

















































































 Биология
БиологияПохожие презентации:










Империя эукариоты
1.
ИМПЕРИЯЭУКАРИОТЫ
Молчанов А.Ю.
СОШ «Во имя апостола и евангелиста Иоанна Богослова»
2.
Четыре царства эукариотI. Царство Простейшие
II. Царство Растения
III. Царство Животные
IV. Царство Грибы
3.
П рокариоты• 0,5-5 мкм, питаются только
осмотрофно.
• Одноклеточные и колониальные
• Нет оформленного ядра и ЭПР.
Кольцевая ДНК.
• Имеется клеточная стенка из
полисахаридов
• Дыхание и фотосинтез происходят
на цитоплазматической мембране;
• Рибосома имеет константу
седиментации 70S и включают три
разных рРНК: 23S, 16S и 5S.
• Не имеют актина и миозина.
Эукариоты
• 7-50 мкм. Питаются осмотрофно,
пиноцитозом и фагоцитозом.
• Фотолитоавтотрофы
• хемоорганогетеротрофы
• Одноклеточные, колониальные и
многоклеточные
• Есть ядро, ЭПР, ап. Гольджи,
лизосомы, митохондрии, пластиды
• Клеточная стенка только у грибов и
растений.
• Дыхание в митохондриях
• Фотосинтез в пластидах;
• Рибосомы типа 80S и включают четыре
разных рРНК: 28S, 168, 5S и 5.8S.
• Имеют актин и миозин. Могут менять
форму, активнодвигаться.
• Митоз и мейоз
4.
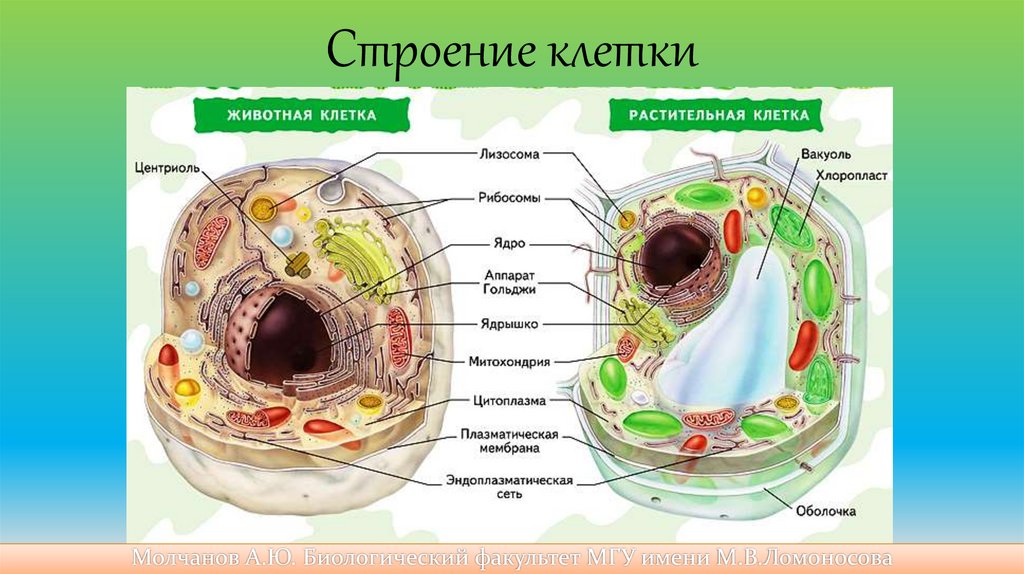
Строение эукариотическойклетки
• Ядро – центральная часть клетки. Имеет собственную
оболочку, состоящую из двух мембран. Хранит
наследственной материал;
• Цитоплазма – основной материал клетки, в котором
осуществляются все обменные процессы;
• Мембрана – образует контур клетки и обеспечивает контакт с
внешней средой.
5.
Плазматическая мембранаСтроение, функции
6.
ПЛАЗМАТИЧЕСКАЯ МЕМБРАНА7.
Строение мембраны• Состоит из жиров, белков и углеводов.
– Жиры – фосфолипиды, сфинголипиды, холестерин и др. Первые
включают водорастворимую (гидрофильную) головку и
водонерастворимые (гидрофобные)хвостики.
– Хвостики представлены остатками жирных кислот.
– Головка в своем составе содержит остаток одной фосфорной
кислоты и либо спирт (напр., холин) или аминокислоту (напр.,
серин).
– Между собой остатки кислот соединены остататком глицерина
(глицеролом).
– Остатками вещества названы так, потому что при образовании
связи некоторые из них потеряли водород (-Н), а некоторые
гидроксильную группу (-ОН).
• Мембрана восстанавливается за счет встраивающихся в нее
пузырьков из ЭПС и аппарата Гольджи.
8.
9.
Мембранны е белки• Биологическая мембрана включает и различные белки:
– интегральные (пронизывающие мембрану насквозь),
– полуинтегральные (погружённые одним концом во внешний или
внутренний липидный слой),
– поверхностные (расположенные на внешней или прилегающие к
внутренней сторонам мембраны).
• Некоторые белки являются точками контакта клеточной
мембраны с цитоскелетом внутри клетки и клеточной стенкой (если
она есть) снаружи.
• Некоторые из интегральных белков выполняют
функцию ионных каналов, различных транспортеров и рецепторов.
10.
Заряд на мембране• Для поддержания тока
ионов.
• Снаружи больше хлора и
натрия.
• Внутри много калия.
• Заряд нужен для
поддержания активности.
* Снаружи NaCl как в океане.
11.
Транспорт через мембрану• Мембрана проницаема только для неполярных веществ: гормоны
жироподобное природы (тестостерон, эстроген), воды, аммиака и др.
• Мембрана не непроницаема для крупных молекул и молекул имеющих
заряд: сахара, аминокислоты, ионы щелочных металлов.
• За перенос через мембрану отвечают мембранные транспортные белки.
• Транспорт этих соединений в клетку делится на активный и
пассивный. Пассивный транспорт — транспорт веществ из области с высокой
концентрацией в область низкой (по градиенту концентрации) без затрат
энергии, то есть диффузия. Она делится на 2 варианта: простая и облегчённая.
– В облегчённой диффузии участвуют белки-переносчики. Например транслоказа.
Переносчиком может быть один или группа белков.
– Простая диффузия обеспечивается каналообразующими белками (например
коннексины и паннексины) .
• Транспорт веществ против градиента концентрации называется активным.
12.
Активны й транспорт• Активный транспорт происходит против градиента концентрации и протекает с затратой
энергии.
• В активном транспорте участвуют белки-переносчики. Энергия, которая требуется для
осуществления активного транспорта, обычно получается транспортными белками при
расщеплении АТФ.
• Один из наиболее изученных белков, осуществляющих активный транспорт — Na+/K+аденозинтрифосфатаза. За полный цикл работы этого насоса в клетку попадают из внешней
среды 2 иона K+ и выбрасывается наружу 3 иона Na+. Или например транспорт глюкозы в
клетку.
13.
Гликокаликс (надмембранный слой)• Гликокаликс — (glycocalyx) [греч. glykys — сладкий и лат. callum — толстая кожа]
«заякоренные» в плазмалемме молекулы олигосахаридов, полисахаридов,
гликопротеинов и гликолипидов.
• Гликокаликс выполняет функцию связи клеток животных с внешней средой и со
всеми окружающими ее веществами. Он не выполняет опорной функции.
• К гликокаликсу относятся олигосахариды рецепторов, которые как правило
связаны с канналообрзующими белками или с ферментами. Поэтому гликокаликс
выполняет рецепторную и маркерную функции.
• Он также участвует в обеспечении избирательности транспорта веществ и
пристеночном (примембранном) пищеварении.
• Наличие гликокаликса характерно для клеток животных (его нет у прокариотов,
растений и грибов).
• Благодаря гликокаликсу обеспечивается адгезия клеток.
14.
Подмембранны й слой• Подмембранный компонент- это периферические белки и элементы
цитоскелета, в первую очередь нитей микрофиламентов актина.
• Нити актина (через периферические белки анкирин и спектрин) связаны
с интегральными белками и образуют плотную сеть пятигранной формы. К
узлам этой сети могут прикрепляться микротрубочки.
• Подмембранные белки участвуют в образовании специфических
клеточных контактов – десмосом.
• У одноклеточных подмембранное пространство ассоциировано с
эктоплазмой. В этом пространстве располагается пелликула, которая
поддерживает форму одноклеточных.
