

















")



 состоит из:")












")

 Биология
БиологияПохожие презентации:










Транспорт газов кровью. Газообмен в тканях. Регуляция дыхания
1.
Лекция по нормальной физиологии длястудентов 2-го курса 1-го и 2-го медицинского
факультета, обучающихся по специальности
«Лечебное дело»
2016
В.М.
Система дыхание
Лекция № 2
2. ДЫХАНИЕ.
Транспорт газов кровьюГазообмен в тканях
Регуляция дыхания
3.
Газообменчерез легочную
мембрану
зависит от:
поверхности,
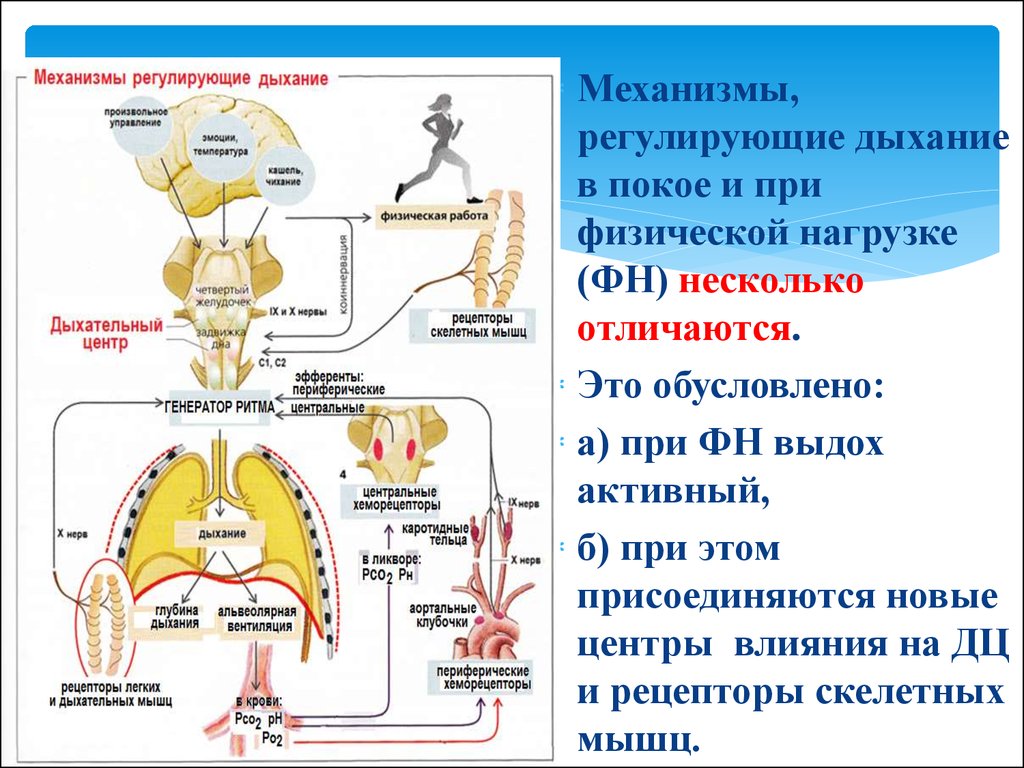
через которую
осуществляется
диффузия (S),
толщины
мембраны (L),
Градиента
давления газов
в альвеолах
и крови ( P),
коэффициента
диффузии (k),
Состояния
мембраны
4. Закон Фика
ЗАКОН ФИКА: М = k S/L P:где, коэффициент диффузии (k) зависит от природы
газа, температуры и среды, в которой происходит
диффузия (см. учебник).
К примеру, СО2 в жидкости диффундирует в 13.000
раз, а О2 в 300.000 раз медленнее, чем в газовой среде.
Поэтому О2 растворяется в 23 раза хуже, чем СО2.
В результате: в 100 мл артериальной крови
растворено лишь 0,3 мл кислорода! Но этого
количества достаточно чтобы создать РаО2 – 100 мм
рт.ст.
5. КЕК и гемоглобин
В крови легких привзаимодействие О2 с Fе2+
гема , образуется легко
диссоциирующее
соединение
(оксигемоглобин) HbO2
Количество О2, которое
транспортируется кровью
(КЕК) всецело
определяется
концентрацией в крови
гемоглобина (см. рис.).
6. Газообмен в эритроцитах в крови легких
В легких в эритроцит поступает О2 , а из него выходит СО2Учитывая, что в идеальных условиях 1 г. Нb может связать
1,34 мл О2, можно подсчитать кислородную емкость (КЕК).
