








































































 Биология
БиологияПохожие презентации:
")



")



")

Метаболизм ДНК
1. Метаболизм ДНК
2. Метаболизм ДНК
• Репликация – процесс точного копированиямолекулы ДНК.
• Репарация – поддержание целостности
биологического материала в ДНК.
• Рекомбинация –перестройка заключенной в
молекуле ДНК генетической информации.
3. Репликация
• Процесс самовоспроизведениямакромолекул нуклеиновых кислот,
обеспечивающий точное копирование
генетического материала и передачу его от
поколения к поколению.
• В основе репликации лежит понятие
матрицы – это макромолекулярная
структура для синтеза комплементарной
копии макромолекулы.
4.
• Репликация ДНК происходит вопределенной фазе клеточного цикла.
• Основные свойства процесса репликации
ДНК и каталитические механизмы этого
процесса практически идентичны у всех
видов организмов.
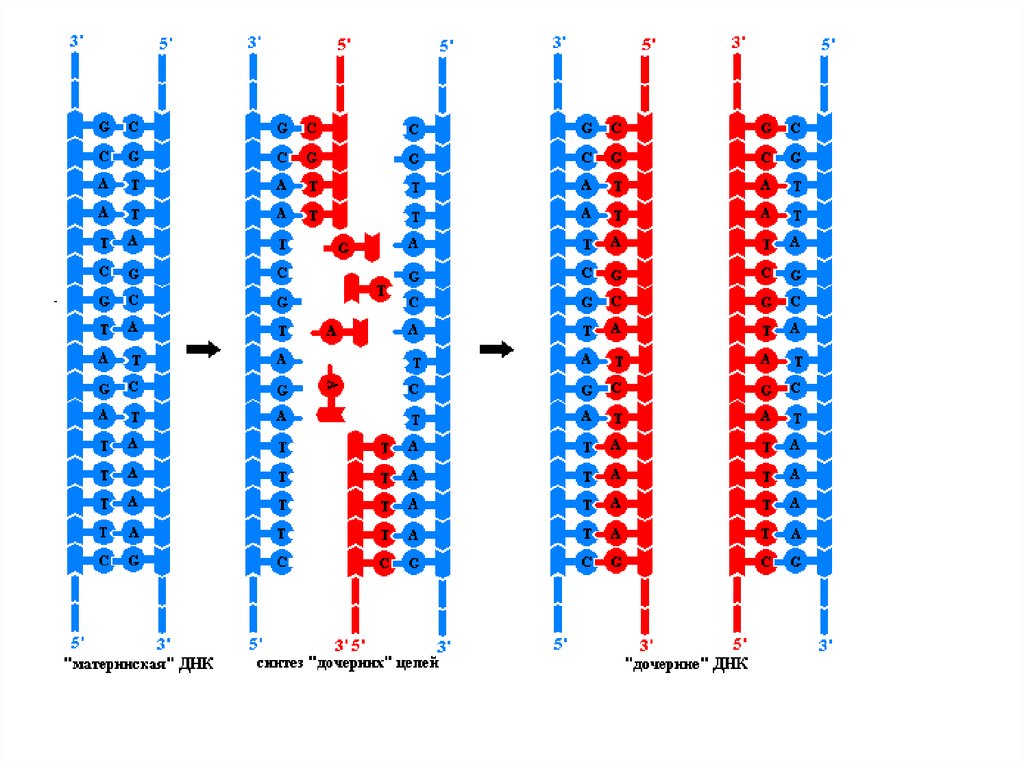
5. Основные принципы репликации ДНК
• Полуконсервативный механизм- каждая из 2 цепей ДНК служит
матрицей для образования новой
цепи. Образуются две новые
двухцепочечные молекулы ДНК,
каждая из которых состоит из
одной новой и одной старой цепей.
6.
7.