15.
Мембрана и смерть• Переход отдельной молекулы из одного слоя в другой (так
называемый флип-флоп) затруднён, но может происходить
спонтанно, примерно раз в 6 месяцев или с помощью белковфлиппаз и скрамблазы плазматической мембраны. Если в наружном
слое появляется фосфатидилсерин, это является сигналом
для макрофагов о необходимости уничтожения клетки.
• Лимфоциты специально таким образом проверяют
жизнеспособность клеток: если клетка не смогла вернуть
фосфотидилсерин вовнутрь, то она подлежит уничтожению.
16.
Функции клеточной мембраны• Барьерная – избирательное пропускание веществ. Через мембрану
свободно проходят вода и неполярные молекулы;
• Структурная (механическая) – мембрана определяет форму и границу
клетки.
• Транспортная – через каналы в мембране осуществляется пассивный или
активный транспорт;
• Ферментативная — мембранные белки нередко являются ферментами.
Например, плазматические мембраны эпителиальных клеток кишечника
содержат пищеварительные ферменты. А также в мембране каждой клетки
имеются фосфолипазы и G-белки (АТФазы);
• Иммунная - маркировка клетки с помощью антигенов;
• Рецепторная — некоторые белки, находящиеся в мембране,
являются рецепторами.
• Образование клеточных контактов.
17.
Дополнительны е функции• Эти функции выполняет плазматическая мембрана в составе
одно- и двумембранных органелл:
– Энергетическая — при фотосинтезе в хлоропластах и клеточном
дыхании в митохондриях в их мембранах действуют системы
переноса энергии, в которых также участвуют белки.
– Компартмелизующая – разделяет внутреннее пространство клетки
на отсеки для осуществления реакций и накопления веществ.
18.
19.
ЯДРО20.
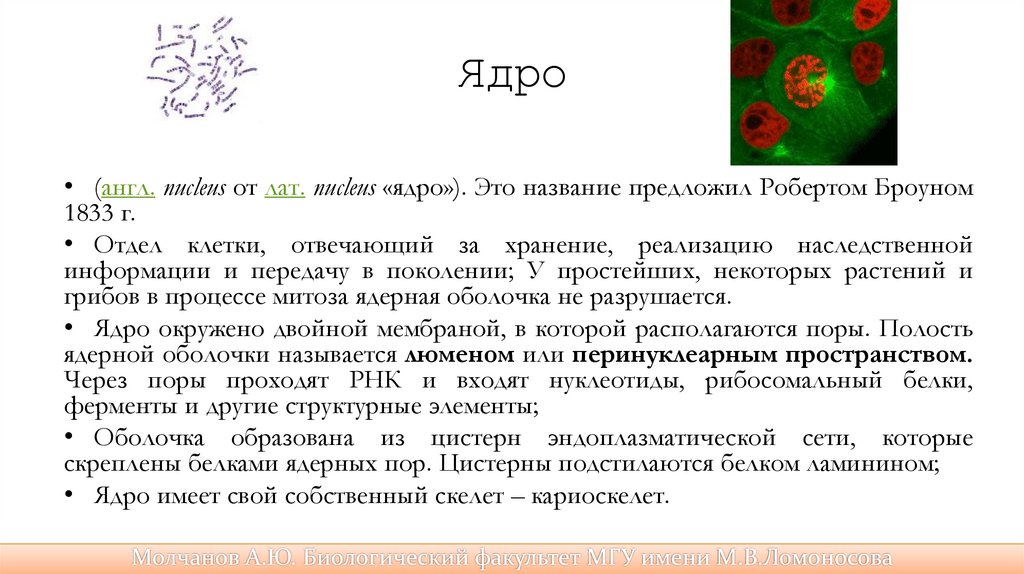
Ядро• (англ. nucleus от лат. nucleus «ядро»). Это название предложил Робертом Броуном
1833 г.
• Отдел клетки, отвечающий за хранение, реализацию наследственной
информации и передачу в поколении; У простейших, некоторых растений и
грибов в процессе митоза ядерная оболочка не разрушается.
• Ядро окружено двойной мембраной, в которой располагаются поры. Полость
ядерной оболочки называется люменом или перинуклеарным пространством.
Через поры проходят РНК и входят нуклеотиды, рибосомальный белки,
ферменты и другие структурные элементы;
• Оболочка образована из цистерн эндоплазматической сети, которые
скреплены белками ядерных пор. Цистерны подстилаются белком ламинином;
• Ядро имеет свой собственный скелет – кариоскелет.
21.
Клеточное ядро22.
Схема строения клеточного ядра1 – ядерная оболочка (две
мембраны – внутренняя и
внешняя – и перинуклеарное
пространство),
2 – ядерная пора,
3 – хроматин
конденсированный
(гетерохроматин),
4 – хроматин диффузный
(эухроматин),
5 – ядрышко,
6 – рибосомы,
7 – фибриллы,
8 - кариоплазма
23.
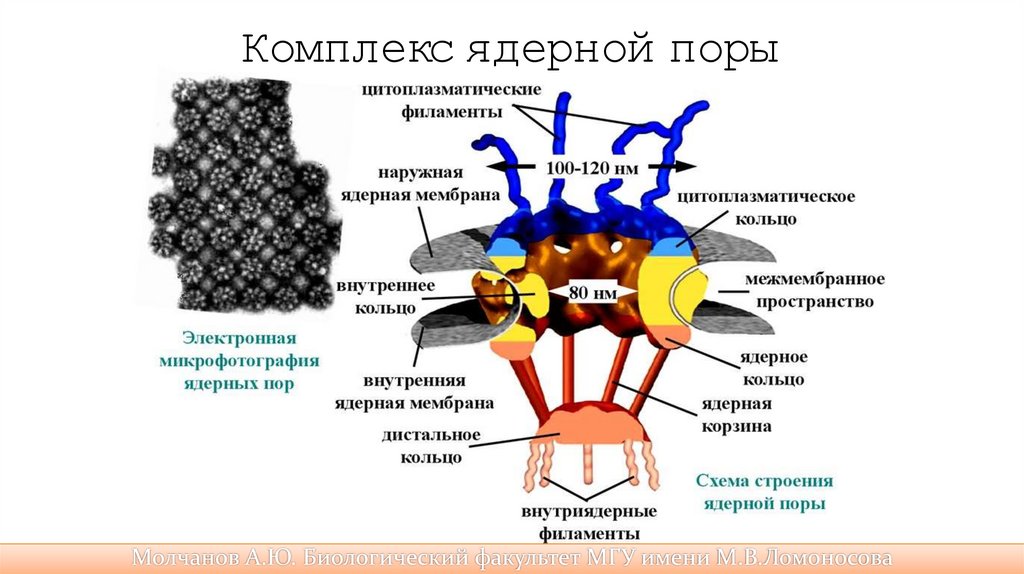
Строение ядра• Внутренняя поверхность ядерной оболочки подстилается ядерной
ламиной, жесткой белковой структурой, образованной белкамиламинами, к которой прикреплены нити хромосомной ДНК.
• В ядре есть ядрышко.
• Гистоны регулируют активность (экспрессию) генов.
• Поры пронизывают обе ядерные мембраны и формируют сквозной
канал, через который малые молекулы и ионы проходят свободно, а
крупные молекулы активно транспортируются с участием белковпереносчиков.
• Транспортом через ядерную пору занимаются белки импортины(в ядро)
и экспортины (из ядра).
24.
Ядрышко• Ядрышко — это отдельная плотная структура в ядре. Она не окружена
мембраной и формируется в области расположения рДНК. Они
представлены участками хромосом, имеющими ядрышковые организаторы
(13, 14, 15, 21, 22).
• На первой стадии образования рибосом фермент РНК-полимераза
I транскрибирует рДНК и образует пре-рРНК, которая далее разрезается
на 5,8S, 18S и 28S рРНК. Транскрипция и посттранскрипционный
процессинг рРНК происходят в ядрышке при участии малых ядрышковых
РНК.
• Сборка рибосом также происходит в ядре, в специальных образованиях,
называемых ядрышками. Открыты Барбарой Мак-Клинток.
• По структуре ядрышка можно судить о физиологической активности
клетки.
25.
Тельце Кахаля• Тельце Кахаля (ТК) — ядерное тельце, имеющееся у всех
эукариот. Оно идентифицируется по наличию сигнатурного
белка коилина и специфических РНК (scaРНК). В ТК также
содержится белок SMN (англ. survival of motor neurons). В ТК
наблюдается высокая концентрация сплайсирующих малых
ядерных рибонуклеопротеинов (мяРНП) и других факторов
процессинга РНК, поэтому считается, что ТК служат местами
сборки и/или посттранскрипционной
модификации факторов сплайсинга. ТК присутствует в ядре
во время интерфазы, но исчезает в митозе
26.
