










 цепь ДНК выступает в качестве матрицы для синтеза дочерней цепи")










































")







 Биология
БиологияПохожие презентации:





")




Реализация наследственной информации. Репликация, транскрипция, трансляция, генетический код
1. Тема лекции:
Реализация наследственнойинформации. Репликация,
транскрипция, трансляция,
генетический код.
Недорепликация концов
линейных молекул ДНК.
Теломеры, теломеразы
2. 2. Цель:
Сформироватьу
студентов
современные представления и знания о
направлении и механизмах передачи
наследственной информации в живых
системах, их роли и значении в
функционировании живых организмов в
нормальных и патологических условиях.
3. План лекции:
1. Центральная догма молекулярной биологии (основнойпостулат Крика). Типы переноса генетической информации в
живых системах: общий, специализированный,
запрещенный.
2. Репликация, определение, принципы.
3. Основные ферменты, участвующие в репликации и их
функции.
4. Проблема концевой недорепликации и ее решение.
5.Транскрипция. Механизмы транскрипции у про- и
эукариот.
6. Процессинг и сплайсинг. Альтернативный сплайсинг.
7. Трансляция. Механизмы трансляции
8. Особенности биосинтеза белков у про- и эукариот
9. Генетический код, определение, свойства.
4. Центральная догма
РепликацияТранскрипция
ДНК
Обратная
транскрипция
Трансляция
РНК
белок
Репликация
РНК
Ретро-РНК-вирусы
Только РНК-вирусы
Другие организмы тоже получили от
них этот фермент и используют в
некоторых случаях
5. Матричные синтезы, разрешенные по центральной догме
РНКДНК
Не обнаружен
белок
6. Запрещенные матричные синтезы
РНКДНК
белок
Белки никогда не бывают матрицами
7.
8.
Универсальный биологическийпроцесс передачи генетической
информации в поколениях клеток и
организмов, благодаря созданию
точных копий ДНК.
ДНК – единственная молекула
клетки, способная к самоудвоению.
9. Скорость репликации ДНК
У прокариот – 1000 нуклеотидов /секУ эукариот – 100 нуклеотидов /сек
(медленнее, потому что ДНК сложно упакована –
нуклеосомы и другие уровни упаковки)
10. Место репликации в клеточном цикле
Репликация ДНК всегда предшествуетделению клетки.
Интерфаза
Деление
Репликация
S-период
(Synthesis)
Каждая дочерняя клетка получает точную копию всей ДНК
11. Принципы репликации
1. Полуконсервативность2. Комплементарность
3. Антипараллельность
4. Униполярность
5. Прерывистость
12. Полуконсервативность – каждая исходная (материнская) цепь ДНК выступает в качестве матрицы для синтеза дочерней цепи
Старые цепочки ДНКВновь синтезированные
Полуконсервативный
Консервативный
Дисперсионный
13. Комплементарность
Вновь синтезируемая ( дочерняя) цепь ДНКстроится по принципу комплементарности. В состав
растущей цепи включается тот нуклеотид , который
комплементарен нуклеотиду родительской цепи
(аденин с тимином, гуанин с цитозином).
14. Антипараллельность – синтез дочерней цепи ДНК происходит в противоположном от материнской цепи направлении
15. Униполярность:
Удвоение цепи ДНК идет в направлении от 5` концак 3` концу, следовательно новый нуклеотид
присоединяется к 3 ` концу растущей цепи.
5'
3'
16. Прерывистость репликации
Репликация может идти одновременно в нескольких местахмолекулы ДНК.
ori
ori
ДНК одной
хромосомы
Репликон
Репликон – расстояние между двумя сайтами
начала репликации ori ~ 100 тыс. н.п.
У прокариот вся кольцевая молекула –
один репликон
17. Прерывистость репликации
oriori
ДНК одной
хромосомы
Репликативные вилки
18. Репликация ДНК
19. Репликативная вилка
Направление движения вилки3'
3'
Лидирующая
цепь
5'
3'
Запаздывающая
цепь
Фрагменты Оказаки
20. ДНК ГеликазА – Фермент разделяющий цепи двухцепочечной ДНК на одинарные ДНК ТОПОИЗОМЕРАЗА – фермент, изменяющий степень сверхспиральнос
Основные ферменты репликацииДНК ГЕЛИКАЗА – Фермент разделяющий цепи
двухцепочечной ДНК на одинарные
ДНК ТОПОИЗОМЕРАЗА – фермент, изменяющий степень
сверхспиральности, возникающее при раскручивании двух
цепей в репликативной вилке
ПРАЙМАЗА – фермент, обладающий РНК –
полимеразной активностью; служит для образования РНКпраймеров, необходимых для инициации синтеза ДНК
ДНК ПОЛИМЕРАЗА – синтезирует новую цепь ДНК по
принципу комплементарности
ДНК ЛИГАЗА – фермент, образующий фосфодиэфирную
связь между двумя полинуклеотидами
SSB (single-strand binding protein)-белки –
связывающиеся с одноцепочечными нитями ДНК и
предотвращают комплементарное спаривание
21. Свойства ДНК-полимеразы
1. Присоединяет по одномунуклеотиду с 3‘ конца
растущей цепочки.
3'
2. Требует для начала работы
спаренного 3‘ конца.
3. Отщепляет один нуклеотид
назад, если он не спарен – т.е.
исправляет свои ошибки.
Логически
связанные
свойства !
