











































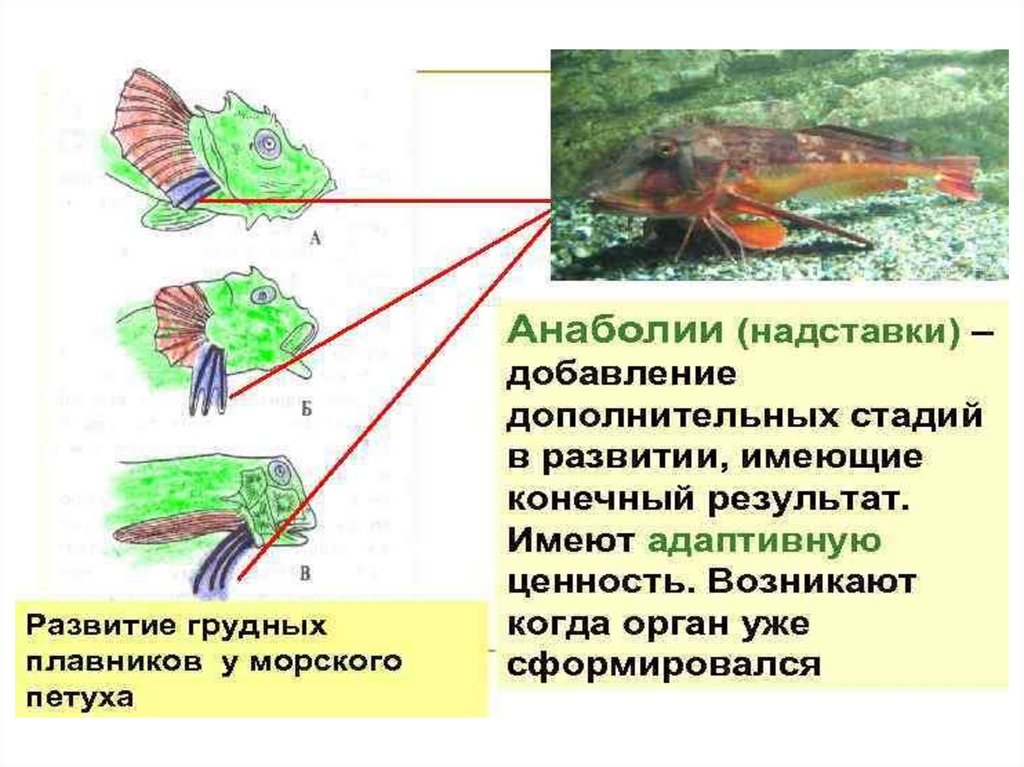





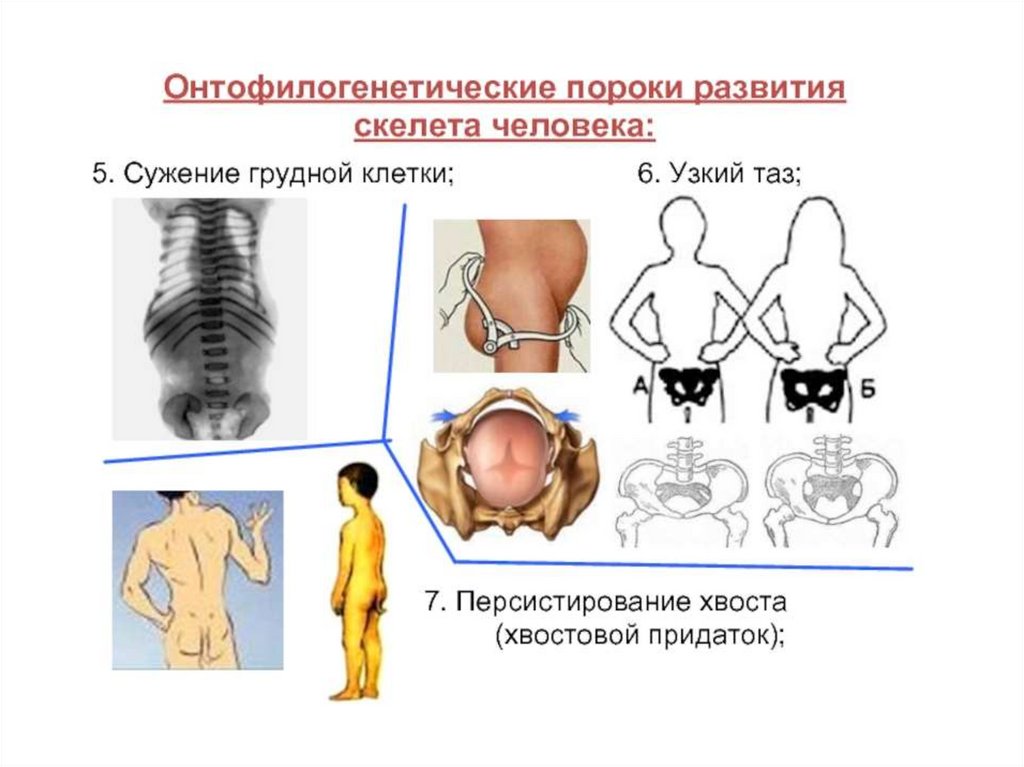
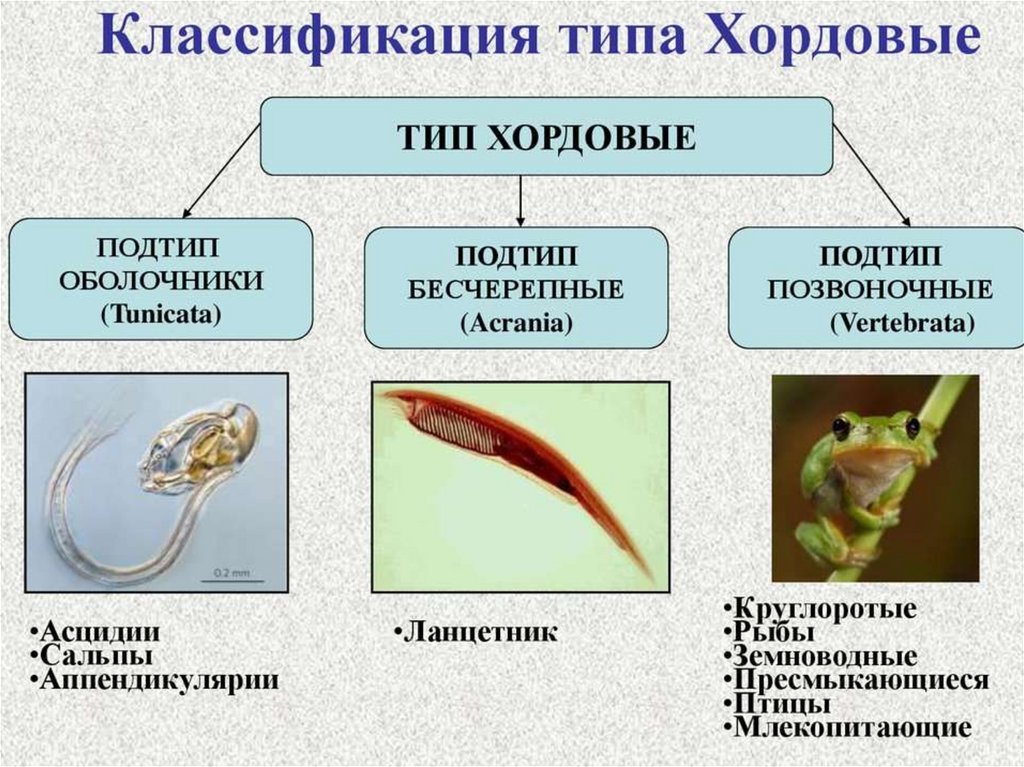





































 Биология
БиологияПохожие презентации:
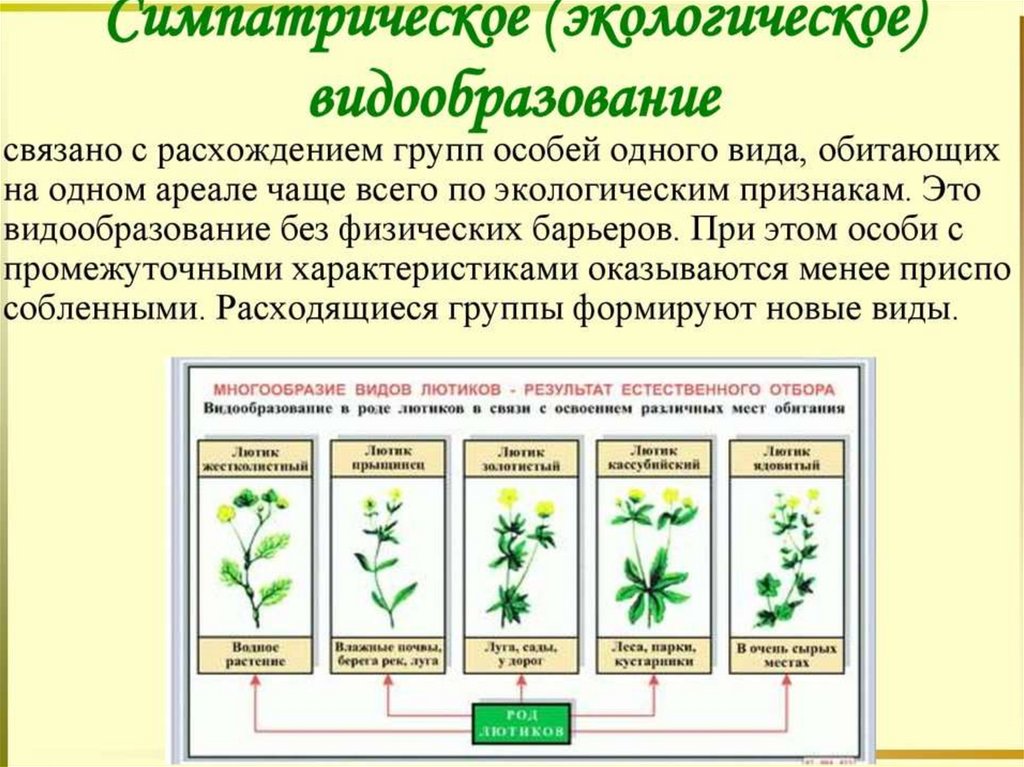