Функции ядра• В ядре происходит репликация — удвоение молекул ДНК,
а также транскрипция — синтез молекул РНК на матрице
ДНК.
• Синтез и сборка малой и большой субъединиц рибосом;
• Хранение наследственной информации;
• Реализация наследственной информации – образование
РНК.
• Регуляции жизнедеятельности клетки;
27.
Комплекс ядерной поры28.
29.
Ламины• Ламины - это фибриллярные белки, обеспечивающие
структурную функцию и регуляцию транскрипции в ядре клеток.
Ламины взаимодействуют с мембранными белками, чтобы
образовать ядерную ламину на внутренней стороне ядерной
оболочки. Они задействованы в дезинтеграции и образовании
оболочки ядра в ходе митоза, а также в позиционировании
ядерных пор.
• В процессе митоза ламины фосфорилируются митозстимулирующим фактором (МСФ), при этом происходит
дезинтеграция ламинов и оболочки ядра. После сегрегации
хромосом дефосфолирование ядерных ламинов обеспечивает
сборку ядерной оболочки.
30.
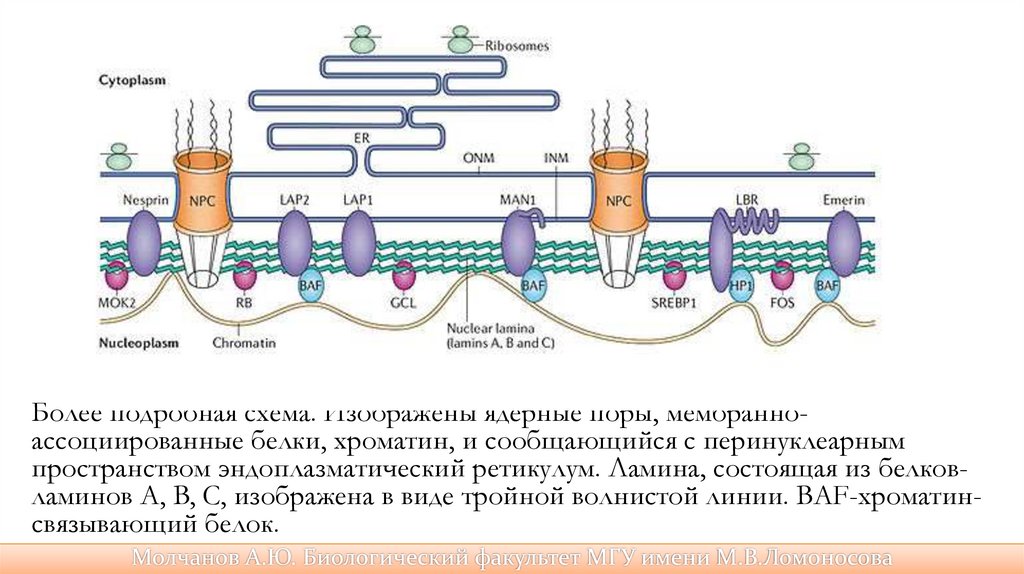
Более подробная схема. Изображены ядерные поры, мембранноассоциированные белки, хроматин, и сообщающийся с перинуклеарнымпространством эндоплазматический ретикулум. Ламина, состоящая из белковламинов A, B, C, изображена в виде тройной волнистой линии. BAF-хроматинсвязывающий белок.
31.
Цитоплазма32.
Строение клетки33.
Цитоплазма• Цитопла́зма
(от греч. κύτος «клетка» и πλάσμα зд.
«содержимое») — внутренняя среда живой клетки, кроме ядра,
ограниченная
плазматической
мембраной.
Термин
«цитоплазма» ввёл Эдуард Страсбургер в 1882 году.
Цитоплазма включает в себя
– Гиалоплазму – основное прозрачное вещество цитоплазмы;
– Органеллы – обязательные клеточные компоненты;
– Включения – также различные непостоянные структуры —.
34.
Органоиды эукариотическойклетки
Немембранные
Мембранные
1.
2.
3.
4.
1.
2.
Одномембранные
Эндоплазматическая сеть
Аппарат Гольджи
Лизосомы
Вакуоли
Двумембранные
Митохондрии
Пластиды
1.
2.
3.
4.
5.
Рибосомы
Микротрубочки
Жгутики
Реснички
Клеточный центр
35.
Мембранные органеллы• Общим свойством мембранных органелл является то, что все
они построены из липопротеидных пленок (биологических
мембран), замыкающихся сами на себя так, что образуются
замкнутые полости, или отсеки. Внутреннее содержимое этих
отсеков всегда отличается от гиалоплазмы
36.
Функции Цитоплазмы• Накопление веществ
• Обеспечение процессов энергетического и пластического
обмена
• передача сигнала от мембраны к ядру.
• тургор клетки.
• объединение всех клеточных структур (компонентов) и
обеспечение их химического взаимодействия
37.
МИТОХОНДРИЯ38.
Митохондрии• Митохондрии—неотъемлемые
компоненты
всех
эукариотических клеток. Они представляют собой гранулярные
или нитеподобные структуры толщиной 0,5 мкм и длиной до 7—
10 мкм.
• Митохондрии — особые органеллы клетки, основной
функцией которых является синтез АТФ — универсального
носителя энергии. Дыхание (поглощение кислорода и
выделение углекислого газа) происходит также за счёт
ферментативных систем митохондрий
39.
Функции митохондрий• Синтез АТФ
• Потребление кислорода
• Выделение углекислого газа
40.
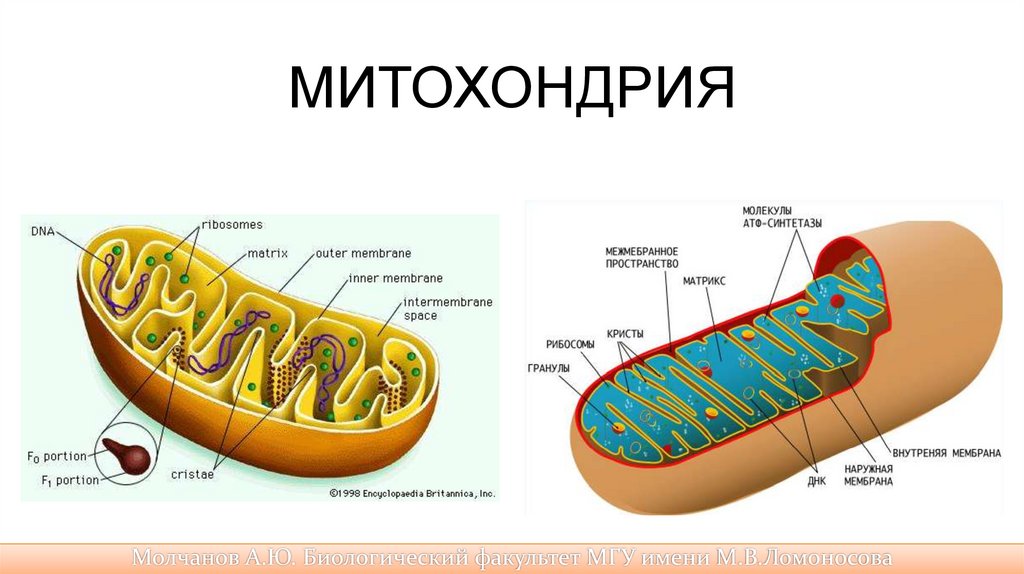
Мембраны митохондрии• Митохондрии имеют две мембраны — наружную и
внутреннюю. Между ними находится перимитохондриалъное
(периплазматическое) пространство, которое является местом
скопления ионов водорода Н+ .
– Наружная митохондриальная мембрана отделяет ее от гиалоплазмы
клетки.
– Внутренняя мембрана образует множество впячиваний внутрь
митохондрий — так называемых крист.
41.
Мембраны митохондрии• На мембране крист или внутри нее располагаются
ферменты, в том числе переносчики электронов и ионов
водорода Н+, которые участвуют в кислородном дыхании.
• Наружная
мембрана
отличается
высокой
проницаемостью, и многие соединения легко проходят
через нее.
• Внутренняя мембрана менее проницаема. Ограниченное
ею внутреннее содержимое митохондрии {матрикс) по
составу близко к цитоплазме. Матрикс содержит
различные белки, в том числе ферменты, ДНК (кольцевая
молекула), все типы РНК, аминокислоты, рибосомы, ряд
витаминов. ДНК обеспечивает некоторую генетическую
автономность митохондрий, хотя в целом их работа
координируется ДНК ядра.