Например: 15 г% х 1,34 мл О2 = 20 мл О2 в 100 мл крови
(20 об%). Учитывая, что те же 100 мл крови содержат лишь
0,3 мл растворенного О2 можно видеть, что основное
количество транспортируемого кровью кислорода - химически
связанный с гемоглобином - оксигемоглобин.
7. Кривая диссоциации оксигемоглобина
Но образование HbО2не прямо
пропорционально
уровню РО2 в крови, а
идет по указанной на
рисунке экспоненте.
Кровь насыщается О2
почти до 100% при РО2
около 100 мм рт. ст.
Даже при подъеме в
горы, когда, к
примеру, РО2 = 80 мм
рт.ст. HbО2
приближается к 100%.
8. Биологическое значение указанной кривой
В смешанной венознойкрови, полученной в покое
из правого предсердия,
РvО2 = 40 мм рт. ст. Но при
этом HbО2 остается еще
более 70% (см. рис.)!
При КЕК в 20 мл/100 мл это
составляет более 15 мл/100
мл крови, что создает
венозный резерв О2.
9. Венозный резерв HbО2
И при снижении РvО2еще до 20 мм рт. ст. в
крови остается лишь
около 30% HbО2. Таким
путем используется
резерв О2 при
мышечной работе.
Кроме того, высокое
сродство Hb к О2 при
относительно низких
уровнях РаО2 позволяет
человеку даже
«поболеть» пневмонией.
10. Кривые диссоциации различных видов Hb
По активности взаимодействия сHb:
1 – СО, но СОHb диссоциирует
очень медленно (возникает
опасность отравления);
2 – О2 с миоглобином;
3 – О2 с гемоглобином плода
(FHb) – активная ассоциация;
О2-метгемоглобин (MtHb окисленное Fe3+ ). Соединение
стойкое - транспорта О2 нет.
Для восстановления в Fe2+ в
эритроците есть фермент –
редуктаза метгемоглобина.
11. Кривые диссоциации НbО2 не постоянны
При увеличении в кровирН, снижении СО2 или
температуры – кривая сдвигается
влево (растет сродство).
При снижении рН, повышении
температуры – сдвиг вправо
(ускоряется диссоциация)
[эффект Бора].
Внутри эритроцита регулятором
диссоциации является уровень
2,3-ДФГ. Так, при анемии в
эритроците возрастает
концентрация 2,3-ДФГ, что
способствует увеличению
диссоциации HbО2. О2 - активнее
поступает в ткани.
12. Газообмен в тканях
Доставка О2 к тканям происходит кровотоком путем конвекции.А в тканях снова происходит диффузия газов.
Газообмен в тканях также, как и газообмен в легких,
зависит от 5 основных факторов:
1) площади диффузии; 2)градиента парциального
давления газов между кровью и клетками; 3)
расстояния, которое проходит газ; 4) коэффициента
диффузии и 5) состояния мембран.
13. Доставка О2 тканям
Количество О2, поступившее к органу, может бытьопределено по Арт. вен. Разнице содержания О2 (АВРО2). Зная объем кровотока и содержание О2 в
приносящей артерии и выносящей вене можно
определить количество О2 поглощенного
органом.
Для определения количества О2 в крови необходимо
использовать кривую диссоциации HbО2.
Кровоток и АВР-О2 зависят и от уровня метаболизма
органа: чем интенсивнее обмен веществ, тем больше
потребляется кислорода, а значит и больше АВР-О2.
14. Цилиндр Крога
В связи с плохой растворимостьюО2 в жидкостях требуется высокие
градиенты РО2.
Обычно РО2: около митохондрий
5-10 мм рт. ст.;
в тканевой жидкости у капилляра 20-40 мм рт. ст.;
а в притекающей крови - более 7080 мм рт. ст.
Это и обеспечивает поступает О2
из крови к митохондриям, где и
используется при окислении.
15. Обмен СО2 между тканями и кровью
Обычно в большинстве тканей уровень РСО2 близок к50-60 мм рт. ст.
В крови, поступающей в артериальный конец
капилляров, РаСО2 составляет около 40 мм рт. ст.
Наличие градиента заставляет СО2 из тканевой
жидкости диффундировать к капиллярам.
РvСО2 в смешанной крови, поступающей в правое
предсердие, составляет 46 мм рт. ст.
Вначале СО2 должен поступить в эритроцит.
16.
ТранспортСО2с
БикарбоКарбУгольная
натный ион гемоглобин
кислота
(НСО3-)
(HHbCO2)
Растворенный
в плазме
17. Транспорт СО2
В венозной крови содержится около 580 мл/л СО2.Поступивший в эритроциты СО2, при участии
имеющейся здесь карбоангидразы , образует про
нестойкую Н2СО3. Образующиеся продукты выходят
в плазму.
