



































 Биология
БиологияПохожие презентации:










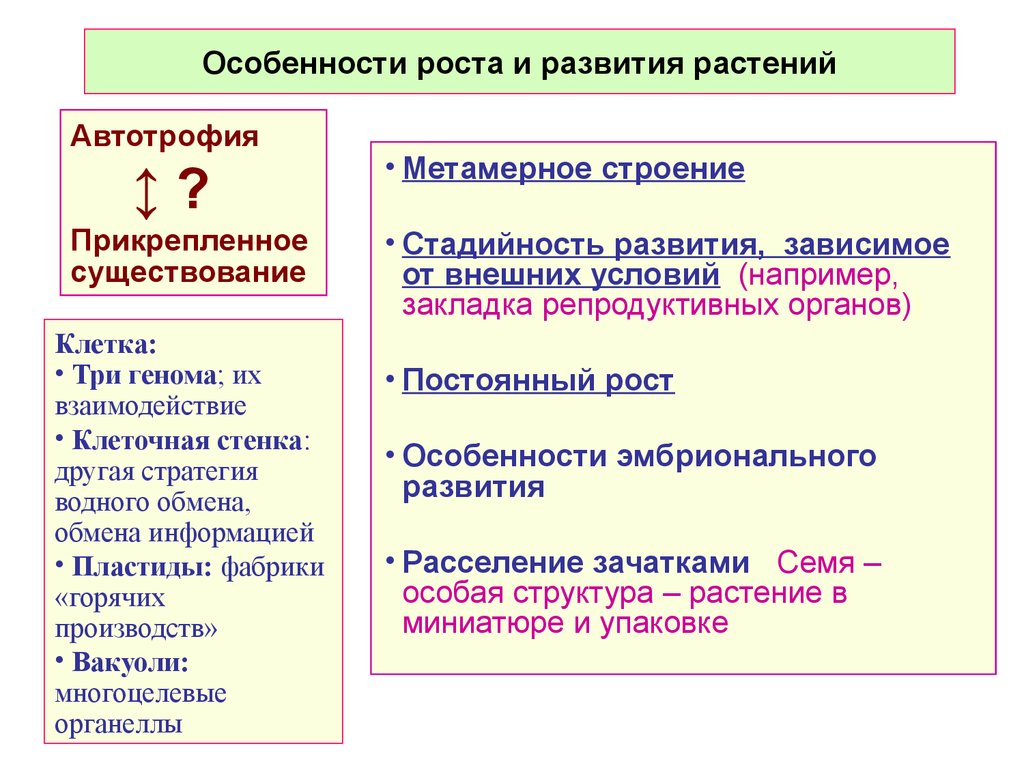
Особенности роста и развития растений
1.
Особенности роста и развития растенийАвтотрофия
↕?
Прикрепленное
существование
Клетка:
• Три генома; их
взаимодействие
• Клеточная стенка:
другая стратегия
водного обмена,
обмена информацией
• Пластиды: фабрики
«горячих
производств»
• Вакуоли:
многоцелевые
органеллы
• Метамерное строение
• Стадийность развития, зависимое
от внешних условий (например,
закладка репродуктивных органов))
• Постоянный рост
• Особенности эмбрионального
развития
• Расселение зачатками. Семя –
особая структура – растение в
миниатюре и упаковке
2.
Гормоны растений – эндогенные регуляторы программ развитияКритерии гормонов:
• Вещество вызывает специфический физиологический ответ у
определенных клеток.
• Разобщено место синтеза и место действия, т.е. необходим транспорт
сигнального вещества по растению (для растений - не строгий критерий)
• Вещество практически не играет роли в основном метаболизме клетки,
используется только для сигнальных целей.
• Вещество должно действовать в низкой концентрации
(начиная с 10-5 моль/л).
Особенности фитогормонов:
• Низкомолекулярны
• Регулируют целые программы развития
• Специфика действия может быть различной в зависимости от места
действия, стадии развития растения, внешних условий.
• В принципе могут синтезироваться любой клеткой растения
• Взаимное действие – «Переговоры гормонов».
3.
Ауксины~0,5 nm
Индолилуксусная кислота
ИУК
наиглавнейший ауксин
Для табака было показано, что треть
всей ИУК находится в хлоропластах,
две трети – в цитозоле.
Триптофан – предшественник ИУК –
образуется в хлоропластах
Индолилмасляная кислота (ИМК) – у горчицы и пшеницы,
хлоро-индолилуксусная (горох), фенилуксусная (ФУК)
кислоты – природные соединения, обладающие
ауксиновой активностью. Присутствуют не у всех
растений
2,4-Д, α–НУК – синтетические соединения.
В малых дозах – ауксины, в больших - гербициды
4.
Что важно для ауксиновой активности и пути формирования активной ИУК5.
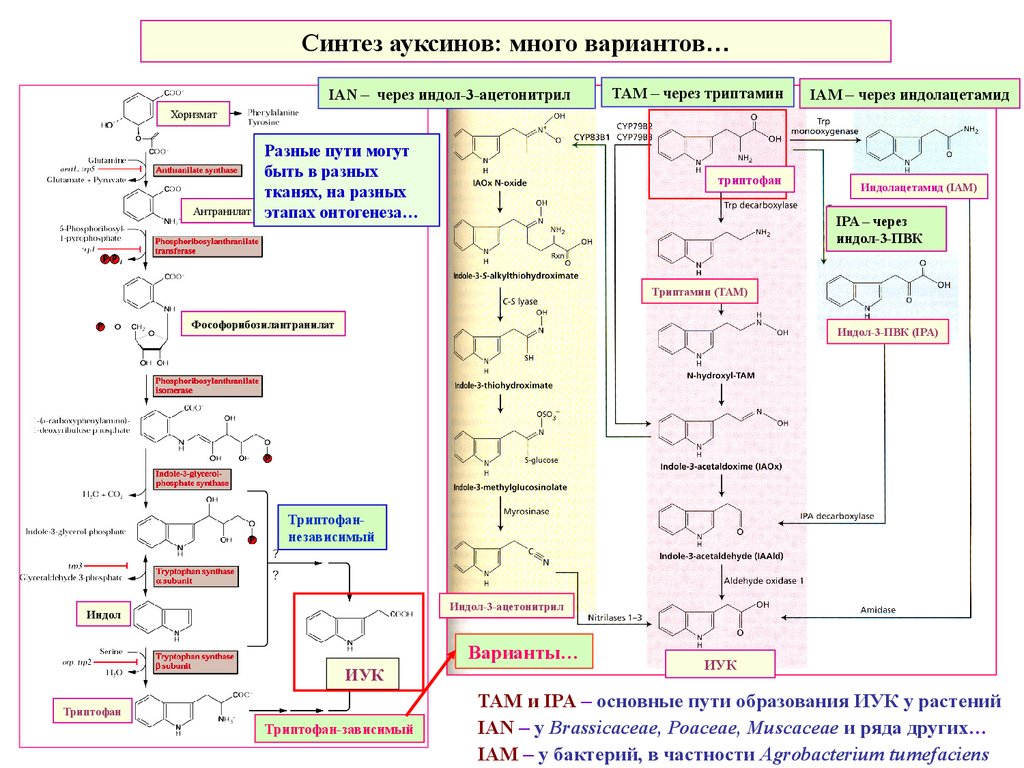
Синтез ауксинов: много вариантов…IAN – через индол-3-ацетонитрил
TAM – через триптамин
IAM – через индолацетамид
Хоризмат
Антранилат
Разные пути могут
быть в разных
тканях, на разных
этапах онтогенеза…
триптофан
Индолацетамид (IAM)
IPA – через
индол-3-ПВК
Триптамин (ТАМ)
Фософорибозилантранилат
Индол-3-ПВК (IPA)
Триптофаннезависимый
Индол-3-ацетонитрил
Индол
Варианты…
ИУК
Триптофан
Триптофан-зависимый
ИУК
TAM и IPA – основные пути образования ИУК у растений
IAN – у Brassicaceae, Poaceae, Muscaceae и ряда других…
IAM – у бактерий, в частности Agrobacterium tumefaciens
6.