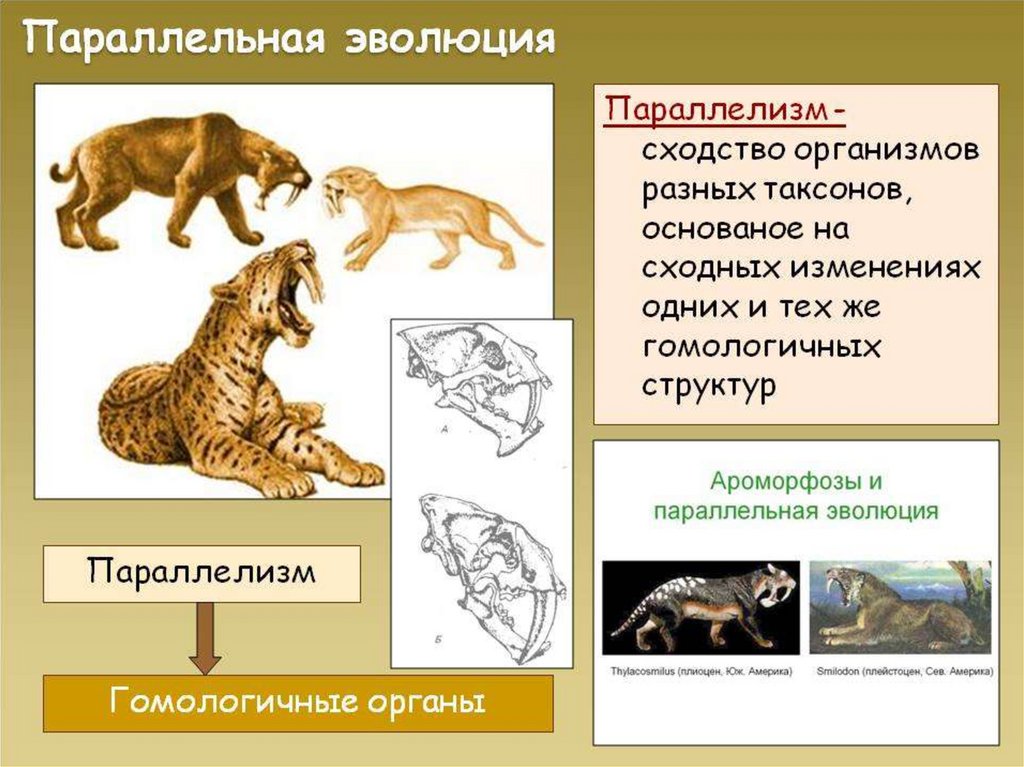
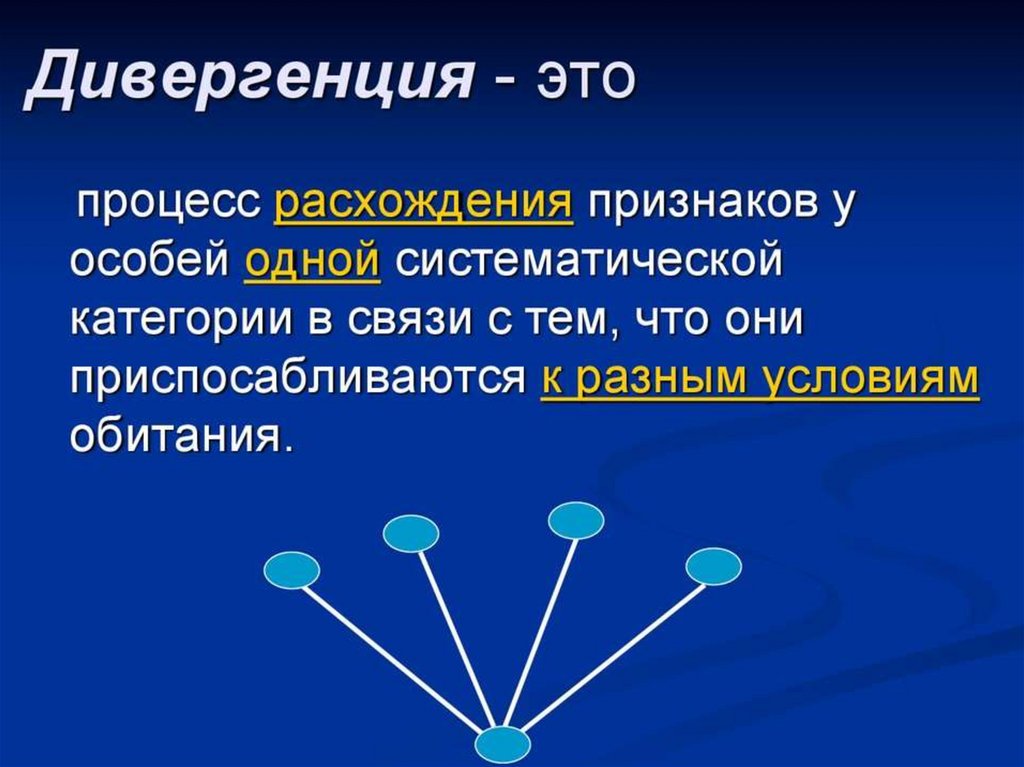
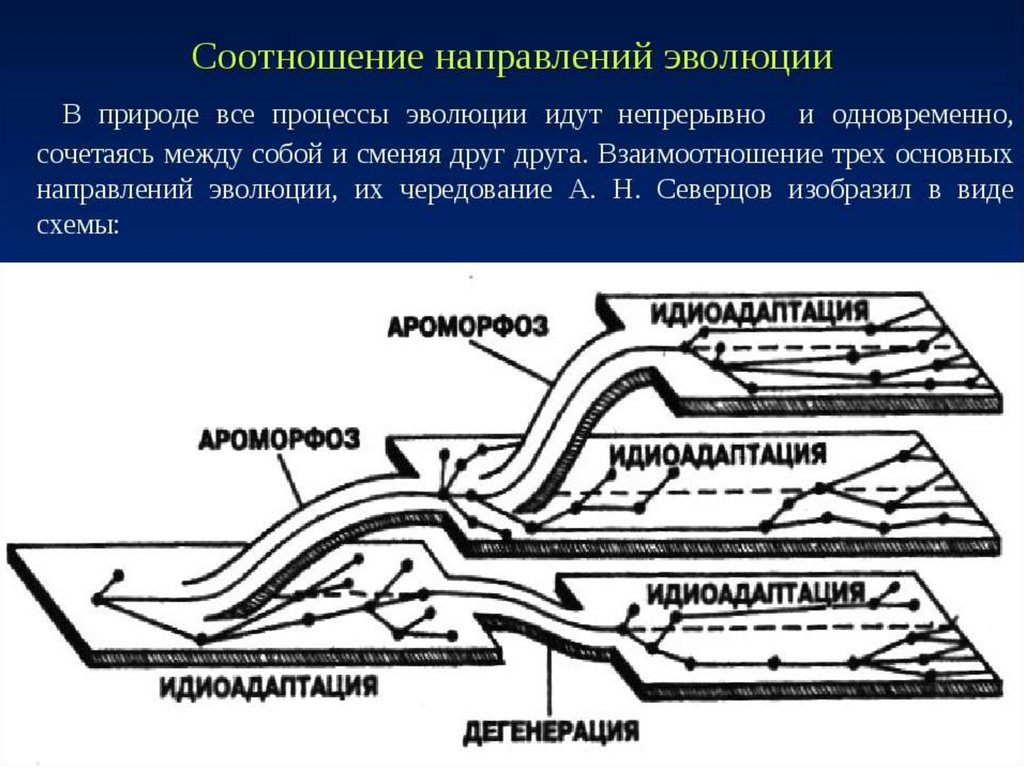
")









Эволюция. Филогенез
1. Эволюция. Филогенез
Доцент кафедры биологиимедицинской Жукова А.А.
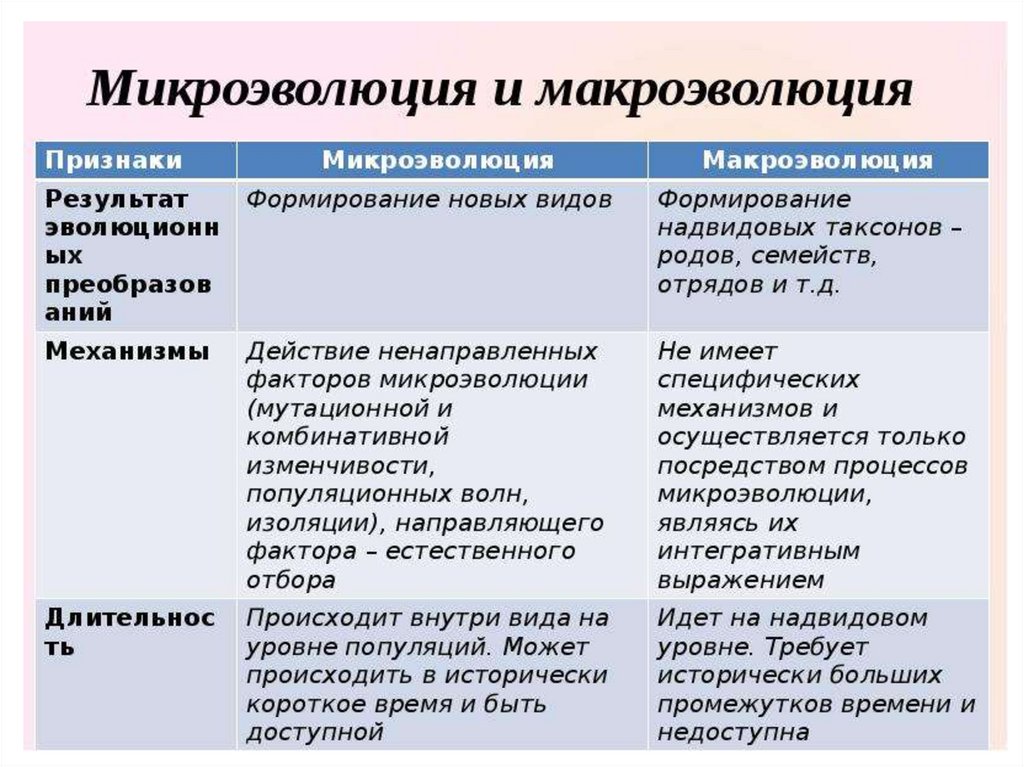
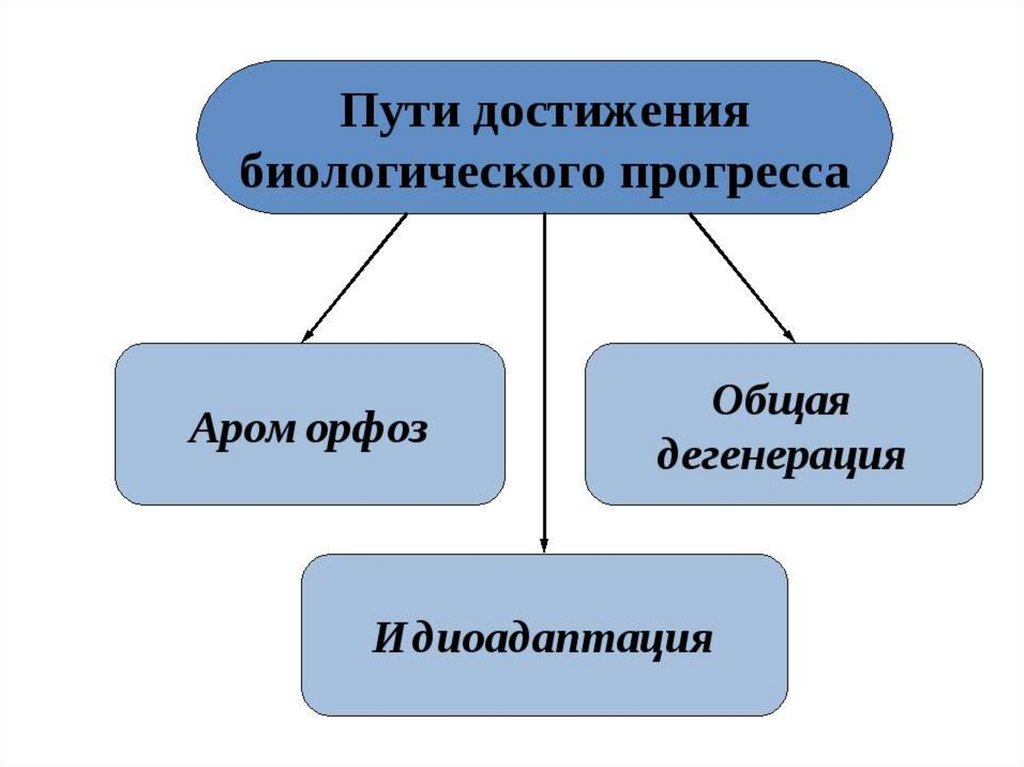
2.