Двуокись углерода в крови находится в трех формах:
а) в плазме часть CO2 остается в растворенном виде
- 2,5 мл/100 мл,
б) связанной с глобином гемоглобина
(карбгемоглобин HbCO2) - 3,5-4,5 мл /100 мл крови,
в) в плазме связанной в виде угольной кислоты и ее
солей - 51 мл /100 мл.
18. Диссоциация СО2
Для СО2 такжеимеется
зависимость
скорости
диссоциации, как
от уровня РСО2 в
крови, так и от
уровня РО2 (рис) .
Но выраженность
ее менее значима
для транспорта
СО2, чем для О2.
19. Реакции, идущие в эритроцитах в венозной крови малого круга кровообращения (в легких)
*а) Н++НСО3-<==>H2СО3<==>Н2О+СО2
б) ННbCO2 + O2 <==> HHbO2 +
CO2 <==> HbO2 + H+ + CO2
* - участие
карбоангидразы
На нижней кривой представлена
зависимость газообмена в
легких от времени: СО2
выводится быстро, а О2
поступает более медленно.
Поэтому при увеличении
скорости тока крови (физ.
работа) эритроцит может
недополучать О2.
20. РЕГУЛЯЦИЯ ДЫХАНИЯ
Дыхательные движения выполняются сокращениемскелетных мышц, а они иннервируются
мотонейронами (С4-Т7) спинного мозга.
Поэтому дыхание можно изменить сознательно
(например, при РЕЧИ!), либо влиянием других
центров коры или подкорки (например, лимбической
системой при эмоциях) .
Но обычно дыхание регулируется и как
вегетативные органы (бессознательно).
21.
Дыхание, являясь вегетативной функцией, новыполняется скелетными мышцами. Поэтому
механизмы регуляции его имеют черты,
характерные для регуляции как вегетативных
органов, так и скелетной мускулатуры.
Необходимость осуществления постоянного
дыхания обеспечивается непроизвольной,
автоматической активностью дыхательного
центра. Однако, в силу того, что дыхательные
движения выполняют скелетные мышцы, возможно и
произвольное изменение характера дыхания,
обусловленное воздействием коры больших
полушарий на дыхательный центр.
22.
Механизмы,регулирующие дыхание
в покое и при
физической нагрузке
(ФН) несколько
отличаются.
Это обусловлено:
а) при ФН выдох
активный,
б) при этом
присоединяются новые
центры влияния на ДЦ
и рецепторы скелетных
мышц.
23.
Если во внутренних органах (сердце, кишечник)автоматия обусловлена лишь свойствами водителей
ритма, то в дыхательном центре периодическая
деятельность определяется более сложными
механизмами.
Здесь она обусловлена:
1) согласованным сопряжением активности
различных отделов самого дыхательного центра,
2) поступлением сюда импульсов от рецепторов,
3) поступлением сигналов от других отделов ЦНС,
вплоть до коры.
24. Дыхательный центр (генератор дыхательного ритма) состоит из:
2 -дорсального ядра (ведущее),3 – вентрального ядра,
4 - апнейстического центра (?),
5 - пневмотаксического центра.
25. Дыхательные нейроны.
Обнаружено 11-типовнейронов, возбуждение в
которых можно
зарегистрировать во время
дыхания.
Если они возбуждаются в фазу
вдоха, то именуются
инспираторными.
Если возбуждаются в фазу
выдоха – называются
экспираторными.
26. Автоматия дыхательного центра
Бульбарный отдел дыхательного центра всеже обладает и собственной, пейсмекерной
автивностью, хотя она и не столь выражена,
как в других вегетативных органах с
полноценным автоматизмом. Так, при
полном отделении бульбарного отдела
дыхательного центра, в нем можно
зарегистрировать вспышки активности с
частотой, намного меньшей, чем в
естественных условиях целостного
организма.
27. Межнейронные взаимодействия дыхательного центра /ГЕНЕРАТОР РИТМА ДЫХАНИЯ/
Вентральное ядроИ
Дорсальное ядро
Iα
I-α
I-β
Э
Спинной мозг
Мотонейроны
вспомогательных
мышц вдоха и
выдоха
Спинной мозг
Мотонейроны основных
мышц вдоха
Возбуждение
Торможение
28. Регуляция дыхания в покое
Началовозбуждение I -нейронов: вдох;
выдох : торможение I -нейронов
возбужденными I -нейронами, ( но для этого
нужна суммация: импульсов от I -нейронов +
пневмотаксического центра + рецепторов
растяжения легких)
29. Регуляция дыхания при одышке
Начало –возбуждение I -нейронов +
возбуждение И-нейронов вентрального центра –
глубокий вдох;
Форсированный выдох
торможение I нейронов, возбужденными
I -нейронами + возбуждение Э-нейронов
вентрального центра (здесь дополнительно активно
присоединяются рефлекторные механизмы).
30. Хеморецепторы
Центральныехеморецепторы
расположены в самом
продолговатом мозгу. Они
контролируют метаболизм
мозга.
