










")











 заполняет пространство между нейронами, составляет 40% от объема мозга. Глиальные клетки по размерам в")












")

































 Биология
БиологияПохожие презентации:










Общая ЦНС
1. Общая ЦНС
2.
План лекции:1. Методы исследования ЦНС.
2. Нейрон, как структурная и функциональная единица
ЦНС.
3. Рефлекторная теория.
4. Понятие о нервном центре и его свойства.
5. Торможение в ЦНС.
6. Координационные принципы рефлекторной деятельности
в ЦНС.
3. Функции ЦНС:
1. взаимосвязь органов и систем2. связь организма с внешней средой
3. психическая деятельность
4. МЕТОДЫ ИССЛЕДОВАНИЯ ЦНС
ПеререзокЭкстирпации
Раздражения и регшистраций
Стереотаксиса
Анатомо-клинических наблюдений
Нейрогистохимя
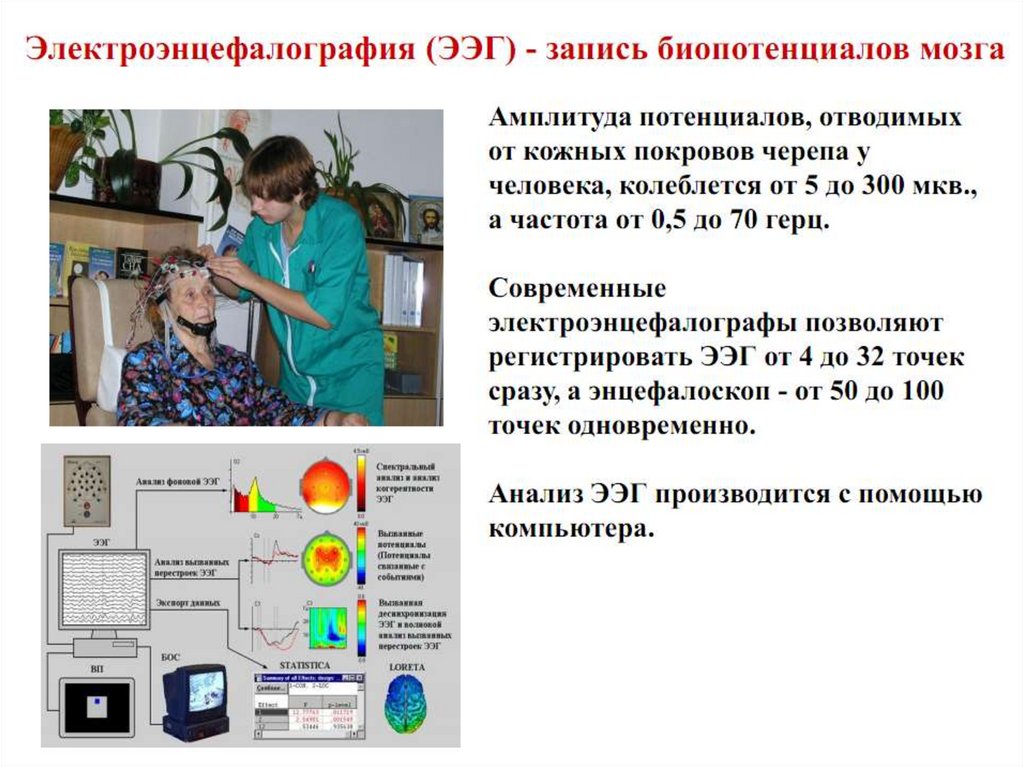
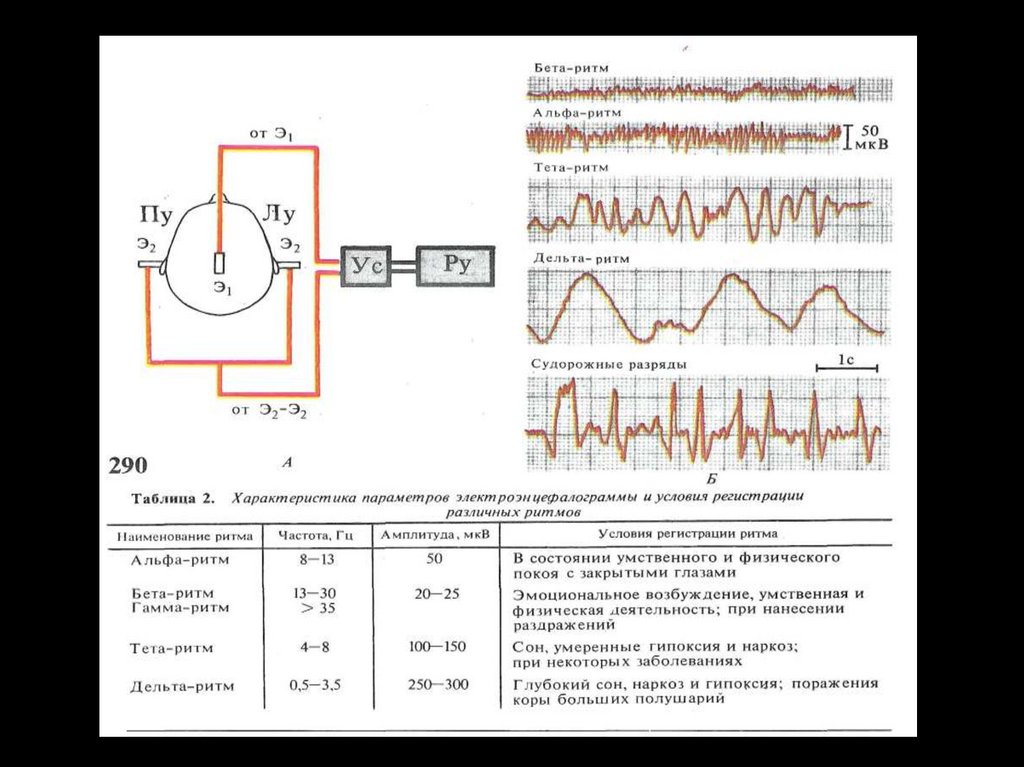
Регистрации электропотенциалов (ЭЭГ)
Компьютерная томография (рентгеновская,
ЯМР, МРТ)
9. Радиоизотопные методы
10. Метод условных рефлексов.