3.
4.
5.
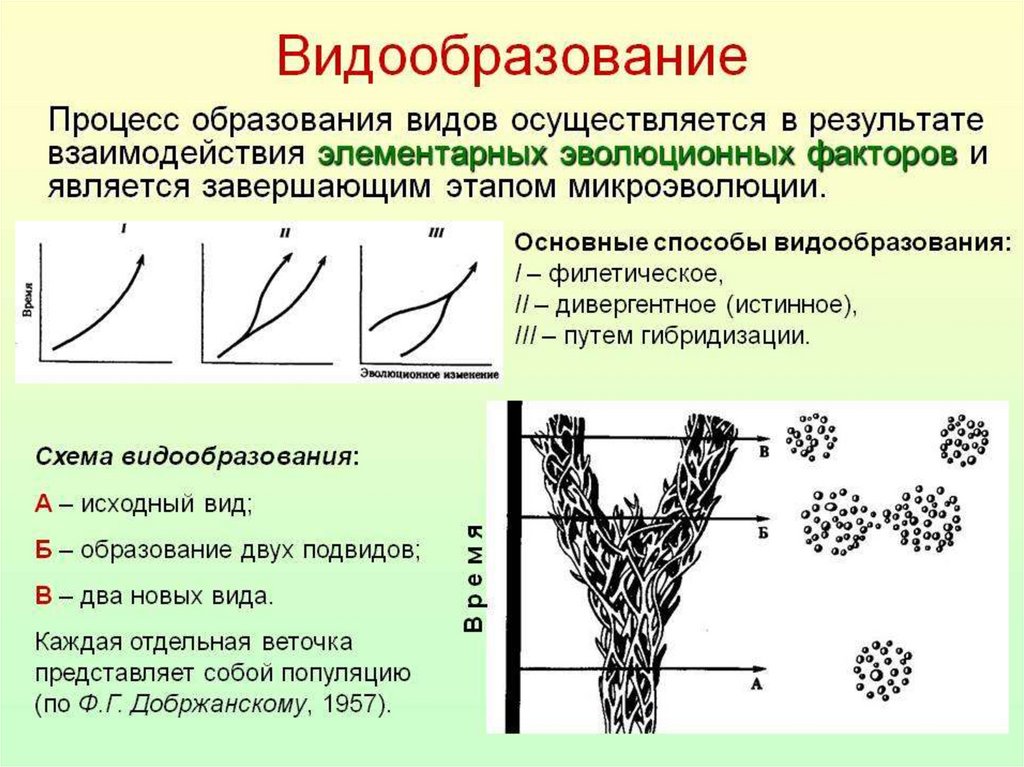
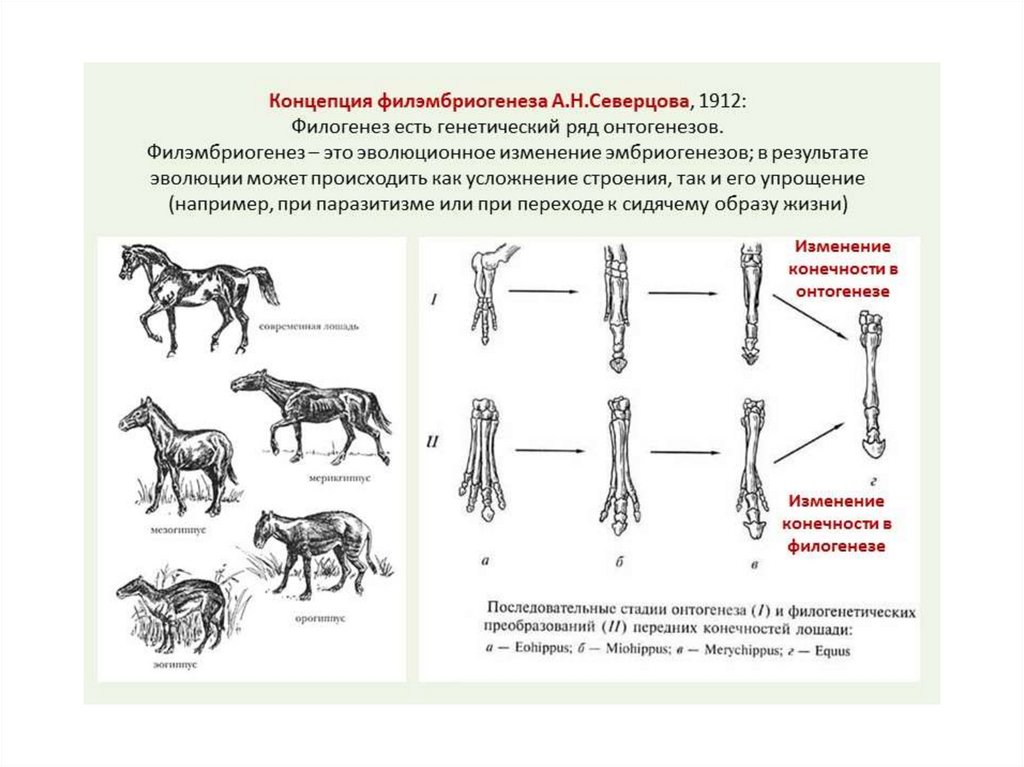
6.
7.
8.
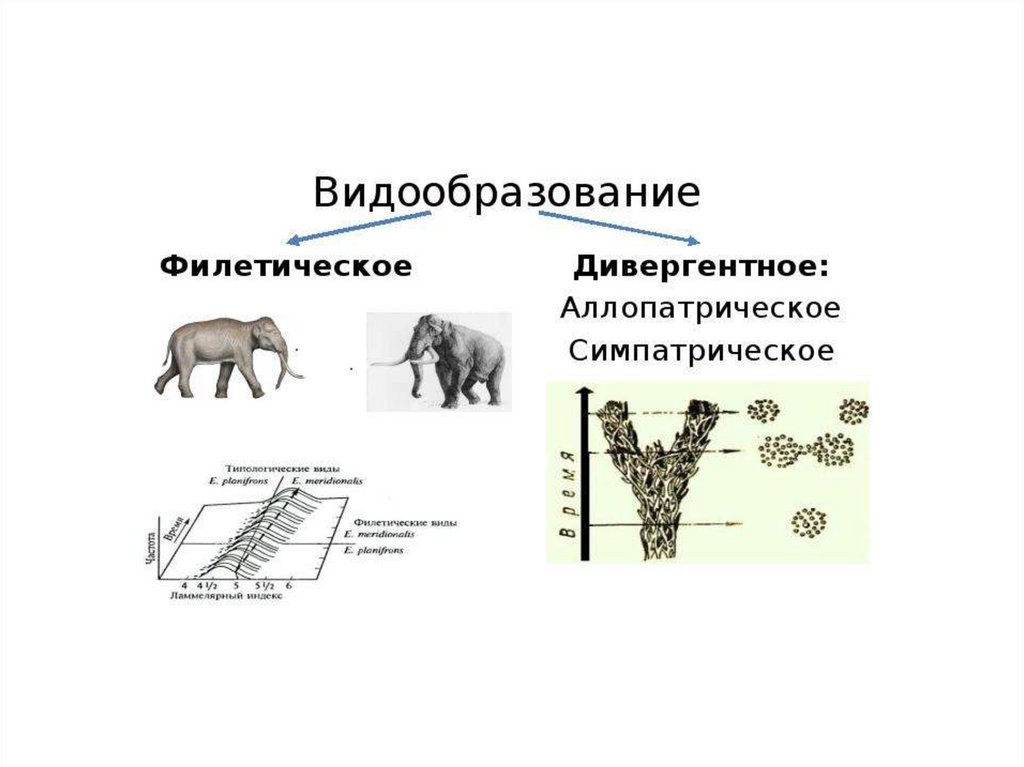
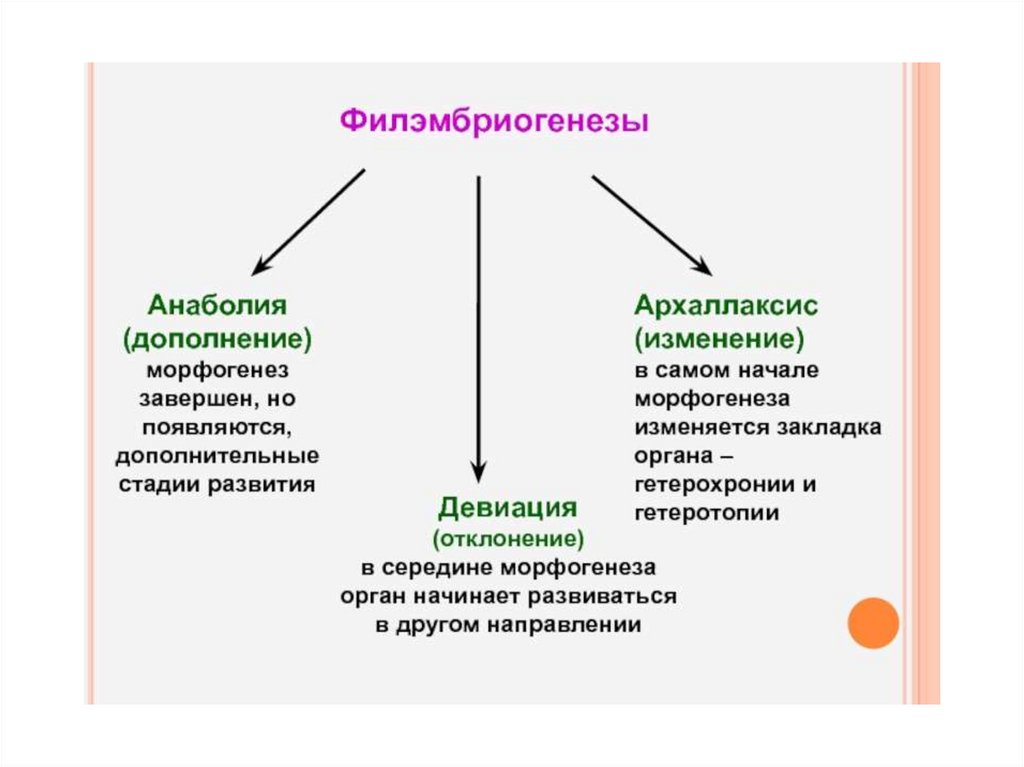
9.
10.
