



















 Медицина
Медицина Биология
БиологияПохожие презентации:










Гормональний стан самиць жуйних під час вагітності
1. ГОРМОНАЛЬНИЙ СТАН САМИЦЬ ЖУЙНИХ ПІД ЧАС ВАГІТНОСТІ
Неплідність при інфекційних таінвазійних захворюваннях
Виконала студентка 2 курсу магістратури
Косьянчук Таїса
2. 1.Гормональні зміни 2.Фолікуло- і овогенез 3.Перебіг вагітності та гормональний стан організму
1.ГОРМОНАЛЬНІ ЗМІНИ2.ФОЛІКУЛО- І ОВОГЕНЕЗ
3.ПЕРЕБІГ ВАГІТНОСТІ ТА
ГОРМОНАЛЬНИЙ СТАН ОРГАНІЗМУ
3. 1.Гормональні зміни
1.ГОРМОНАЛЬНІ ЗМІНИВнаслідок розвитку
зародка відбувається
перебудова не тільки
статевої системи, а й
гормонального балансу
самиці. Це відбувається
під впливом специфічних
для періоду вагітності
органів: жовтого тіла і
плаценти, які виділяють
гормони. В першій
половині вагітності
відіграють роль гормони
жовтого тіла, у другій –
гормони плаценти.
4.
В перші дні вагітності зростає концентрація в крові вільнихестрогенів, до 7 пг/мл на 2–5-йдень і 5 нг/мл на18-йдень.
Особливо вона зростає з 70-го дня вагітності, що зв’язане з
розростанням плаценти, яка продукує цей гормон. Вважають,
що висока концентрація естрогенів необхідна для збільшення
кровообігу у вагітному розі матки.
Висока концентрація естрогенів у молоці з 100-годня після
осіменіння є ефективним діагностичним тестом (РІА) на
вагітність.
З 10-годня зародок виробляє низькомолекулярну непротеїнову
речовину, яка стимулює секрецію жовтим тілом прогестерону, а
слизова оболонка матки виробляє щонайменше чотири інших
специфічних білки, які відіграють певну роль у захисті зародка
від дії маткових протеаз, а також імуносупресії.
З 13-годня зародок корови продукує стероїди, простагландини та
специфічні білки. На16–17-йдень зародок надсилає сигнал до
підтримання активності жовтого тіла ,а також, що він гальмує
вироблення ПГ-F2α у відповідь на введення окситоцину.
5.
Впродовж вагітності у рогатої худоби жовте тіло єголовним продуцентом прогестерону,
концентрація якого у циркулюючій крові з 20годня вагітності становить біля 10 нг/мл і
утримується на такому рівні майже до родів. З27го по111-йдень вагітності він з’являється в
амніотичній та алантоїсній рідині в концентрації
0,2–1,0нг/мл. З 200-годня вагітності прогестерон
виробляють також наднирники в концентрації1–
4нг/мл.
З 21–24-годня вагітності концентрація
прогестерону в молоці вагітних тварин буває
вищою, ніж у невагітних.
Концентрація ЛГ у сироватці крові утримується
протягом вагітності на мінімальному рівні (0,7–
1,0нг/мл). За два тижні до родів різко зростає
концентрація пролактину (з 80 до 200–400нг/мл),
підготовляючи молочну залозу до лактації.
Протягом перших 3–4місяців вагітності
концентрація плацентарного лактогену у
сироватці крові коливається нижче 50 нг/мл, а
з160–200-го дня зростає до 1000 нг/мл і
утримується на цьому рівні до родів. Цей гормон
проявляє вплив на розвиток молочної залози,
стероїдогенез у яєчнику та плаценті, а також на
розвиток плода .
6.
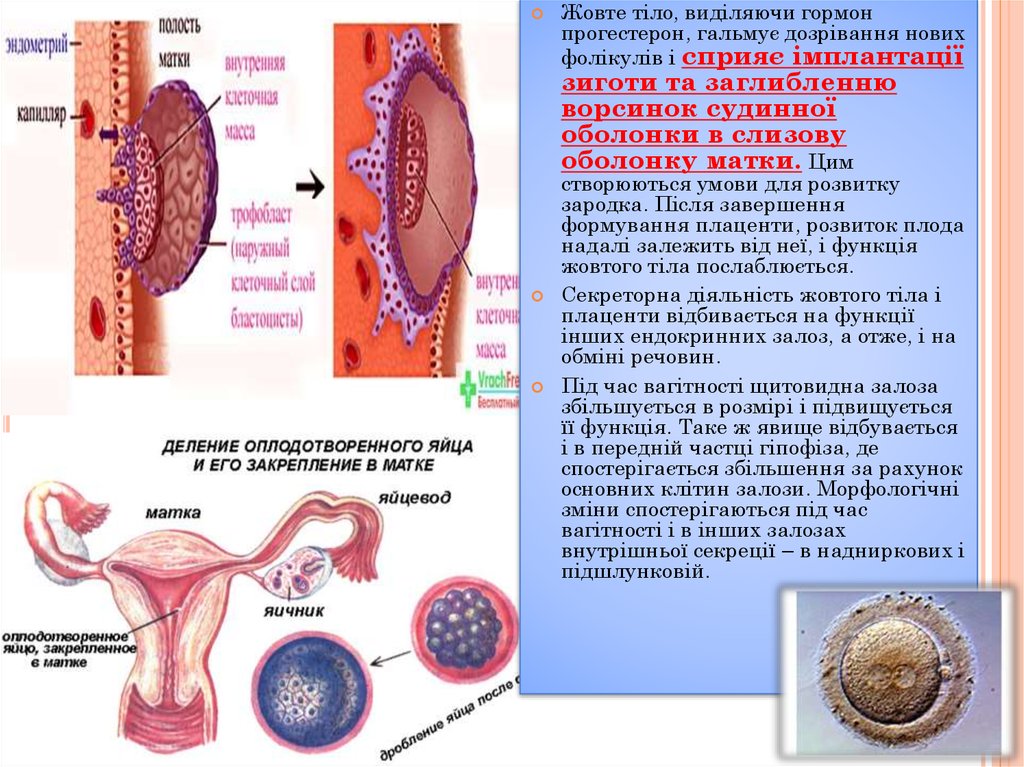
Жовте тіло, виділяючи гормонпрогестерон, гальмує дозрівання нових
фолікулів і сприяє імплантації
зиготи та заглибленню
ворсинок судинної
оболонки в слизову
оболонку матки. Цим
створюються умови для розвитку
зародка. Після завершення
формування плаценти, розвиток плода
надалі залежить від неї, і функція
жовтого тіла послаблюється.
Секреторна діяльність жовтого тіла і
плаценти відбивається на функції
інших ендокринних залоз, а отже, і на
обміні речовин.
Під час вагітності щитовидна залоза
збільшується в розмірі і підвищується
її функція. Таке ж явище відбувається
і в передній частці гіпофіза, де
спостерігається збільшення за рахунок
основних клітин залози. Морфологічні
зміни спостерігаються під час
вагітності і в інших залозах
внутрішньої секреції – в надниркових і
підшлунковій.