22. ДНК-полимераза исправляет ошибки
Если новый нуклеотид не спарен – фермент не можетдвигаться дальше.
Тогда он выедает неверный
нуклеотид и ставит другой.
23. Выводы по репликации ДНК
В результате репликации каждаядочерняя клетка получает точную
копию всей ДНК содержавшейся в
материнской клетке.
ДНК всех клеток одного
организма – одинаковая, как по
количеству молекул, т.е. хромосом, так и
по их нуклеотидному составу.
24. Проблема укорочения концов у линейных ДНК
Сформулирована – А.М. Оловников, 1971При каждой репликации новые цепи должны
укорачиваться с 5‘ концов
Почему? – Там выедается РНК-затравка, а
достроить брешь ДНК-полимераза не может –
нет спаренного конца.
При каждом делении хромосома теряет 50 н.п.
на концах – теломерах.
25. Гипотеза Оловникова
Укорочение концов – это внутренниечасы, отмеряющие время жизни
многоклеточного организма – число
отпущенных ему делений, начиная с
зиготы.
Как только теломеры «закончатся» –
клетка больше не делится и погибает.
26. Но почему тогда клетки зародышевой линии делятся бесконечно?
Оловников: должен существовать механизмудлинения концов хромосом.
Теломераза – фермент, надстраивающий
концы хромосом, содержит РНК длиной 150
нуклеотидов и осуществляет обратную
транскрипцию
Теломераза и обратная транскриптаза –
родственные белки, гомологичные по
структуре и топологии.
27. Теломераза
фермент, надстраивающий концыхромосом, содержит РНК.
удлинение происходит путем
обратной транскрипции:
РНК → ДНК
На концах хромосом находятся длинные
некодирующие повторы 5’ – ГГТ ТАГ – 3’
10-15 тысяч н.п. у человека
28. Репликация теломерной ДНК
29.
Теломераза активна в клеткахзародышевого пути
эмбриональных
стволовых
раковых – поэтому они бессмертны
Теломераза неактивна
в соматических клетках – ген для нее
там, конечно же, есть, но выключен
30.
Значение для медициныИногда в растующую цепь случайно вклинивается
неправильное основание, однако у здоровых клеток
присутствует
пострепликационные
репаративные
ферменты, которые исправляют подобные ошибки.
Патология
пострепликационных
механизмов
репарации
иногда
обусловливает
предрасположенность
пациентов
к
некоторым
онкологическим заболеваниям.
К ним относятся:
31.
Синдроммножественной
(синдром Блума);
Наследственная
ломкости
хромосом
предрасположенность
к
молочной железы, вызванную мутациями
BRCA1 и BRCA2;
Аутосомно-доминантная
раку
генов
форма рака кишечника
(наследственный неполипозный рак толстой кишки)
32.
Значение для медициныПосле
каждого
клеточного
цикла
теломеры укорачиваются на один
повтор, а следовательно, количество
делений клетки ограничено числом
повторов в теломерной цепи. Согласно
этому бесконечный рост и деление
опухолевых клеток происходят из-за
присутствия
активных
мутантных
теломераз,
которые
препятствуют
разрушению теломер
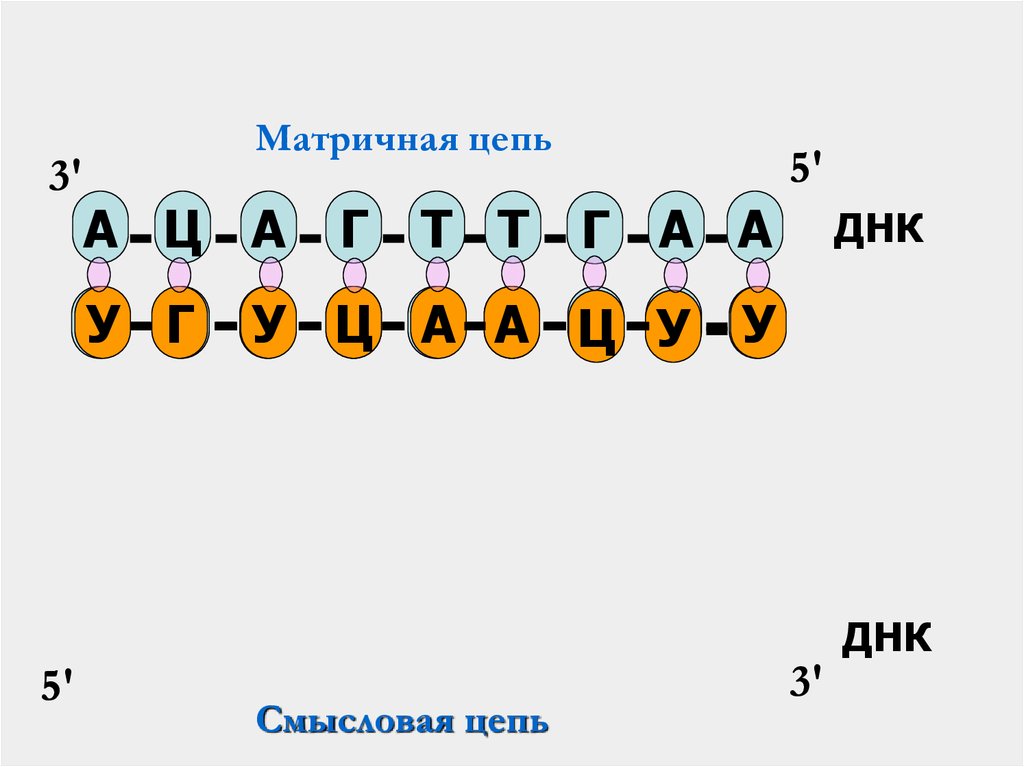
33. Транскрипция
Транскрипция- это первый этап реализации
генетической информации, при котором в
клетках осуществляется биосинтез РНК на
матрице
ДНК,
т.е.