1.
2.
3.
4.
5.
6.
7.
8.
5. Стереотаксис
6. Картография нервных ядер с помощью метода стереотаксиса
7. Кортикография и метод раздражений
8. Микроэлектродный метод
9.
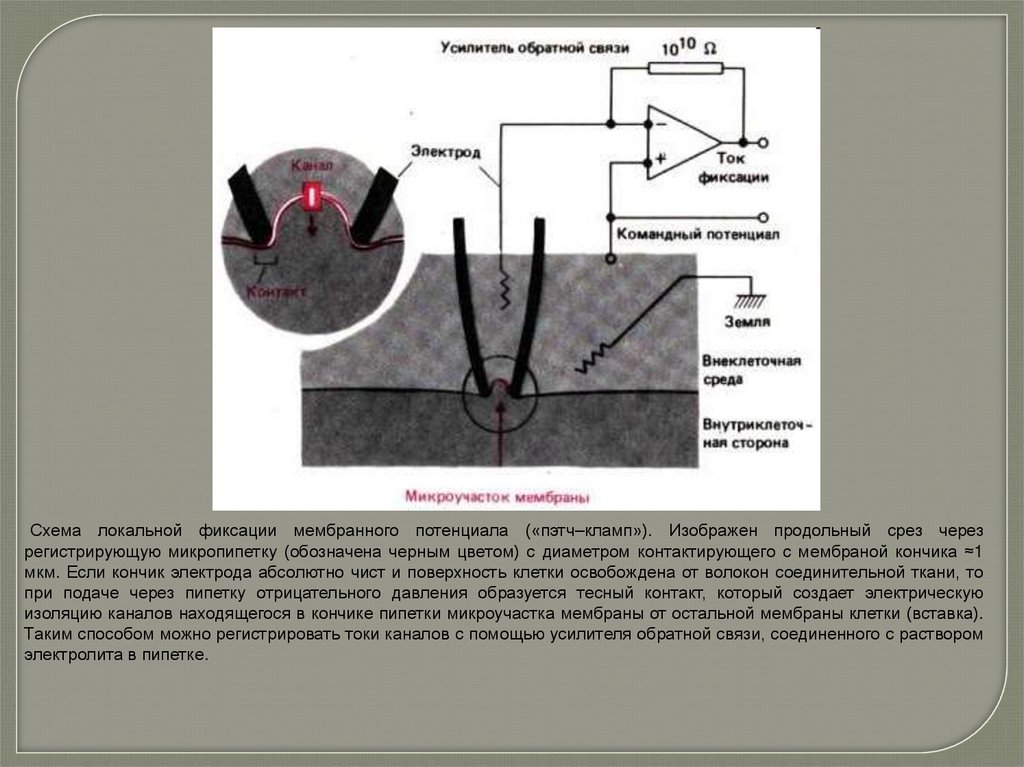
Схема локальной фиксации мембранного потенциала («пэтч–кламп»). Изображен продольный срез черезрегистрирующую микропипетку (обозначена черным цветом) с диаметром контактирующего с мембраной кончика ≈1
мкм. Если кончик электрода абсолютно чист и поверхность клетки освобождена от волокон соединительной ткани, то
при подаче через пипетку отрицательного давления образуется тесный контакт, который создает электрическую
изоляцию каналов находящегося в кончике пипетки микроучастка мембраны от остальной мембраны клетки (вставка).
Таким способом можно регистрировать токи каналов с помощью усилителя обратной связи, соединенного с раствором
электролита в пипетке.
10.
11.
12. Позитронно-эмиссионная томография
Магнитно-резонанснаятомография
13. Нейрон – структурная и функциональная единица нервной системы. Он способен принимать, обрабатывать, кодировать, передавать и
хранить информацию, устанавливатьконтакты с другими клетками, организовывать реакции на раздражение.
Число нейронов мозга человека ~ 10 11. Размеры нейронов – 6-120 мкМ.
На одном нейроне может быть до 10 000 синапсов. Если эти элементы
принять за ячейки хранения информации, то общий объем памяти нервной
системы составит 10 19 бит.
14.