• Субстратами для синтеза ДНКявляются
дезоксирибонуклеозидтрифосфаты
– dАТФ, dГТФ, dТТФ, dЦТФ,
выполняющие роль строительного
материала и источников энергии.
8.
• Точка начала репликации – origin(ориджин).
• У бактерий в кольцевом геноме имеется
только одна точка «origin», тогда как у
эукариотических хромосом их множество.
• У человека репликация начинается в
точках, удаленных друг от друга от 30 до
300тыс.п.н.
9.
10.
• Репликон – это участок ДНК междудвумя «ориджинами» репликации.
11.
Родительская ДНКРепликационный глазок
Дочерние молекулы ДНК
Репликативные вилки
12. Репликация идет в двух направлениях
13.
• Синтез новых цепей ДНК можетпротекать только в направлении
5’ 3’, так как достраивается 3’конец цепи ДНК.
14.
5’3’
3’
5’
15. Синтез ДНК
16.
• Вся сложность ферментативного аппаратарепликации связана с требованиями
высокой точности синтеза.
• Весь этот комплекс называется ДНКрепликативной системой, или
• Реплисомой.
17. Ферменты репликации
• Синтез новой цепи ДНКосуществляется при помощи ДНКполимеразы.
• Фермент катализирует синтез
дочерних цепей на матрице ДНК по
принципу комплементарности.
18. Синтез ДНК
• У прокариот найдено 3 типа этихферментов:
• ДНК-полимераза I
• ДНК-полимераза II
• ДНК-полимераза III
19.
• ДНК-полимераза I выполняет функциипроверки поддержания порядка во время
репликации, репарации и рекомбинации.
• Обладает 5’ 3’ экзонуклеазной
активностью, может заменить участок ДНК
(или РНК), что используется в методах по
гибридизации нуклеиновых кислот.
20.
• ДНК-полимераза II – участвует впроцессах репарации ДНК при
повреждении ДНК ультрафиолетовым
облучением.
21.
• ДНК-полимераза III – главный ферментрепликации у E.coli.
• Состоит из субъединиц 10 разных типов.
• Обладает полимеразной активностью
(присоединяет 250-1000 нуклеотидов/с).
• Корректирующей активностью, т.е. 3’
5’ экзонуклеазной активностью, благодаря
которой каждый нуклеотид после присоединения
проверяется дважды.
22.
• Для создания Матрицы – одноцепочечнойДНК, необходимы хеликазы –
ферменты, разделяющие цепи
двухцепочечной ДНК на одинарные цепи.
23.
• Хеликазы – это ферменты, способныерасплетать две комплементарные нити в
ДНК с использованием энергии,
полученной при гидролизе АТФ.
24.
• Топоизомеразы – ферменты,изменяющие степень
сверхспирализации ДНК, путем
внесения одноцепочечных
или двухцепочечных разрывов в ДНК.
25.
• Топоизомеразы находятся передрепликативной вилкой, разрезают молекулу
ДНК для облегчения ее расплетания и
раскручивания молекулы ДНК, после чего
непрерывность ее восстанавливается.
26.
• Антибиотики – ингибиторы топоизомеразы:• Хинолоны, фторхинолоны
• Норфлоксацин
27.
• белки, стабилизирующиеразделенные нити ДНК – SSB
(single strand binding)
28.
• Роль SSB -белков заключается в том, что онисвязываются с однонитчатой ДНК, выпрямляют
ее и блокируют образование шпилечных
двухнитчатых структур.
• Участие SSB в репликации абсолютно
необходимо. Они удерживают матричные цепи
ДНК в репликативной вилке в одноцепочечном
состоянии, а также защищают одноцепочечную
ДНК от действия нуклеаз.
29.
• ДНК-лигаза – фермент катализирующийсшивание одноцепочечных фрагментов
ДНК.
• ДНК-праймаза — это фермент РНКполимераза, синтезирующий короткий
фрагмент РНК, называемый праймером,
комплементарный одноцепочечной матрице
ДНК.
30. Синтез ДНК
Синтез ДНК включает в себятри этапа:
• инициация
• элонгация
• терминация
31. Инициация
• инициация синтеза ДНК у E.coliпроисходит в oriC (245п.н.)