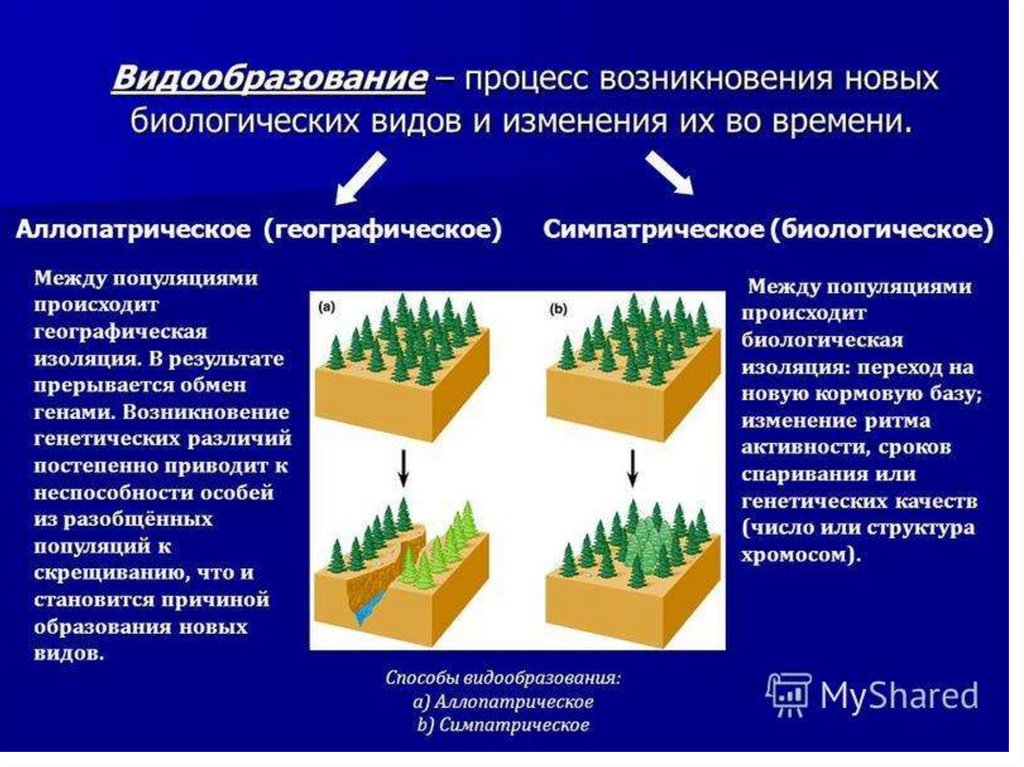
11.
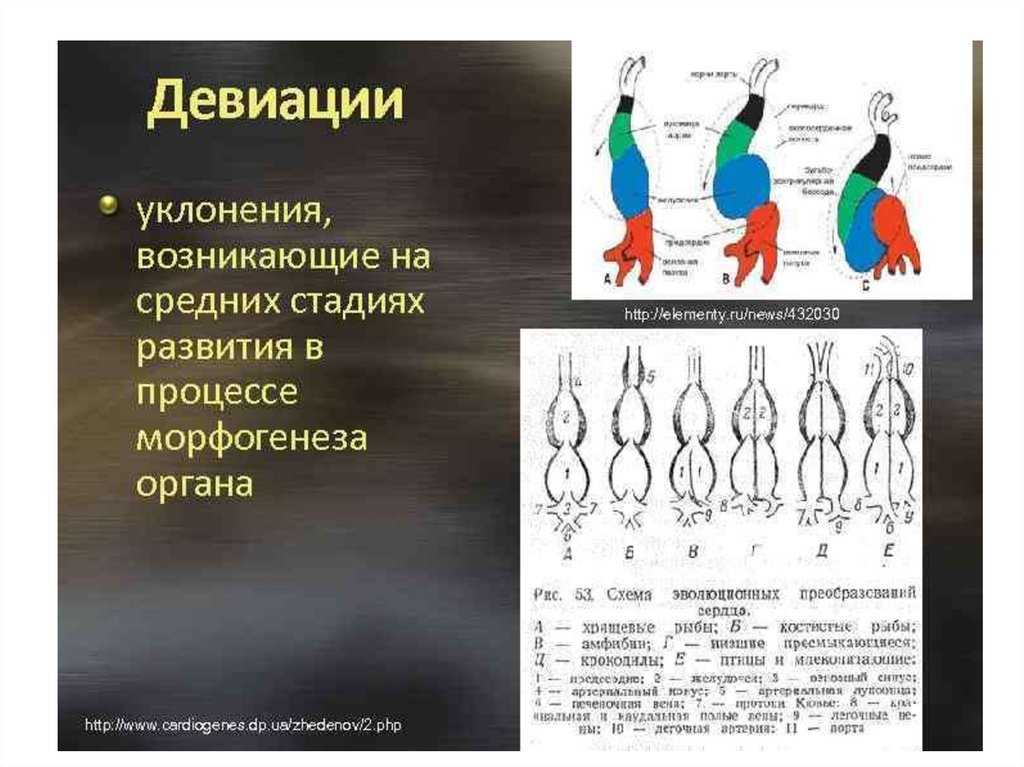
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60.
61.
62.
63.
64.
65.
66.
67.
68.
69.
70.
71.
72.
73.
74.
75.
76.
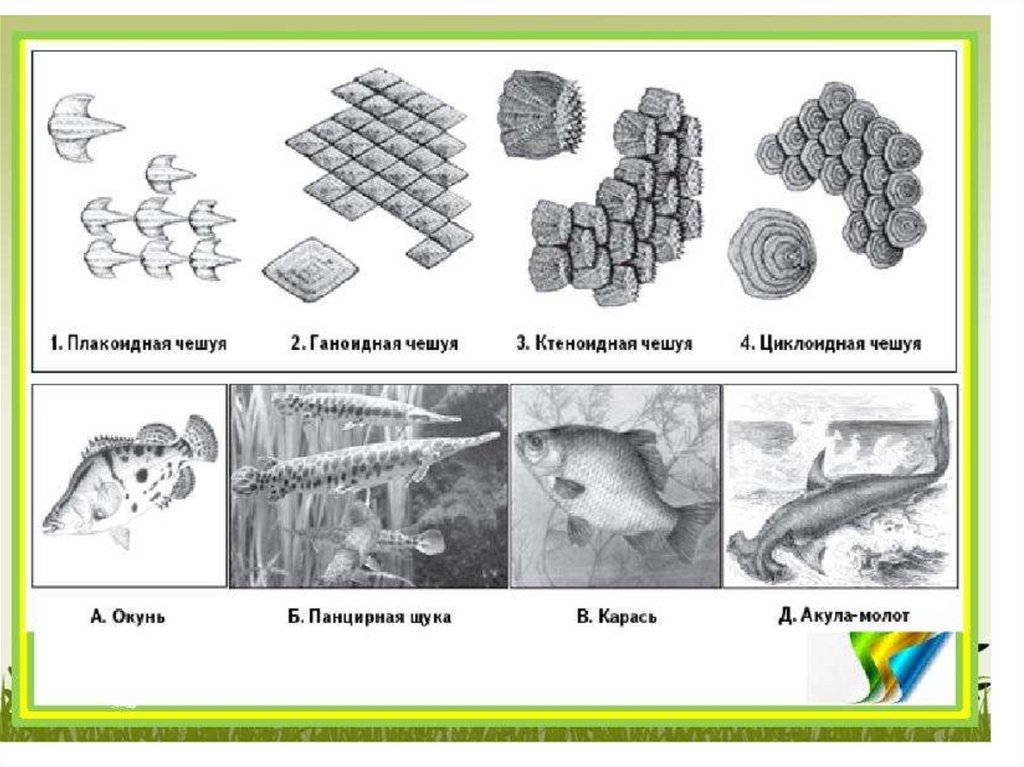
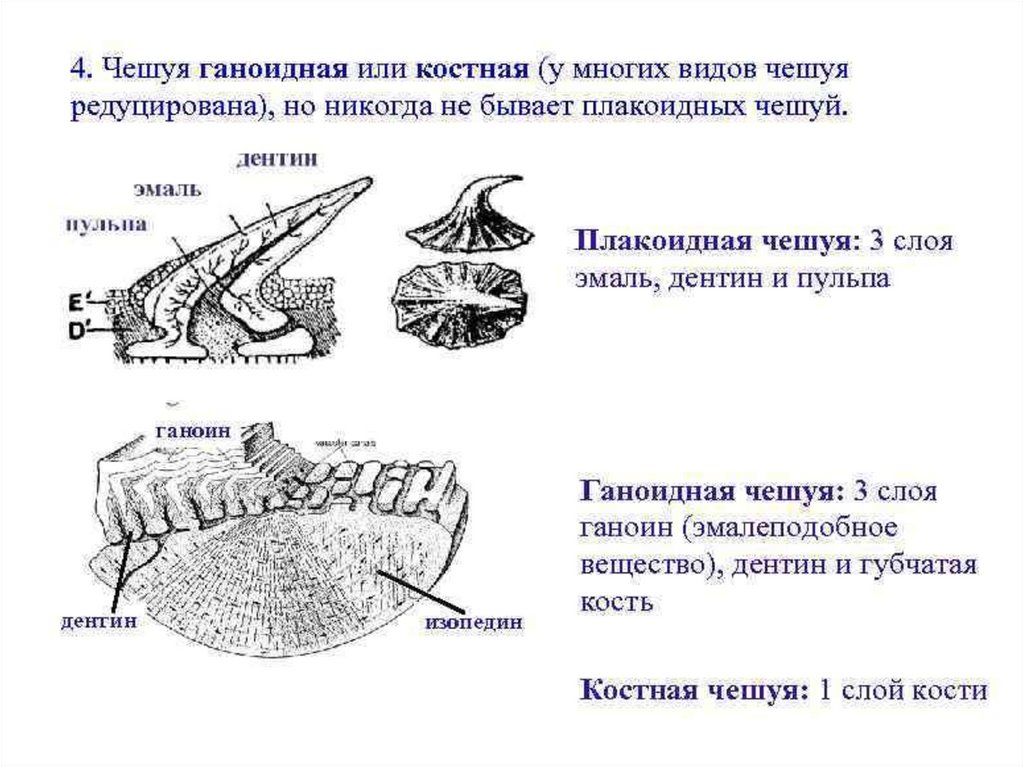
Плакоидная чешуя рыбы ( фото вверху) характерна для современных иископаемых хрящевых рыб – а это акулы и скаты. Каждая такая чешуйка
имеет пластинку и шип, сидящий на ней, остриё которого выходит наружу
через эпидермис. В этой чешуйке основой является дентин. Сам шип покрыт
ещё более твёрдой эмалью. Плакоидная чешуйка внутри имеет полость,
которая заполнена мякотью – пульпой, она имеет кровеносные сосуды и
нервные окончания.
Ганоидная чешуя рыбы имеет вид ромбической пластинки и чешуйки
соединены друг с другом, образуя на рыбе плотный панцирь. Каждая такая
чешуйка состоит из очень твёрдого вещества – верхняя часть из ганоина, а
нижняя из кости. Такой вид чешуи имеют большое количество ископаемой
рыбы, а также верхние части в хвостовом плавнике у современных осетровых
рыб.
Ктеноидная чешуя рыбы тоже встречается у костистых рыб и не имеет слоя
ганоина, на тыльной стороне у неё шипы. Обычно чешуя у этих рыб
расположена черепицообразно, и каждая чешуйка прикрыта спереди и с
обеих сторон такими же чешуйками. Получается так, что задний конец
чешуйки выходит наружу, но и снизу он подостлан другой чешуйкой и такой
вид покрова сохраняет гибкость и подвижность рыбы. Годовые кольца на
чешуе рыбы позволяют определить её возраст.
Циклоидная чешуя рыбы. Циклоидная чешуя рыбы встречается у костистых
рыб и не имеет слоя ганоина. Циклоидная чешуя имеет округлую шейку с
гладкой поверхностью.
77.
78.
79.
80.
81.
82.
83.
84.
85.
86.
87.
88.
89.
90.
91.
92.
93.
94.
95.
96.