42.
Кардиолипин• Кардиолипин «CL»— фосфолипид,
который является важным
компонентом внутренней мембраны
митохондрий. Молекула
дифосфатидилглицерол.
• Связывается с комплексом 4 дыхательной
цепи (цитохромоксидаза).
• Нужен для поддержания структуры
комплекса 3 (цитохром-bc1-комплекса).
Функционирует как протонная ловушка.
43.
ДНК митохондрии• Митохондрии
имеют
свой
собственный
ДНК-геном
и
прокариотические
рибосомы,
что,
безусловно,
указывает
на симбиотическое происхождение этих органелл.
• В ДНК митохондрий закодированы совсем не все митохондриальные
белки, большая часть генов митохондриальных белков находятся в ядерном
геноме, а соответствующие им продукты синтезируются в цитоплазме, а
затем транспортируются в митохондрии.
• Геномы митохондрий отличаются по размерам: например геном
человеческих митохондрий содержит всего 13 генов. Самое большое число
митохондриальных
генов
(97)
из
изученных
организмов
имеет простейшее Reclinomonas americana.
44.
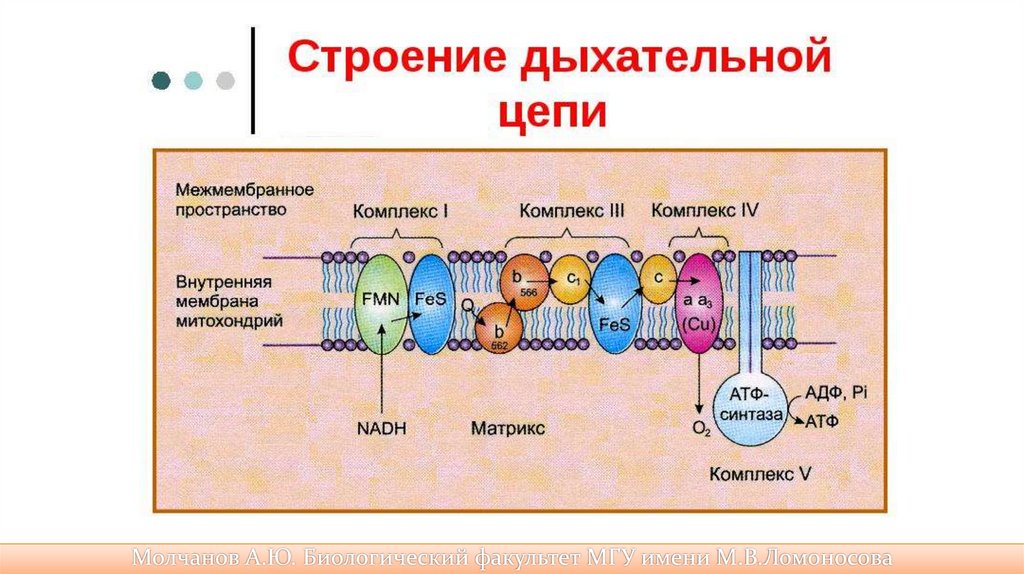
45.
ЭТЦ митохондрий• Включает четыре вида белковых комплекса;
• Один из них не переносит электронов/протонов
• Сукцинат-дегидрогиназа – не переносит электронов, также
участвует в цикле Кребса.
46.
Митохондриальные заболевания• Происходит нарушение тканевого дыхания. Передается только по женской
линии детям обоих полов . Дефект может быть связан с нарушением в цикле
Кребса, в дыхательной цепи, процессах бета-окисления и т. д.
• Мутации митохондриальных генов приводят к следующим заболеваниям
(синдром Барта, синдром Кернса-Сейра, синдром Пирсона, синдром
MELAS, синдром MERRF и другие).
• Вторичные митохондриальные заболевания, включающие нарушение
клеточного энергообмена как важное звено формирования патогенеза (болезни
соединительной ткани, синдром хронической
усталости, гликогеноз, кардиомиопатия, мигрень, печёночная
недостаточность, панцитопения, а также гипопаратиреоз, диабет, рахит и
другие).
47.
Пластиды• Пластиды —характерные органеллы клеток автотрофных эукариотических
организмов. Их окраска, форма и размеры весьма разнообразны.
• Различают хлоропласты, хромопласты и лейкопласты. Хлоропласты имеют
зеленый цвет, обусловленный присутствием основного пигмента — хлорофилла.
• Хлоропласты содержат также вспомогательные пигменты —
каротиноиды (оранжевого цвета).
• Наружная мембрана отграничивает жидкую внутреннюю гомогенную среду
хлоропласта — строму (матрикс). В строме содержатся белки, липиды, ДНК
(кольцевая молекула), РНК, рибосомы и запасные вещества (липиды, крахмальные
и белковые зерна) а также ферменты, участвующие в фиксации углекислого газа.
• Хлоропласты в клетке осуществляют процесс фотосинтеза.
48.
Строение хлоропластаХлоропласты ограничены двумя мембранами — наружной и внутренней
По форме хлоропласты — это овальные линзовидные тельца размером (5—10) х (2—4)
мкм. В одной клетке листа может находиться 15—20 и более хлоропластов, а у некоторых
водорослей — лишь 1 -2 гигантских хлоропласта (хроматофора) различной формы.
49.
Прохлорон (прохлорофиты)• Содержит, как эукариоты хлорофилл типа А и В,
поэтому они способны к оксигенному
фотосинтезу.
• Кислород также выделяют цианобактерии, но они
имею отличные пигменты. Фиксирует углерод в
пентозофосфатном цикле.
• В экосистеме является коменсалом по отношению
к асцидиям. Живет на поверхности и потребляет
аминокислоты.
• Их считают наиболее вероятными
предшественниками хлоропластов.
50.
Внутреннее строение хлоропласта• Внутренняя мембрана хлоропласта образует впячивания внутрь
стромы —тилакоиды, или ламеллы, которые имеют форму
уплощенных мешочков (цистерн).
• Несколько таких тилакоидов, лежащих друг над другом, образуют
грану, и в этом случае они называются тилакоидами граны.
• Именно в мембранах тилакоидов локализованы
светочувствительные пигменты, а также переносчики электронов и
протонов, которые участвуют в поглощении и преобразовании
энергии света.
• Пространство внутри тилакоидов называется люмен.
51.
Хлоропласт• Это цветная трансмиссионная
электронная микрофотография
(ТЭМ) двух хлоропластов,
видимых в листьях растения
гороха Pisum sativum.
• Свет и углекислый газ
превращаются в углеводы
хлоропластом. Большие участки
крахмала, образующиеся в
процессе фотосинтеза, видны
как темные круги внутри каждого
хлоропласта.
52.
Разнообразие пластид• Лейкопласты — мелкие бесцветные пластиды различной формы. Они бывают
шаровидными, эллипсоидными, гантелевидными, чашевидными и т. д. По
сравнению с хлоропластами у них слабо развита внутренняя мембранная система.
• Лейкопласты в основном встречаются в клетках органов, скрытых от
солнечного света (корней, корневищ, клубней, семян). Они осуществляют
вторичный синтез и накопление запасных питательных веществ — крахмала,
реже жиров и белков.
• Хромопласты отличаются от других пластид своеобразной формой
(дисковидной, зубчатой, серповидной, треугольной, ромбической и др.) и
окраской (оранжевые, желтые, красные). Хромопласты лишены хлорофилла и
поэтому не способны к фотосинтезу. Внутренняя мембранная структура их слабо
выражена.
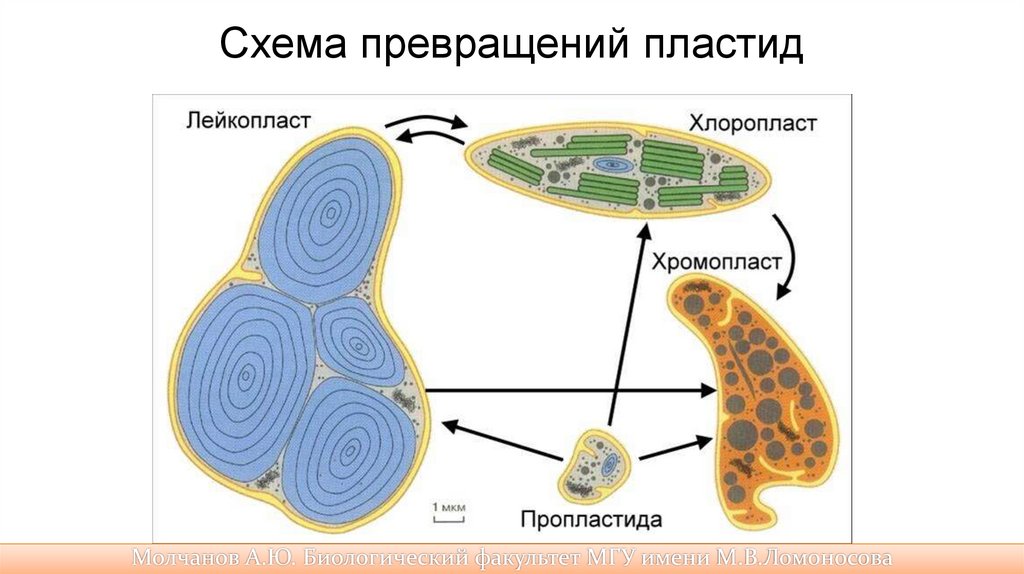
53.