КЛАССИФИКАЦИИ НЕЙРОНОВпо функции:
Афферентные нейроны проводят возбуждение от рецепторов периферических
органов в структуры ЦНС
Эфферентные нейроны осуществляют передачу сигналов от ЦНС к органамэффекторам (мышцам и железам)
Ассоциативные нейроны проводят возбуждение между нейронами
15.
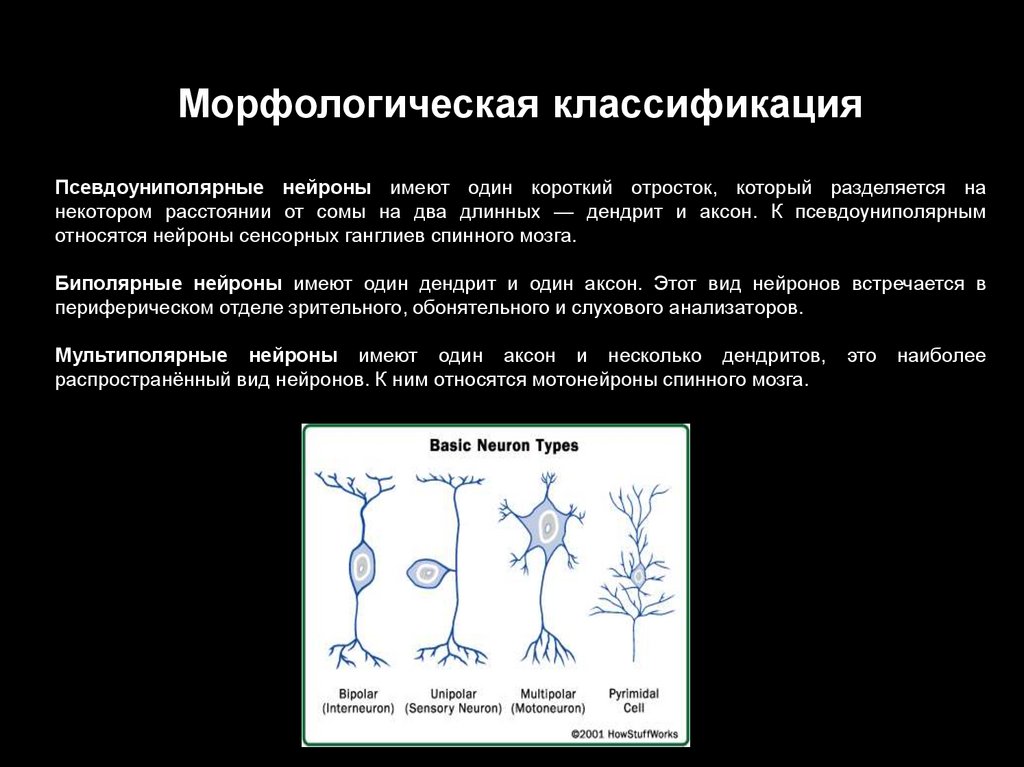
Морфологическая классификацияПсевдоуниполярные нейроны имеют один короткий отросток, который разделяется на
некотором расстоянии от сомы на два длинных — дендрит и аксон. К псевдоуниполярным
относятся нейроны сенсорных ганглиев спинного мозга.
Биполярные нейроны имеют один дендрит и один аксон. Этот вид нейронов встречается в
периферическом отделе зрительного, обонятельного и слухового анализаторов.
Мультиполярные нейроны имеют один аксон и несколько дендритов,
распространённый вид нейронов. К ним относятся мотонейроны спинного мозга.
это
наиболее
16. КЛАССИФИКАЦИЯ НЕЙРОНОВ ПО МОДАЛЬНОСТИ (чувствительности)
1. Моносенсорные нейроны – располагаются в первичных проекционных
зонах коры и реагируют только на сигналы своей модальности. Например,
нейроны первичной зрительной коры – реагируют только на световые
раздражения сетчатки.
2. Бисенсорные нейроны – кроме своей модальности способны реагировать и
на другие сигналы. Например, нейроны вторичной зрительной коры реагируют
на зрительные и слуховые раздражения.
3. Полисенсорные нейроны – располагаются чаще всего в ассациативных
зонах мозга. Они способны реагировать на раздражения слуховой, зрительной,
кожной и других анализаторных систем.
17. по медиатору:
Холинергические,
Адренергические,
ГАМКергические и т.д.
Принцип Дейла (1936): в одних и тех же нейронах
выделяется один вид медиатора.
В настоящее время пересмотрен:
1.
2.
В двигательной концевой пластине вместе с Ах выделяется АТФ .
Часто НА выделяется с пептидом (медиатор-спутник)
18. КЛАССИФИКАЦИЯ ПО МЕСТУ КОНТАКТА
- аксосоматические,
- аксоаксональные,
- аксодендретические
- дендросоматические,
- дендродендретические
- дендроаксональные
химические синапсы
электрические синапсы
19. по активности :
1. молчащие2. спонтанноактивные или фоновоактивные
a. возбуждающиеся
b. тормозящиеся
молчащие
спонтанноактивные
возбуждающиеся
тормозящиеся
20. Классификация нейронов по электрической активности
Фоновая активность:1.) Непрерывно-аритмичные (замедляют и увеличивают частоту разряда). Обеспечивают тонус нервного
центра.