Обратимая инактивация ауксинов (образование конъюгатов) и их катаболизмЗначительная часть ауксинов
образует неактивные конъюгаты,
которые хранятся в запасающих
органах и семенах
Низкомолекулярные конъюгаты:
с глюкозой, инозитолом, кислотами
Катаболизм:
а/ декарбоксилирующий - за
счет работы пероксидазы,
б/ без декарбоксилирования
Высокомолекулярные:
с глюканами (7–50
остатков glu) с пептидами,
с гликопротеинами
гликозилирование
7.
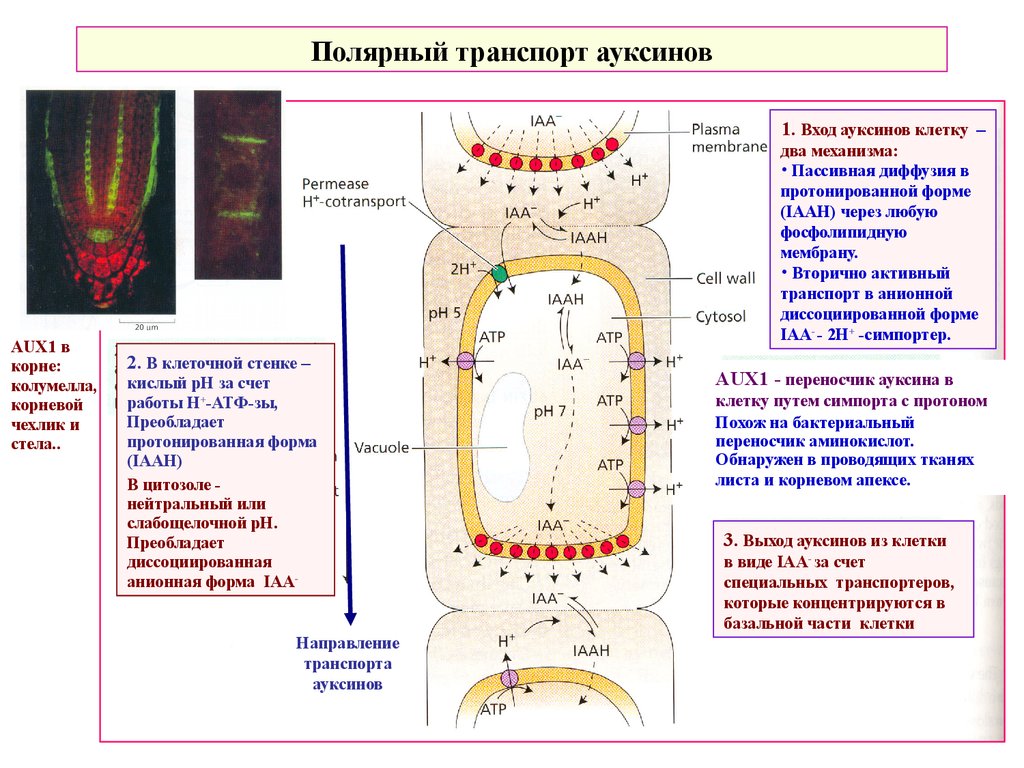
Полярный транспорт ауксинов1. Вход ауксинов клетку –
AUX1 в
корне:
колумелла,
корневой
чехлик и
стела..
два механизма:
• Пассивная диффузия в
протонированной форме
(IAAH) через любую
фосфолипидную
мембрану.
• Вторично активный
транспорт в анионной
диссоциированной форме
IAA- - 2H+ -симпортер.
2. В клеточной стенке –
кислый рН за счет
работы Н+-АТФ-зы,
Преобладает
протонированная форма
(IAAH)
В цитозоле нейтральный или
слабощелочной рН.
Преобладает
диссоциированная
анионная форма IAAНаправление
транспорта
ауксинов
AUX1 - переносчик ауксина в
клетку путем симпорта с протоном
Похож на бактериальный
переносчик аминокислот.
Обнаружен в проводящих тканях
листа и корневом апексе.
3. Выход ауксинов из клетки
в виде IAA- за счет
специальных транспортеров,
которые концентрируются в
базальной части клетки
8.
Полярный транспорт ауксиновВыход ауксинов из клетки
В цитозоле рН 7,2, ауксин в
диссоциированной форме,
поэтому он концентрируется в
клетке (так как IAA- плохо
проходит через мембрану).
Выход – семейство переносчиков PIN
(название от pin-shaped
inflorescences – булавковидное
соцветие, по мутанту PIN1)
Они располагаются как и предсказано
– по линии транспорта ауксинов.
Разные белки – в разных тканях.
ИУК, формирующийся в
листьях,
транспортируется также
неполярно по флоэме.
При этом скорость
транспорта выше, чем
полярного.
Загрузка и разгрузка –
переносчиками. При
этом неполярный
транспорт видимо
важен для камбиального
деления клеток,
формирования каллозы
в ситовидных трубках и
формирования боковых
корней. Он может
вносить значительный
вклад в поступление
ауксина в корни.
Работа PIN связана с другими белками
– регуляторными и
транспортными.
PIN1 – вертикальный
транспорт ИУК от
побега к корню
PIN3 – возврат ауксина
в сосудистую паренхиму
В частности, АТФ-зависимые
переносчики, принадлежащие к
семейству MDR/PGR белков
(multidrug resistance/Pglycoprotein - PGPs) -. Это
интегральные белки –анионные
транспортеры.
У арабидопсиса – 21, у риса – 17 PGP.
Три PGPs – это транспортеры
ауксина.
PIN и PGPs действуют синергично.
9.