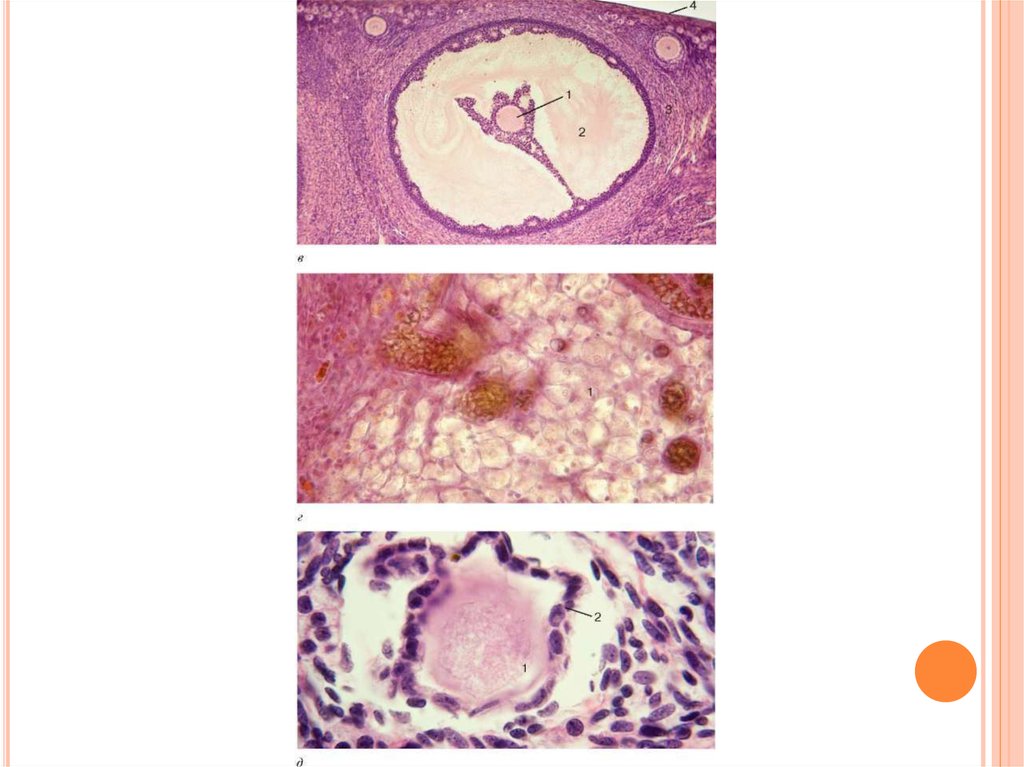
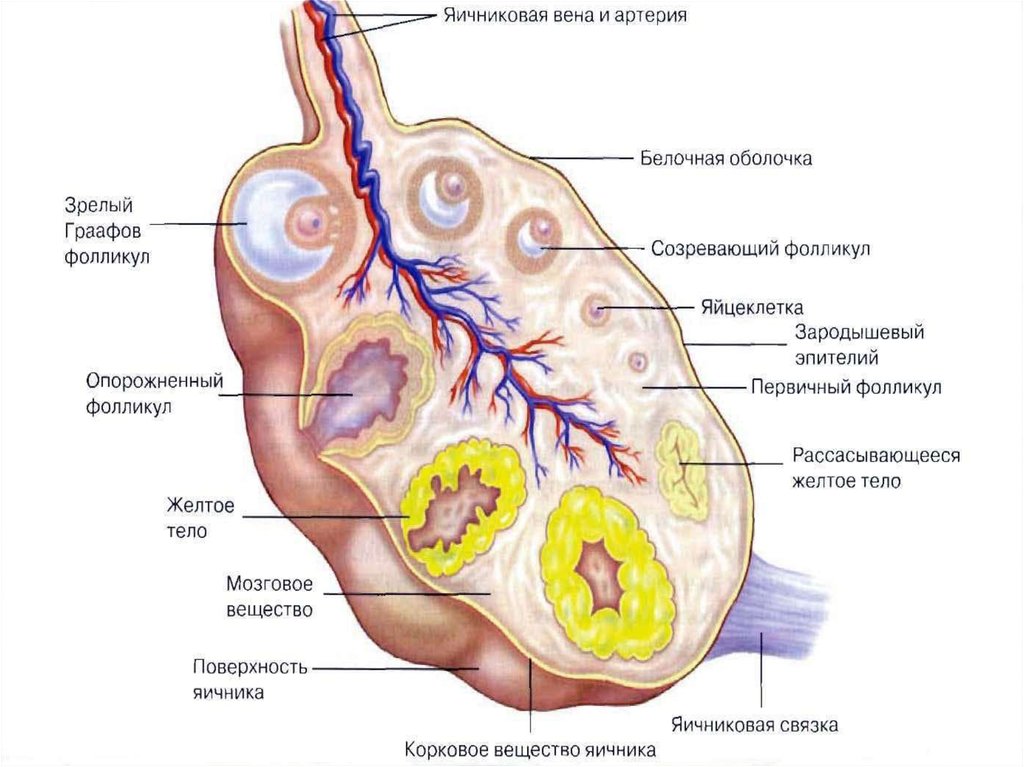
7. 2.Фолікуло- і овогенез
2.ФОЛІКУЛО- І ОВОГЕНЕЗФолікулогенез. Ріст овоцита супроводжується ростом і дозріванням
фолікула (фолікулогенезом), який умовно можна поділити на два
періоди: а) безперервного, генетично обумовленого росту фолікула, від
первинного
до
антрального
(порожнинного);
б)
гонадотропінзалежногоросту у фолікулінову стадію циклу. Фолікулярні клітини, що
оточують овоцит, при цьому збільшуються у об’ємі, перетворюються з
плоских у кубічні, а опісля – у циліндричні. Їх кількість різко зростає,
вони оточують овоцит декількома шарами (до18–20),які розташовуються
радіально і утворюють навколо яйця так званий променевий вінець.
Такий фолікул називається вторинним або фолікулом ІІ стадії. Його
діаметр у корови досягає від 300 мкм до 5 мм. Процес перетворення
первинного фолікула у вторинний досить тривалий, у великої рогатої
худоби і овець для цього необхідно біля 130 діб.
8.
9.
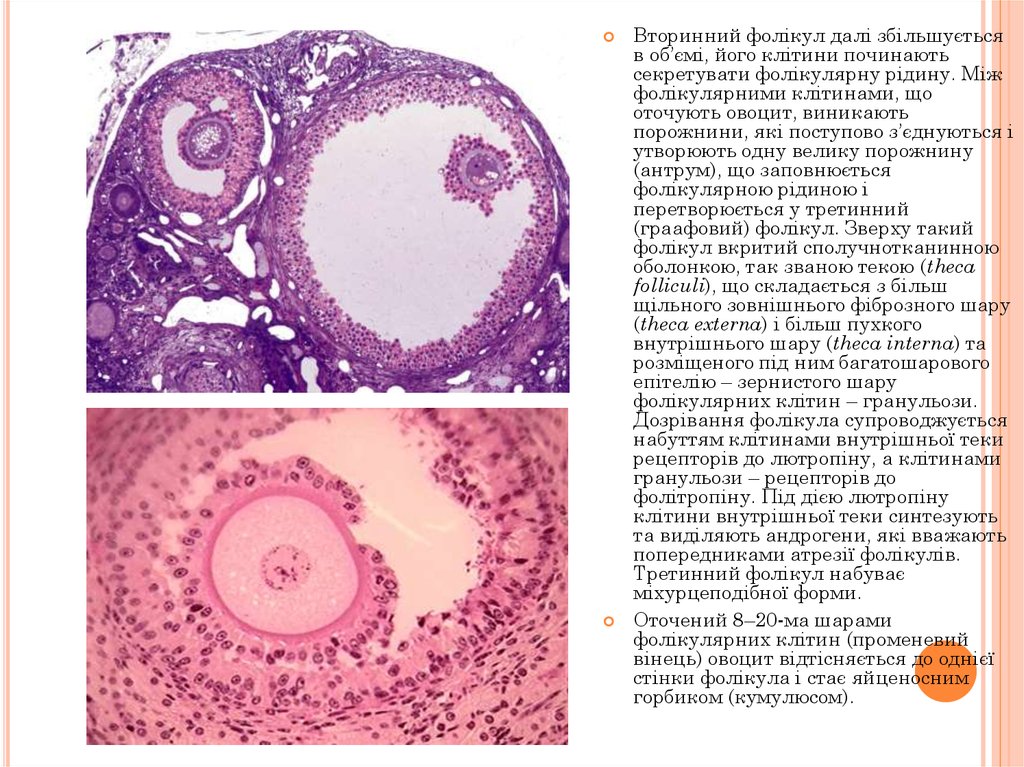
Вторинний фолікул далі збільшуєтьсяв об’ємі, його клітини починають
секретувати фолікулярну рідину. Між
фолікулярними клітинами, що
оточують овоцит, виникають
порожнини, які поступово з’єднуються і
утворюють одну велику порожнину
(антрум), що заповнюється
фолікулярною рідиною і
перетворюється у третинний
(граафовий) фолікул. Зверху такий
фолікул вкритий сполучнотканинною
оболонкою, так званою текою (theca
folliculi), що складається з більш
щільного зовнішнього фіброзного шару
(theca externa) і більш пухкого
внутрішнього шару (theca interna) та
розміщеного під ним багатошарового
епітелію – зернистого шару
фолікулярних клітин – гранульози.
Дозрівання фолікула супроводжується
набуттям клітинами внутрішньої теки
рецепторів до лютропіну, а клітинами
гранульози – рецепторів до
фолітропіну. Під дією лютропіну
клітини внутрішньої теки синтезують
та виділяють андрогени, які вважають
попередниками атрезії фолікулів.
Третинний фолікул набуває
міхурцеподібної форми.
Оточений 8–20-ма шарами
фолікулярних клітин (променевий
вінець) овоцит відтісняється до однієї
стінки фолікула і стає яйценосним
горбиком (кумулюсом).
10.