Уникальная особенность организации хордовых - филогенетическая,эмбриогенетическая, а также функциональная связь пищеварительной и
дыхательной систем. Действительно, только у хордовых дыхательная система
развивается на базе пищеварительной и на первых этапах эволюции
функционирует совместно с ней.
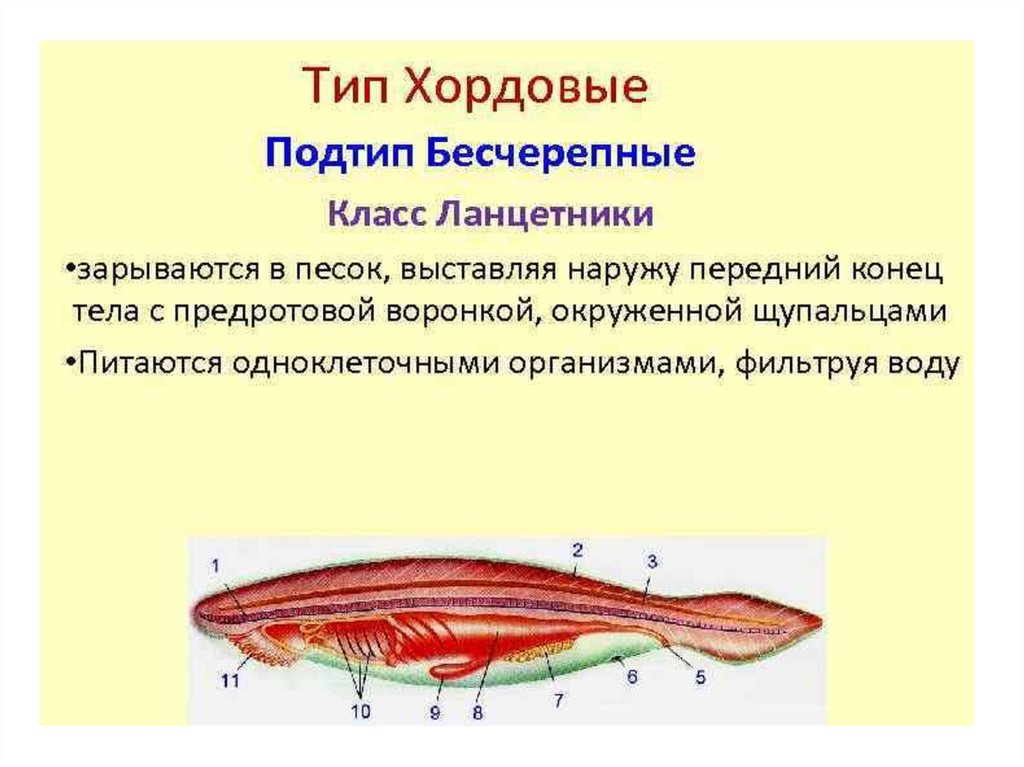
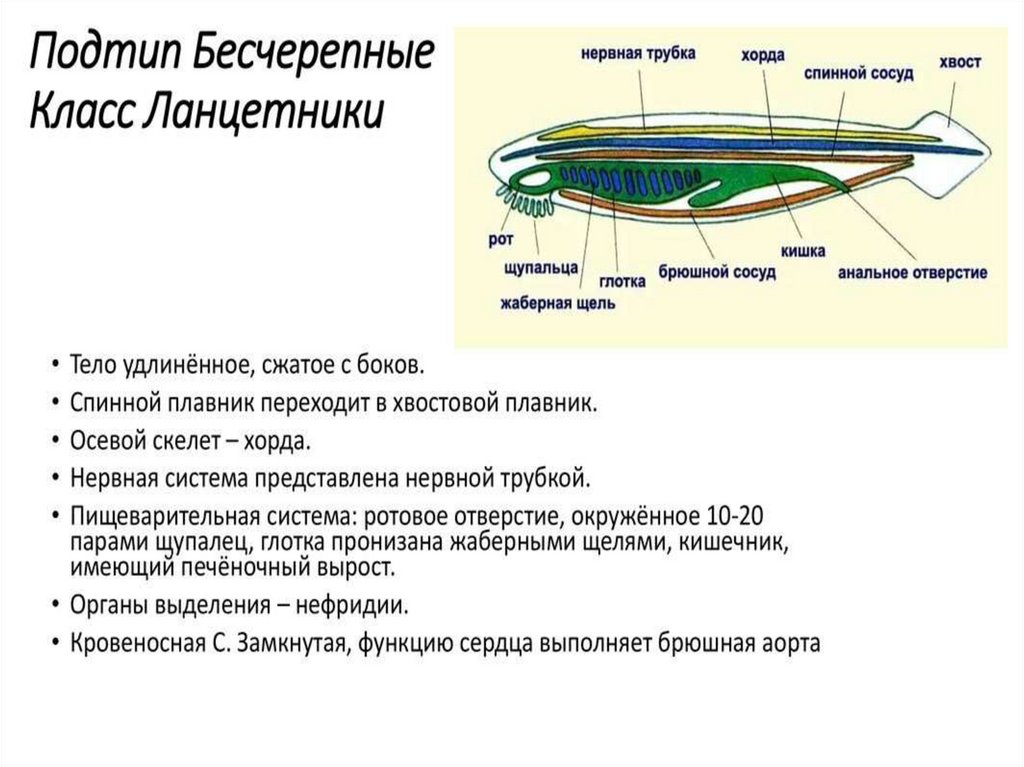
Так, у ланцетника, сохранившего в значительной степени черты организации
предковых форм, специализированных органов дыхания нет, а дыхательную
функцию выполняет глотка - передняя часть пищеварительной трубки,
пронизанная сквозными отверстиями - жаберными щелями, главной функцией
которых является фильтрация воды.
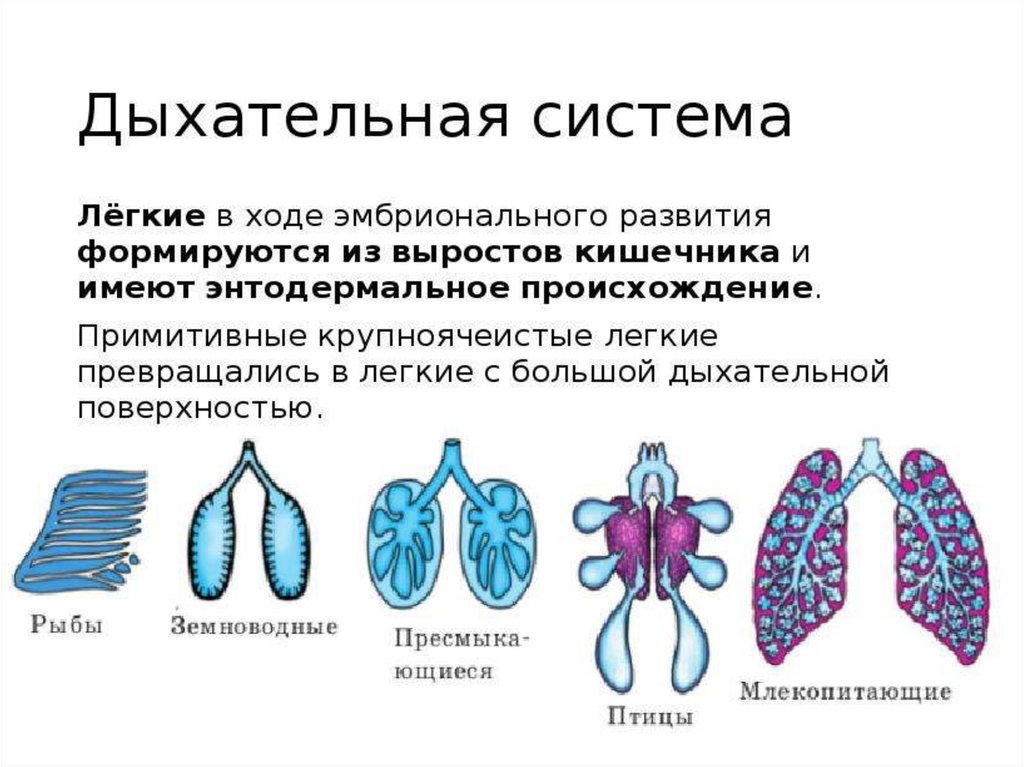
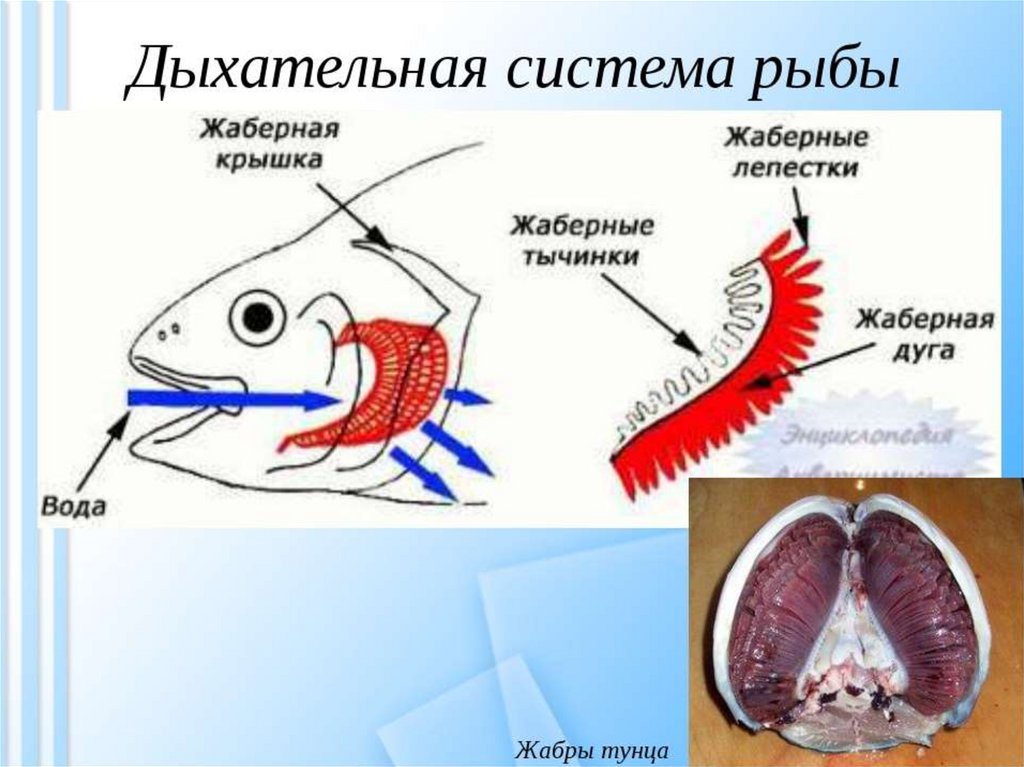
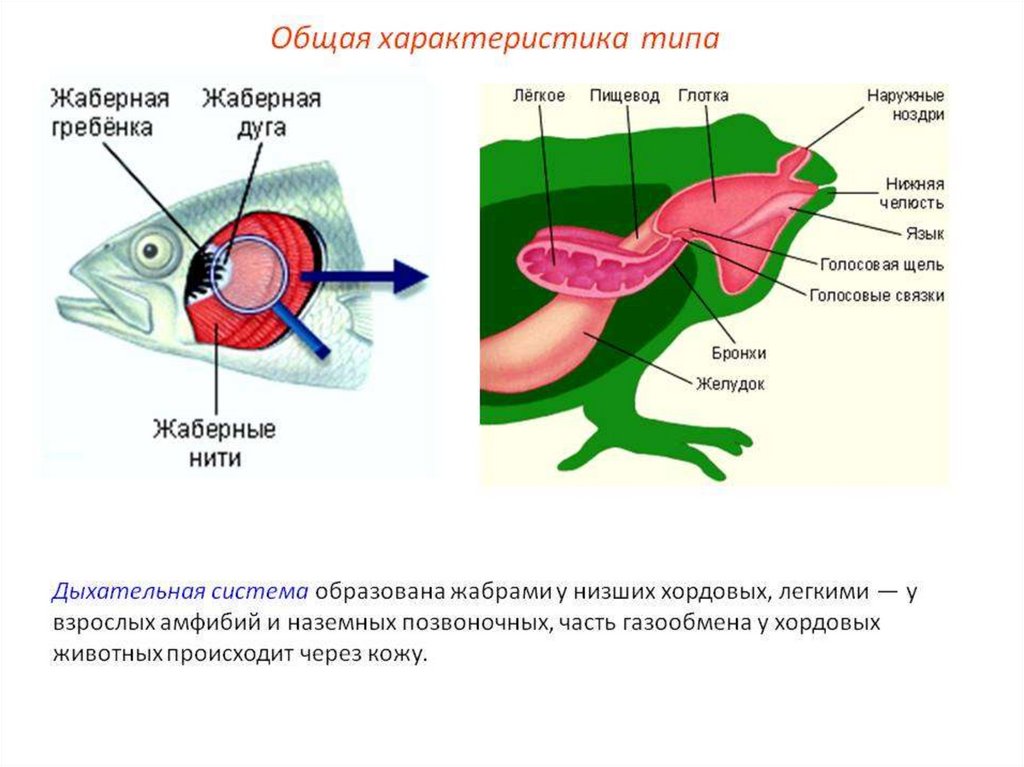
Органами дыхания рыб являются - жабры. Жабры- наиболее ранние
специализированные органы дыхания, появляющиеся среди хордовых впервые
у рыб. Они представляют собой тонкие складки слизистой оболочки глотки,
лежащие на жаберных дугах, снабжаемые венозной кровью через жаберные
артерии и распадающиеся здесь на капилляры. Позади последней жаберной
дуги у кистеперых рыб за счет выпячивания вентральной стороны глотки
формируется парное образование - плавательный пузырь, который выполняет
в первую очередь гидростатические функции, уравновешивая тело рыбы в
толще воды. Связь между ним и глоткой, имеющаяся в эмбриогенезе, не
прерывается и позже, поэтому воздух при попадании в глотку может
беспрепятственно проходить в плавательный пузырь. Эта
морфофизиологическая особенность при изменении условий существования
кистеперых рыб стала предпосылкой преобразования плавательного пузыря в
легкие.