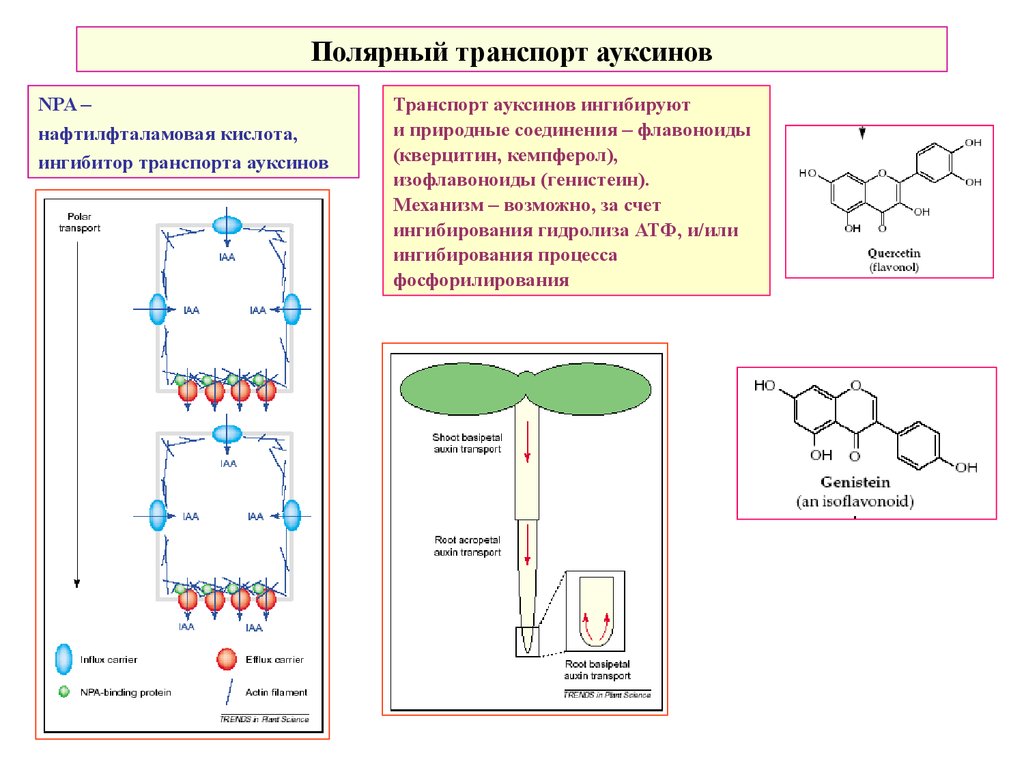
Полярный транспорт ауксиновNPA –
нафтилфталамовая кислота,
ингибитор транспорта ауксинов
Транспорт ауксинов ингибируют
и природные соединения – флавоноиды
(кверцитин, кемпферол),
изофлавоноиды (генистеин).
Механизм – возможно, за счет
ингибирования гидролиза АТФ, и/или
ингибирования процесса
фосфорилирования
10.
Содержание свободной ИУК в контрольном и трансгенном растении табакаКонтрольное растение
Трансгенное растение с двумя
бактериальными генами, кодирующими
ферменты IAM-пути синтеза ИУК –
Trp-монооксигеназы и IAM-гидролазы
11.
«Работа» ауксинов1. Деление клеток - образование CDKs (совместно с цитокининами, которые «запускают» циклины)
2. Удлинение клетки.
Ауксин способствует росту стеблей и колеоптилей, но ингибирует рост корней.
Оптимальная концентрация ИУК для роста растяжением колеоптиля овса и стебля гороха – 10 -6 - 10-5 М. Для
других растений – несколько ниже.
Для корня сложнее. Ауксин индуцирует продукцию этилена, коий ингибирует рост. Однако, если
заингибировать синтез этилена, то показано, что концентрации 10 -9 - 10-10 М - стимулирует рост корня,
а концентрации которые стимулируют рост стебля (10-6 ) – строго ингибируют. Лаг-фаза действии ауксина –
от 10 минут.
Гипотеза кислого роста – ауксин активирует Н+АТФ-зу плазмалеммы. Действие ауксина на изолированную
плазмалемму – повышение активности Н+АТФ-зы на 20%. Интактные клетки – на 40%.Действует и на
синтез Н+АТФ-зы и на ее встраивание в плазмалмму (t – 40 -50 минут)
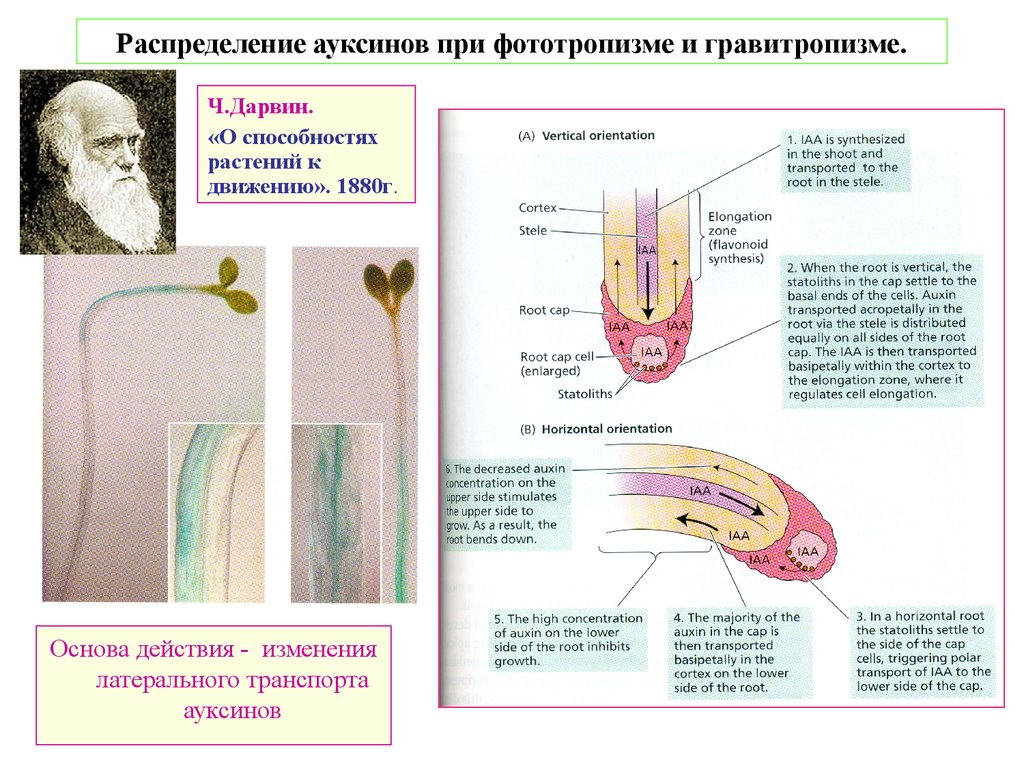
3. Фототропизм и гравитропизм.
Фототропизм. Рецепторы – фототропины 1 и 2.