2.) Пачечные – выдают группу импульсов с короткими межимпульсными интервалами. Они создают условия
для проведения сигналов при снижении функциональных возможностей проводящих или воспринимающих
структур.
3.) Групповой тип - апериодическое появление в фоне группы импульсов, сменяющихся периодом молчания.
21. КОДИРОВАНИЕ ИНФОРМАЦИИ В НЕЙРОННОЙ ЦЕПИ
• ИМПУЛЬСНОЕ:НЕПРЕРЫВНОЕ, ПАЧЕЧНОЕ, ЧАСТОТНОЕ,
ИНТЕРВАЛЬНОЕ, ДЛИТЕЛЬНОСТЬЮ
АКТИВНОСТИ, ВАРИАБЕЛЬНОСТЬЮ
АКТИВНОСТИ
• НЕИМПУЛЬСНОЕ:
СОСТАВ И СКОРОСТЬ ТОКА
АКСОПЛАЗМЫ
22. Функционально нейрон состоит из:
1.воспринимающей (дендриты и сома)части,
2.интегративной (сома) части,
3.передающей (аксон) части
23.
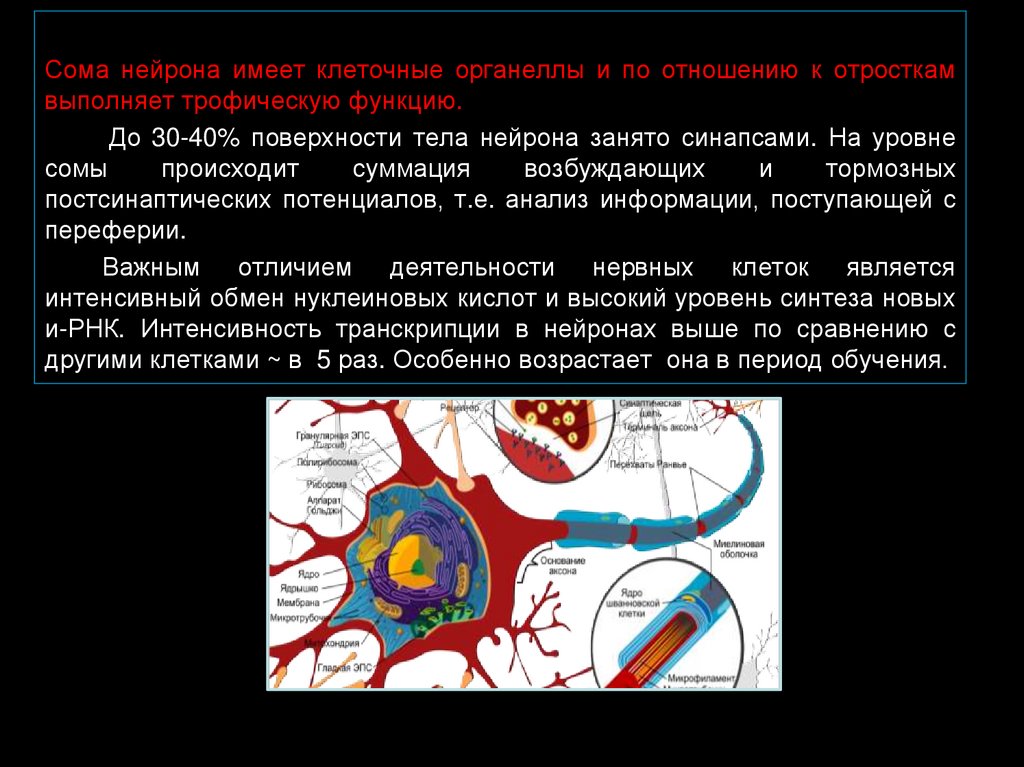
Сома нейрона имеет клеточные органеллы и по отношению к отросткамвыполняет трофическую функцию.
До 30-40% поверхности тела нейрона занято синапсами. На уровне
сомы
происходит
суммация
возбуждающих
и
тормозных
постсинаптических потенциалов, т.е. анализ информации, поступающей с
переферии.
Важным отличием деятельности нервных клеток является
интенсивный обмен нуклеиновых кислот и высокий уровень синтеза новых
и-РНК. Интенсивность транскрипции в нейронах выше по сравнению с
другими клетками ~ в 5 раз. Особенно возрастает она в период обучения.
24. Генерация ПД в нейронах
ПД на соме и дендритахвозникнуть не могут, так
как очень большой порог
возбуждения.
Здесь возникают лишь ПСП
(ВПСП и ТПСП) в виде ЛО,
которые суммируются.
Таким образом происходит
анализ и синтез
поступившей информации.
25. Генерация ПД в нейронах
Суммированный ПСП может достигатьаксонального холмика и только тогда
возникает ПД (Ео = - 70 мВ, Ек = - 50
мВ).
Распространение возбуждения идет в
обе стороны, но на соме затухает изза большого порога.
По аксону ПД распространяется на
большие расстояния.
Особенностью ПД нейронов является
его Nа+ -Са 2+– К + – природа.
Повышенный вход Са 2+ в клетку во
время деполяризации приводит к
длительной
следовой
гиперполяризации, за счет активации Са 2+
-зависимых К+ - каналов.
Это
защищает
нерон
от
перевозбуждения.
26. Функции аксона
Основная функция аксона – проведение нервного импульса (ПД).
