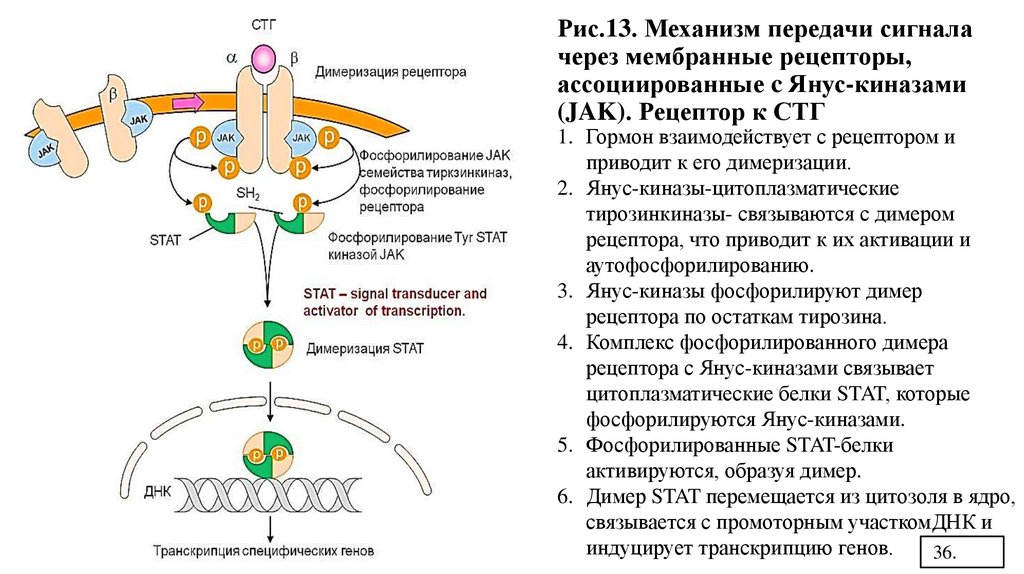
Рис.13. Механизм передачи сигналачерез мембранные рецепторы,
ассоциированные c Янус-киназами
(JAK). Рецептор к СТГ
1. Гормон взаимодействует с рецептором и
приводит к его димеризации.
2. Янус-киназы-цитоплазматические
тирозинкиназы- связываются с димером
рецептора, что приводит к их активации и
аутофосфорилированию.
3. Янус-киназы фосфорилируют димер
рецептора по остаткам тирозина.
4. Комплекс фосфорилированного димера
рецептора с Янус-киназами связывает
цитоплазматические белки STAT, которые
фосфорилируются Янус-киназами.
5. Фосфорилированные STAT-белки
активируются, образуя димер.
6. Димер STAT перемещается из цитозоля в ядро,
связывается с промоторным участкомДНК и
индуцирует транскрипцию генов.
36.
37.
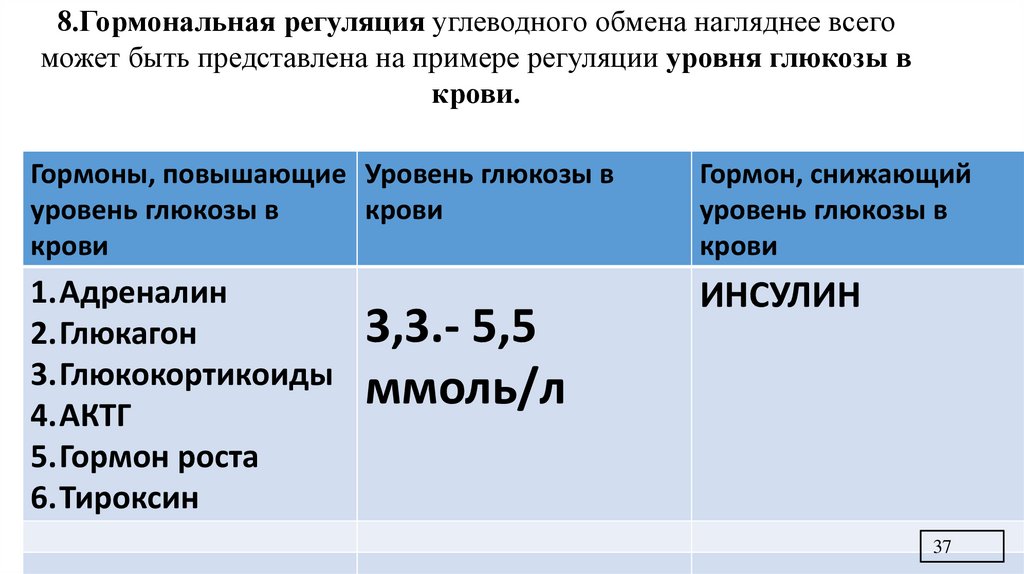
8.Гормональная регуляция углеводного обмена нагляднее всегоможет быть представлена на примере регуляции уровня глюкозы в
крови.
Гормоны, повышающие Уровень глюкозы в
уровень глюкозы в
крови
крови
Гормон, снижающий
уровень глюкозы в
крови
1.Адреналин
2.Глюкагон
3.Глюкокортикоиды
4.АКТГ
5.Гормон роста
6.Тироксин
ИНСУЛИН
3,3.- 5,5
ммоль/л
37
38.