переписывание
информации о структуре белка с ДНК на
специальный посредник – м РНК.
34.
3'Матричная цепь
А Ц А
Т
У Г
5'
Г
Т Т
5'
Г А А
ДНК
У
Т Ц А А Ц У
Т Т
У
Смысловая цепь
3'
ДНК
35.
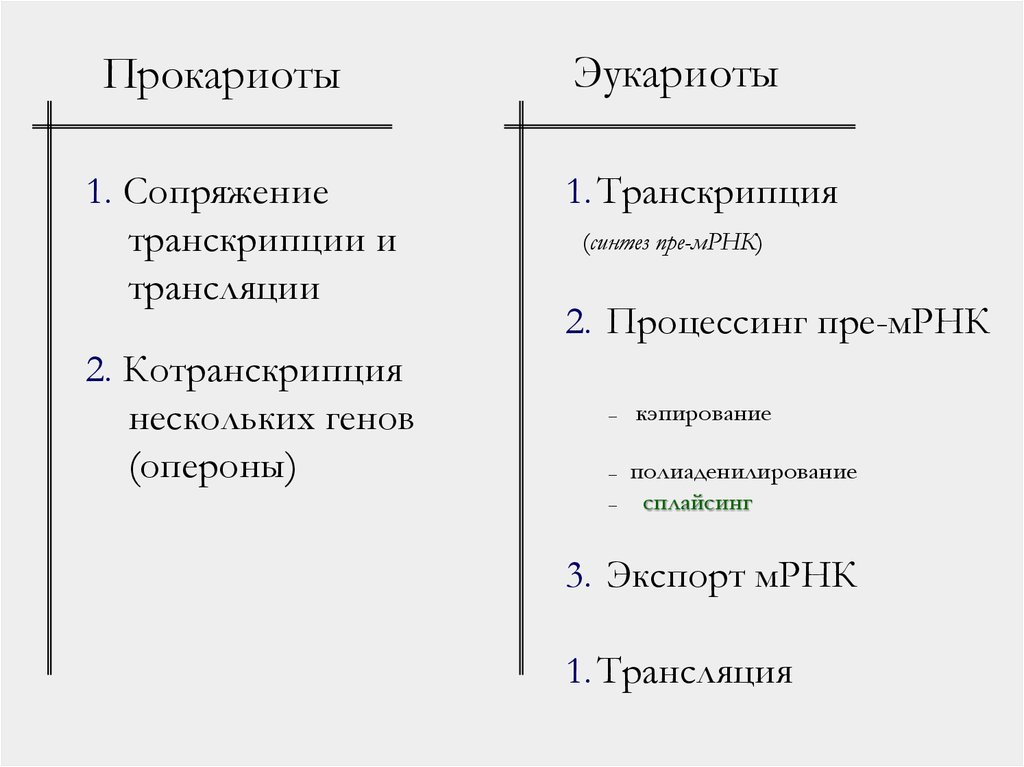
Прокариоты1. Сопряжение
транскрипции и
трансляции
2. Котранскрипция
нескольких генов
(опероны)
Эукариоты
1. Транскрипция
(синтез пре-мРНК)
2. Процессинг пре-мРНК
–
–
–
кэпирование
полиаденилирование
сплайсинг
3. Экспорт мРНК
1. Трансляция
36. РНК-полимераза
Кор ферментα
Холо фермент
ά
α
ω
ά
ω
β
βІ
β
δ
βІ
37. Гены – транскрибируемые участки ДНК
Транскрибируется не вся ДНК, алишь отдельные ее участки – гены.
ДНК одной
хромосомы
РНК
Некодирующая ДНК между генами
38.
39. РНК-полимераза движется по гену
2. Элонгация (рост цепочки РНК)РНК-полимераза движется по гену
40.
3. Терминациянаправление транскрипции
Терминатор (знак конца
транскрипции)
В области терминатора
находится инвертированный
повтор, который приводит к
образованию петли на РНК
41. Общие параметры транскрипции
Скорость – около 30 нуклеотидов / секЧастота ошибок – 1 на 104 нуклеотидов,
т.е. на пять порядков выше, чем при
репликации.
Синтез РНК – гораздо менее точный
процесс, чем синтез ДНК.
42.
ДеградацияТранспорт
ДНК
Транскрипция
Модификация
белка
РНК
Созревание РНК
м-РНК
Ядро
Экспорт м-РНК в
цитоплазму
Трансляция
43. Особенности транскрипции у эукариот
Транскрипцияядро
Созревание м-РНК
1. Кэп и поли-Ахвост
2.Сплайсинг
цитоплазма
44. Матричная РНК
Лидернаяпоследовательность
Прокариота
5'
3'
Старт-кодон
Стоп-кодон
Кодирующая часть
5'
Эукариота
Кэп –
модифицированные
нуклеотиды
Выполняет роль лидерной
последовательности
Созревание
3'
Поли-А-хвост
30 – 300 А
45.