Периферические
в кровеносных сосудах:
дуге аорты и каротидном
тельце. Т.е. рецепторами
осуществляется контроль за
кровью, поступающей в
мозг.
31. Центральные хеморецепторы
У вентральной поверхности продолговатого мозга на глубине200-400 мкм расположены центральные хеморецепторы
(ЦХР). Нахождение их в мозгу можно объяснить
необходимостью контроля за снабжением О2 нейронов ЦНС.
Омываются они внеклеточной жидкостью, состав которой
определяется метаболизмом окружающих нейронов и
местным кровотоком. Кроме того, состав межклеточной
жидкости во многом зависит от состава спинномозговой
жидкости. Ведущим фактором раздражения этих рецепторов
является концентрация Н+. Они наиболее чувствительны к
изменению параметров крови.
ЦХР стимулируют инспираторные и экспираторные нейроны,
усиливая как вдох, так и выдох. Поэтому, например, при
снижении рН СМЖ лишь на 0,01 вентиляция легких
увеличивается на 4 л/мин.
32. Периферические хеморецепторы
ПХР находятся в бифуркации общих сонных артерий ив аортальных тельцах, находящихся на верхней и
нижней поверхности дуги аорты. Наибольшее
значение для регуляции дыхания принадлежит
каротидным тельцам, контролирующим газовый
состав крови, поступающей к мозгу.
В основе рецепции лежит то, что каротидное тельце
имеет собственное интенсивное кровоснабжение (до
20 мл/мин/г). Но так как О2 в них используется мало, то
АВР-О2 небольшая. Поэтому хеморецепторы более
чувствительны к увеличению РаСО2, чем снижению
РаО2 .
33.
Возбуждаются периферические хеморецепторытакже при снижении рН или повышении РаСО2.
Гипоксия и гиперкапния взаимно усиливают
импульсацию от этих рецепторов.
Импульсы от хеморецепторов достигают
инспираторных нейронов продолговатого мозга и
задерживают выключение вдоха, углубляя дыхание.
Рефлексы, приводящие к изменению активности
дыхания, начинают возникать при уменьшении РаО2
ниже 90 мм рт. ст.
Меньшее значение в регуляции дыхания принадлежит
аортальным хеморецепторам, которые более
заметную роль играют в регуляции кровообращения.
34. Рецепторы легких и дыхательных путей
а)Р. растяжения легких – расположены в гладких
мышцах воздухоносных путей (активируя I -нейроны,
тормозящие активность I -нейронов, они
останавливают глубокий вдох );
В каждом легком имеется до 1000 рецепторов.
Механизм раздражения рецепторов заключается в
следующем. Мелкие бронхи растягиваются эластической тягой, возникающей в альвеолах: чем больше
расширяются альвеолы, тем сильнее растяжение
структурно связанных с ними воздухоносных путей.
Крупные воздухоносные пути структурно не связаны с
легочной тканью и растягиваются "отрицательностью
давления" в плевральной полости.
35. Другие рецепторы легких
б) Рецепторы дыхательных мышц . Функционируютпо принципу гамма-петли межреберных и мышц
стенок живота (при внешнем затруднении
дыхательных движений они автоматически
усиливают силу сокращения мышц).
Примечательно, что их почти нет в диафрагме!
в) ирритантные рецепторы – находятся в
эпителиальном и субэпителиальном слоях;
г) J-рецепторы (юкстамедуллярные рецепторы)
называются так потому, что залегают в стенках
альвеол около капилляров.
36. Ирритантные рецепторы
Раздражителями этих рецепторов являются едкие газыРаздражение ирритантных рецепторов вызывает у человека
такие неприятные ощущения, как першение, жжение,
кашель.
Раздражение ирритантных рецепторов через блуждающий
нерв может вызвать и рефлекторное сокращение гладких
мышц бронхов. К примеру, этот рефлекс лежит в основе
бронхоспазма при возбуждении рецепторов гистамином,
образующимся при бронхиальной астме.
Физиологическое значение указанного рефлекса
заключается в том, что при вдыхании токсических
веществ сужением просвета бронхов, снижая
вентиляцию альвеол, уменьшить поступление этих
веществ в альвеолы, а значит и в кровь.
37. J-рецепторы (юкстамедуллярные рецепторы)
Раздражаются они при поступлении биологическиактивных веществ в сосуды малого круга
кровообращения, а так же при увеличении объема
интерстициальной жидкости в легочной ткани.
Вероятно, они вместе с ирритантными рецепторами
при отеке легких вызывают одышку.
Роль их в регуляции дыхания у здорового человека
не очень понятна.
38. Схема всех механизмов, которые могут участвовать в регуляции дыхания
На дыхательныйцентр оказывают и
неспецифическое
влияние многие
нервные центры и
воздействия.