• В точках начала репликации отмечено
большое количество А=Т пар и
участки связывания ключевого
инициаторного белка DnaA.
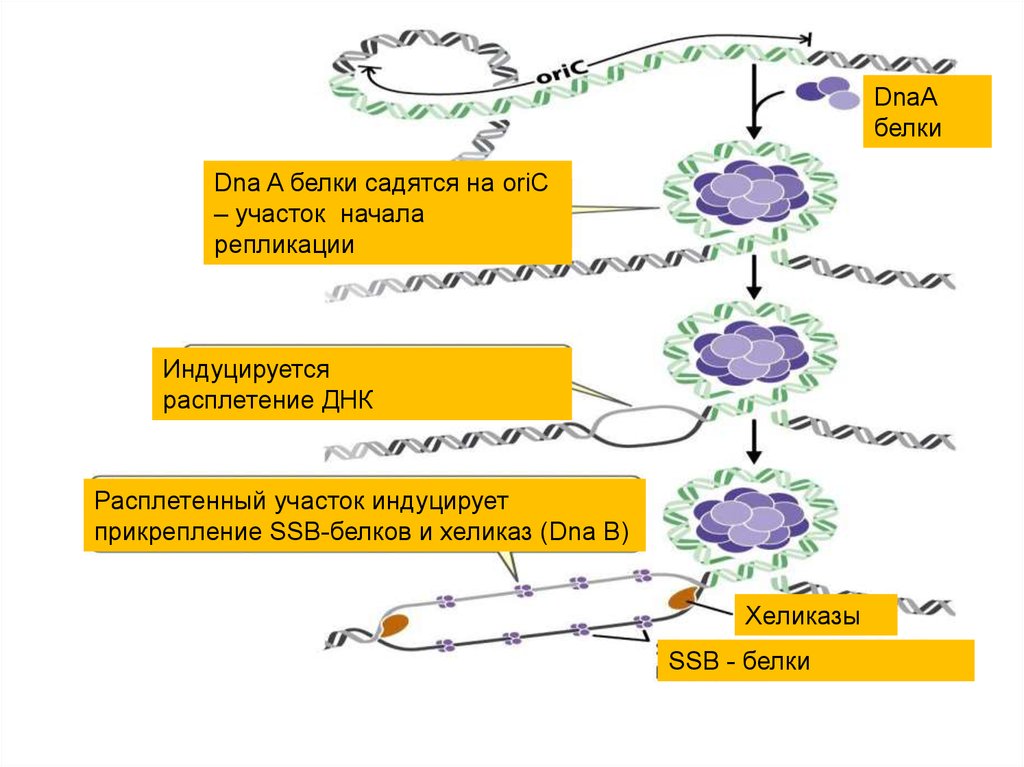
32.
• Процесс инициации начинается сприсоединения к хромосоме белка DnaA
(в origin области).
• 8 молекул DnaA, каждая из которых
связана с АТФ, образуют спиральный
комплекс (положительная
суперспираль). В соседних участках
ДНК это вызывает денатурацию А=Т
богатого участка.
33.
DnaАбелки
Dna A белки садятся на oriC
– участок начала
репликации
Индуцируется
расплетение ДНК
Расплетенный участок индуцирует
прикрепление SSB-белков и хеликаз (Dna B)
Хеликазы
SSB - белки
34.
• Это приводит к разделению цепей испособствует работе основного
расплетающего белка - хеликазы (DnaB).
• С образовавшейся одноцепочечной ДНК
связываются белки SSB, которые
стабилизируют вилку репликации.