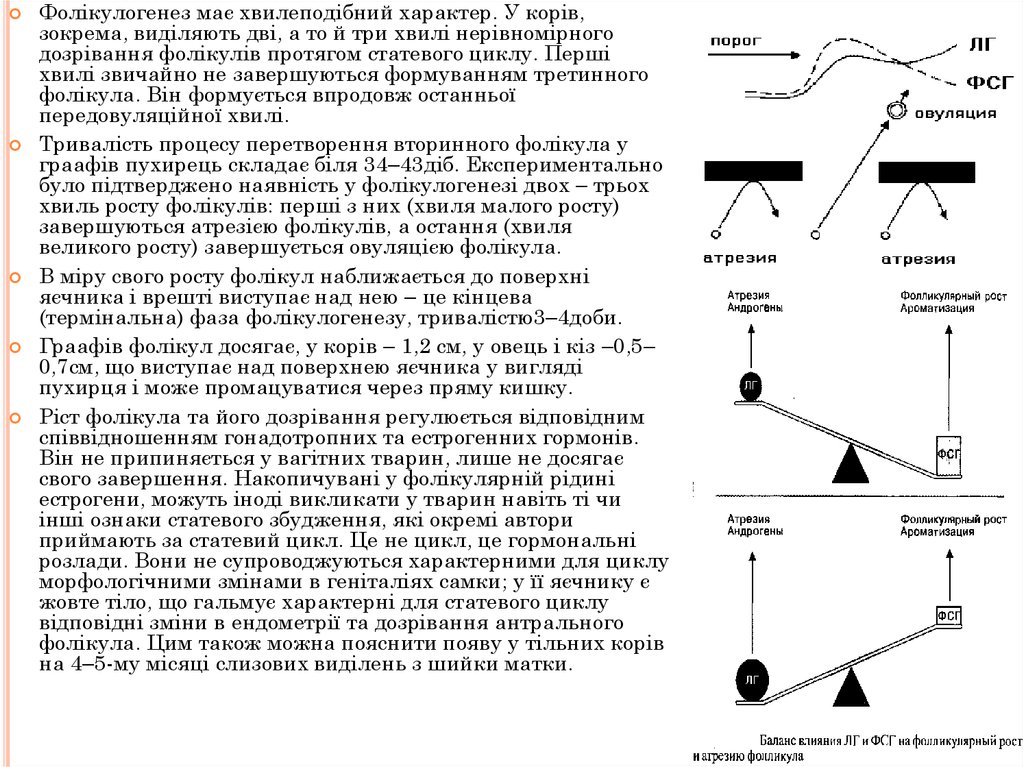
Фолікулогенез має хвилеподібний характер. У корів,зокрема, виділяють дві, а то й три хвилі нерівномірного
дозрівання фолікулів протягом статевого циклу. Перші
хвилі звичайно не завершуються формуванням третинного
фолікула. Він формується впродовж останньої
передовуляційної хвилі.
Тривалість процесу перетворення вторинного фолікула у
граафів пухирець складає біля 34–43діб. Експериментально
було підтверджено наявність у фолікулогенезі двох – трьох
хвиль росту фолікулів: перші з них (хвиля малого росту)
завершуються атрезією фолікулів, а остання (хвиля
великого росту) завершується овуляцією фолікула.
В міру свого росту фолікул наближається до поверхні
яєчника і врешті виступає над нею – це кінцева
(термінальна) фаза фолікулогенезу, тривалістю3–4доби.
Граафів фолікул досягає, у корів – 1,2 см, у овець і кіз –0,5–
0,7см, що виступає над поверхнею яєчника у вигляді
пухирця і може промацуватися через пряму кишку.
Ріст фолікула та його дозрівання регулюється відповідним
співвідношенням гонадотропних та естрогенних гормонів.
Він не припиняється у вагітних тварин, лише не досягає
свого завершення. Накопичувані у фолікулярній рідині
естрогени, можуть іноді викликати у тварин навіть ті чи
інші ознаки статевого збудження, які окремі автори
приймають за статевий цикл. Це не цикл, це гормональні
розлади. Вони не супроводжуються характерними для циклу
морфологічними змінами в геніталіях самки; у її яєчнику є
жовте тіло, що гальмує характерні для статевого циклу
відповідні зміни в ендометрії та дозрівання антрального
фолікула. Цим також можна пояснити появу у тільних корів
на 4–5-му місяці слизових виділень з шийки матки.
11.
12.
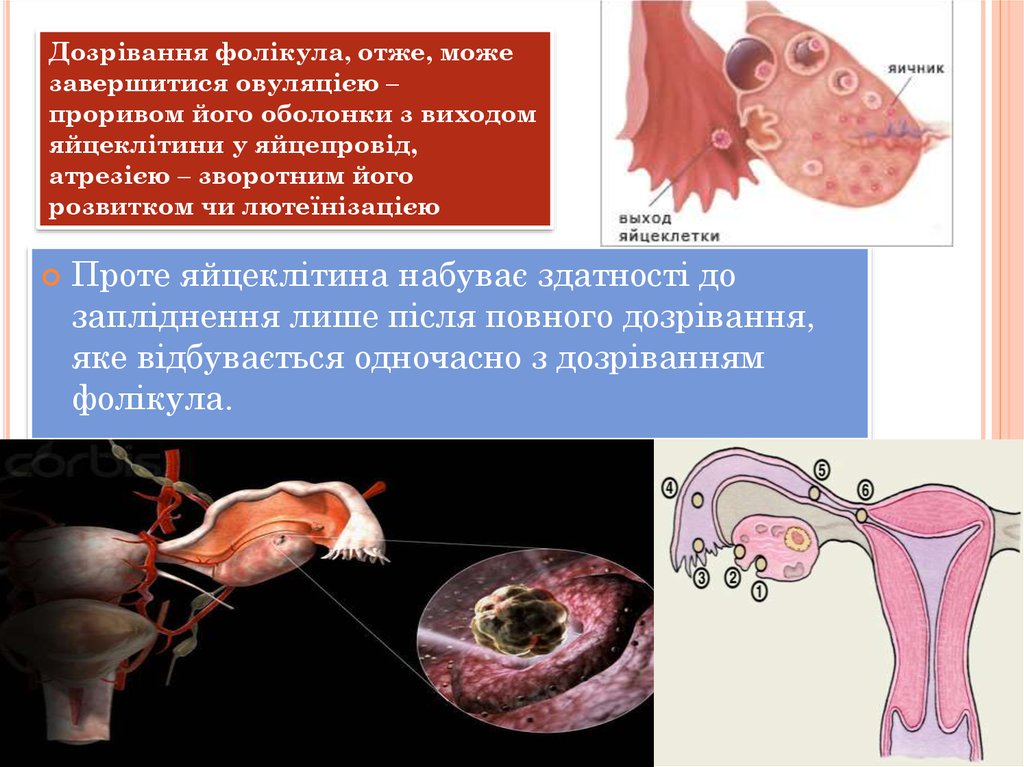
Дозрівання фолікула, отже, можезавершитися овуляцією –
проривом його оболонки з виходом
яйцеклітини у яйцепровід,
атрезією – зворотним його
розвитком чи лютеїнізацією
Проте яйцеклітина набуває здатності до
запліднення лише після повного дозрівання,
яке відбувається одночасно з дозріванням
фолікула.
13.