Механизм – за счет неравномерного латерального распределения ауксинов. Это происходит за счет
неравномерного распределения латерально расположенных PIN3 и дестабилизации
базально расположенных PIN1. Затем происходит неравномерное подкисление апопласта.
4. Ауксин влияет на процессы развития.
- Апикальное доминирование.
Механизм- скорее всего, за счет влияния на развитие ксилемы и межпучковой склеренхимы.
- Развитие флоральных почек и филлотаксис.
- Увеличивает закладку и количество боковых и адвентивных корней.
- Индукция дифференциацию проводящих систем.
- Ауксин участвует в опадении листьев и плодов.
В молодых листьях ауксина много, в зрелых он прогрессивно уменьшается
и его очень мало в листьях, «готовых» к опадению.
- Ауксин способствует развитию плодов.
12.
Распределение ауксинов при фототропизме и гравитропизме.Ч.Дарвин.
«О способностях
растений к
движению». 1880г.
Основа действия - изменения
латерального транспорта
ауксинов
13.
«Клеточки памяти» для ауксинов[С]
[С]
14.
Цитокинины и их биосинтез?
В отличие от ауксинов, среди цитокининов
нет «наиглавнейшего».
В рамочках – синтетические соединения с
цитокининовой активностью. 6-BAP - ?
In vivo присутствуют и рибозиды и
риботиды. Но активная форма –
«свободное» основание…
15.
Цитокинины найдены в т-РНК. Зачем?16.
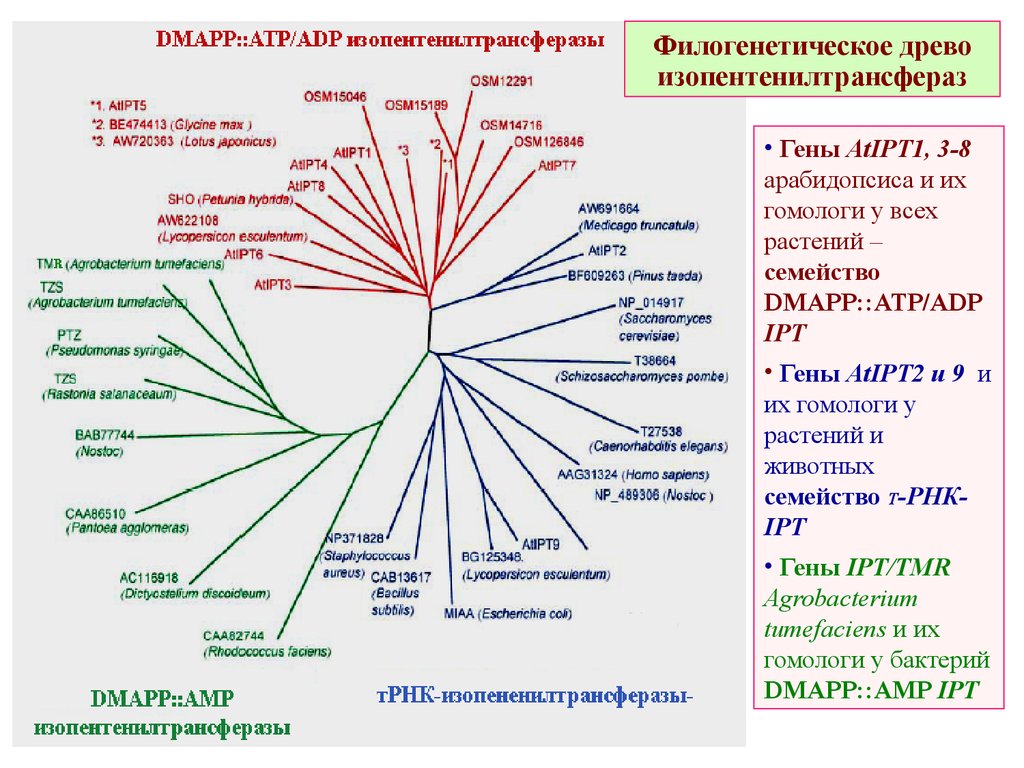
Филогенетическое древоизопентенилтрансфераз
• Гены AtIPT1, 3-8
арабидопсиса и их
гомологи у всех
растений –
семейство
DMAPP::ATP/ADP
IPT
• Гены AtIPT2 и 9 и
их гомологи у
растений и
животных
семейство Т-РНКIPT
• Гены IPT/TMR
Agrobacterium
tumefaciens и их
гомологи у бактерий
DMAPP::AMP IPT
17.
Тканеспецифичность экспрессии некоторых генов AtIPTв корне
AtIPT7 –
эндодерма зоны
элонгации корня
AtIPT1 –
предшественники
ксилемы в
прокамбии
AtIPT3 – флоэма,
молодые сосуды и
перицикл.
AtIPT5 – кончик
корня и зачатки
боковых корней
18.
Конъюгаты цитокининов19.
Транспорт цитокининов2. Z, IP, ZOG
1. ZR, (также ZMP, Z, iPMP, iPA, iP)
A. Транспорт между
органами растения:
1. Ксилемный транспорт. Основная
транспортная форма - зеатинрибозид. Транспорт от корня к
стеблю.
2. Флоэмный транспорт.
Свободные цитокинины и
гликозиды. В обоих
направлениях.
Б. Транспорт между клетками:
PUP
ENT
1. Белки PUP свободные
цитокинины (Z, IP)
2. Белки ENT цитокининнуклеозиды (ZR, IPR)
20.
Межклеточные переносчики свободных цитокининов – белки PUP1 и PUP2Относятся к семейству
пуринпермеаз – осуществляют
также транспорт аденина
• Гены AtPUP1 и AtPUP2
различаются по пространственному
характеру экспрессии,
имеют разные функции
предотвращение
потери цитокининов
через гидатоды при
гуттации
PUP2::GUS
PUP1::GUS
обмен цитокининами
между проводящей
системой и окружающими
тканями
21.