ТерминаторПромотор
ДНК
одного
гена
П
Э
И
Э
И
Э
И
Э
Т
Транскрипция
м-РНК
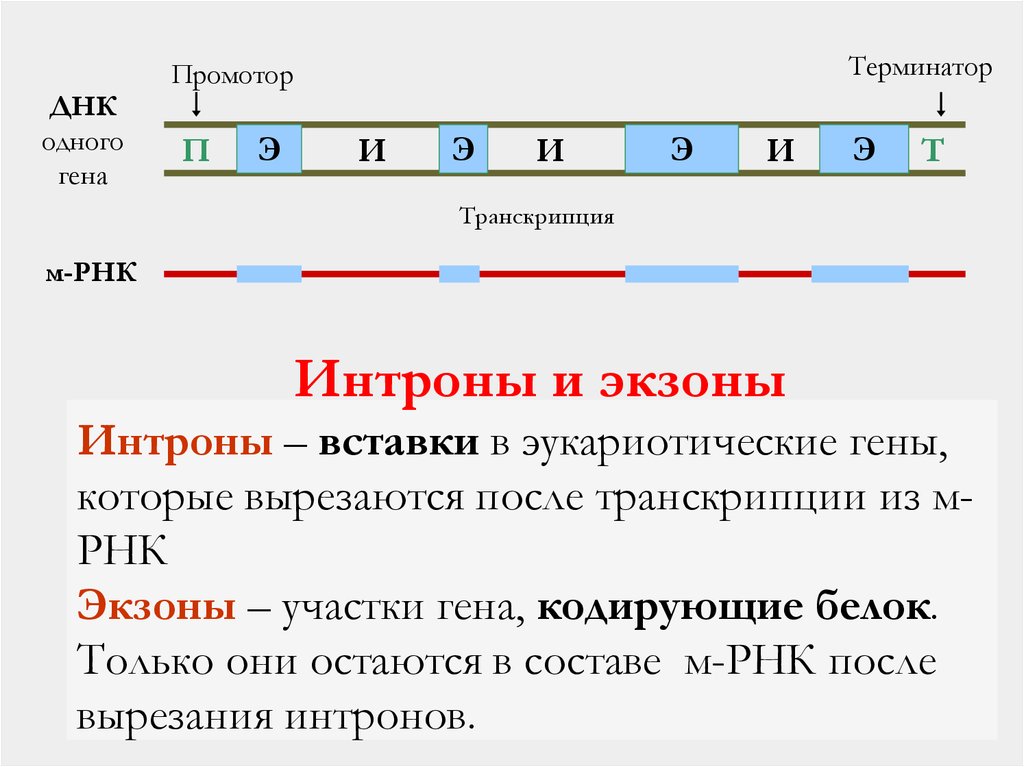
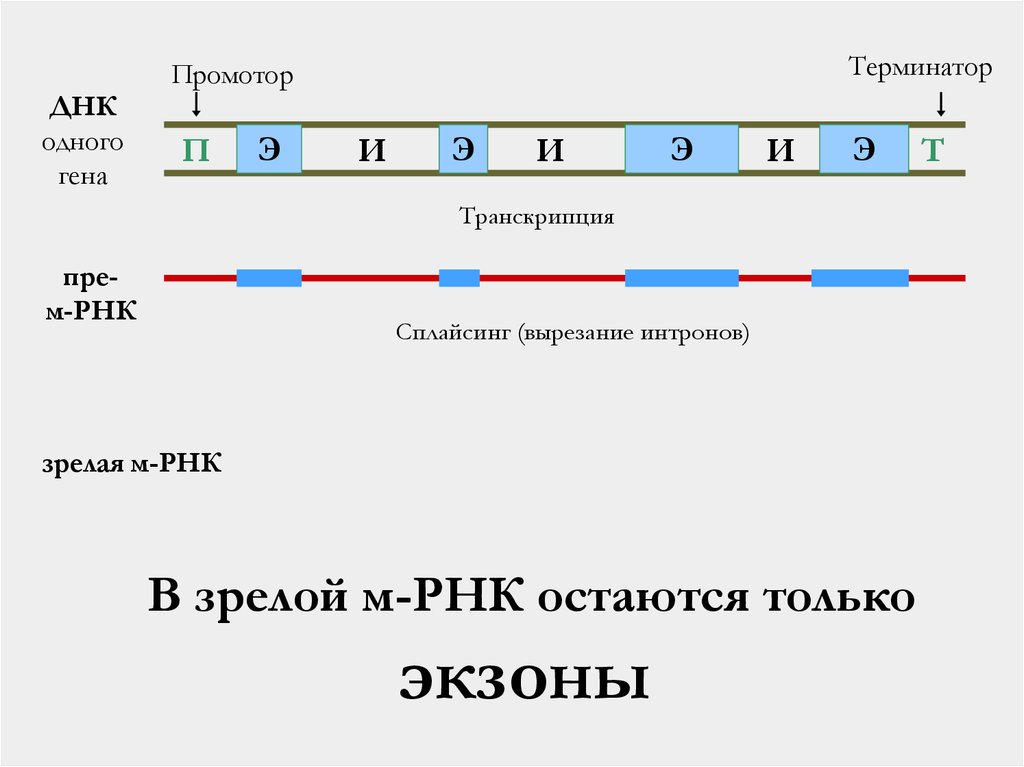
Интроны и экзоны
Интроны – вставки в эукариотические гены,
которые вырезаются после транскрипции из мРНК
Экзоны – участки гена, кодирующие белок.
Только они остаются в составе м-РНК после
вырезания интронов.
46.