8.1. Снижение уровня глюкозы в крови. ИНСУЛИН.1.Повышает проницаемость клеточных мембран для глюкозы за счет
перемещениия в мембрану из цитоплазмы переносчиков глюкозы.
2. Повышает окисление глюкозы в клетке за счет активации гексокиназы,
индуцирует синтез глюкокиназы, посредством активации генома путем каскада
реакции фосфорилирования.
3. Усиливает синтез гликогена. Эффект активации осуществляется путем
дефосфорилирования гликогенсинтетазы.
4. Усиливает процесс липогенеза (синтеза жира) - под действием инсулина 30%
глюкозы переходит в жир. Усиление липогенеза происходит за счет активации
фермента синтеза жирных кислот ацетил-КоА - карбоксилазы путем
дефосфорилирования и также путем индукции синтеза синтетазы жирных кислот.
5. Инсулин тормозит процесс глюконеогенеза за счет репрессии синтеза
ключевых ферментов глюконеогенеза.
38
39.
8.2 ПОВЫШЕНИЕ УРОВНЯ ГЛЮКОЗЫ В КРОВИ1. АДРЕНАЛИН-способствует мобилизации гликогена печени и мышц Механизм действия
реализуется через аденилатциклазную систему.
2. ГЛЮКАГОН-а)способствует мобилизации гликогена печени. Механизм действия
реализуется через аденилатциклазную систему; б) активирует глюконеогенез - при
повышении концентрации цАМФ усиливаются процессы фосфорилирования, что приводит
к инактивации пируватоксидазного комплекса, накоплению пирувата, который используется
в глюконеогенезе.
3. ГЛЮКОКОРТИКОИДЫ-кортикостерон, гидрокортизон, кортизон - усиливают процесс
глюконеогенеза. Гормоны проникают внутрь клетки, что способствует индукции ключевых
ферментов глюконеогенеза (фосфоенолпируваткарбоксикиназы, глюкозо-6-фосфатазы и
фруктозо-1,6-дифосфатазы, пируваткарбоксилазы); ферментов распада аминокислот.
4. ТИРОКСИН-повышает уровень глюкозы в крови, усиливает всасывание глюкозы в
кишечнике, потенциирует действие адреналина, повышает активность окислительных
ферментов.
5. АКТГ-повышает уровень глюкозы в крови ( в основном за счет увеличения синтеза
гормонов коры надпочечников).
39.
6. ГОРМОН РОСТА – усиливает глюконеогенез и всасывание глюкозы в кишечнике
40.
Рекомендуемая литература1. Биохимия : учебник / под ред. Е.С. Северина. – М. : ГЭОТАР-Медиа, 2011. – С. 355–358; 534–580.
2. Биохимия [Электронный ресурс] : учебник / под ред. Е.С. Северина. – 5-е изд., испр. и доп. – М. : ГЭОТАР-Медиа,
2015. – http://www.studmedlib.ru / book / ISBN9785970433126.html (разд. 7, п. Х–ХI; разд. 11, п. I–V; с. 355–358; 534–
580).
3. Биологическая химия. Биохимия полости рта : учебник / Т.П. Вавилова, А.Е. Медведев. – М. : ГЭОТАР-Медиа,
2014. – С. 277–278.
4. Биологическая химия. Биохимия полости рта [Электронный ресурс] : учебник / Т.П. Вавилова, А.Е. Медведев. –
М. : ГЭОТАР-Медиа, 2014. – http://www.studmedlib.ru / book / ISBN9785970430392.html (ч. V, гл. 13; с. 277–278).
5. Метаболические нарушения и зубочелюстная система : учебно-методическое пособие для студентов
стоматологических факультетов медицинских вузов / под ред. Л.Б. Гайковой, Ж.В. Антоновой, Р.Н. Павловой. – СПб. :
Изд-во СЗГМУ им. И.И. Мечникова, 2017. – С. 95–111.
6. Обмен углеводов. Энергетический обмен. Гормональная регуляция обмена веществ и функций организма : учебнометодическое пособие к практическим и лабораторным занятиям по биологической химии – биохимии полости рта для
студентов стоматологических факультетов медицинских вузов / под ред. Ж.В. Антоновой, Р.Н. Павловой, Л.Б.
Гайковой. – СПб. : Изд-во СЗГМУ им. И.И. Мечникова, 2019. – С. 56–73.
7. Сборник тестовых заданий по разделам биологической химии и биохимии полости рта: учебно-методическое
пособие для студентов стоматологического факультета / под ред. В.А. Дадали, Ж.В. Антоновой, Р.Н. Павловой. – СПб. :
Изд-во СЗГМУ им. И. И. Мечникова, 2017. – С. 70–76.
8. Биологическая химия / Т.Т. Березов, Б.Ф. Коровкин. – М. : Медицина, 1998. – С. 248–283, 289–298, 357–361.
40.
41.
Благодарю за внимание!Пройдите тестирование.
41/Ж.В.