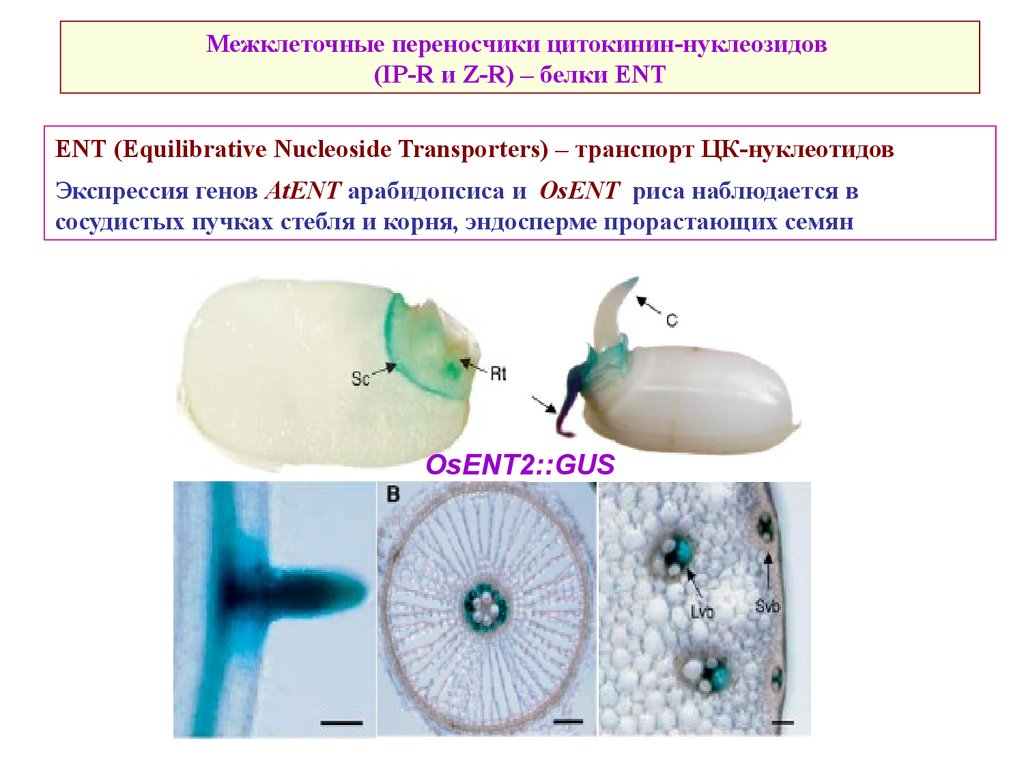
Межклеточные переносчики цитокинин-нуклеозидов(IP-R и Z-R) – белки ENT
ENT (Equilibrative Nucleoside Transporters) – транспорт ЦК-нуклеотидов
Экспрессия генов AtENT арабидопсиса и OsENT риса наблюдается в
сосудистых пучках стебля и корня, эндосперме прорастающих семян
OsENT2::GUS
22.
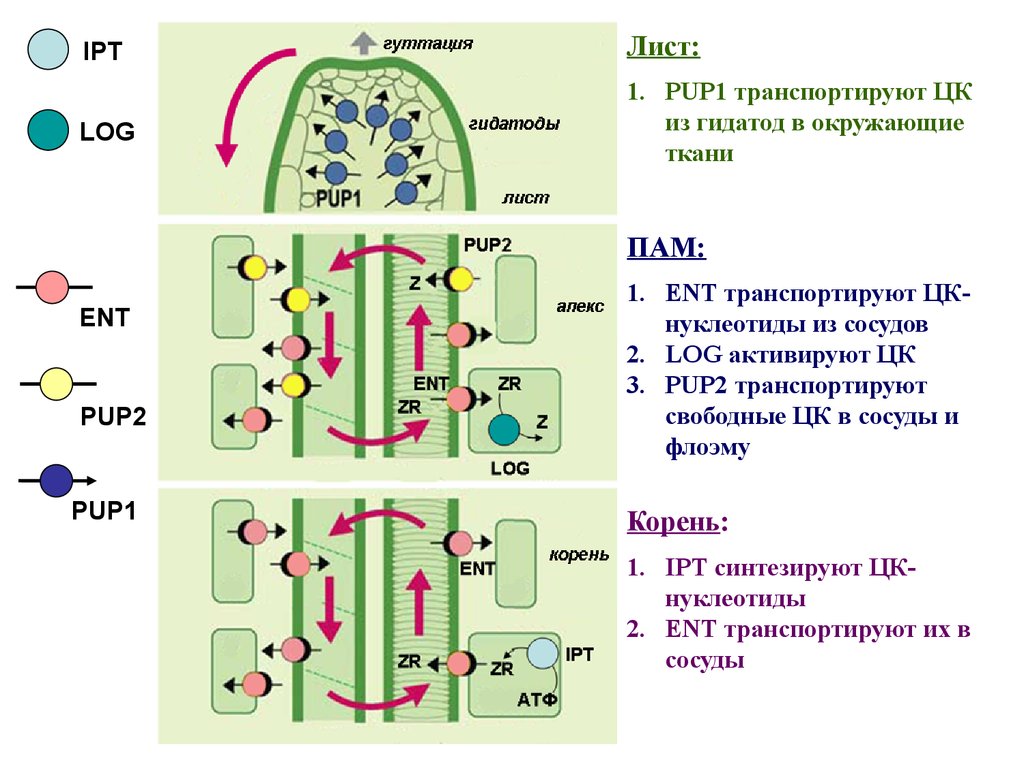
IPTЛист:
LOG
1. PUP1 транспортируют ЦК
из гидатод в окружающие
ткани
ПАМ:
ENT
PUP2
PUP1
1. ENT транспортируют ЦКнуклеотиды из сосудов
2. LOG активируют ЦК
3. PUP2 транспортируют
свободные ЦК в сосуды и
флоэму
Корень:
1. IPT синтезируют ЦКнуклеотиды
2. ENT транспортируют их в
сосуды
23.
«Клеточки памяти» для цитокининов24.
Количество известных гиббереллинов превышает сотню (136 штук…)Ent-гиббереллан:
С6-С5-С6-С5
Две группы GAs:
С20 и С19.
C19 обычно имеют
лактонное кольцо
от С4 к С10
Активные С19 GAs
содержат лактонное
кольцо, карбоксил
у С6 и β-гидроксил
у С3
С2-гидроксил
снимает
активность…
25.
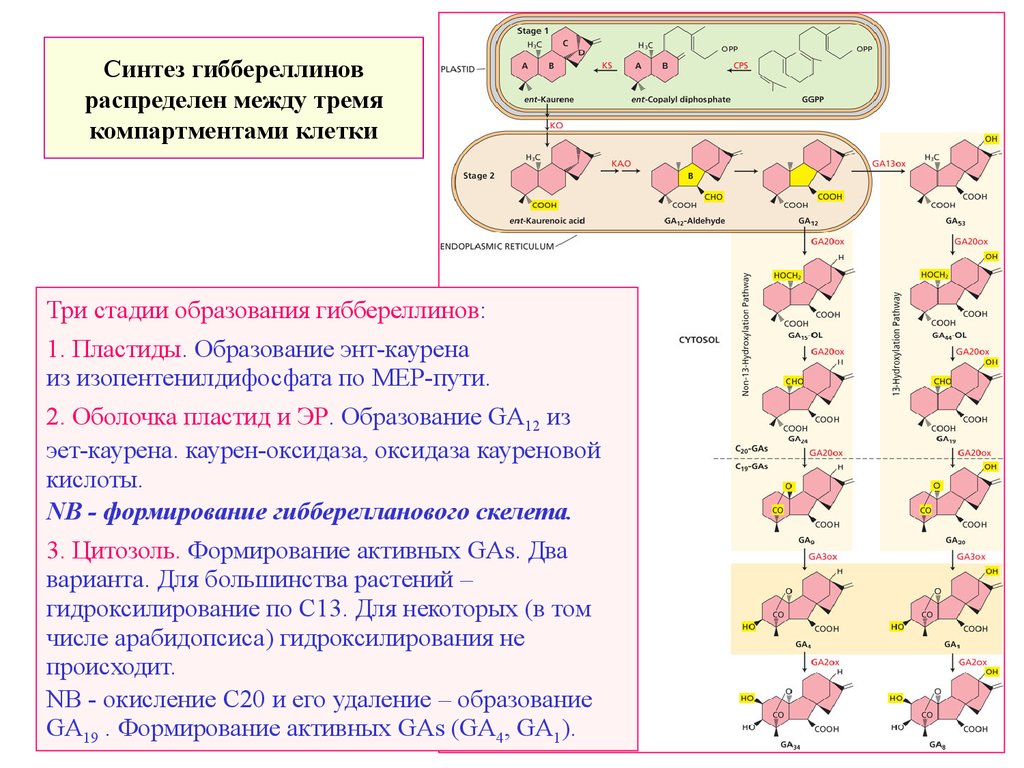
Синтез гиббереллиновраспределен между тремя
компартментами клетки
Три стадии образования гиббереллинов:
1. Пластиды. Образование энт-каурена
из изопентенилдифосфата по MEP-пути.