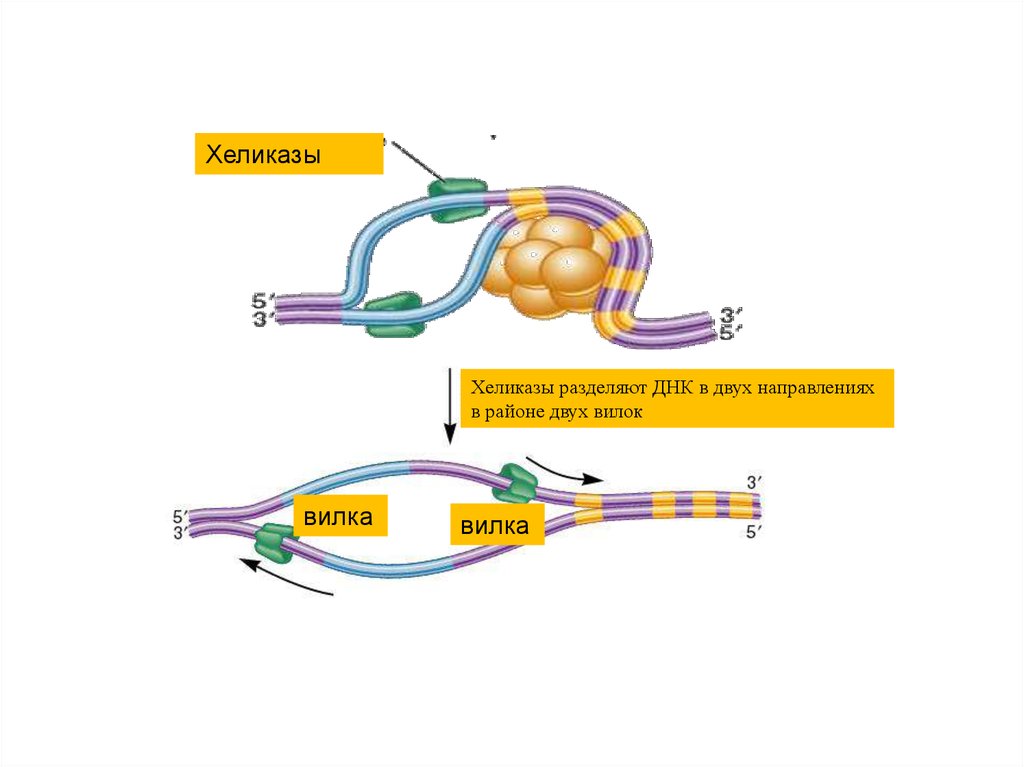
35.
ХеликазыХеликазы разделяют ДНК в двух направлениях
в районе двух вилок
вилка
вилка
36.
37. Синтез ДНК
• В результате действия хеликазы,топоизомеразы, SSB- белков,
ионов Mg+2 образуется
репликативная вилка - участок
ДНК, в пределах которого спираль
раскручена и разделена на
отдельные цепи.
38. Синтез ДНК
• ДНК-полимераза III не способнаначинать синтез новой цепи с ее
первого нуклеотида. Поэтому
репликация начинается с синтеза
праймера (РНК-затравки) на
обеих цепях расплетенной ДНК.
39. Синтез ДНК
• Праймер - короткий сегмент РНК,комплементарный матричной цепи
ДНК.
• Праймер синтезируется при
участии ДНК- зависимой РНКполимеразы (праймазы).
40. Синтез ДНК
• от 3’ –конца праймера начинаетсясинтез новой цепи ДНК при
помощи ДНК- полимераза III.
Синтез идет в направлении 5’ 3’
41. Синтез ДНК
• ДНК-полимераза III удлиняетРНК-затравку, присоединяя к ней
один за другим нуклеотиды,
комплементарные матричной цепи
42. Элонгация
• Синтез лидирующей цепи.• Синтез отстающей цепи.
43.
• Синтез лидирующей цепи начинается ссинтеза праймера, дезоксирибонуклеотиды
добавляются к этому праймеру ДНКполимеразой III, связанной с DnaВхеликазой.
• Синтез идет непрерывно со скоростью,
соответствующей скорости раскручивания
ДНК в репликативной вилке.
44. Синтез отстающей цепи
• Синтез осуществляется в виде короткихфрагментов ОКАЗАКИ.
• Каждый фрагмент Оказаки состоит
примерно из 1000 нуклеотидов
(у прокариот).
• Праймаза синтезирует РНК-праймер.