97.
98.
99.
У земноводных, связанных по происхождению с кистеперыми рыбами, вличиночном состоянии функционируют жабры, а во взрослом - органами
дыхания становятся легкие. В связи с отсутствием грудной клетки и
диафрагмы воздух в них попадает из ротовой полости за счет глотательных
движений, осуществляющихся подбородочно-подъязычной мышцей.
Материал жаберных дуг, следующих за подъязычной дугой, частично
редуцируясь, входит в состав хрящей гортани, которая, появляясь впервые у
земноводных, является первым органом, относящимся к нижним
дыхательным путям. Легкие начинаются непосредственно от гортани. Они
крупноячеисты и имеют относительно малую дыхательную поверхность, в
связи с чем газообмен в большей степени осуществляется через кожные
покровы.
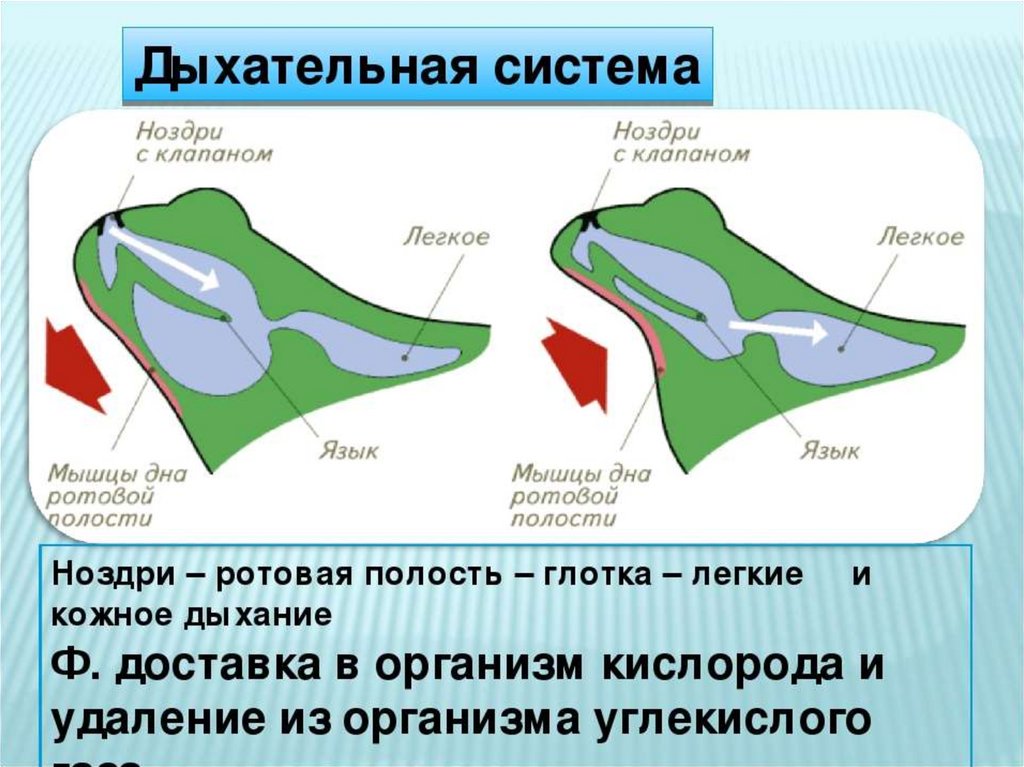
У Земноводных есть внутренние ноздри (хоаны), которые проводят воздух в
лёгкие. Горло лягушки несколько раз в секунду оттягивается вниз, благодаря
чему в ротовой полости создаётся разреженное пространство. Тогда лягушка
совершает глотательное движение, и воздух проникает через ноздри в
ротовую полость, а оттуда в лёгкие.
100.
101.
102.
103.
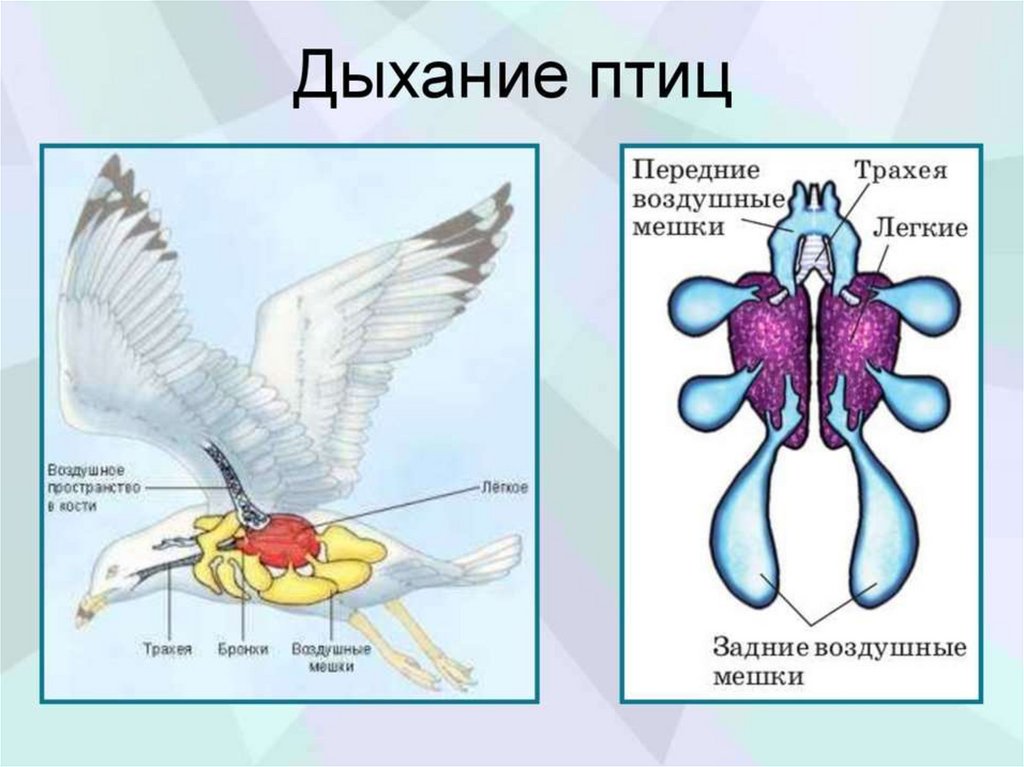
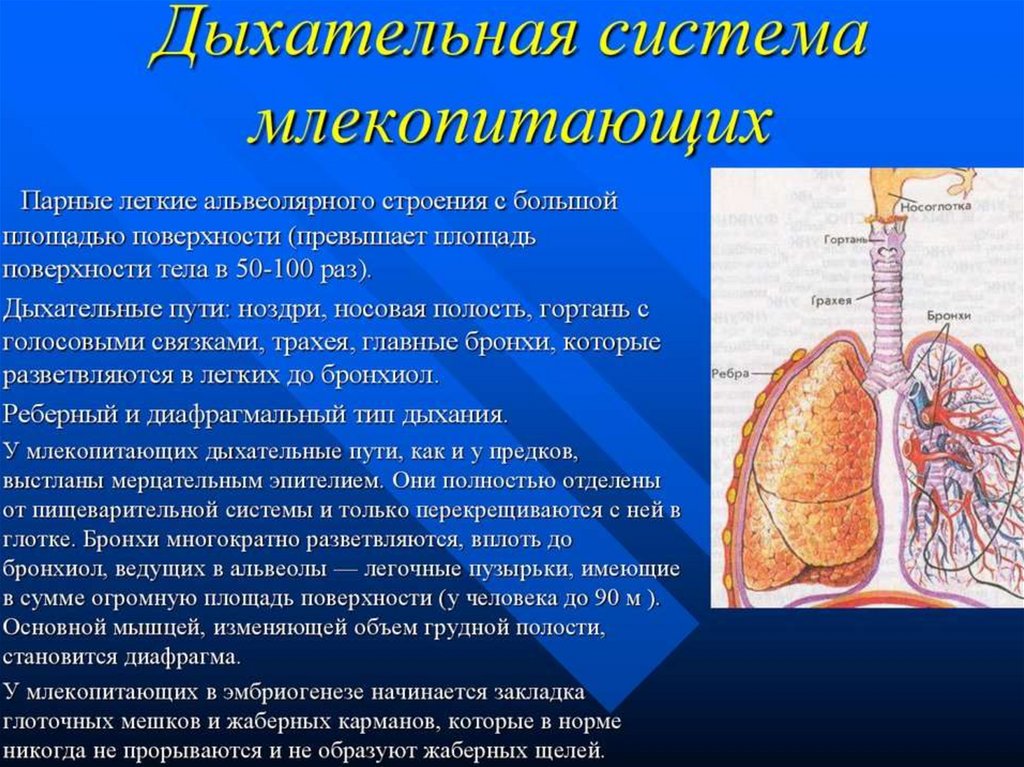
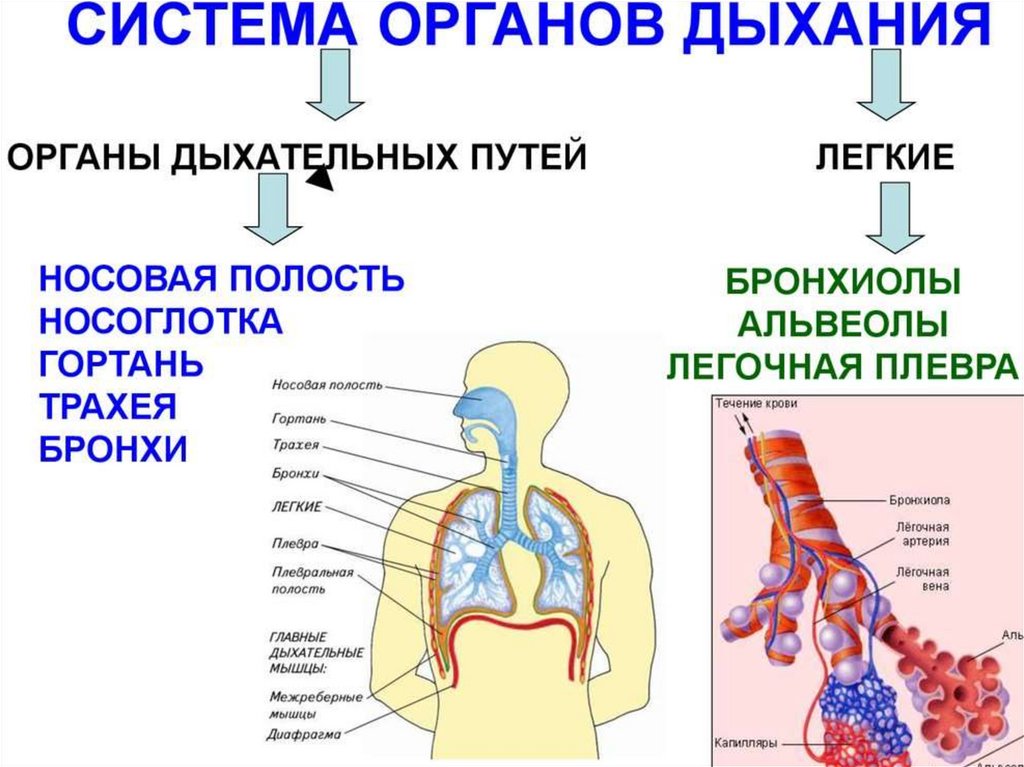
Дыхательная система образована дыхательными путями и лёгкими своздушными мешками.