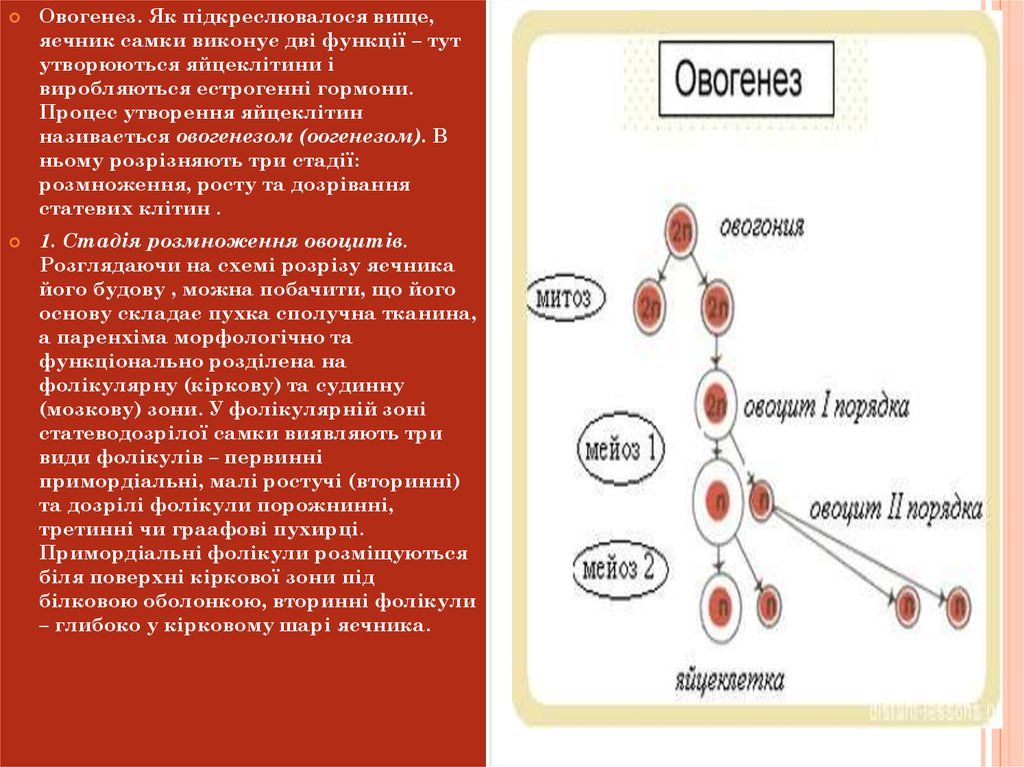
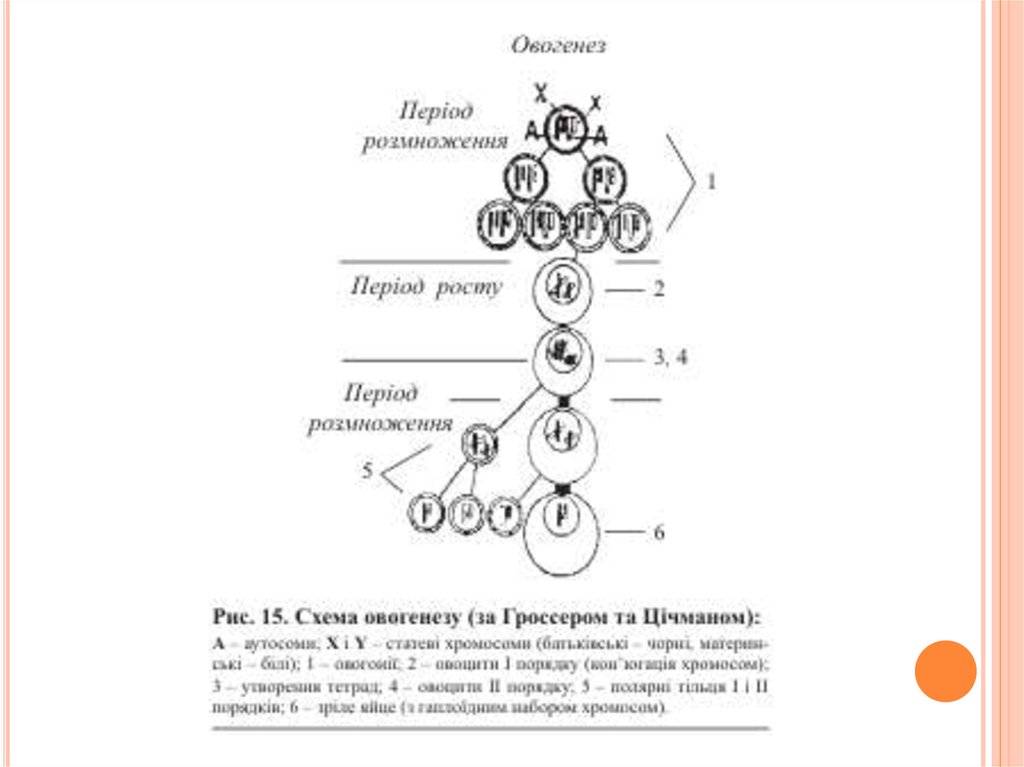
3. Стадія дозрівання. На перших двох стадіях овогенезу овоцитиволодіють властивим для кожного виду тварин повним, диплоїдним
набором хромосом. Незадовго до овуляції починається дозрівання
овоцита – складні якісні зміни його з перебудовою ядра, протоплазми
та обміну речовин, що супроводжуються двома послідовними
поділами: редукційним, спрямованим на зменшення кількості
хромосом, і екваційним – вирівнюючим. В результаті редукційного
мітотичного поділу, що зачіпає лише ядро, утворюються два нерівні
за величиною ядра – велике та маленьке (перше напрямне чи
полярне тільце). У велике ядро переходить половина хромосом і
основна маса каріоплазми, а в друге – абортивне – друга половина
хромосом і незначна частина каріоплазми. Напрямне тільце при
цьому виштовхується у позажовтковий простір. Таким чином, в
результаті редукційного поділу овоцита І-гопорядку утворюється
овоцит другого порядку з половинним набором хромосом.
Перший (редукційний) поділ звичайно буває незадовго до овуляції.
Другий (екваційний) поділ, що відбувається вже в яйцепроводі після
проникнення уяйце спермія, також буває нерівномірним: знову ядро
ділиться абортивно на велике та друге полярне тільце, яке також
викидається у позажовтковий простір. Оскільки другий (екваційний)
поділ гомотипний, то новоутворена клітина – дозріла яйцеклітина
також має половинну кількість хромосом.
Перше полярне тільце може також розділитись на два. Отже, в
процесі овогенезу овоцит 1 порядку перетворюється на дозріле яйце з
гаплоїдним набором хромосом та двома – трьома полярними
тільцями. Вважають, що напрямні тільця не беруть участі у процесі
запліднення. Вони просто служать сховищем для збереження
“зайвого” набору хромосом.
14.
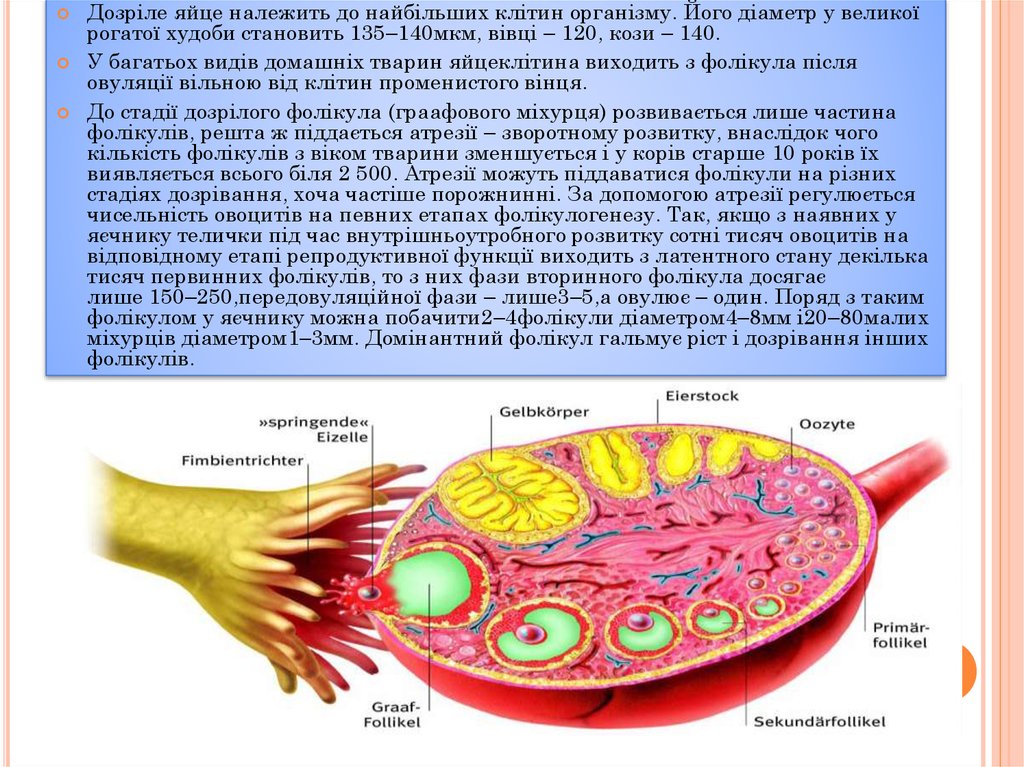
Дозріле яйце належить до найбільших клітин організму. Його діаметр у великоїрогатої худоби становить 135–140мкм, вівці – 120, кози – 140.
У багатьох видів домашніх тварин яйцеклітина виходить з фолікула після
овуляції вільною від клітин променистого вінця.