ТерминаторПромотор
ДНК
одного
гена
П
Э
И
Э
И
Э
И
Э
Транскрипция
прем-РНК
Сплайсинг (вырезание интронов)
зрелая м-РНК
В зрелой м-РНК остаются только
экзоны
Т
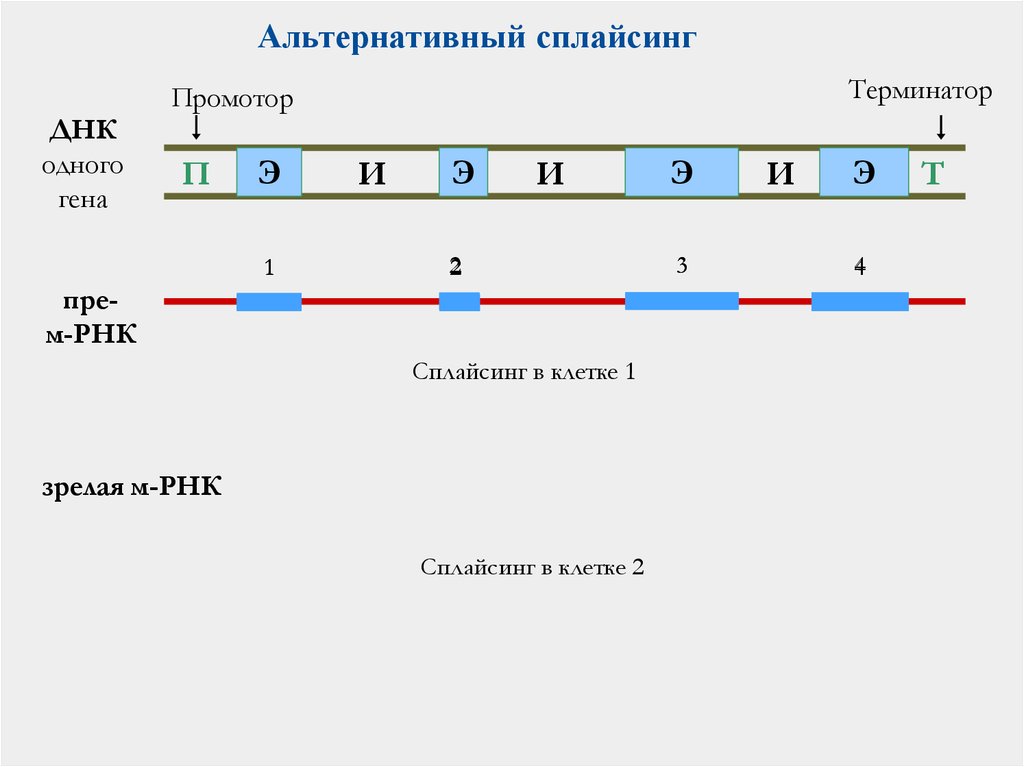
47.
Альтернативный сплайсингТерминатор
Промотор
ДНК
одного
гена
П
Э
1
И
Э
И
2
прем-РНК
Сплайсинг в клетке 1
зрелая м-РНК
Сплайсинг в клетке 2
Э
3
И
Э
4
Т
48. Альтернативный сплайсинг
в разных органахна разных стадиях развития
в разных состояниях клетки
Человек
≈
25 000 генов
≈ 300 000 белков
94% генов человека проходит
альтернативный сплайсинг
49.
Значение для медициныИногда
в
некоторых
транскриптах
обнаруживают
альтернативные
механизмы сплайсинга, однако ошибки в
данном процессе играют важную роль в
развитии
многих
генетических
заболеваний.
50.
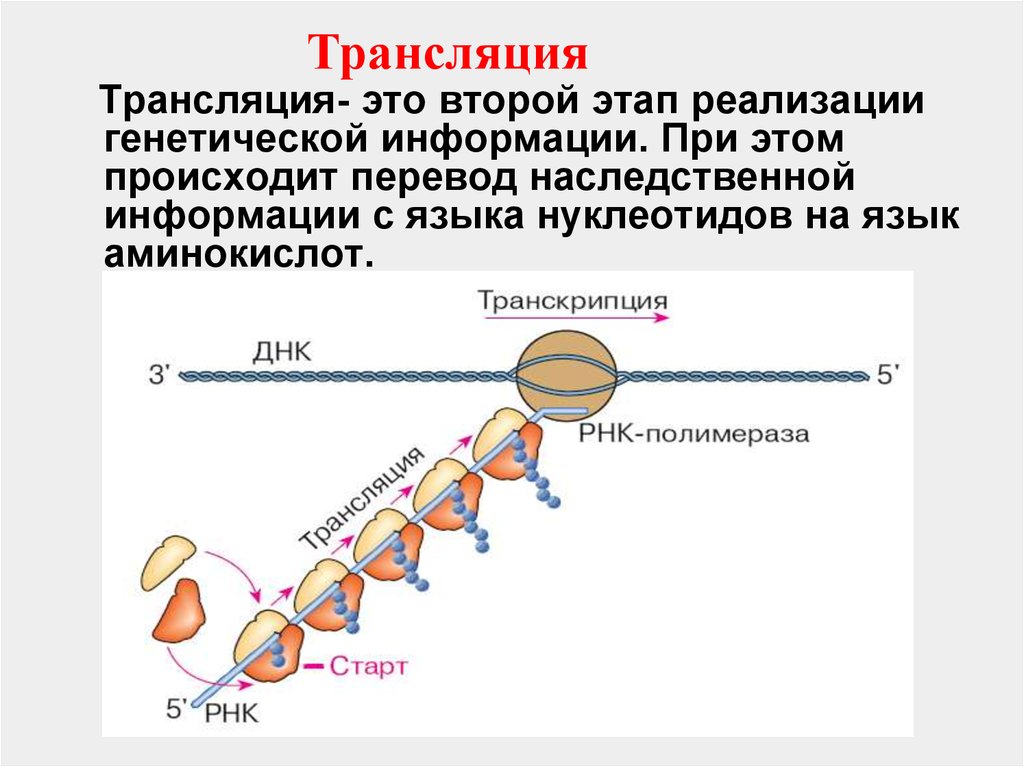
ТрансляцияТрансляция- это второй этап реализации
генетической информации. При этом
происходит перевод наследственной
информации с языка нуклеотидов на язык
аминокислот.