45.
• ДНК-полимераза III связывается спраймером и присоединяет
дезоксирибонуклеотиды.
• Синтез фрагментов Оказаки происходит с
участием ферментативного аппарата –
праймасомы ( DnaB-хеликаза и DnaGпраймаза) в составе репликативного
комплекса.
46.
• Реплисома присоединяет по 1000нуклеотидов в секунду к каждой цепи
(лидирующей и отстающей).
• После завершения сборки одного
фрагмента Оказаки его РНК-праймер
удаляется и замещается
последовательностью ДНК с помощью
ДНК-полимеразы I.
47.
• Оставшийся разрыв «сшивает» ДНК-лигаза.• ДНК-лигаза катализирует образование
фосфодиэфирной связи между 3’гидроксильной группой на конце одной
цепи ДНК и 5’-фосфатом на конце другой
цепи.
• Для этой реакции используется НАДкофактор в качестве источника АМФ (у
прокариот).
48.
49. Терминация
• У прокариот есть специальныетерминаторы (ter) – специальные
последовательности нуклеотидов,
прекращающие синтез цепи ДНК
• Ter-последовательности служат
участками связывания белка Tus
(terminus utilization substance)
50.
• Комплекс Ter-Tus может задержатьрепликативную вилку, движущуюся только
в одном направлении, т.е. останавливает
одну из вилок с которой сталкивается.
Другая вилка останавливается, когда
встречается с первой задержанной вилкой.
51.
• Для терминации репликации линейныхэукариотических хромосом на концах
каждой хромосомы синтезируются
специальные структуры, называемые
теломерами.
• Иначе с каждым клеточным делением
хромосомы становились бы короче и
короче.
52.
• Это многократно повторяющиесяпоследовательности нуклеотидов ( у
одноклеточных эукариот от 20 до100, у
млекопитающих превышает 1500).
• У человека такая последовательность
(TTAGGG)n .
53.
• Теломеры присоединяются к концамэукариотических хромосом с помощью
фермента теломеразы.
54.
• Одна из причин старения — закон делимости клеток,открытый американским биологом Леонардом
Хейфликом (Leonard Hayflick) в 1961 году. Суть его в
том, что клетки человеческого организма не могут
делиться бесконечно. Максимально возможное
количество делений в среднем составляет 50±10 (так
называемый предел или лимит Хейфлика).
• Вызвано это тем, что при делении клетки репликация ДНК
происходит "с потерями" — молекула копируется не до
конца. До определенного момента в этом ничего
страшного нет — кончики хромосом защищены
теломерами, которые и теряются при копировании..
55.
• есть клетки, которые могут делитьсябесконечно, например кроветворные или
раковые. В этих случаях механизм
ограничения количества делений не
срабатывает благодаря теломеразе, которая
"чинит" ДНК, достраивая теломеры.
56. Источники повреждения ДНК
УФ излучение
Радиация
Химические вещества
Ошибки репликации ДНК
Апуринизация - отщепление азотистых
оснований от сахарофосфатного остова
• Дезаминирование - отщепление аминогруппы от
азотистого основания
57. Репарация ДНК
• Репарация- процесс восстановленияповреждений ДНК.
• Распознавание дефекта
сопровождается непосредственно при
репликации, все ДНК-полимеразы
обладают 3’ 5’ экзонуклеазной
активностью.
58.
• Репарация осуществляется с помощью:специфического набора ферментов, постоянно
присутствующих в нормально функционирующих
клетках (фотореактивационная, эксцизионная)
• активации группы генов, контролирующих
различные клеточные функции –
SOS-репарация.
59.
• У бактерий имеются 2 ферментныесистемы, ведущие репарацию:
прямая
эксцизионная
60. Прямая репарация ДНК
• Фотореактивация. Расщеплениепиримидиновых димеров (приУФ
облучении) осуществляется
ферментом
ДНК - фотолиазой.
Реакция расщепления связей зависит от
видимого света. У человека отсутствует.
61.