До стадії дозрілого фолікула (граафового міхурця) розвивається лише частина
фолікулів, решта ж піддається атрезії – зворотному розвитку, внаслідок чого
кількість фолікулів з віком тварини зменшується і у корів старше 10 років їх
виявляється всього біля 2 500. Атрезії можуть піддаватися фолікули на різних
стадіях дозрівання, хоча частіше порожнинні. За допомогою атрезії регулюється
чисельність овоцитів на певних етапах фолікулогенезу. Так, якщо з наявних у
яєчнику телички під час внутрішньоутробного розвитку сотні тисяч овоцитів на
відповідному етапі репродуктивної функції виходить з латентного стану декілька
тисяч первинних фолікулів, то з них фази вторинного фолікула досягає
лише 150–250,передовуляційної фази – лише3–5,а овулює – один. Поряд з таким
фолікулом у яєчнику можна побачити2–4фолікули діаметром4–8мм і20–80малих
міхурців діаметром1–3мм. Домінантний фолікул гальмує ріст і дозрівання інших
фолікулів.
15.
Такимчином,
протягом
статевого
циклу
відбувається дві – три хвилі росту і регресії (чи
овуляції) домінантних фолікулів, а саме: з 3-гопо13й,13-гопо18-йі
з18-гопо0-й.(“нульовим”
днем
статевого циклу вважають1-йдень нового циклу), що
супроводжується
трьома
хвилями
виділення
естрадіолу (тривалістю 7 днів) дояєчниковоматковоїсудини.
Перший і другий домінантний фолікул піддаються
регресії, а третій (діаметром 12,8 мм) овулює.
Роль перших домінантних фолікулів невідома.
Вважають, що перший з них може бути джерелом
естрогенів,
необхідних
для
транспортування
яйцеклітини до матки, а естрогени другого
домінантного фолікула можуть мати значення у
стимулюванні рецепторів окситоцину в матці та
лютеолізу жовтого тіла.
Поява домінантного фолікула супроводжується
інтенсивною секрецією естрадіолу та інгібіну.
Останній гальмує ріст інших фолікулів через
зменшення
виділення
гіпофізом
ФСГ.
Така
черговість може проявлятися декілька раз протягом
статевого циклу, що кожен раз завершується
атрезією домінуючого фолікула, крім останнього,
передовуляційного.
Дві третини фолікулів піддаються атрезії.
Механізм атрезії остаточно не з’ясований. Одні
автори вважають, що вона розпочинається із
загибелі
та
десквамації
клітин
гранульози,
відшарування
її
від
базальної
капсули
і
розсіювання
клітин
променевого
вінця.
Це
позбавляє овоцит живлення і він дегенерує. Інші
вважають, що фолікул припиняє свій ріст, наявна у
ньому яйцеклітина піддається морфологічним
змінам (каріопікноз, каріолізис, гіаліноз), що
врешті викликає її загибель з подальшим розкладом
зернистого шару та сполучнотканинної оболонки,
розсмоктуванням
фолікулярної
рідини.
У
атретичних фолікулах стероїдогенез зупиняється на
стадії андрогенів без ароматизації їх у естрогени.
16.
Порожнина атретичного фолікула спадається, фолікул проростаєсполучною тканиною і піддається гіаліновому переродженню,
продукти розкладу фолікула розсмоктуються і на його місці
виявляється фіброзне тільце, яке згодом зникає.
Атрезія фолікулів як фізіологічний процес, може проявлятися на
різних стадіях ово та фолікулогенезу, як під час внутрішньоутробного,
постфетального, так і протягом репродуктивного періоду життя самки.
Механізм овуляції досить складний і остаточно не вивчений. Його
можна уявити собі таким чином: незадовго до овуляції зростає
приплив крові до яєчника, у фолікулі збільшується кількість
фолікулярної рідини, він досягає властивої для кожного виду тварин
величини, відтісняє на боки навколишні тканини, наближається до
поверхні яєчника і випинається над ним у вигляді щільного
флуктуючого міхурця. Оболонка фолікула при цьому сильно
розтягується, тоншає, в її гранульозі з’являються типові лютеїнові
клітини з вакуолізованою цитоплазмою, його стінки піддаються
частковому лізису; відбувається сильний приплив крові як до
фолікула, так і до лійки яйцепроводу, вона щільно обхоплює яєчник;
тоді судини звужуються, стінка фолікула тоншає, на її поверхні
з’являється позбавлене судин та фолікулярних клітин одне або
декілька вторинних підвищень, крізь які просвічується рідина.
Врешті під впливом розрихлюючої дії ферменту колагенази та
зростаючого внутрішньофолікулярного тиску фолікул розривається, в
ньому утворюється овальний отвір, крізь який повільно витікає
фолікулярна рідина, змиваючи у яйцепровід частину клітин
фолікулярного епітелію разом з яйцеклітиною. Попаданню
яйцеклітини у яйцепровід сприяють його скорочення.
17.
18.
19.
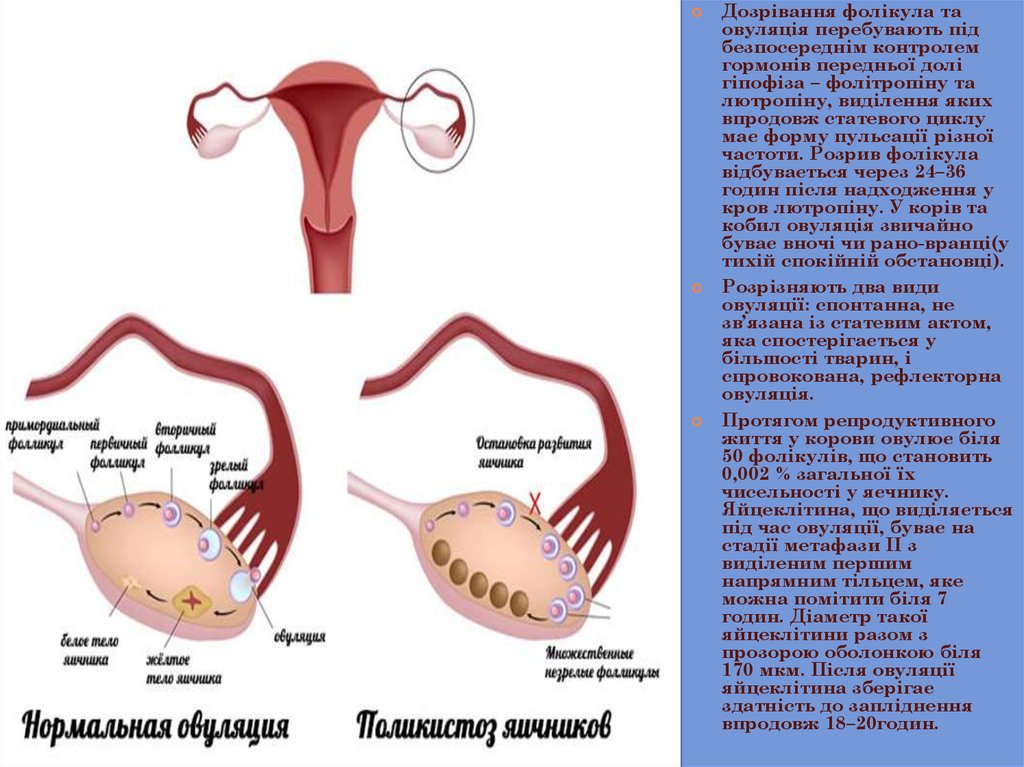
Дозрівання фолікула таовуляція перебувають під
безпосереднім контролем
гормонів передньої долі
гіпофіза – фолітропіну та
лютропіну, виділення яких
впродовж статевого циклу
має форму пульсації різної
частоти. Розрив фолікула
відбувається через 24–36
годин після надходження у
кров лютропіну. У корів та
кобил овуляція звичайно
буває вночі чи рано-вранці(у
тихій спокійній обстановці).
Розрізняють два види
овуляції: спонтанна, не
зв’язана із статевим актом,
яка спостерігається у
більшості тварин, і
спровокована, рефлекторна
овуляція.
Протягом репродуктивного
життя у корови овулює біля
50 фолікулів, що становить
0,002 % загальної їх
чисельності у яєчнику.
Яйцеклітина, що виділяється
під час овуляції, буває на
стадії метафази ІІ з
виділеним першим
напрямним тільцем, яке
можна помітити біля 7
годин. Діаметр такої
яйцеклітини разом з
прозорою оболонкою біля
170 мкм. Після овуляції
яйцеклітина зберігає
здатність до запліднення
впродовж 18–20годин.
20.