Длина аксона может составлять до нескольких десятков сантиметров.
Наиболее возбудимой структурой аксона является аксональный
холмик – начальный сегмент аксона, лишенный миелиновой оболочки.
Кроме этого по аксону в ортодромном (или антероградное ) и
антидромном (или ретроградное ) направлениях постоянно идет
транспорт веществ. Выделяют: быстрый – 410 мм/сут, медленный - 1-5
мм/сут и промежуточный.
27.
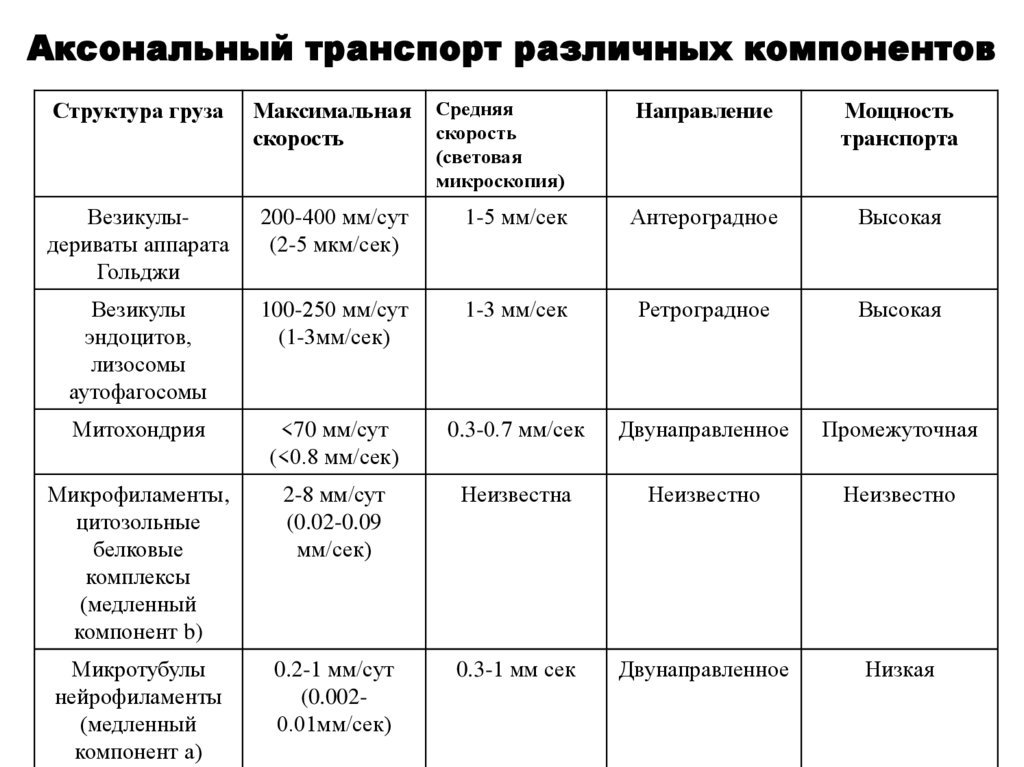
Аксональный транспорт различных компонентовСтруктура груза
Максимальная
скорость
Средняя
скорость
(световая
микроскопия)
Направление
Мощность
транспорта
Везикулыдериваты аппарата
Гольджи
200-400 мм/сут
(2-5 мкм/сек)
1-5 мм/сек