2. Оболочка пластид и ЭР. Образование GA12 из
эет-каурена. каурен-оксидаза, оксидаза кауреновой
кислоты.
NB - формирование гибберелланового скелета.
3. Цитозоль. Формирование активных GAs. Два
варианта. Для большинства растений –
гидроксилирование по С13. Для некоторых (в том
числе арабидопсиса) гидроксилирования не
происходит.
NB - окисление С20 и его удаление – образование
GA19 . Формирование активных GАs (GA4, GA1).
26.
Варианты конъюгатов и пути катаболизма гиббереллинов различны27.
Синтез гиббереллинов происходит в разных органах и в разное время(GA1-промотор фермента копалил-дифосфатсинтазы CPS, циклизующего GGPP + GUS)
28.
Гиббереллины индуцируют синтез гидролаз при прорастании…29.
«Клеточки памяти» для гиббереллинов30.
Абсцизовая кислота – АБКВ растениях присутствуют
S-формы АБК
У печеночных мхов функцию
АБК, возможно,
выполняет стильбен –
лунулариевая кислота
31.
Синтез абсцизовой кислоты и ее «работа».Синтез имеет
ритмичност
ь – ночью
может быть
в 50 – 60
раз больше,
чем днем
32.
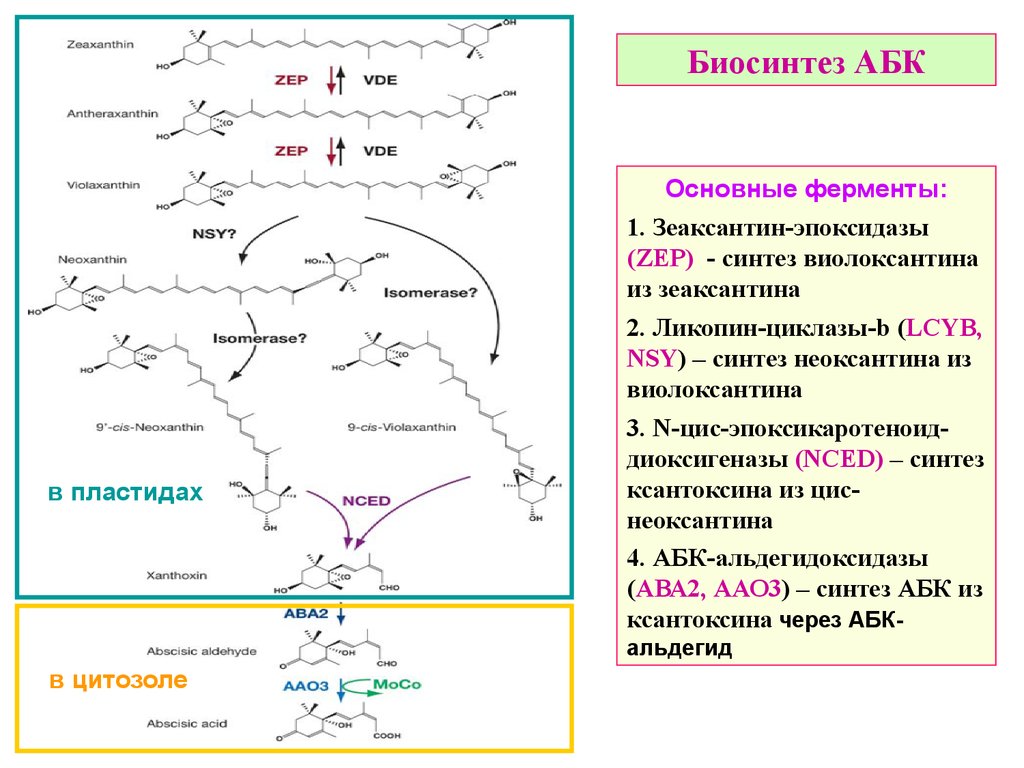
Биосинтез АБКОсновные ферменты:
1. Зеаксантин-эпоксидазы
(ZEP) - синтез виолоксантина
из зеаксантина
2. Ликопин-циклазы-b (LCYB,
NSY) – синтез неоксантина из
виолоксантина
в пластидах
3. N-цис-эпоксикаротеноиддиоксигеназы (NCED) – синтез
ксантоксина из циснеоксантина
4. АБК-альдегидоксидазы
(АВА2, ААО3) – синтез АБК из
ксантоксина через АБКальдегид
в цитозоле
33.
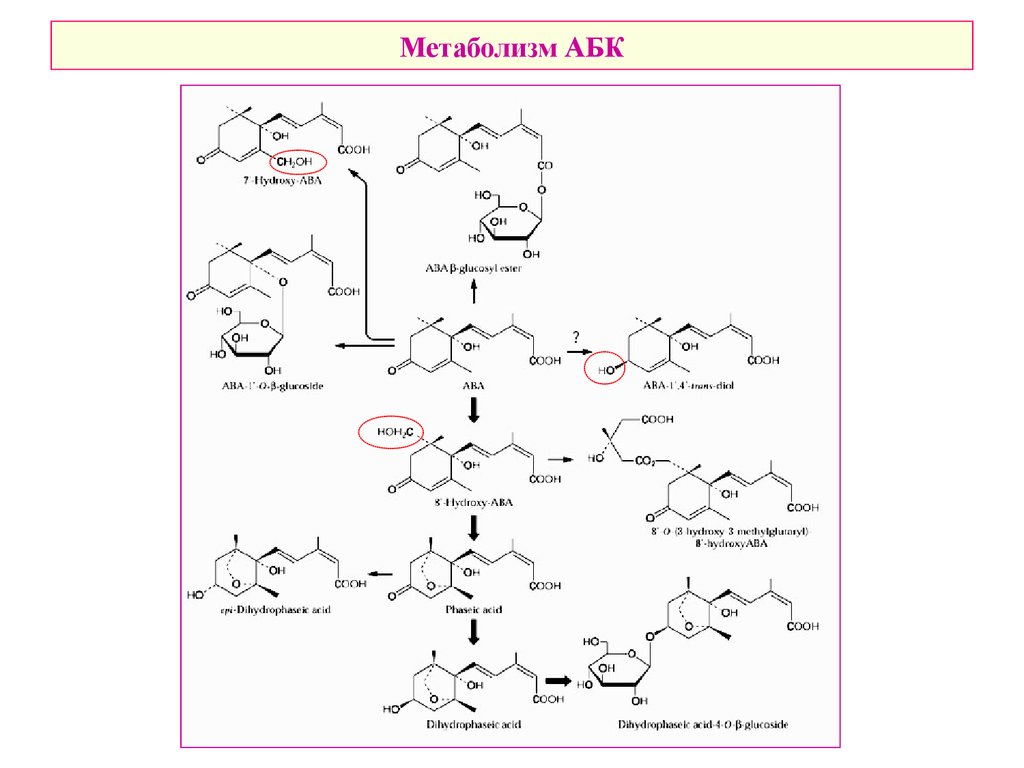
Метаболизм АБК34.