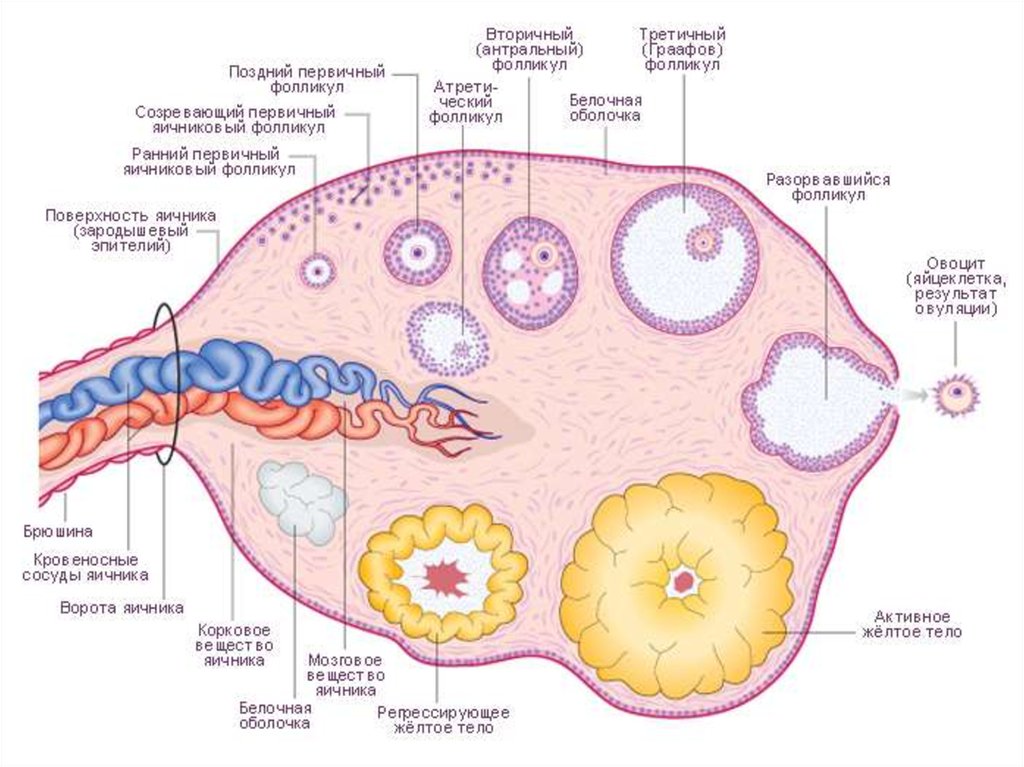
Овогенез. Як підкреслювалося вище,яєчник самки виконує дві функції – тут
утворюються яйцеклітини і
виробляються естрогенні гормони.
Процес утворення яйцеклітин
називається овогенезом (оогенезом). В
ньому розрізняють три стадії:
розмноження, росту та дозрівання
статевих клітин .
1. Стадія розмноження овоцитів.
Розглядаючи на схемі розрізу яєчника
його будову , можна побачити, що його
основу складає пухка сполучна тканина,
а паренхіма морфологічно та
функціонально розділена на
фолікулярну (кіркову) та судинну
(мозкову) зони. У фолікулярній зоні
статеводозрілої самки виявляють три
види фолікулів – первинні
примордіальні, малі ростучі (вторинні)
та дозрілі фолікули порожнинні,
третинні чи граафові пухирці.
Примордіальні фолікули розміщуються
біля поверхні кіркової зони під
білковою оболонкою, вторинні фолікули
– глибоко у кірковому шарі яєчника.
21.
22.
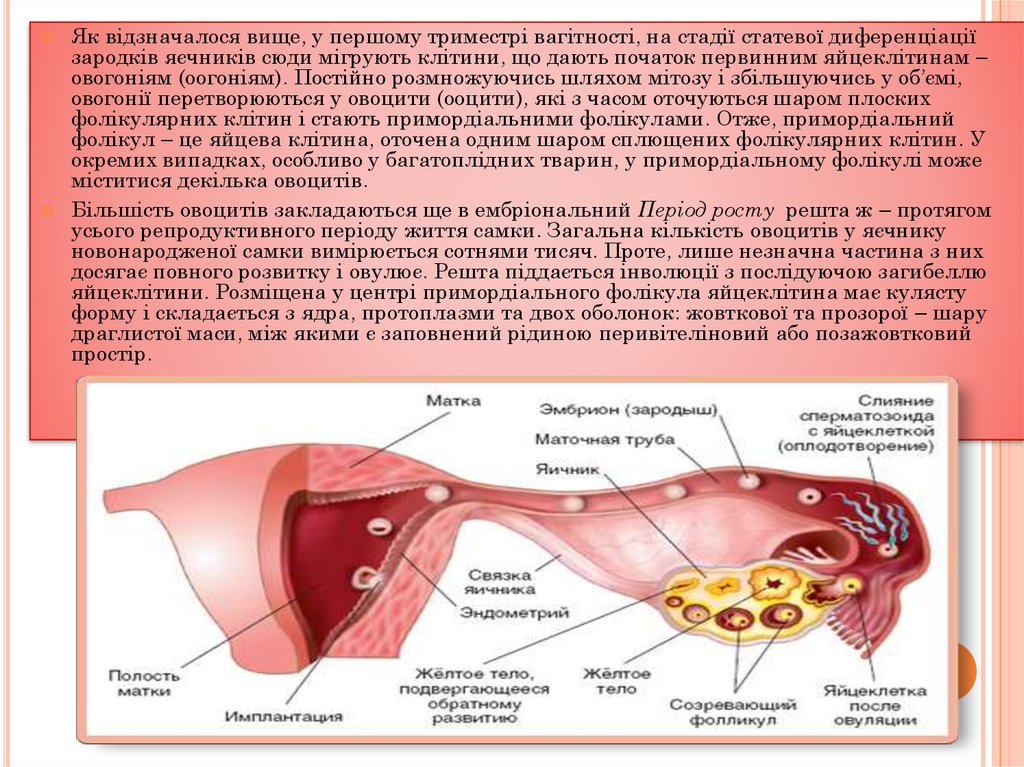
Як відзначалося вище, у першому триместрі вагітності, на стадії статевої диференціаціїзародків яєчників сюди мігрують клітини, що дають початок первинним яйцеклітинам –
овогоніям (оогоніям). Постійно розмножуючись шляхом мітозу і збільшуючись у об’ємі,
овогонії перетворюються у овоцити (ооцити), які з часом оточуються шаром плоских
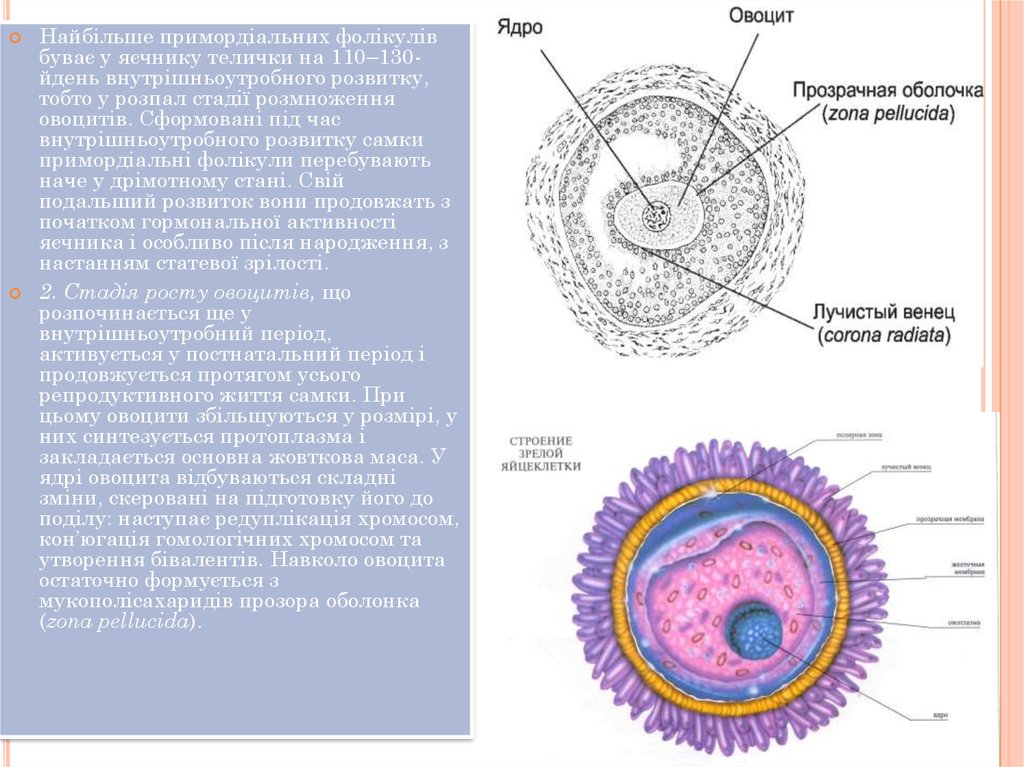
фолікулярних клітин і стають примордіальними фолікулами. Отже, примордіальний
фолікул – це яйцева клітина, оточена одним шаром сплющених фолікулярних клітин. У
окремих випадках, особливо у багатоплідних тварин, у примордіальному фолікулі може
міститися декілька овоцитів.