Схема превращений пластид54.
Пластиды• Все типы пластид генетически родственны друг другу, и одни их
виды могут превращаться в другие:
• Таким образом, весь процесс взаимопревращений пластид можно
представить в виде ряда изменений, идущих в одном направлении
— от пропластид до хромопластов (хлоропласты, амилопласты,
лейкопласты).
• Лейкопласты образуются из протопластиды в том случае, если на
клетку не попадает ультрафиолет.
• Если в хлоропласте разрушается хлорофилл и накапливаются
другие пигменты, например каратиноиды, цианины и др., то он
необратимо изменяется в хромопласт (лепестки, осенние листья).
Хромопласты не фотосинтезируют.
55.
Разнообразие пластидАмилопласт
Элайопласт
Лейкопласт
Протиопласт
Пластида
Хромопласт
Этиопласт
Хлоропласт
56.
Несколько мембран• У высших растений хлоропласты
двумембранные.
• У эвглены трехмембранные хлоропласты (С).
• У бурых водорослей четырехмембранные
хлоропласты.
• У животного элизии изумрудной трех
мембранные хлоропласты.
57.
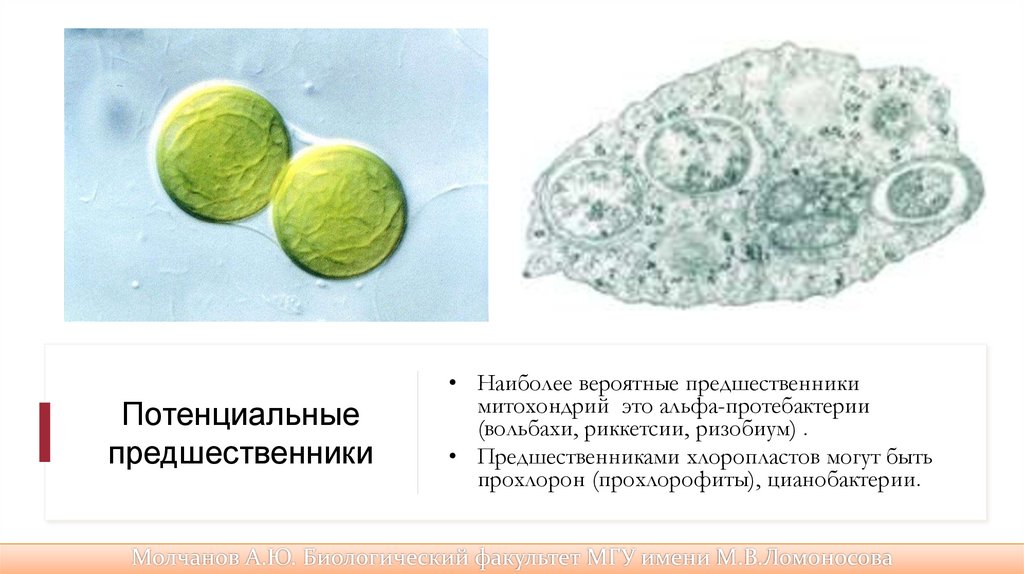
Потенциальныепредшественники
• Наиболее вероятные предшественники
митохондрий это альфа-протебактерии
(вольбахи, риккетсии, ризобиум) .
• Предшественниками хлоропластов могут быть
прохлорон (прохлорофиты), цианобактерии.
58.
Одномембранные органеллы.• В клетке синтезируется огромное количество различных веществ.
Часть из них потребляется на собственные нужды (синтез АТФ,
построение органелл, накопление питательных веществ), часть
выводится из клетки и используется на построение оболочки (клетки
растений и грибов), гликокаликса (животные клетки).
• Клеточными секретами являются также ферменты, гормоны,
коллаген, кератин и т. д. Накопление этих веществ и перемещение их
из одной части клетки в другую либо выведение за ее пределы
происходит в системе замкнутых цитоплазматических мембран —
эндоплазматической сети, или эндоплазматическом ретикулуме, и
комплексе Гольджи, составляющих транспортную систему клеток.
59.
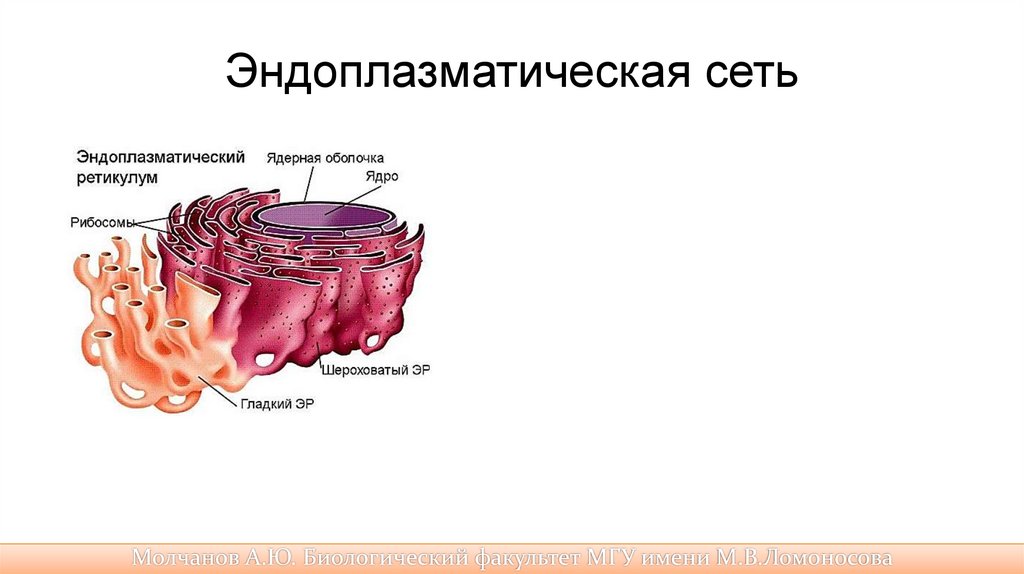
Эндоплазматическая сеть• была открыта с помощью электронного микроскопа в 1945 г. Она
представляет собой систему разветвленных каналов, цистерн (вакуолей),
пузырьков, создающих подобие рыхлой сети в цитоплазме. Стенки каналов
и полостей образованы элементарными мембранами.
• В клетке существует два типа эндоплазматического
ретикулума: гранулярный (шероховатый) и агранулярный (гладкий). Гранулярный
эндоплазматическая сеть (ретикулум) густо усеяна рибосомами, на которых
осуществляется биосинтез белка.
• Синтезируемые белки проходят через мембрану в каналы и полости
эндоплазматического ретикулума, изолируются от цитоплазмы,
накапливаются там, дозревают и перемещаются в другие части клетки либо
в комплекс Гольджи в специальных мембранных пузырьках, которые
отшнуровываются от цистерн эндоплазматической сети
60.
Функции ЭПС• В мембранах гранулярного эндоплазматического ретикулума накапливаются и
изолируются белки, которые после их синтеза могли оказаться вредными для клетки.
Например, синтез гидролитических ферментов и их свободный выход в цитоплазму привел
бы к самоперевариванию клетки и ее гибели. Однако этого не происходит, потому что
подобные белки надежно изолированы в полостях эндоплазматического ретикулума.
• На рибосомах гранулярной эндоплазматической сети синтезируются также
интегральные и периферические белки мембран клетки и некоторая часть белков
цитоплазмы.
• Цистерны шероховатого эндоплазматического ретикулума связаны с ядерной оболочкой,
причем некоторые из них являются прямым продолжением последней. Считается, что после
деления клетки оболочки новых ядер образуются из цистерн эндоплазматического
ретикулума.
• На мембранах гладкого эндоплазматического ретикулума протекают процессы синтеза
липидов и некоторых углеводов (например, гликогена).