51. и-РНК → БЕЛОК
Синтез белка по матрице и-РНК,осуществляемый на рибосомах
Самый сложный из матричных
синтезов
– не просто копирование, а перевод
с языка нуклеиновых кислот на язык
белков. Словарь – генетический код.
52. Участники трансляции:
Аминокислоты;т – РНК;
Аминоацил- т – РНК синтетазы;
м – РНК;
Рибосомы;
АТФ, ГТФ
Белковые факторы инициации, элонгации, терминации
Ионы
53. Матричная РНК
Лидернаяпоследовательность
Шайна-Дальгарно
5'
Знак начала
трансляции
3‘ нетранслируемый
район
АУГ
STOP
Кодирующая часть,
транслируется
а.к. а.к. а.к. а.к. а.к. а.к. а.к. а.к.
БЕЛОК
3'
54. Транспортные РНК
Молекула-адаптор.Аминокислота
3'
Один ее конец узнает
кодон в м-РНК, а другой
– несет аминокислоту.
Антикодон
т-РНК 3'
5'
Г Ц У
Антикодон
м-РНК 5'
Ц Г А
Кодон
3'
55.
Этапы трансляции1. Инициация (начало)
2. Элонгация (удлинение)
3. Терминация (окончание)
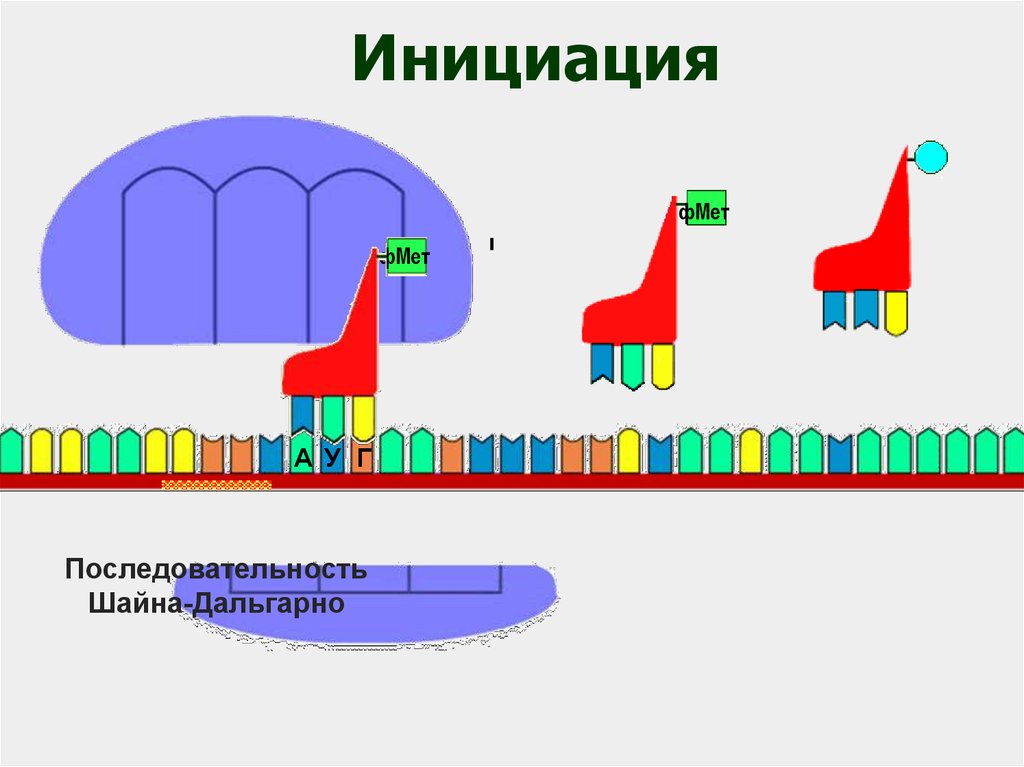
56.
ИнициацияфМет
фМет
А У Г
Последовательность
Шайна-Дальгарно
57.