Більшість овоцитів закладаються ще в ембріональний Період росту решта ж – протягом
усього репродуктивного періоду життя самки. Загальна кількість овоцитів у яєчнику
новонародженої самки вимірюється сотнями тисяч. Проте, лише незначна частина з них
досягає повного розвитку і овулює. Решта піддається інволюції з послідуючою загибеллю
яйцеклітини. Розміщена у центрі примордіального фолікула яйцеклітина має кулясту
форму і складається з ядра, протоплазми та двох оболонок: жовткової та прозорої – шару
драглистої маси, між якими є заповнений рідиною перивітеліновий або позажовтковий
простір.
23.
24.
Найбільше примордіальних фолікулівбуває у яєчнику телички на 110–130йдень внутрішньоутробного розвитку,
тобто у розпал стадії розмноження
овоцитів. Сформовані під час
внутрішньоутробного розвитку самки
примордіальні фолікули перебувають
наче у дрімотному стані. Свій
подальший розвиток вони продовжать з
початком гормональної активності
яєчника і особливо після народження, з
настанням статевої зрілості.
2. Стадія росту овоцитів, що
розпочинається ще у
внутрішньоутробний період,
активується у постнатальний період і
продовжується протягом усього
репродуктивного життя самки. При
цьому овоцити збільшуються у розмірі, у
них синтезується протоплазма і
закладається основна жовткова маса. У
ядрі овоцита відбуваються складні
зміни, скеровані на підготовку його до
поділу: наступає редуплікація хромосом,
кон’югація гомологічних хромосом та
утворення бівалентів. Навколо овоцита
остаточно формується з
мукополісахаридів прозора оболонка
(zona pellucida).
25. 3.Перебіг вагітності та гормональний стан організму
3.ПЕРЕБІГ ВАГІТНОСТІ ТАГОРМОНАЛЬНИЙ СТАН ОРГАНІЗМУ
Складність змін у гормональній системі вагітної визначається тим,
що на діяльність залоз внутрішньої секреції самки дуже великий
вплив мають гормони плаценти, а також плода.
Гіпофіз збільшується при вагітності в 2 – 3 рази. Це ще раз
підтверджує той факт, що при вагітності ендокринна система
трудиться над забезпеченням всіх систем організму. Гіпофіз, як
«диригент» ендокринної системи, збільшується в розмірах і
починає працювати інтенсивніше. Перш за все це виражається в
різкому зменшенні вироблення гормонів, що регулюють роботу
статевих залоз(фолікулостимулюючого (ФСГ) і лютеїнізуючого
(ЛГ) гормонів). Це супроводжується природним гальмуванням
дозрівання яйцеклітин в яєчниках; процес овуляції – виходу
яйцеклітин у черевну порожнину – також припиняється.
Вироблення пролактину, відповідального за становлення лактації,
під час вагітності, навпаки, зростає і до моменту родів збільшується
в 5 – 10 разів у порівнянні з показниками, характерними для
невагітних самок. Оскільки зростання вироблення пролактину
починається в першому триместрі вагітності, то зміни молочних
залоз з'являються вже в цей період.
У самок спостерігаються підвищення вироблення тиреотропного
гормону (ТТГ), що регулює функцію щитовидної залози.
Під час вагітності збільшується також
вироблення адренокортикотропного гормону (АКТГ), який
регулює вироблення гормонів надниркових залоз.
26.
27.
Концентрація що утворюється вгіпофізі окситоцину зростає в кінці вагітності і під час
пологів, що є одним із пускових механізмів родової
діяльності. Основною властивістю окситоцину є здатність
викликати сильні скорочення мускулатури матки,
особливо вагітної. Існує синтетичний аналог цього
гормону, який вводять самкам при розвитку слабкості
родової діяльності. Окситоцин сприяє також виділенню
молока з молочних залоз.
Виникнення і розвиток вагітності пов'язані з роботою
нової залози внутрішньої секреції – жовтого тіла
вагітності. Ця залоза формується в яєчнику на місці
яйцеклітини. У жовтому тілі виробляються статеві
гормони (прогестерон і естроген), яким належить
величезна роль в імплантації та подальший розвиток
вагітності. Особливу роль у першому триместрі
вагітності грає прогестерон, що виділяється жовтим
тілом: він є основним гормоном цього періоду, тому
обумовлює імплантацію й утримання плодового яйця в
матці. У разі нестачі прогестерону виникає загроза
переривання вагітності. Для корекції цього стану
застосовують препарати, що містять синтетичні аналоги
прогестерону, – Дюфастон, УТРОЖЕСТАН. З 3 – 4-го
місяця вагітності жовте тіло піддається інволюції
(зворотному розвитку) і його функцію цілком бере на
себе плацента. Стимуляція жовтого тіла
здійснюється хоріонічним гонадотропіном, який
виробляється в гіпофізі.
28.
У тварини після запліднення змінюєтьсяобмін речовин і появляється добрий апетит,
що сприяє кращій їх вгодованості. Це видно з
того, що шерстний покрив стає гладким і
блискучим, форми тіла округляються і
повніють. У другій половині вагітності,
незважаючи на те, що апетит зберігається,
тварина марніє, бо вона не встигає
асимілювати достатню кількість поживних
речовин, потрібну для себе, і для плода, який
швидко розвивається. Через це відбувається
витрачання резервів, пластичних речовин,
зібраних протягом першої половини
вагітності. Це чергування доброї вгодованості
і похудання наочно відбивається на рості
копитець та рогів тварини. Потовщення на
рогах (“кільце”) з’являється у вагітних
тварин у першій половині вагітності, а поява
кільцевих жолобків – в другій половині
вагітності (по кільцях визначають, скільки
було отелень у корови).
Поведінка тварини значно змінюється в
другій половині вагітності. Вона стає
спокійнішою, а в міру розвитку вагітності
спостерігається обережність у рухах; тварина
швидко стомлюється і в русі порівняно
швидко починає потіти.
29.
Кількість крові у вагітних тварин збільшена, і серце трохи змінюється за рахунокдеякого збільшення маси м’язових елементів. Кількість кальцію в крові звичайно
зменшена, бо на початку вагітності він скупчується в органах і в плаценті, а в другій
половині йде на побудову скелета плода. Лужність крові знижується.
Дихання у вагітних тварин прискорюється, бо потреба в кисні збільшується за
рахунок плода, що швидко розвивається. А проте воно утруднюється, бо плід тисне
на діафрагму з боку черевної порожнини, і тип дихання переходить зреберно-черевного у реберний. Відзначають навіть випадки задишки, яка зумовлюється наявністю
в матці дуже великого плода або багатоплідністю.
Удругій половині вагітності тварини часто виділяють невеликими порціями сечу й
кал, бо матка, збільшуючись, дедалі зменшує місткість черевної порожнини і
посилює тиск на розміщені в ній внутрішні органи. В свою чергу, внутрішні органи
тиснуть на черевні стінки, внаслідок чого об’єм черева збільшується.
В останню чверть вагітності матка дуже тисне на великі венозні стовбури, що
відводять кров від задніх кінцівок і з черевної порожнини. Це спричиняє деяку
набряклість в ділянці тазових органів та статевих губ.
30.
Посилений приплив крові до судин тазової порожнини спричиняє розтягнення стінок капілярів,через що рідка частина крові виходить з них і просочує навколишні тканини. Це в певній мірі
сприяє розм’якшенню зв’язок, які сполучають крижі з тазом, а потім крижі западають.
Розслаблення зв’язок та інших тканин підготовляє родові шляхи до наступного процесу родів.
Вагітність впливає на ріст молочної залози. Особливо це помітно у нетелей і кіз, у яких уже в
середині вагітності спостерігається збільшення залозистої тканини вим’я. У дійних корів і кіз
внаслідок здоювання не помічається збільшення залозистої тканини, і під час сухостійного
періоду вим’я протягом тривалого часу відзначається своїм малим розміром і дряблістю. Тільки
за кілька тижнів чи навіть днів до родів помічається збільшення залозистої тканини вим’я.
Виникнення та розвиток вагітності позначається і на імунному стані організму самки .Імунна
система, як відомо, здійснює імунологічний нагляд за появою в організмі сторонніх антигенів.
Кожна тканина, в тому числі й сперма,яйцеклітина, ембріон та плід є носіями видоспецифічних,
стадіоспецифічних чи органо(тканино)специфічних антигенів. Безумовно, вони викликають на
себе імунні реакції, в результаті яких розвивається стан толерантності (терпимості) чи
несприйнятності (відторгнення).
31.