61.
ЭПС• Является продолжением внешней ядерной мембраны.
Представлена большим количество плоских полостей
(цистерн). Жидкость может перетекать из одной
цистерны в другую. На концах полостей могут
формировать пузырьки с белками;
• Гладкая эпс
– Накопление кальция и сахаров;
– В ней происходит синтез жиров и гликогена;
• Шероховатая ЭПС
– Связана с рибосомами
– На ней происходит синтез белков
– Запускает микровизикуляторный транспорт
62.
В мышечных клетках• Мембрана мышечных волокон называется
саркоплазматический ретикулум. В нем накапливаются ионы
кальция.
63.
Эндоплазматическая сеть64.
Комплекс (аппарат) Гольджи• открыт в 1898 г. итальянским
ученым К. Гольджи. Он
представляет собой систему
плоских дисковидных замкнутых
цистерн, которые располагаются
одна над другой в виде стопки и
образуют диктиосому.
• От цистерн отходят во все
стороны мембранные трубочки и
пузырьки. Число диктиосом в
клетках варьирует от одной до
нескольких десятков в
зависимости от типа клеток и
фазы их развития.
65.
Диктиосомы ап. Гольджи• К комплексу Гольджи доставляются белки,
синтезируемые в ЭПС. От цистерн ЭПС
отшнуровываются пузырьки, которые
соединяются с цистернами комплекса
Гольджи, где эти вещества модифицируются
и дозревают.
• Пузырьки, выделяемые комплексом
Гольджи, участвуют в формировании
цитоплазматической мембраны и стенок
клеток растений после деления, а также в
образовании вакуолей и первичных
лизосом.
• Пузырьки заполненные секретом.
Содержимое таких пузырьков либо
используется самой клеткой, либо выводится
за ее пределы.
66.
Участие в секреции• Пузырьки Гольджи
подходят к
плазматической
мембране, соединяются
с ней и изливают свое
содержимое наружу, а
их мембрана
включается в
плазматическую
мембрану и таким
образом происходит ее
обновление.
67.
Диктиосомы ап. Гольджи• Цистерны комплекса Гольджи активно извлекают моносахариды из
цитоплазмы и синтезируют из них более сложные олиго- и полисахариды. У
растений в результате этого образуются пектиновые вещества, гемицеллюлоза
и целлюлоза, используемые для построения клеточной стенки, слизь корневого
чехлика. У животных подобным образом синтезируются гликопротеины и
гликолипиды гликокаликса, вырабатываются секрет поджелудочной железы,
амилаза слюны, пептидные гормоны гипофиза, коллаген.
• Комплекс Гольджи участвует в образовании лизосом, белков молока в
молочных железах, желчи в печени, веществ хрусталика, зубной эмали и г.
п.
• Комплекс Гольджи и эндоплазматический ретикулум тесно связаны между
собой; их совместная деятельность обеспечивает синтез и преобразование
веществ в клетке, их изоляцию, накопление и транспорт.
68.
Комплекс Гольджи69.
Аппарат Гольджи• Состоит из нескольких несвязанных полостей (диктиосом), от
которых тянутся отростки;
• К ап. Гольджи со стороны ядра подходят пузырьки от ЭПС;
1. В ап. Гольджи происходит созревание белков. По мере
продвижения с помощью визикулярного транспорта по
диктиосомам
белки
активируются
и
приобретают
четвертичную структуру;
2. Дистальные (со стороны плазматической мембраны)
диктиосомы отвечают за распределение готовых веществ
(в цитоплазму, в ядро, к мембране и для секреции);
3. Ап. Гольджи образует лизосомы.
70.
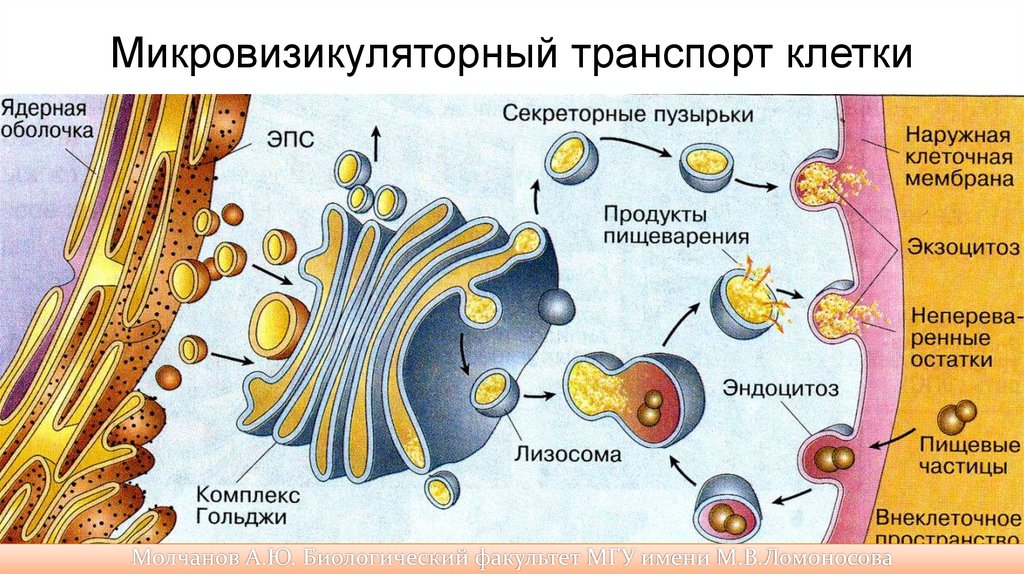
Микровизикуляторный транспорт клетки71.
Лизосомы• Лизосомы — это мембранные пузырьки величиной до 2 мкм. Внутри
лизосом содержатся гидролитические ферменты, способные переваривать
белки, липиды, углеводы, нуклеиновые кислоты. Лизосомы образуются из
пузырьков, отделяющихся от комплекса Гольджи, причем предварительно
на шероховатом эн до плазматическом ретикулуме синтезируются
гидролитические ферменты.
• Сливаясь
с
эндоцитозными
пузырьками,
лизосомы
образуют пищеварительную вакуоль (вторичная лизосома), где происходит
расщепление органических веществ до составляющих их мономеров.
Последние через мембрану пищеварительной вакуоли поступают в
цитоплазму клетки. Именно так происходит, например, обезвреживание
бактерий в клетках крови — нейтрофилах.
72.
Лизосомы• Вторичные лизосомы, в которых закончился процесс переваривания, практически не
содержат ферментов. В них находятся лишь непереваренные остатки, т. е. негидролизуемый
материал, который либо выводится за пределы клетки, либо накапливается в цитоплазме.
• Расщепление лизосомами чужеродного, поступившего путем эндоцитоза материала
называется гетерофагией. Лизосомы участвуют также в разрушении материалов клетки,
например запасных питательных веществ, а также макромолекул и целых органелл,
утративших функциональную активность (аутофагия).
• При патологических изменениях в клетке или ее старении мембраны лизосом могут
разрушаться: ферменты выходят в цитоплазму, и осуществляется самопереваривание
клетки —автолиз.
• Иногда с помощью лизосом уничтожаются целые комплексы клеток и органы.
Например, когда головастик превращается в лягушку, лизосомы, находящиеся в клетках
хвоста, переваривают его: хвост исчезает, а образовавшиеся во время этого процесса
вещества всасываются и используются другими клетками тела
73.
Вакуоли• Вакуоли — крупные мембранные пузырьки или полости в
цитоплазме, заполненные клеточным соком.
• Вакуоли образуются в клетках растений и грибов из
пузыревидных расширений эндоплазматического ретикулума
или из пузырьков комплекса Гольджи.
• В меристематических клетках растений вначале возникает
много мелких вакуолей. Увеличиваясь, они сливаются
в центральную вакуоль, которая занимает до 70—90% объема
клетки и может быть пронизана тяжами цитоплазмы
74.
Вакуоли• Содержимое вакуолей —клеточный сок. Он представляет собой водный
раствор различных неорганических и органических веществ.
• Большинство из них являются продуктами метаболизма протопласта,
которые могут появляться и исчезать в различные периоды жизни клетки.
Химический состав и концентрация клеточного сока очень изменчивы и
зависят от вида растений, органа, ткани и состояния клетки.
• В клеточном соке содержатся соли, сахара (прежде всего сахароза,
глюкоза, фруктоза), органические кислоты (яблочная, лимонная, щавелевая,
уксусная и др.), аминокислоты, белки. Эти вещества являются
промежуточными продуктами метаболизма, временно выведенными из
обмена веществ клетки в вакуоль. Они являются запасными веществами
клетки
75.
