














 Биология
БиологияПохожие презентации:










Роль, распределение и миграция кальция в клетке
1.
Кафедра биохимииРОЛЬ, РАСПРЕДЕЛЕНИЕ
И МИГРАЦИЯ КАЛЬЦИЯ
В КЛЕТКЕ
2. Роль кальция в организме
Основнойстроительный материал
для твёрдых
минерализованных
тканей
(входит в состав
гидроксиапатита
Ca10(PO4)6(OH)2)
Участник мышечного
сокращения
(связывание с
тропонином,
активация каталитических
центров для расщепления
АТФ на головках миозина)
Участник процесса
свертывания крови
(IV фактор свёртывания,
активирует
протромбиназу,
превращение
протромбина в тромбин,
ингибирует фибринолиз)
КАЛЬЦИЙ
Участник формирования
возбуждения в клетке
(формирует фазу роста
потенциала действия в
нейронах, определяет
возбудимость клеточной
мембраны)
Посредник
в передаче
гормонального сигнала
(выполняет роль
вторичного и третичного
мессенджера)
Аллостерический
модулятор активности
более 300 ферментов
(активирует АТФазу,
киназу фосфорилазы,
фосфодиэстеразу,
ингибирует енолазу,
дипептидазы)
3. Распределение кальция
I пул– экстрацеллюлярный кальций
(кальций внеклеточного пространства).
II пул
– интрацеллюлярный кальций
(внутриклеточный кальций).
III пул
– кальций внутриклеточного пространства
(кальций митохондрий и ЭПР)
4. Кальцийсвязывающие белки
Кальцийсвязывающиебелки
Депонирующие
белки
(кальсеквестрин,
кальрегулин,
кальретикулин)
Каналообразующие
белки
(Са2+-АТФаза)
Функциональные
белки
(тропонин С,
кальмодулин)
5. Миграция кальция
1. Медленныепотенциалозависимые
кальциевые каналы
сарколеммы L-типа
2
4
Na+-K+-АТФ-аза
Na+
3. Ca2+-АТФаза сарколеммы
Ca2+
1
3
5
2. Кальциевые каналы
саркоплазматического
ретикулума, связанные с
рианодиновым рецептором
K+
Na+
4. Ca2+-АТФаза
саркоплазматического
ретикулума
5. Na+/Ca2+-обменник
6. Потенциалозависимые кальциевые каналы
Состав:o Входные ворота
o Пора
o
o
o
o
Виды:
L-типа
N-типа
P-типа
T-типа
Потенциалозависимый кальциевый канал
L- типа носит название
дигидропиридинового рецептора,
т.к. чувствителен к действию
дигидропиридинов.
Цитоплазматическая петля, формирующая
ножку, которая является сенсором
напряжения, взаимодействующим с
рианодиновым рецептором
7. Рианодиновый рецептор
Рианодиновые рецепторы (RyR) особый тип хемоактивируемых Са2+-каналов,имеющихся в мембране саркоплазматического ретикулума.
Выполняют важнейшую функцию
сопряжения потенциала действия с мышечным сокращением.
RyR активируется:
(в микромолярных концентрациях),
АТФ,
кофеином,
жирными кислотами,
рианодином (в низкой концентрации).
Са2+
Ryania speciosa
Рианодин –
алкалоид
растительного
происхождения
RyR инактивируется:
Са2+ (в миллимолярных концентрациях),
рианодином (в высокой концентрации).
Изоформы рецепторов, кодируемые разными генами.
RyR1 – в скелетной мускулатуре,
RyR2 – в миокарде,
RyR3 – в головном мозге и диафрагме.
В скелетных мышцах рианодиновые рецепторы активируются
посредством специализированного механизма прямого
электромеханического сопряжения,
а сокращение сердечной мышцы запускается по механизму
Са2+-индуцированного выброса Са2+ .
8. Рианодиновый рецептор
Трехмерная модель рианодинового рецептора,построенная по данным электронной микроскопии
Цитоплазматический
домен
Вид сбоку
Гидрофобная часть,
погруженная в
мембрану СПР
Центральный
ионопроводящий
канал
Вид со
стороны
цитоплазмы
Радиальный
ионопроводящий
канал
Базальная
платформа
Вид со
стороны
мембраны
9. Рианодиновый рецептор
АБ
Ca2+
В
Механизм высвобождения
из цистерн саркоплазматического ретикулума.
А - рецептор рианодина в мембране цистерны связан с Са2+-каналом. Работу канала контролируют
рецепторы дигидропиридина плазмолеммы, в ответ на деполяризацию активирующие рецепторы
рианодина, что и приводит к высвобождению Ca2+ (скелетные мышцы);
Б - рецептор рианодина в мембране цистерны непосредственно не связан с белками
плазмолеммы, но активируется Ca2+ при незначительном повышении концентрации этого катиона в
цитозоле за счёт поступления в клетку через потенциалозависимые Ca2+-каналы (миокард);
В - Ca2+ освобождается из цистерн при активации рецепторов ИТФ. Этот второй посредник
образуется при участии фосфолипазы C в ответ на взаимодействие агониста со своим рецептором в
плазмолемме (гладкие мышцы)
10. Рианодиновый рецептор
АБ
В
Механизм высвобождения Ca2+ из
цистерн саркоплазматического
ретикулума.