Воздух проходит через ноздри в дыхательные пути: носовую
полость, гортань, трахею, бронхи — и далее в лёгкие и воздушные мешки.
Газообмен происходит в ветвящихся бронхиолах лёгких.
Лёгкие птиц отличаются сложным губчатым строением.
Бронхи, заходя в лёгкие, многократно ветвятся и заканчиваются в полостях, а
некоторые, выйдя из лёгких, расширяются, образуя воздушные мешки.
Функции воздушных мешков:
участвуют в дыхании при полёте (двойное дыхание);
обеспечивают охлаждение организма в полёте;
уменьшают плотность (и вес) тела.
Двойное дыхание
Когда птица находится в состоянии покоя (не в полёте), дыхание происходит
без участия воздушных мешков.
В полёте воздух, насыщенный кислородом, проходит через лёгкие птиц 2 раза
— при вдохе и при выдохе (при выдохе он перемещается в лёгкие из
воздушных мешков). Такой механизм дыхания птиц называется двойным
дыханием.
104.
105.
106.
107.
108.
109.
110.
Кровеносная система всех хордовых замкнутая и состоит из двух основных артериальныхсосудов: брюшной и спинной аорт. По брюшной аорте венозная кровь продвигается
кпереди, обогащается кислородом в органах дыхания, а по спинной - кзади. Из спинной
аорты кровь через систему капилляров возвращается по венам в брюшную аорту. Брюшная
аорта или ее часть, периодически сокращаясь, проталкивает кровь по сосудам
У ланцетника кровеносная система наиболее проста. Круг кровообращения один. По
брюшной аорте венозная кровь поступает в приносящие жаберные артерии, число которых
соответствует числу межжаберных перегородок (до 150 пар), где и обогащается кислородом.
По выносящим жаберным артериям кровь поступает в корни спинной аорты, расположенные
симметрично с двух сторон тела. Они продолжаются как вперед, неся артериальную кровь к
переднему концу тела, так и назад. Передние ветви этих двух сосудов называются сонными
артериями. На уровне заднего конца глотки задние ветви, срастаясь, образуют спинную
аорту, которая разветвляется на многочисленные артерии, направляющиеся к органам и
распадающиеся на капилляры. После тканевого газообмена кровь поступает в
парные передние или задние кардинальные вены, расположенные симметрично.
Передняя и задняя кардинальные вены с каждой стороны впадают в кювьеров проток. Оба
кювьеровых протока впадают с двух сторон в брюшную аорту. От стенок пищеварительной
системы венозная кровь оттекает по воротной вене печени в печеночный вырост, где
формируется система капилляров. Затем капилляры вновь собираются в венозный сосуд печеночную вену, по которой кровь поступает в брюшную аорту. Таким образом, несмотря
на простоту кровеносной системы в целом, уже у ланцетника имеются основные
магистральные артерии, характерные для позвоночных, в том числе для человека: это
брюшная аорта, преобразующаяся позже в сердце, восходящую часть дуги аорты и корень
легочной артерии; спинная аорта, становящаяся позже собственно аортой, и сонные
артерии. Основные вены, имеющиеся у ланцетника, также сохраняются у более
высокоорганизованных животных. Так, передние кардинальные вены станут
позже яремными венами, правый кювьеров проток преобразуется в верхнюю полую
вену, а левый, сильно редуцировавшись, - в коронарный синус сердца. Для того чтобы
111.
Активный образ жизни рыб предполагает более интенсивный метаболизм. В связи с этим на фоне олигомеризацииих артериальных жаберных дуг, в конечном счете, до четырех пар в них отмечается высокая степень
дифференцировки: жаберные сосуды распадаются на капилляры, пронизывающие жаберные лепестки. В процессе
интенсификации сократительной функции брюшной аорты часть ее преобразовалась в двухкамерное
сердце, состоящее из предсердия и желудочка и располагающееся под нижней челюстью, рядом с жаберным
аппаратом. В остальном кровеносная система рыб соответствует строению ее у ланцетника.
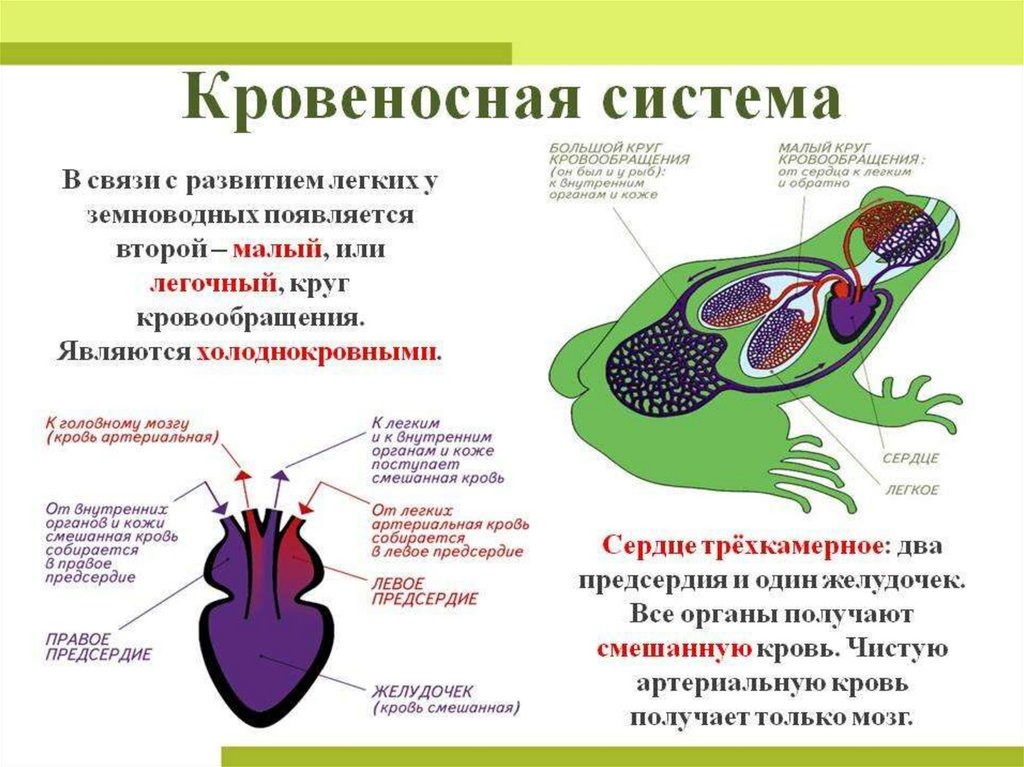
В связи с выходом земноводных на сушу и появлением легочного дыхания у них возникает два круга
кровообращения. Соответственно
этому в строении сердца и артерий появляются приспособления, направленные на разделение артериальной и
венозной крови. Перемещение земноводных в основном за счет парных конечностей, а не хвоста обусловливает
изменения в венозной системе задней части туловища.
Сердце амфибий расположено каудальнее, чем у рыб, рядом с легкими; оно трехкамерное, но, как и у рыб, от
правой половины единственного желудочка начинается единственный сосуд - артериальный
конус, разветвляющийся последовательно на три пары сосудов: кожно-легочные артерии, дуги аорты и сонные
артерии. Как и у всех более высокоорганизованных классов, в правое предсердие впадают вены большого круга,
несущие венозную кровь, в левое - малого с артериальной кровью. При сокращении предсердий в желудочек,
внутренняя стенка которого снабжена большим количеством мышечных перекладин, одновременно попадают обе
порции крови. Полного их смешения из-за своеобразного строения стенки желудочка не происходит, поэтому при
его сокращении первая порция венозной крови поступает в артериальный конус и с помощью спирального
клапана, находящегося там, направляется в кожно-легочные артерии. Кровь из середины желудочка, смешанная,
поступает таким же образом в дуги аорты, а оставшееся небольшое количество артериальной крови, последней
попадающей в артериальный конус, направляется в сонные артерии.
Две дуги аорты, несущие смешанную кровь, огибают сердце и пищевод сзади, образуя спинную аорту,
снабжающую все тело, кроме головы, смешанной кровью. Задние кардинальные вены сильно редуцируются и
собирают кровь только с боковых поверхностей туловища. Функционально их замещает возникшая заново задняя
полая вена, собирающая кровь в основном из задних конечностей. Она располагается рядом со спинной аортой и,
находясь позади печени, вбирает в себя печеночную вену, которая у рыб впадала непосредственно в венозный
синус сердца. Передние кардинальные вены, обеспечивая отток крови от головы, называют теперь яремными
венами, а кювьеровы потоки, в которые они впадают вместе с подключичными венами, - передними полыми
венами.