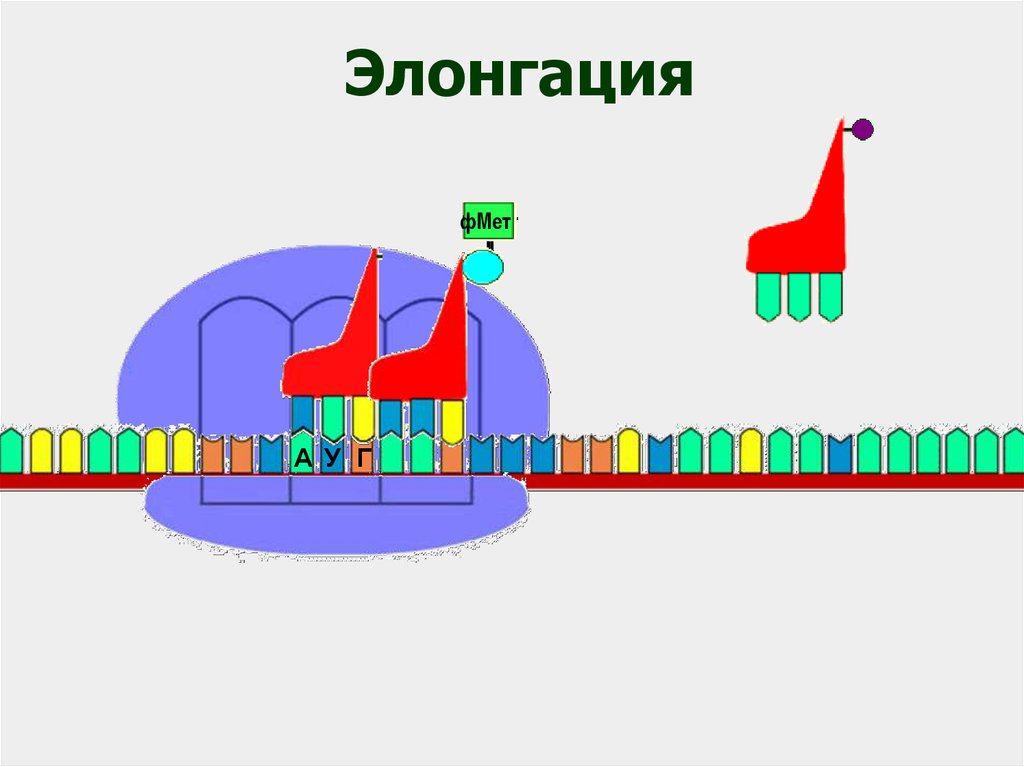
ЭлонгацияфМет
фМет
А У Г
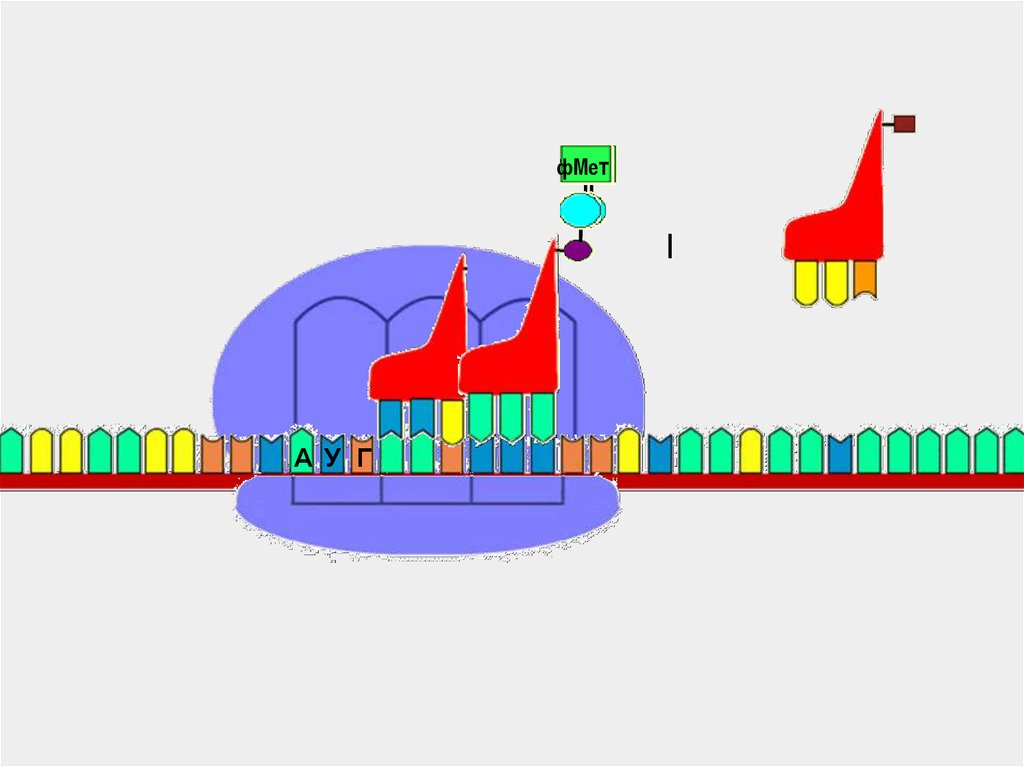
58.
фМетфМе
т
А У Г
59.
ТерминацияфМет
А У Г
стоп
60.
МетА У Г
стоп
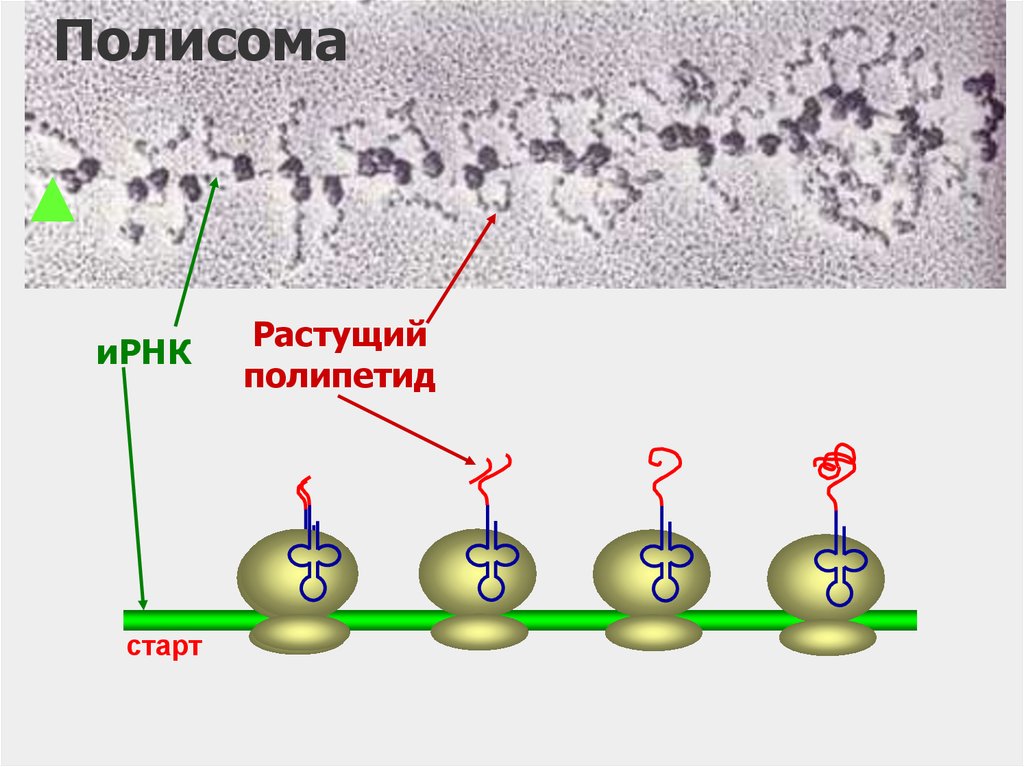
61.