А - рецептор рианодина в мембране
цистерны связан с Са2+-каналом. Работу
канала
контролируют
рецепторы
дигидропиридина плазмолеммы, в ответ на
деполяризацию активирующие рецепторы
рианодина,
что
и
приводит
к
высвобождению Ca2+ (скелетные мышцы);
Б - рецептор рианодина в мембране
цистерны непосредственно не связан с
белками плазмолеммы, но активируется
Ca2+ при незначительном повышении
концентрации этого катиона в цитозоле за
счёт
поступления
в
клетку
через
2+
потенциалозависимые
Ca -каналы
(миокард);
В - Ca2+ освобождается из цистерн при
активации рецепторов ИТФ. Этот второй
посредник
образуется
при
участии
фосфолипазы C в ответ на взаимодействие
агониста
со
своим
рецептором
в
плазмолемме (гладкие мышцы)
11. Ca2+-АТФаза
Механизм работы Са2+-АТФазы.(1) - Когда протеин находится в исходной конформации, два иона Ca2+ по очереди
связываются с высокоаффинными участками поверхности протеина, обращенной в сторону
цитозоля. Затем АТФ связывается с другим определенным участком поверхности протеина,
обращенной в сторону цитозоля.
(2) - Одновременное присутствие ионов Ca2+ и Mg2+-АТФ запускает гидролиз АТФ.
(3) - Освобожденный фосфат (концевая фосфатная группа в АТФ) переносится от АТФ к
карбоксильной группе остатка аспарагиновой кислоты протеина (фосфорилирование белка по
карбоксильной
группе
аспарагиновой
кислоты),
формируя
высокоэнергетическую
ацилфосфатную связь.
(4) - Происходит образование фосфорилированного протеина (или ферментфосфатного
комплекса). Это изменение конформации протеина ведет к одновременному проталкиванию
двух ионов Ca2+ на другую сторону протеина и соответственно мембраны.
12. Na+/Ca2+-обменник
Натрий-кальциевый обменник трансмембранный белок цитоплазматической мембраны, транспортирующий ионы кальция изклетки в обмен на ионы натрия, которые поступают в клетку (механизм антипорта).
Обменник использует энергию, накопленную в электрохимическом градиенте натрия,
пропуская три иона Na+ в клетку по градиенту концентрации
и выводя один ион Ca2+ из клетки против градиента концентрации (вторично-активный транспорт).
Эта система работает обратимо: при искусственном понижении внеклеточной концентрации Na+
либо при повышении концентрации этого иона в цитоплазме может происходить вход Са2+ снаружи
в обмен на внутриклеточный натрий. Поскольку этот процесс является электрогенным, он в
большой степени зависит от мембранного потенциала: деполяризация мембраны наряду с
уменьшением трансмембранного градиента ионов натрия также способствует входу Са2+ в клетку.
13.
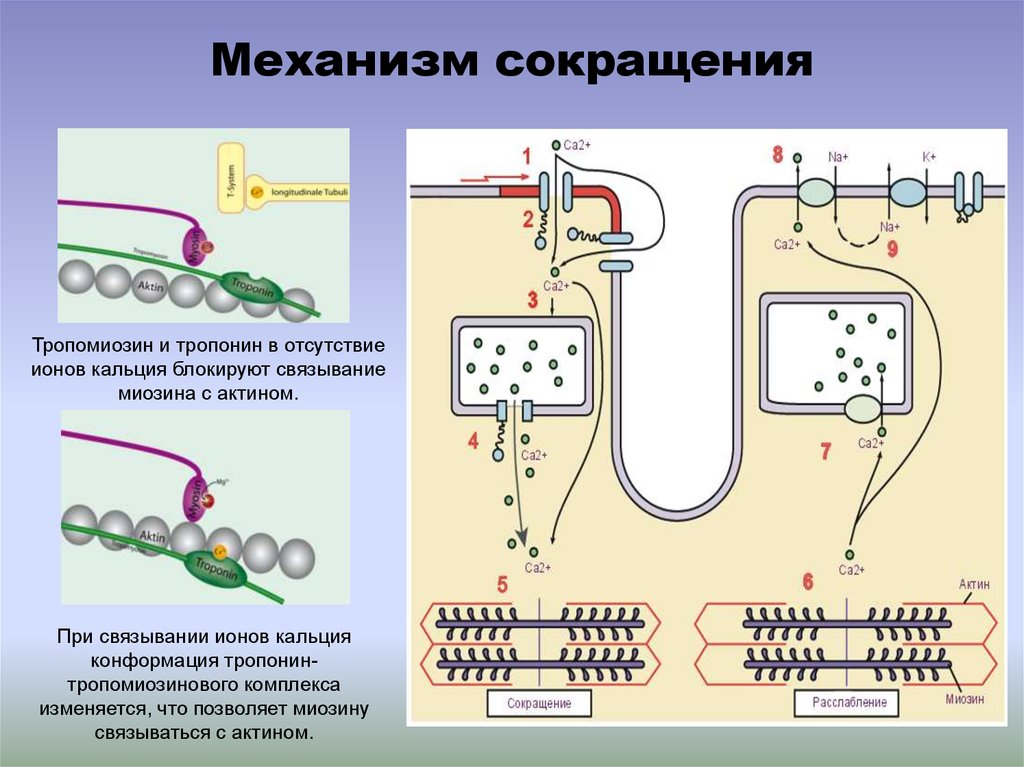
Механизм сокращенияТропомиозин и тропонин в отсутствие
ионов кальция блокируют связывание
миозина с актином.