В імунних реакціях беруть участь три типи імунокомпетентних клітин:Т-лім-фоцити,Влімфоцитита макрофаги.Т-лімфоцитирозпізнають генетично сторонній матеріал і передаютьінформацію про нього макрофагам(1-йсигнал), які захоплюють цей матеріал, переробляють і
посилають інформаціюВ-лімфоцитам(2-йсигнал). Останні у відповідь розмножуються і
створюють клони плазматичних клітин, здатних синтезувати комплементарні антитіла і
розвиваються реакції відторгнення або ж виникають відносини толерантності (терпимості).
Процес відтворення організмів, що включає стадії гаметогенезу, осіменіння, запліднення,
вагітності, родів та післяродову, супроводжується появою в організмі сторонніх антигенів і
розвитком відповідних імунних реакцій.
Гаметогенез завершується утворенням нових статевих клітин, що відрізняються своїм
антигенним складом від сім’яників (спермії) та яєчників (яйцеклітини), але вони не викликають
щодо себе імунних реакцій. Чому?
– Спермії вкрилися у придатку ліпопротеїдним шаром;гемато-тестикулярнийбар’єр сім’яників не
допускає проникнення сперміїв у кров’яне русло, під час еякуляції спермії вкриваються
плазмою. При пошкодженні цього бар’єра виникає аутоімунний орхіт.
– У яйцеклітини в яєчнику формується прозора оболонка, вона огортається шаром фолікулярних
клітин у фолікулі, омивається фолікулярною рідиною і відгороджується від крові ендотелієм
капілярів. При порушенні цих захисних структур виникає автоімунний оварит.
32.
Після запліднення формується оболонка запліднення.Статеві органи добре васкуляризовані, під впливом імунних подразників змінюється
проникність судин, надходження імунокомпетентних клітин, напрямок імунних реакцій.
Статевий цикл, роди, післяродовий період супроводжуються змінами епітелію ендометрію,
відбувається деструкція, первинна епітелізація, проліферація та регенерація, скеровані на
обмеження проникнення сперміїв у кров’яне русло, обмеження виходу макрофагів чи,
навпаки, сприяння цьому процесу. Введені в геніталії самки спермії під час осіменіння
оточені плазмою та секретом матки, що обмежує контакт антигену з імунокомпетентними
клітинами. Секрету матки властивий супресивний ефект. Супресивна властивість притамана
і спермі. Осіменіння, отже, може завершитися заплідненням, або ж воно може бути
ненавмисною імунізацією (в антигенному плані сперма є чужорідною для самки), що
викликає утворення антитіл.
В геніталіях самки відбувається капацитація сперміїв та імунна селекція в результаті чого
настає супресія одних та толерантність до інших.
Ліпопротеїдний покрив сперміїв маскує їх антигени, проте капацитація руйнує цей покрив.
33.
Є ще один фактор імунного захисту – лімфоцити заряджені негативно, а наявні у прозорій оболонцігіалуронова та сіалова кислоти володіють електровідштовхувальною властивістю щодо лімфоцитів.
Після розриву прозорої оболонки зростає імуногенність ембріона. Він тепер уже не захищений
прозорою оболонкою. Роль імунологічного фільтра тепер виконує трофобласт, що вступає в
безпосередній контакт зі слизовою оболонкою матки. Він виділяє супресорний фактор, що захищає
ембріон від відторгнення. Розміщені на поверхні трофобласта антигени утворюють комплекси з
антитілами, по яких материнський організм розпізнає, чи сумісний ембріон з материнським
організмом чи ні. Згадані комплексиантиген-антитілозахищають ембріон або ж активують
цитотоксичну дію лімфоцитів. В результаті ембріон зберігається або ж гине.
Вирішальну роль у подальшому збереженні вагітності відіграютьплодово-мате-ринські відносини.
Плацента і плодові води виконують тепер роль масивного тканинного бар’єра між плодом і
материнським організмом.
Амніотична та алантоїсна рідини відзначаються антигенною подібністю з ембріоном .Під час
вагітності зростає продукція Т-лімфоцитів(головно супресорів), що пригнічують синтезВклітинамиантитіл, збільшується в крові вміст кортикостероїдів – головних інгібіторів синтезу
антитіл і встановлюється рівновага між факторами стимулювання та пригнічення синтезу антитіл.
Координуючий вплив на динаміку цих процесів проявляють гормони: естрадіол стимулює імунну
систему ендометрію, продукцію тут імуноглобулінів, синтез рецепторів окситоцину, а прогестерон
проявляє протилежну дію.
Потрібно мати на увазі, що ембріони також продукують гормони, які сигналізують про настання
вагітності і проявляють вплив на перебіг ембріогенезу. Саме ці гормони пригнічують синтез у матці
простагландинів F2α – природного ендогенного лютеолізину.
34.
Продукований жовтим тілом прогестерон підсилює імуносупресивну дію ендометрію іпригнічує продукцію інтерлейкіну.
Таким чином, плацента – це не лише додаткове джерело прогестерону, це також орган
імунного захисту плода. Вона запускає проліферацію Т- і В-лімфоцитів,підтримує напругу
місцевого імунітету. Певну роль також відіграє імуносупресивний вплив так званих білків
зони вагітності, що виявляються в крові вагітних тварин.
Таким чином, організм самки виявляється імунокомпетентним стосовно змін, що виникають у
ньому в зв’язку з розвитком вагітності.
З наближенням родів порушується імунологічна рівновага і замість толерантності
розвиваються реакції відторгнення.
Зміна фізіологічного стану самки, неповноцінна годівля, незадовільні умови догляду та
утримання, розвиток патологічних процесів порушує імунну рівновагу в організмі і вагітність
ускладнюється чи переривається.
Зміни в статевій системі. Під впливом дедалі наростаючого розвитку плода помітні зміни
відбуваються у статевій системі, зокрема в матці. Матка поступово збільшується в об’ємі. Ріст
матки відбувається за рахунок гіпертрофії (збільшення) кожного зокрема м’язового волокна
як у довжину, так і в товщину. Сполучна тканина, закладена між шарами м’язових волокон,
розм’якшується, стає еластичною, через що більш рухомими стають м’язові волокна.
Судини матки під час вагітності здовжуються і розширюються, сприяючи цим постачанню до
плаценти материнської крові. Маткові артерії збільшують свій просвіт. Незважаючи на
збільшення м’язових волокон, розм’якшення сполучної тканини і припливу великої кількості
крові, товщина стінок матки поступово зменшується. Це відбувається за рахунок сильного
збільшення її поверхні через розтягнення.
Слизова оболонка матки під час вагітності від великого припливу крові трохи набухає, але
складки її згладжуються внаслідок збільшення об’єму матки. Миготливий епітелій слизової
оболонки зазнає жирового переродження, беручи участь в утворенні маткового молока, яке,
як відомо, є поживним середовищем для зародка. У матковому молоці є білок, жири, невелика
кількість солей, лейкоцити та різні епітеліальні клітини, що відокремилися від слизової
оболонки матки, а також ферменти і біологічно активні речовини.
На слизовій оболонці матки у кобили й свині збільшуються заглибини крипт, у які входять
ворсинки судинної оболонки плода. У корів на слизовій оболонці розростаються карункули,
розміщені в кожному розі в чотири ряди. На них утворюються крипти, куди також входять
ворсинки хоріона плода.
35.
Укорів, овець ікіз вагітна матка опускається в черевну порожнину і в кінці вагітностірозміщується на нижній стінці живота, доходячи до грудної кістки, і звичайно випинає
праву здухвинну ділянку. В тих випадках, коли є плід і в лівому розі, він розміщується
під рубцем. У міру росту плода шийка матки і передній край маткових зв’язок теж
просуваються далі вперед. При двійнях плоди містяться в обох рогах, але бувають
випадки, коли обидва плоди лежать в одному розі. В кінці вагітності товщина стінок
матки доходить у дрібних жуйних до 1 мм. Це слід брати до уваги під час акушерської
допомоги, бо небезпека розриву матки буває дуже велика. Треба відзначити, що
яєчники в період вагітності теж трохи змінюють своє положення під впливом опускання
матки.
Широкі маткові зв’язки в період вагітності товщають за рахунок гіпертрофії м’язових
волокон і набухання сполучної тканини. До стінок піхви припливає багато крові, через
що тканини поступово набрякають, і слизова оболонка червоніє. Внаслідок опускання
матки вниз черевної порожнини піхва здовжується і доходить у великих тварин до краю
таза. Слизова оболонка піхви покрита густим слизом. Набряклість тканин піхви
поширюється і на вульву, через що складки на ній згладжуються. Набряклість вульви у
корів в останній стадії вагітності буває така велика, що статеві губи розкриваються.