Вакуоли• Помимо запасных веществ, которые могут вторично
использоваться в метаболизме, клеточный сок содержит
фенолы, танины (дубильные вещества), алкалоиды,
антоцианы, которые выводятся из обмена в вакуоль и таким
путем изолируются от цитоплазмы.
• Танины особенно часто встречаются в клеточном соке (а
также в цитоплазме и оболочках) клеток листьев, коры,
древесины, незрелых плодов и семенных оболочек.
Алкалоиды присутствуют, например, в семенах кофе
(кофеин), плодах мака (морфин) и белены (атропин), стеблях
и листьях люпина (люпинин) и др. Считается, что танины с
их вяжущим вкусом, алкалоиды и токсичные полифенолы
выполняют защитную функцию: их ядовитый (чаще горький)
вкус и неприятный запах отталкивают растительноядных
животных, что предотвращает поедание этих растений
76.
Вакуоли• В вакуолях также часто накапливаются конечные продукты
жизнедеятельности клеток (отходы). Таким веществом для клеток
растений является щавелевокислый кальций (оксалат), который
откладывается в вакуолях в виде кристаллов (друз) различной формы.
• В клеточном соке многих растений содержатся пигменты (меланин,
каратиноид), придающие клеточному соку разнообразную окраску.
Пигменты и определяют окраску венчиков цветков, плодов, почек и
листьев, а также корнеплодов некоторых растений (например, свеклы).
• Клеточный сок некоторых растений содержит физиологически
активные вещества — фитогормоны (регуляторы роста), фитонциды,
ферменты. В последнем случае вакуоли действуют как лизосомы. После
гибели клетки мембрана вакуоли теряет избирательную проницаемость,
и ферменты, высвобождаясь из нее, вызывают автолиз клетки.
77.
Функции вакуолей• Вакуоли играют главную роль в поглощении воды растительными клетками.
Вода путем осмоса через ее мембрану поступает в вакуоль, клеточный сок
которой является более концентрированным, чем цитоплазма, и оказывает
давление на цитоплазму, а следовательно, и на оболочку клетки. В результате в
клетке развивается тургорное давление, определяющее относительную
жесткость растительных клеток и обусловливающее растяжение клеток во
время их роста.
• В запасающих тканях растений вместо одной центральной часто бывает
несколько вакуолей, в которых скапливаются запасные питательные вещества
(жиры, белки). Сократительные (пульсирующие) вакуолислужат для осмотической
регуляции, прежде всего, у пресноводных простейших, так как в их клетки
путем осмоса непрерывно поступает вода из окружающего гипотонического
раствора (концентрация веществ в речной или озерной воде значительно
ниже, чем концентрация веществ в клетках простейших). Сократительные
вакуоли поглощают избыток воды и затем выводят ее наружу путем
сокращений.
78.
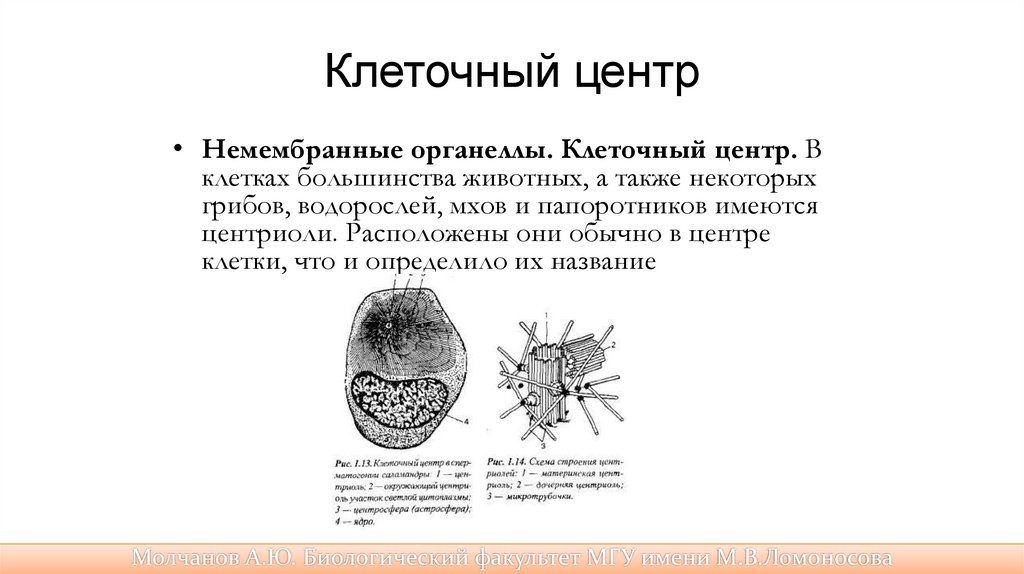
Клеточный центр• Немембранные органеллы. Клеточный центр. В
клетках большинства животных, а также некоторых
грибов, водорослей, мхов и папоротников имеются
центриоли. Расположены они обычно в центре
клетки, что и определило их название
79.
Немембранные органеллы• Центриоли представляют собой полые цилиндры
длиной не более 0,5 мкм. Они располагаются парами
перпендикулярно одна к другой. Каждая центриоль
построена из девяти триплетов микротрубочек.
• Основная функция центриолей — организация
микротрубочек веретена деления клетки.
• Центриолям по структуре идентичны базальные
тельца, которые всегда обнаруживаются в основании
жгутиков и ресничек. По всей вероятности, базальные
тельца образуются путем удвоения цен-триолей.
Базальные тельца, как и центриоли, являются центрами
организации микротрубочек, входящих в состав жгутиков
и ресничек.
80.
Жгутики• Жгутики и реснички — органеллы движения у клеток многих видов
живых существ. Они представляют собой подвижные цитоплазм
этические отростки, служащие либо для передвижения всего организма
(многие бактерии, простейшие, ресничные черви) или репродуктивных
клеток (сперматозоидов, зооспор), либо для транспорта частиц и
жидкостей (например, реснички мерцательных клеток слизистой
оболочки носовых полостей и трахеи, яйцеводов и т. д.).
• Жгутики эукариотических клеток по всей длине содержат 20
микротрубочек: 9 периферических дуплетов и 2 центральные
одиночные. У основания жгутика в цитоплазме располагается базальное тельце.
• Жгутики имеют длину около 100 мкм и более. Короткие жгутики
(10—20 мкм), которых бывает много на одной клетке,
называются ресничками.
• Скольжение микротрубочек, входящих в состав жгутиков или
ресничек, вызывает их биение, что обеспечивает перемещение клетки
либо продвижение частиц.
81.
Рибосомы• Рибосомы — это мельчайшие сферические гранулы диаметром 15—
35 нм, являющиеся местом синтеза белка из аминокислот. Они
обнаружены в клетках всех организмов, в том числе прокариотических.
Рибосомы представлены в клетке огромным числом: за клеточный
цикл их образуется около 10 млн. штук.
• В состав рибосом входит множество молекул различных белков и
несколько молекул рРНК. Рибосома состоит из малой и большой
субъединиц.
• Большая субьединица похожа на полусферу с тремя торчащими
выступами. Большая субъединица имеет показатель седиментации 45S. В
состав большой субъединицы входит три молекулы рРНК 5,8S, 18S, 28S.
• Малая субъединица имеет палочковидную форму с несколькими
выступами. В состав малой субъединицы входит одна молекула РНК.
• При объединении в рибосому малая субъединица ложится одним
концом на один из выступов большой субъединицы.
82.
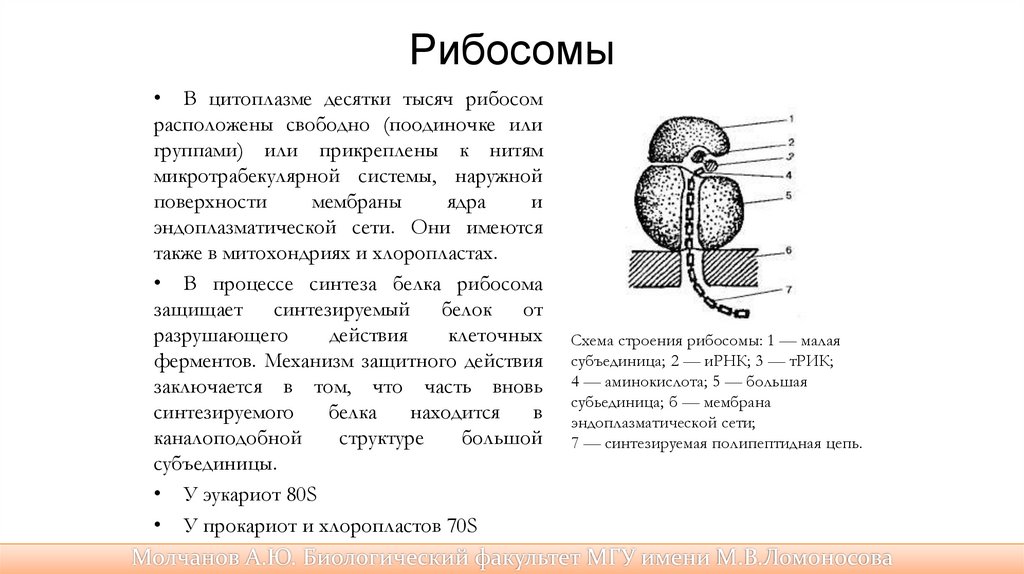
Рибосомы• В цитоплазме десятки тысяч рибосом
расположены свободно (поодиночке или
группами) или прикреплены к нитям
микротрабекулярной системы, наружной
поверхности
мембраны
ядра
и
эндоплазматической сети. Они имеются
также в митохондриях и хлоропластах.