«Клеточки памяти» для АБК35.
Этилен – газообразный гормон растений…Синтез этилена,
его деградация и
его «работа»
CH2 = CH2 этилен
CH2 – CH2 окись этилена
O
CH2 – CH2 этиленгликоль
Ι
OH
Ι
OH
36.
Уровень этилена в тканях регулируется через стабильностьАСС-синтазы (ACS белков)
А – Деградация ACS белков с помощью 26S-протеасомы. Начинается с присоединения
убиквитин-лигазного комплекса ЕТО1-CUL3 к С-концевой последовательности ACS.
Мутации по С-концу ACS (eto2, 3) дают стабильный ACS белок
В – стабилизация ACS белков путем фосфорилирования по консервативным остаткам
серина: с помощью Са2+-зависимых киназ CDPK (все ACS) и MAPКК киназы МРК6 (ACS2 и
6).
37.
Эффекты этиленаА. «Тройной ответ» у
6-дневных
проростков
гороха,
обработанных
10ppm этилена
В. Эпинастии у
томатов
С. «Старение»
цветков
(STS –тиосульфат
серебра,
ингибитор
действия
этилена)
D. Формирование
корневых
волосков у 2дневных
проростков
салата-латука,
обработанных
10ppm этилена
38.
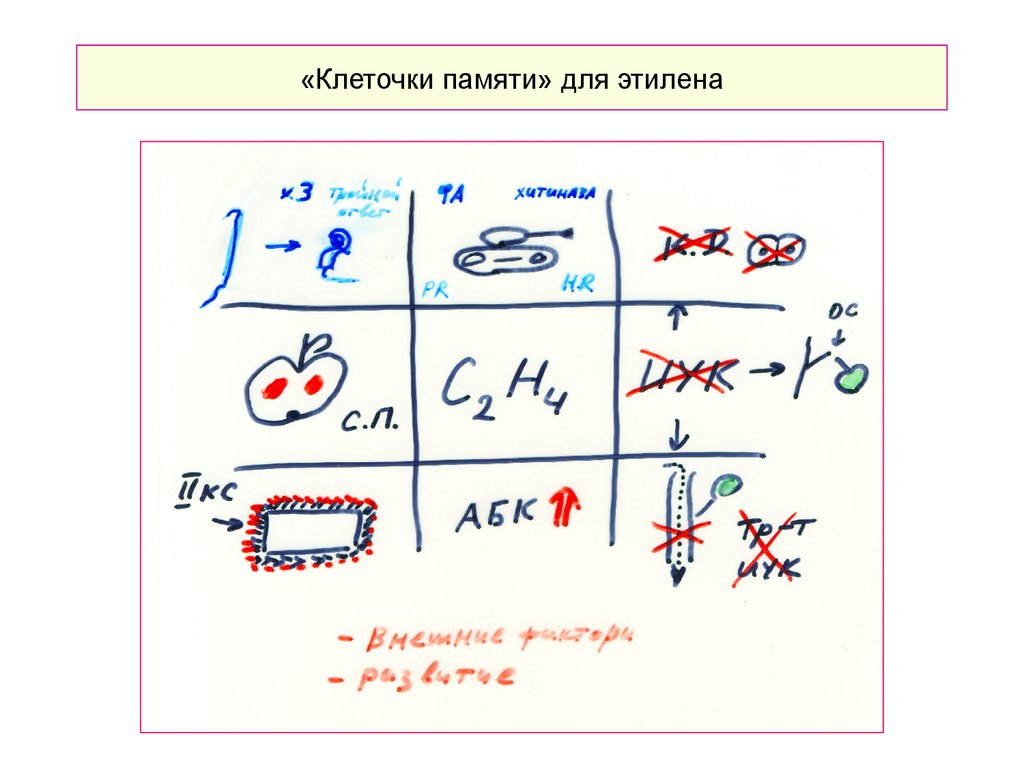
«Клеточки памяти» для этилена39.
Структура и действие брассиностероидов (брассинолидов)• Карликовость
• Образование ксилемы
• Мужская фертильность
• Удлинение клеток за счет активации ХЕТ
• Деление клеток - транскрипция CycD3
Мутанты по синтезу и рецепции
брассинолидов
40.
Структура брассиностероидов1
Вариации в кольце А
2
3
Вариации боковой цепи
Вариации в
кольце В
41.
Синтезбрассиностероидов
Существуют
альтернативные пути
синтеза – «позднее» и
«раннее» окисление по
С6…
42.
Метаболизм эпи-брассинолида43.
Действие брассинолидов• Усиливают растяжение проростков, но медленнее ауксинов (начало через 30 минут,
длительность 1,5 – 2 часа, для ауксинов – 10 мин. и 30 -45 мин). Синергизм действия с
ауксинами. .
• На молекулярном уровне эффект обусловлен активацией генов ксилоглюканэндотрансгликозилаз (КсЭТ). Если ауксины запускают процесс растяжения, то
брассиностероиды важны для его длительного поддержания.
• Частичная и полная мужская стерильность при недостаточности брассиностероидов.
У многих растений, мутантных по генам биосинтеза брассиностероидов, тычиночные
нити не достигают нужной длины, достаточной для самоопыления. Но даже в том случае,
когда пыльцевые зерна попадают на поверхность рыльца, рост пыльцевой трубки
существенно замедлен.
• Брассиностероиды взаимодействуют не только с ауксинами, но и с гиббереллинами,
усиливая растяжение клеток.
• Обсуждают участие брассиностероидов при передаче световых сигналов и "переговоры"
путей фоторецепции и брассиностероидов.
• Брассиностероиды регулируют процессы клеточной дифференцировки. У мутантов bri 1
нарушено формирование столбчатого мезофилла. Кроме того, уменьшено количество
проводящих элементов ксилемы. Брассинолиды регулируют вторичный запуск синтеза
ФАЛ и ГКК с дальнейшей сильной лигнификацией и программированной гибелью
клеток.