фермент ДНК – фотолиаза - мономерный флавинзависимый фермент и 2 кофактора (хромофоры).• 5,10-метенилтетрагидрофолат (5,10-MTГФ) –
поглощает фотоны синего цвета (300-500 нм) и
передает энергию возбуждения на FADH- .
• Возбужденный FADH- отдает электрон
пиримидиновому димеру, устраняя повреждение.
62. Репарация ДНК
63. Репарация ДНК
• Темновая эксцизионнаярепарация
• Не нуждается в энергии видимого
света
64.
Каждая из систем репарации включает следующиекомпоненты:
• фермент, «узнающий» химически изменённые участки в
цепи ДНК и осуществляющий разрыв цепи вблизи от
повреждения.
• фермент, удаляющий повреждённый участок.
• фермент, синтезирующий соответствующий участок цепи
ДНК взамен удалённого.
• фермент, замыкающий последнюю связь в полимерной
цепи и тем самым восстанавливающий её непрерывность.
65. Эксцизионная репарация
• Base excision repair – BER• ДНК гликозилазы, распознают аномальные
основания ДНК и катализируют гидролитическое
расщепление N-glycosyl связи, между основанием
и сахаром.
• Образуется AP-сайт (apurinic/apyrimidinic).
• АП-сайт распознается АП-эндонуклеазой,
которая вводит в нить ДНК разрыв.
66.
• Фосфодиэстераза отщепляет от ДНКсахарофосфатную группу, к которой не
присоединено основание.
• Брешь размеров в 1 н. застраивается
ДНК-полимеразой I и концы ДНК
соединяются ДНК-лигазой.
67.
• Nucleotide excision repair - NER• Узнавание повреждений.
• Связывание мультисубъединичного комплекса с
поврежденным сайтом.
• Двойное надрезание поврежденной цепи на
несколько нуклеотидов.
от поврежденного сайта в обоих направлениях
5' и 3'.
68.
• Освобождение олигонуклеотида,содержащего повреждение между двумя
надрезами.
• Заполнение образовавшейся бреши ДНК
полимеразой.
• Лигирование.
69.
• При некоторых типах повреждений ДНК(двухнитевые разрывы,поперечные
сшивки).
• Репликативная вилка наталкивается на
нерепарированное повреждение ДНК.
• Результат воздействия ионизирующей
радиации.
• Окислительные реакции.
70. SOS-репарация
• SOS – белки всегда присутствуют в клетке, нопри запуске SOS-ответа их уровень значительно
повышается.
• Мутации, возникающие в результате такой
репликации, приводят к гибели одних клеток и
опасны для других, но репликация не
прекращается, позволяет выжить некоторым
мутантным дочерним клеткам.
71.
• В геноме типичной клетки млекопитающихза 24 ч аккумулируется много тысяч
повреждений. Благодаря репарации менее
одного повреждения из 1000 становится
мутацией. Изменения в генах репарации
значительно повышают чувствительность к
раку.
72.
• Все дефекты генов белков, участвующих вэксцизионной репарации связаны с
онкологическими заболеваниями,
• генетическими заболеваниями, например,
пигментная ксеродерма.
73.
• У человека нет ДНК-фотолиазы иэксцизионная репарация оснований –
единственный способ репарации
пиримидиновых димеров.
• Инактивация этой системы связана с
развитием рака кожи, который
индуцируется солнечным светом.
74. Молекулярные мутации
• Анемия Фанкони – генетическиеотклонения, возникающие при репарации
ДНК.
• Болезнь названа в честь швейцарского
педиатра, Гвидо Фанкони, впервые
описавшего это заболевание.
75.
• У 60—75 % больных также встречаются врожденные дефекты, такиекак низкорослость, ненормальная пигментация, маленькая голова,
аномалии скелета
• Ряд неврологических расстройств (косоглазие, недоразвитие одного
или обоих глаз, опущение века, глазное дрожание, глухота,
умственная отсталость),
• Поражения половых органов (недоразвитие половых органов)
• Почечные аномалии
• Врождённые пороки сердца.
• Средняя продолжительность жизни у больных анемией Фанкони
составляет около 30 лет.