• В процессе синтеза белка рибосома
защищает синтезируемый белок от
разрушающего
действия
клеточных
ферментов. Механизм защитного действия
заключается в том, что часть вновь
синтезируемого
белка
находится
в
каналоподобной
структуре
большой
субъединицы.
• У эукариот 80S
• У прокариот и хлоропластов 70S
• У митохондрий 55S
Схема строения рибосомы: 1 — малая
субъединица; 2 — иРНК; 3 — тРИК;
4 — аминокислота; 5 — большая
субьединица; б — мембрана
эндоплазматической сети;
7 — синтезируемая полипептидная цепь.
83.
ЦИТОСКЕЛЕТ84.
Цитоскелет• Актиновые филаменты. Микрофиламенты представляют собой
две цепочки из мономеров актина, закрученные спиралью (диаметр 7
нм). Они сконцентрированы у внешней мембраны клетки, так как
отвечают за форму клетки и способны образовывать выступы на
поверхности клетки (псевдоподии и микроворсинки). Также они
участвуют в межклеточном взаимодействии (образовании адгезивных
контактов), передаче сигналов и, вместе с миозином — в мышечном
сокращении. С помощью цитоплазматических миозинов по
микрофиламентам может осуществляться везикулярный транспорт.
85.
Цитоскелет• Микротрубочки
представляют собой полые
цилиндры порядка 25 нм диаметром, стенки которых
составлены из 13 глобул гетеродимера тубулина (альфа и
бета). Микротрубочки — крайне динамичные структуры,
потребляющие ГТФ в процессе полимеризации. Они
играют ключевую роль во внутриклеточном транспорте
(служат
«рельсами», по
которым перемещаются
молекулярные моторы — кинезин и динеин).
86.
Промежуточные филаменты• Диаметр промежуточных филаментов составляет от 8 до 11
нанометров. Они состоят из разного рода субъединиц и
являются наименее динамичной частью цитоскелета.
• Классификация:
• Тип I — кератины;
• Тип II - десмин и виментин;
• Тип III – нестин и интернексин:
• Тип IV – белки ядерной ламины.
87.
Кератины• Это фибриллярные белки, обладающие высокой
механической прочностью.
• Выделяют две основные группы — эпителиальные
кератины (цитокератин), включающую около 20 видов
кератинов, и кератины волос (примерно 10 видов), из
которых построены также ногти, рога и чешуя
пресмыкающихся, перья и рамфотека клюва птиц.
• По вторичной структуре различают альфа- и бетакератины.
• Прочность белка альфа-кератина обусловлена высокой
концентрацией цистеина, который обеспечивает
образование дисульфидных мостиков между соседними
полипептидными цепями.
https://ru.wikipedia.org/wiki/Кератины
88.
Десмин• белок промежуточных филаментов, находящихся рядом с Zлинией в саркомерах. Десмин нужен для интеграции белков актина
и тропонина. Содержитсятолько в клетках позвоночных.
• Саркомеры становятся неровными, что приводит к
рассогласованию работы мышечных волокон. Эта мутация
приводит также к гибели мышечных клеток
путём апоптоза и некроза. Миопатия, связанная с
десмином (МСД, или десминопатия), является разновидностью
миофибриллярных миопатий.
89.
Виментин• Виментин — белок промежуточных
филаментов соединительных тканей и других тканей
мезодермального происхождения. Промежуточные
филаменты присутствуют во всех клетках животных и
в бактериях, и наряду с микротрубочками и актином
участвуют в
• Виментин прикрепляется
к ядру, эндоплазматическому ретикулуму (ЭПР)
и митохондриям. Виментин играет значительную роль в
закреплении органелл и поддержании их положения в
цитоплазме построении цитоскелета. Именно виментин
обеспечивает прочность клеток и их устойчивость к
90.
Моторные белкиК моторным белка клетки относятся миозины,
кинезины, динеины. Они участвуют во
внутриклеточном транспорте.
Миозины – находятся мышцах (саркомеры). С их
помощью мышцы сокращаются.
Динеины и кинезины траспортируют вещества в
цитоплазме.
Кинезины - суперсемейство моторных
белков эукариотических клеток, это тубулинзависимые АТФазы. Кинезины участвуют в
осуществлении различных клеточных функций и
процессов, включая митоз, мейоз и везикулярный
транспорт — транспорт мембранных пузырьков с грузом
(карго), в том числе быстрый аксональный транспорт.
91.
СтроениеФорма молекул белков этого семейства может быть разной, но
типичный кинезин — димер, каждая половина которого состоит
из тяжёлой и лёгкой цепи. Тяжёлая цепь образует глобулярную
«головку» (двигательный домен) и соединена с помощью
короткого шарнирного участка со «стеблем». «Стебли» двух
тяжёлых цепей спирально закручены друг относительно друга и
заканчиваются «хвостами», которые состоят из лёгких цепей. Груз
прикрепляется к хвостовому участку, структура которого
определяет тип переносимого груза.
92.
Процесс транспортировкиЭто сложный скоординированный процесс, поскольку
изначально что-то должно получить доступ к библиотеке ДНК
данного организма, распечатать информацию в точно
определенном месте, необходимом для определенной информации
(именно для того вещества, для которого она была произведена),
создать копию информации для данного вещества и доставить его
на «фабрику».
Затем другой клеточный органоид (под названием комплекс
Гольджи) запаковывает необходимое вещество, заворачивая его в
специальный пакет (который называется везикулой) и наносит
«адрес» того места в клетке, куда вещество должно быть доставлено
и распаковано из посылки-везикулы.
Затем кинезин отправляется в путь. Он поднимает посылку и
«идет» по микротрубочкам внутри клетки, доставляя посылку к
месту назначения.
93.
Двигательный доменГлобулярная «головка» всех
кинезинов имеет
консервативную
аминокислотную
последовательность. На ней
есть два центра связывания —
один для тубулина
микротрубочки, а второй для
АТФ.
Связывание и гидролиз АТФ (так же как и высвобождение
АДФ) приводит к изменениям конформации сайта связывания
тубулина и положения шарнирных участков по отношению к
«головкам», что приводит к передвижению кинезина вдоль
микротрубочки.
Несколько
других
доменов
головки
обеспечивают взаимодействие между двумя сайтами связывания и
между ними и шарнирными участками. Кинезин имеет
структурное сходство с G-белками (которые гидролизуют ГТФ).
94.
Направление и скорость движенияКинезины двигаются от минус-концов микротрубочек к плюс-концам, обычно
перемещая груз от центра клетки к периферии. В аксонах такой транспорт называется
антероградным. Транспорт в противоположном направлении осуществляют белки
другого семейства — динеины (в аксонах это ретроградный транспорт). Скорость
антероградного транспорта достигает 500 мм в сутки, скорость ретроградного
транспорта обычно вдвое ниже. Максимальная скорость скольжения микротрубочек
друг по другу, которую могут развивать динеины, тем не менее, намного выше: она
составляет для динеинов около 14 мкм/с, а для кинезинов только около 2-3 мкм/с.
95.
Кинезин и митозВ последние годы выяснилось, что сборка и разборка микротрубочек, в том числе
при формировании веретена деления, происходит при участии различных моторных
белков, в том числе кинезинов. Вероятно, эти белки участвуют также в анафазе II,
когда центриоли удаляются друг от друга за счёт скольжения цитоплазматических
микротрубочек веретена деления.
96.
Оглавление• Общие характеристики клетки эукариот
• Сравнение клеток прокариот и эукариот
• Строение
– Мембраны
– Ядра
– Цитоплазмы
• Физиологические особенности