• Действие на корневую систему брассиностероидов и ауксинов противоположно: если
ауксины стимулируют образование боковых корней, то брассиностероиды ингибируют их
образование.
• В больших дозах брасииностероиды сдерживают рост и повышают устойчивость к
неблагоприятным внешним факторам (перегреву, заморозкам, засухе, инфекции).
44.
«Клеточки памяти» для брассинолидов45.
Структура и метаболизмжасмонатов
46.
Синтез жасмонатов«Запасной» OPDA в
мембранах
хлоропластов – до
90% от всего ОPDА.
OPDA – 12-оксофитодиеновая
кислота
LOX – липоксигеназа
AOS – аллен-оксид
синтаза
АОС – аллен-оксид
циклаза
OPR – OPDA-редуктаза
47.
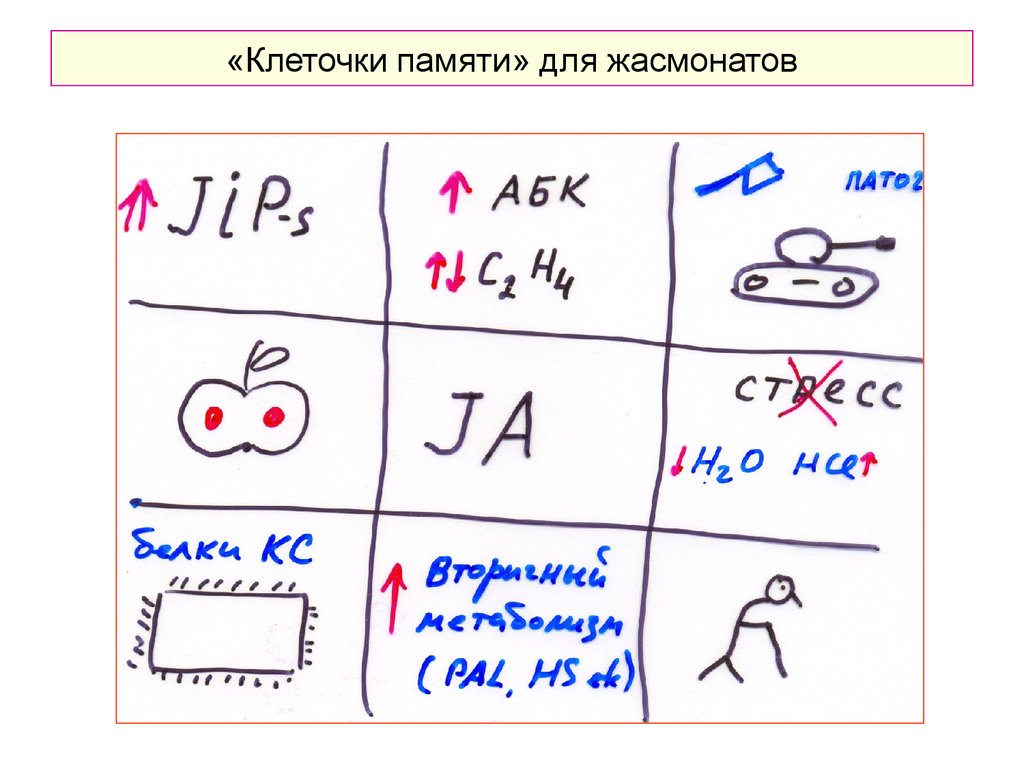
«Клеточки памяти» для жасмонатов48.
Салициловая кислота – «кандидат» в фитогормоны?СА регулирует термогенез у
ароидных лилий
СА антогонист этилена –
блокирует превращение
АСС в этилен
СА участвует (запускает?) в
защитных реакциях
растений от вирусов и
патогенов – участие в
системной устойчивости.
На рисунке – активация
синтеза СА у ВТМинфицированных
устойчивых растений
табака.
Перечеркнутые стрелки – эти
пути синтеза не
задействованы в резком
повышении СА при
инфекции.
49.
Взаимодействие гормоновИсторическая картинка Фольке Скуга…
50.
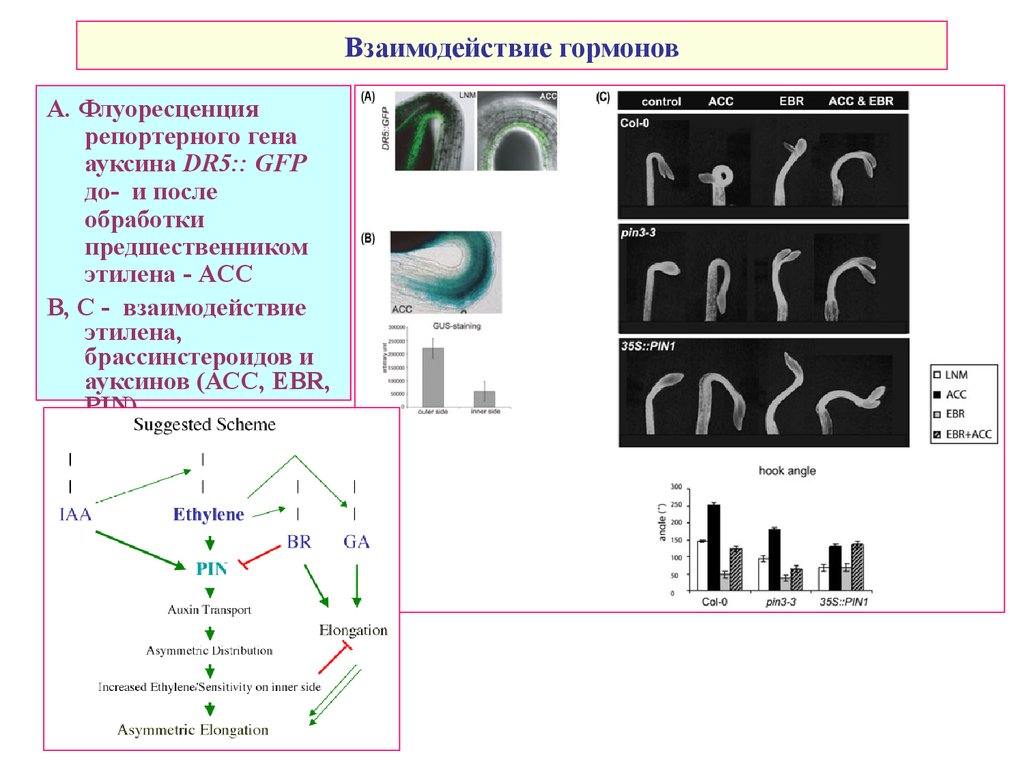
Взаимодействие гормоновА. Флуоресценция
репортерного гена
ауксина DR5:: GFP
до- и после
обработки
предшественником
этилена - ACC
В, С - взаимодействие
этилена,
брассинстероидов и
ауксинов (АСС, EBR,
PIN)