Антероградное
Высокая
Везикулы
эндоцитов,
лизосомы
аутофагосомы
100-250 мм/сут
(1-3мм/сек)
1-3 мм/сек
Ретроградное
Высокая
Митохондрия
<70 мм/сут
(<0.8 мм/сек)
0.3-0.7 мм/сек
Двунаправленное
Промежуточная
Микрофиламенты,
цитозольные
белковые
комплексы
(медленный
компонент b)
2-8 мм/сут
(0.02-0.09
мм/сек)
Неизвестна
Неизвестно
Неизвестно
Микротубулы
нейрофиламенты
(медленный
компонент а)
0.2-1 мм/сут
(0.0020.01мм/сек)
0.3-1 мм сек
Двунаправленное
Низкая
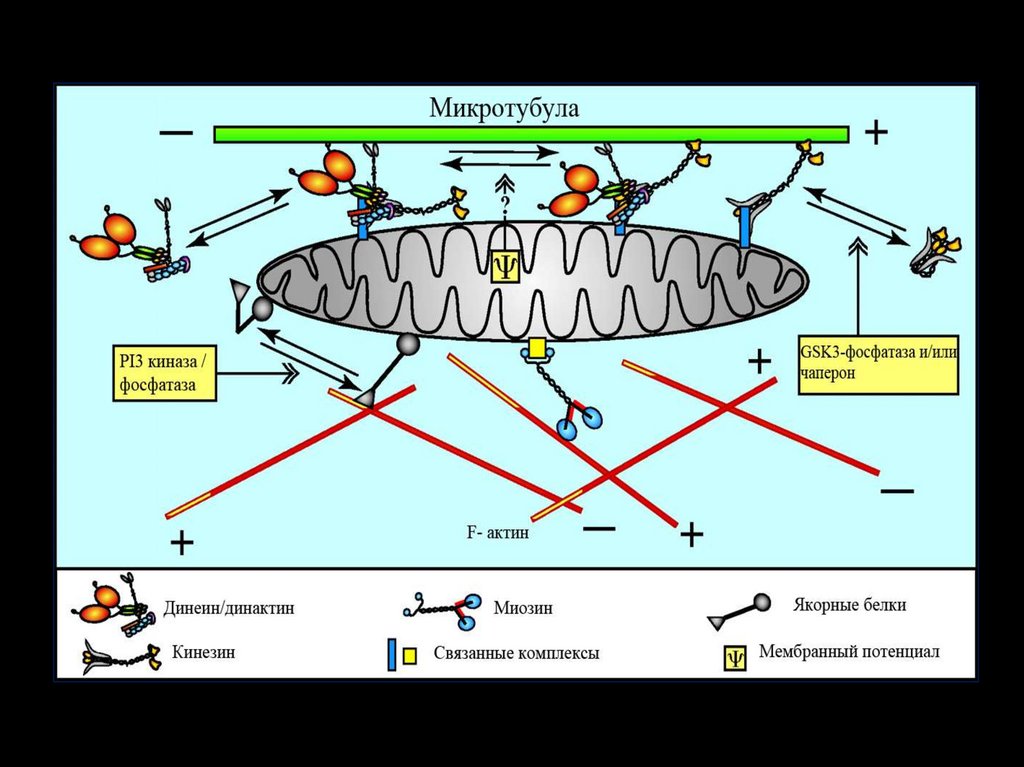
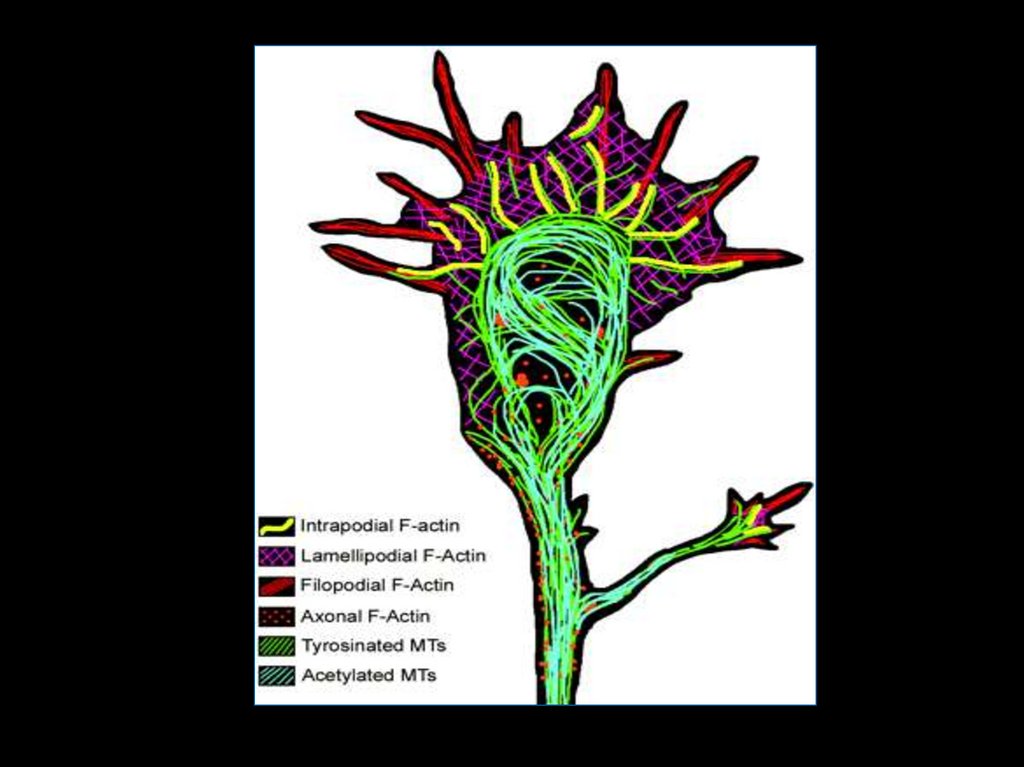
28. Динеин и кинезин обеспечивают транспорт крупных молекул и органоидов. Динеин – ретроградно кинезин – антероградно.
29.
30.
31.
32.
33.