При связывании ионов кальция
конформация тропонинтропомиозинового комплекса
изменяется, что позволяет миозину
связываться с актином.
14.
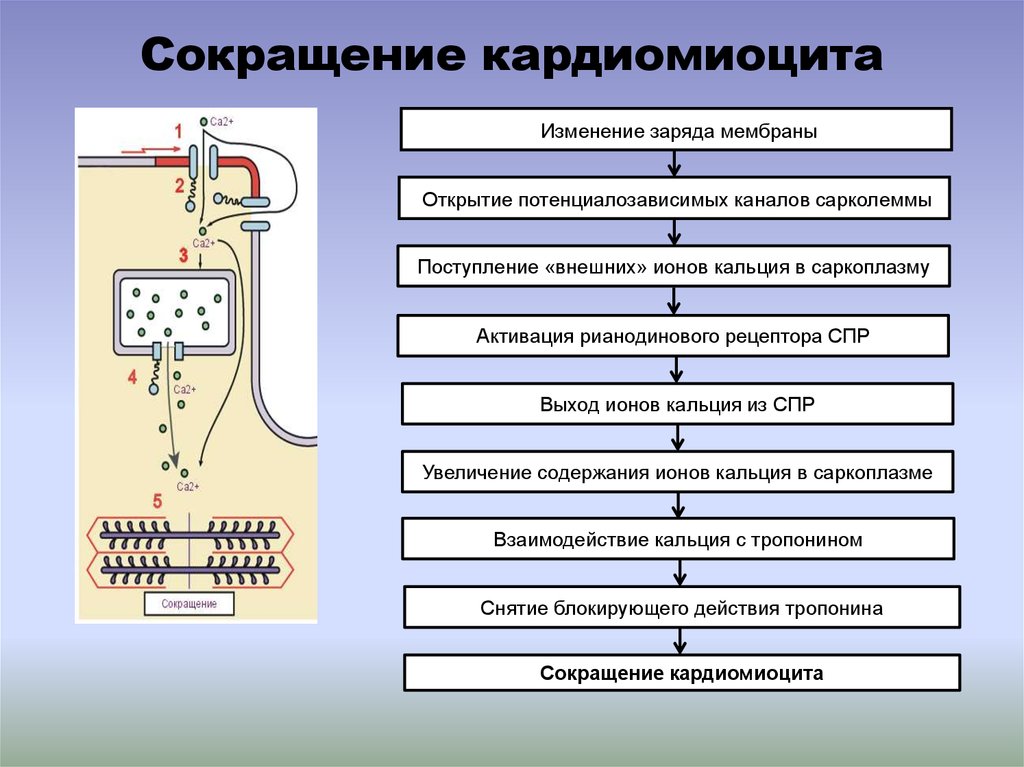
Сокращение кардиомиоцитаИзменение заряда мембраны
Открытие потенциалозависимых каналов сарколеммы
Поступление «внешних» ионов кальция в саркоплазму
Активация рианодинового рецептора СПР
Выход ионов кальция из СПР
Увеличение содержания ионов кальция в саркоплазме
Взаимодействие кальция с тропонином
Снятие блокирующего действия тропонина
Сокращение кардиомиоцита
15. Расслабление кардиомиоцита
Увеличение содержания кальцияв цитозоле
Торможение
процесса
освобождения
ионов кальция
Активация
обратного
захвата
кальция в СПР
Сокращение
Активация обмена ионами на
сарколемме:
2+
ионы Са выходят из клетки, а
ионы Na+ входят в
саркоплазму. Поддержание
концентрационного градиента
осуществляется за счёт Na+,К+АТФазы.
Уменьшение концентрации кальция в цитозоле
Диссоциация комплекса «кальций-тропомиозин»
Тропомиозин подавляет сцепление актина и миозина
Расслабление кардиомиоцита
16. Механизм сокращения
В общем объёме кальция, обеспечивающего сокращение,на долю межклеточного кальция приходится 20%,
на долю саркоплазматического кальция – 80%.
80% кальция возвращаются в полость саркоплазматического ретикулума,
15% выходят из клетки за счёт работы Na+/Ca2+-обменника,
5% - за счёт работы Са2+-АТФазы.
17.
СПАСИБОЗА ВНИМАНИЕ!