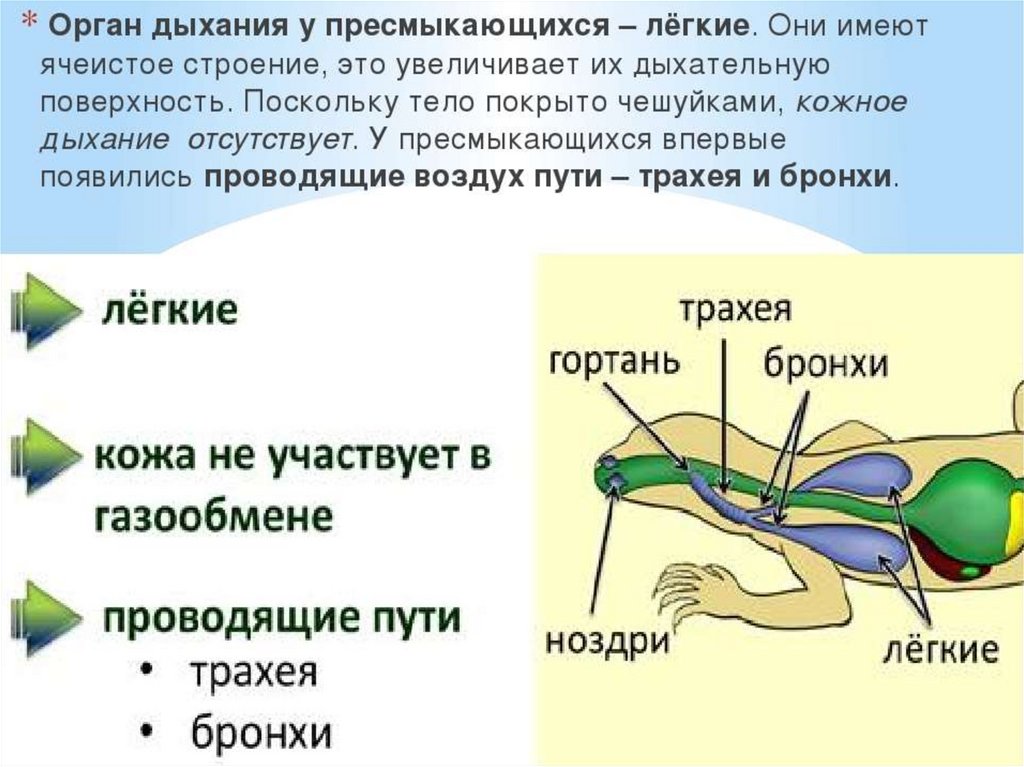
112.
В кровеносной системе пресмыкающихся возникают следующиепрогрессивные изменения: в желудочке их сердца имеется неполная
перегородка, затрудняющая смешение крови, поступающей из
правого и левого предсердий; от сердца отходит не один, а три
сосуда, образовавшихся в результате разделения артериального
ствола. Из левой половины желудочка начинается правая дуга
аорты, несущая артериальную кровь, а из правой - легочная артерия
с венозной кровью. Из середины желудочка, в области неполной
перегородки, начинается левая дуга аорты со смешанной кровью.
Обе дуги аорты, как и у предков, срастаются позади сердца, трахеи
и пищевода в спинную аорту, кровь в которой смешанная, но более
богата кислородом, чем у земноводных, в связи с тем, что до
слияния сосудов только по левой дуге течет смешанная кровь.
Кроме того, сонные и подключичные артерии с обеих сторон берут
начало от правой дуги аорты, в результате чего артериальной
кровью снабжается не только голова, но и передние конечности. В
связи с появлением шеи сердце располагается еще более
каудально, чем у земноводных. Венозная система пресмыкающихся
принципиально не отличается от системы вен земноводных.
113.
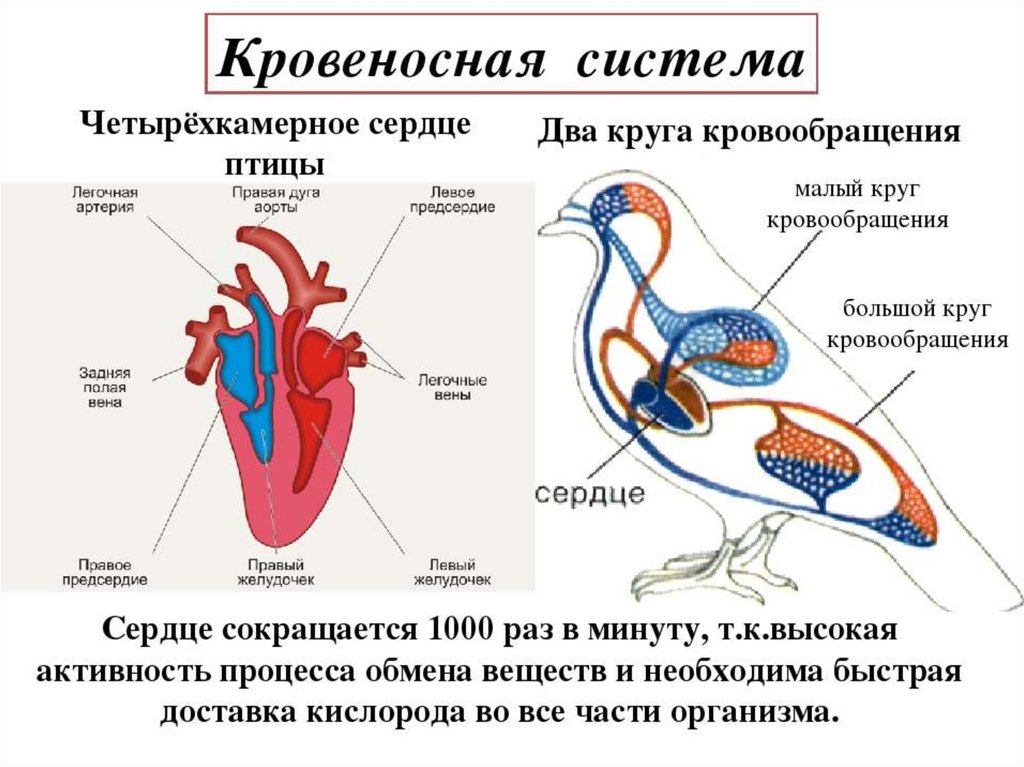
четырёхкамерным сердцем.Сердце состоит из двух предсердий и двух желудочков и полностью разделяет
венозную и артериальную кровь.
Большой и малый круги
Кровообращение птиц имеет большой и малый круги, также как и у
млекопитающих; но в отличие от них, большой круг осуществляется не левой, а
правой дугой аорты.
Выходящая из левого желудочка аорта разветвляется на артерии, несущие кровь в
различные органы, в том числе к ногам и крыльям. От ног кровь поступает в
почечно-портальную систему, а от неё течёт в нижнюю полую вену.
Эти многочисленные артерии, в том числе сонная и подключичная, ответвляются
из двух безымянных артерий, исходящих от аорты; а сама она разворачивается и
продолжается над позвоночным столбом как спинная аорта, от которой также
исходят мощные стволы, питающие органы.
Левая дуга аорты у птиц редуцирована, в сравнении с организмом
пресмыкающихся. В частности, вместо брюшной вены наличествует копчиковобрыжеечная. Расставшаяся с кислородом кровь из головы и других органов
собирается в капилляры, переходящие в мелкие вены, которые в свою очередь
сливаются в крупные стволы.
114.
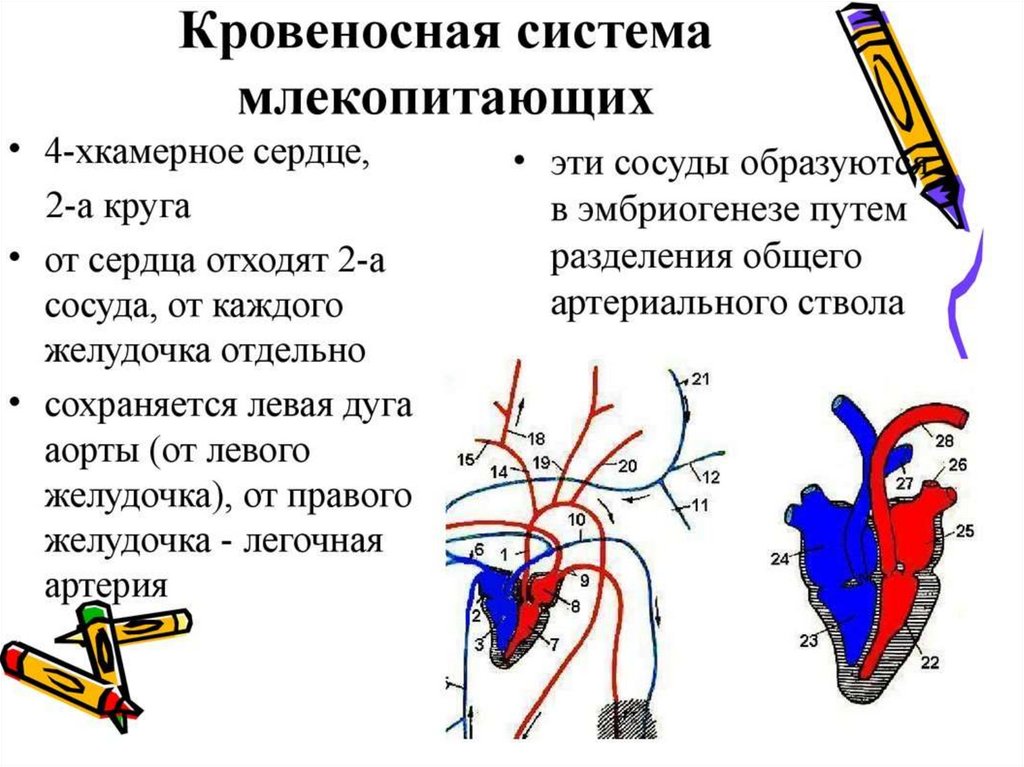
Прогрессивные изменения кровеносной системы млекопитающих сводятся кполному разделению венозного и артериального кровотоков. Это достигается, во-первых,
завершенной четырехкамерностью сердца и, во-вторых, редукцией правой дуги аорты и
сохранением только левой, начинающейся от левого желудочка. В результате все органы
млекопитающих снабжаются артериальной кровью. В венах большого круга
кровообращения также обнаруживаются прогрессивные изменения: возникла безымянная
вена, объединяющая левые яремную и подключичную вены с правыми, в результате чего
остается лишь одна передняя полая вена, располагающаяся справа. Левый кювьеров
проток в виде рудиментарного сосуда sinus coronarius теперь собирает венозную кровь
только от миокарда, а непарная и полунепарная вены - рудименты задних кардинальных
вен, имеют существенное значение в основном в случаях формирования обходных путей
венозного оттока через кава-кавальные анастомозы, формируемые ими.В эмбриональном
развитии млекопитающих, в том числе человека, рекапитулируют закладки сердца и
основных кровеносных сосудов предковых классов.
Сердце закладывается в виде недифференцированной брюшной аорты, которая за счет
изгибания, появления в просвете перегородок и клапанов, становится последовательно
двух-, трех- и четырехка-мерным сердцем. Однако рекапитуляции здесь неполны в связи
с тем, что межжелудочковая перегородка млекопитающих формируется иначе и из другого
материала, чем у рептилий. Поэтому можно считать, что четырехкамерное сердце
млекопитающих формируется на базе трехкамерного сердца, а межжелудочковая
перегородкавляется новообразованием, а не результатом доразвития перегородки
пресмыкающихся. Таким образом, в филогенезе сердца позвоночных проявляется
девиация: в процессе морфогенеза этого органа у млекопитающих рекапитулируют
ранние филогенетические стадии, а затем развитие его идет в ином направлении,
характерном лишь для этого класса.