ПолисомаиРНК
старт
Растущий
полипетид
62. Общие представление о фолдинге белков
Трансляция мРНК приводит к образованиюпептидной цепи со строго определенной
последовательностью аминокислот.
Далее происходит формирование белкафолдинг, т.е сворачивание пептидной цепи в
правильную трехмерную структуру.
Если белок состоит их нескольких
субъединиц, то фолдинг включает и
объединение их в единую макромолекулу.
63. Вспомогательные факторы фолдинга:
а) Фолдазы – белки с каталитическойактивностью:
протеиндисульфидизомераза,
пептидилпролилизомераза.
б) Молекулярные шапероны. К ним
относятся белки с самыми разными
механизмами действия.
64.
65. Генетический код
Способ записи информации опервичной структуре белков через
последовательность нуклеотидов ДНК и
РНК.
«Словарь» перевода с языка
нуклеиновых кислот на язык белков.
Полностью расшифрован к 1966
66. Георгий Антонович Гамов (1904-1968)
История открытия генетического кодаГеоргий Антонович
Гамов
(1904-1968)
Физик-теоретик
1954
Сформулировал
проблему кода и
предположил его
триплетность.
67. Проблема
Алфавит белков20 а.к.
Алфавит ДНК
и РНК
4 нуклеотида
68. Обоснование триплетности кода Гамовым
н. а.к.Сколько а.к. можно
закодировать
Моноплетный
1→1
4
Дуплетный
2→1
16
Триплетный
3→1
64
69.
Свойства генетическогокода
1. Триплетность - каждая аминокислота кодируется последовательностью из 3-х
нуклеотидов.
Определение: триплет или кодон - последовательность из трех нуклеотидов,
кодирующая одну аминокислоту.
2. Неперекрываемость - нуклеотид, входящий в состав кодона не входит в состав
другого кодона
3. Коллинеарность- порядок расположения аминокислот в полипептидной цепи
соответствует порядку расположения кодонов в полинуклеотидной цепи
4. Вырожденность - все аминокислоты, за исключением метионина и триптофана,
кодируются более чем одним триплетом:
5. Униполярность
7. Специфичность - один кодон кодирует одну аминокислоту. Например, кодон
АУГ кодирует метионин.
8. Универсальность - Генетический код един для всех живущих на Земле существ.
5→3
6. Непрерывность - считывание информации происходит непрерывно, без знаков
препинания.
70.
Первая буквав кодоне
У
Ц
А
Г
Вторая буква в кодоне
Третья буква
в кодоне
У
Ц
А
Г
Фен
Фен
Лей
Лей
Сер
Сер
Сер
Сер
Тир
Тир
STOP
STOP
Цис
Цис
STOP
Трп
У
Ц
А
Г
Лей
Лей
Лей
Лей
Про
Про
Про
Про
Гис
Гис
Глн
Глн
Арг
Арг
Арг
Арг
У
Ц
А
Г
Иле
Иле
Иле
Мет
(START)
Тре
Тре
Тре
Тре
Асн
Асн
Лиз
Лиз
Сер
Сер
Арг
Арг
У
Ц
А
Г
Вал
Вал
Вал
Вал
Ала
Ала
Ала
Ала
Асп
Асп
Глу
Глу
Гли
Гли
Гли
Гли
У
Ц
А
Г
71. Значение нарушений трансляции в медицине
72.
Значение для медициныЗнание белковых продуктов различных генов
позволяет успешно лечить многие болезни обмена
посредством введения в организм недостающих
ферментов (лечение болезни Помпе) или недопущения
поступления тех веществ, метаболический путь
которых
нарушен.
Например,
основа
лечения
фенилкетонурии, причиной которой служат мутации
гена фенилаланин-гидроксилазы, - диетотерапия,
исключающая поступление в организм с продуктами
питания аминокислоты фенилаланина.
73. Литература:
1.2.
3.
4.
5.
6.
Жимулев И.Ф. «Общая и молекулярная генетика»
Новосибирск., 2003, стр. 123-126, 172-191.
Гинтер Е.К. «Медицинская генетика» М., 2003. стр.
35-46.
Мушкамбаров Н.Н., Кузнецов С.Н. «Молекулярная
биология». М. 2003., стр. 125-176.
Фаллер Д.М. Шилдс Д. «Молекулярная биология
клетки», пер. с англ. М. 2003. стр
«Генетика». Учебник для ВУЗов (под редакцией
академика РАМН В.И. Иванова – М.2006. стр. 200203.
Албертс Б. и др. «Молекулярная биология клетки».
Пер. с англ. том 2. М. Мир 1994. стр. 253-267.