Нейродегенеративные заболевания, связанныес ослаблением аксонального транспорта
Заболевания
Мутантный ген / нарушения синтеза
белка
Наследственная спастическая
параплегия
Тяжелые цепи кинезина
Семейная моторно-нейрональная
болезнь
Субъединица p 150 динактина
Болезнь Шарко-Мари-Тута типа 2А
(мышечная атрофия перонеального
типа)
b-субъединица кинезина Kif 1B
Болезнь Хантингтона
Хантингтин
Семейные амиотрофические
латеральные склерозы
Супероксид дисмутаза I
Болезнь Паркинсона и
связанные синуклеинопатии
a-синуклин
Болезнь Альцгеймера
АРР, призенилины, Тау
Нейродегенеративные Тауопатии
Таu
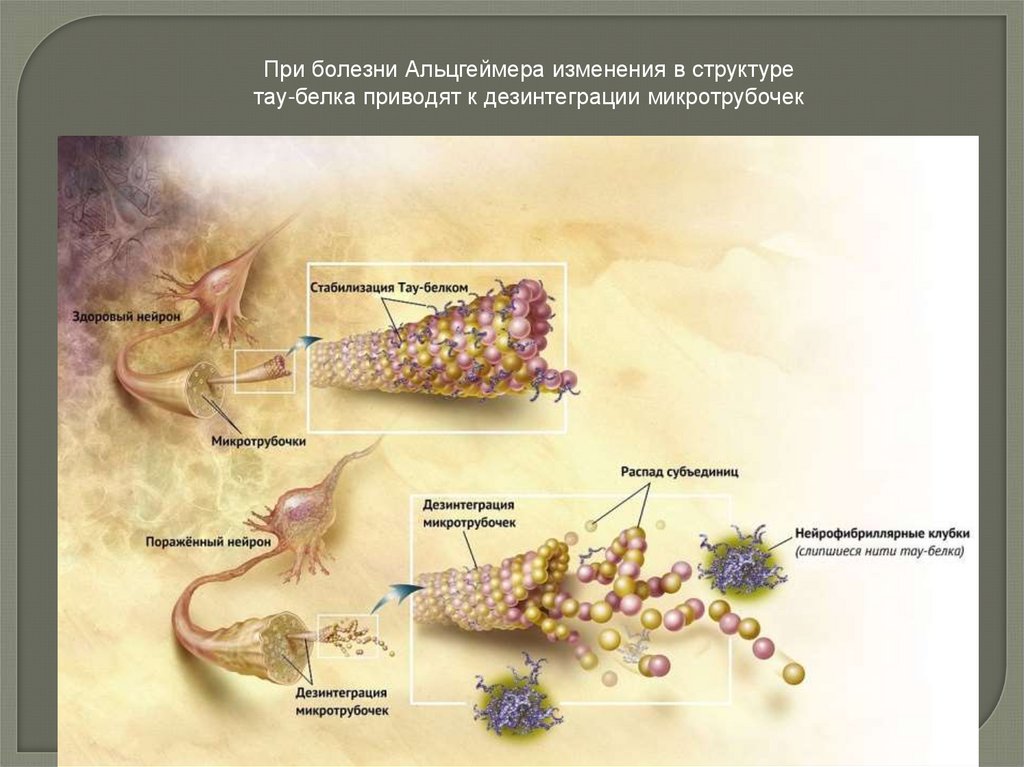
34.
При болезни Альцгеймера изменения в структуретау-белка приводят к дезинтеграции микротрубочек
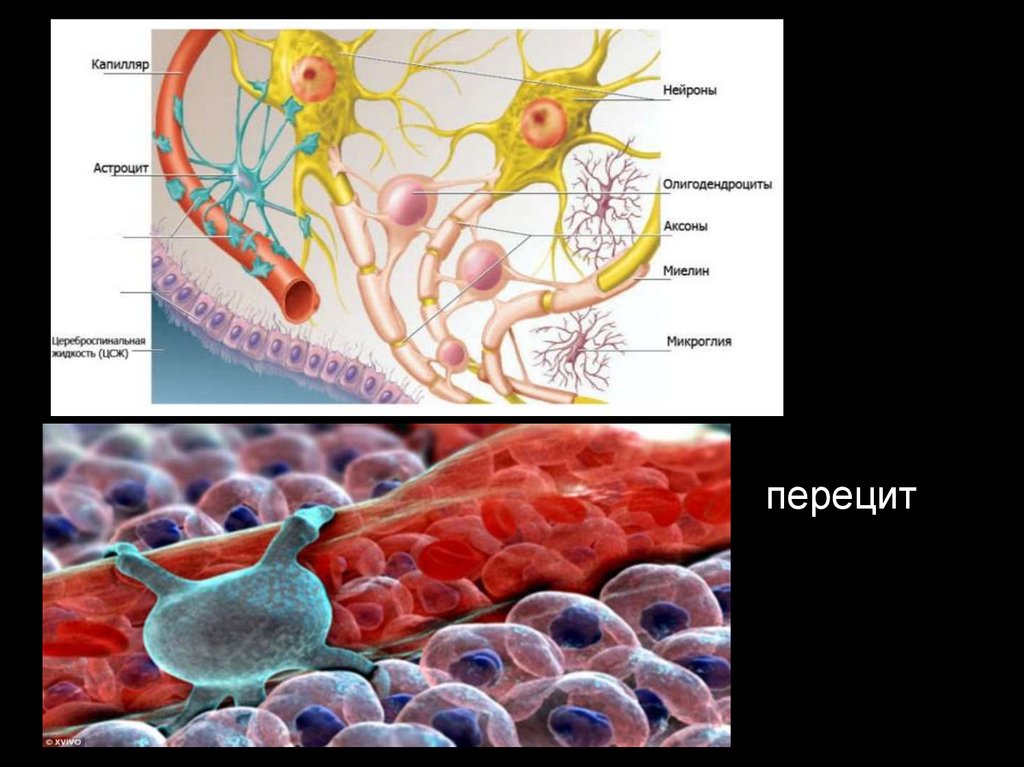
35. Нейроглия (нервный клей) заполняет пространство между нейронами, составляет 40% от объема мозга. Глиальные клетки по размерам в
3-4 раза меньше, чемнервные, число их в ЦНС достигает 140 млрд.
С возрастом у человека в мозге число нейронов
уменьшается, а число глиальных клеток возрастает.
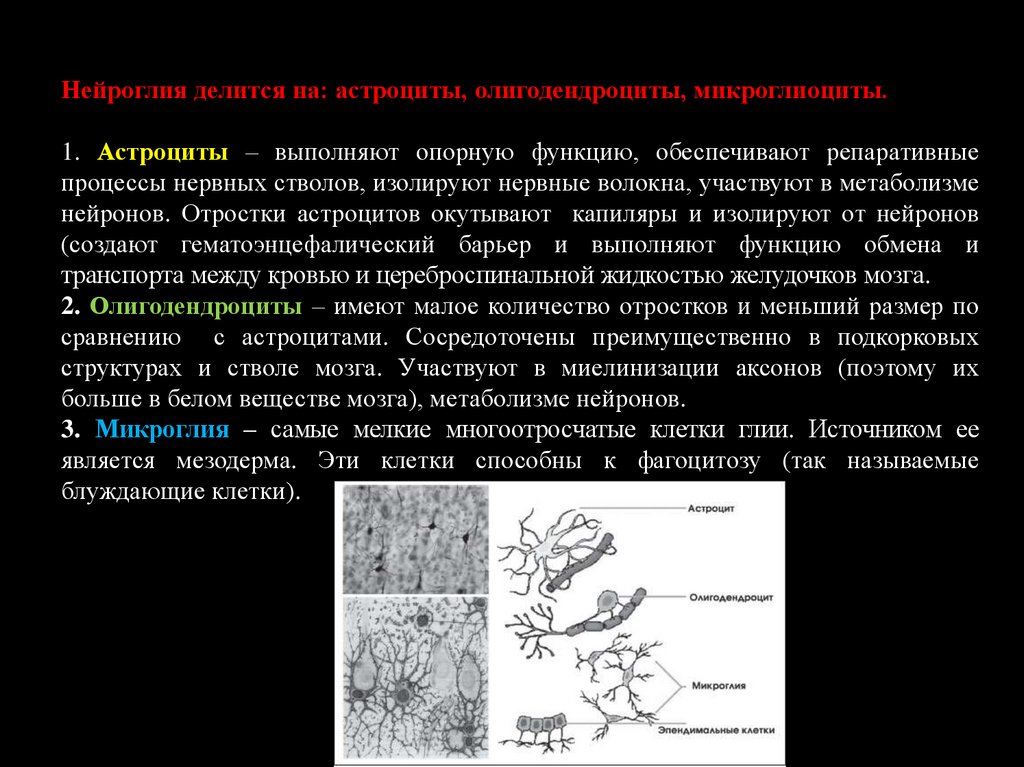
36.
Нейроглия делится на: астроциты, олигодендроциты, микроглиоциты.1. Астроциты – выполняют опорную функцию, обеспечивают репаративные
процессы нервных стволов, изолируют нервные волокна, участвуют в метаболизме
нейронов. Отростки астроцитов окутывают капиляры и изолируют от нейронов
(создают гематоэнцефалический барьер и выполняют функцию обмена и
транспорта между кровью и цереброспинальной жидкостью желудочков мозга.
2. Олигодендроциты – имеют малое количество отростков и меньший размер по
сравнению с астроцитами. Сосредоточены преимущественно в подкорковых
структурах и стволе мозга. Участвуют в миелинизации аксонов (поэтому их
больше в белом веществе мозга), метаболизме нейронов.
3. Микроглия – самые мелкие многоотросчатые клетки глии. Источником ее
является мезодерма. Эти клетки способны к фагоцитозу (так называемые
блуждающие клетки).
37.
ФУНКЦИИНЕЙРОГЛИИ:
1. Опорная функция.
2. Электрическая изоляция.
3. Участие в восстановлении и регенерации нервной ткани
(способны к делению в течение всей жизни).
4. Участие в онтогенетическом развитии нервной системы.
5. Участие в обмене веществ нейронов и транспорте.
6. Буфферная функция
38. БУФЕРНАЯ ФУНКЦИЯ ГЛИИ
Тесный контакт глии с нейронами - обусловливает изменение мембранного
потенциала глии при возбуждении нейронов. Так, выход ионов К+ во
внеклеточную среду во время реполяризации нейронов – изменяет его
концентрацию вокруг глии и приводит к ее деполяризации. Поглащая ионы
К+, глиальные клетки снижают внеклеточную концентрацию К+, образуя как
бы буфер.
Затем они медленно выделяют К+. Таким образом, во внеклеточной среде
поддерживается постоянная концентрация К+, что важно для поддержания
нормальной возбудимости нейронов.
39. Пульсации глии
Одной из особенностей глиальных клеток является их способность к
изменению размера, который носит ритмический характер.
Физиологическая роль такого рода «пульсаций» состоит в
проталкивании аксоплазмы нейрона и влиянии на ток жидкости в
межклеточном пространстве.
Периодичность глиальных «пульсаций» может изменяться под
действием БАВ. Например, норадреналин – усиливает их, а серотонин –
уменьшает.
40. Гематоэнцефалический барьер
41.
перецит42. Структура гематоэнцефалического барьера
Артериальные капилярыЭндотелий
Сосудистые сплетения
Мембраны
Образование ЦСЖ
Перециты
Желудочки мозга
Нейроглия
Внеклеточное пространство мозга
Нейроны
Венозные капиляры
Субарахиодальная жидкость
Венозная система
43.
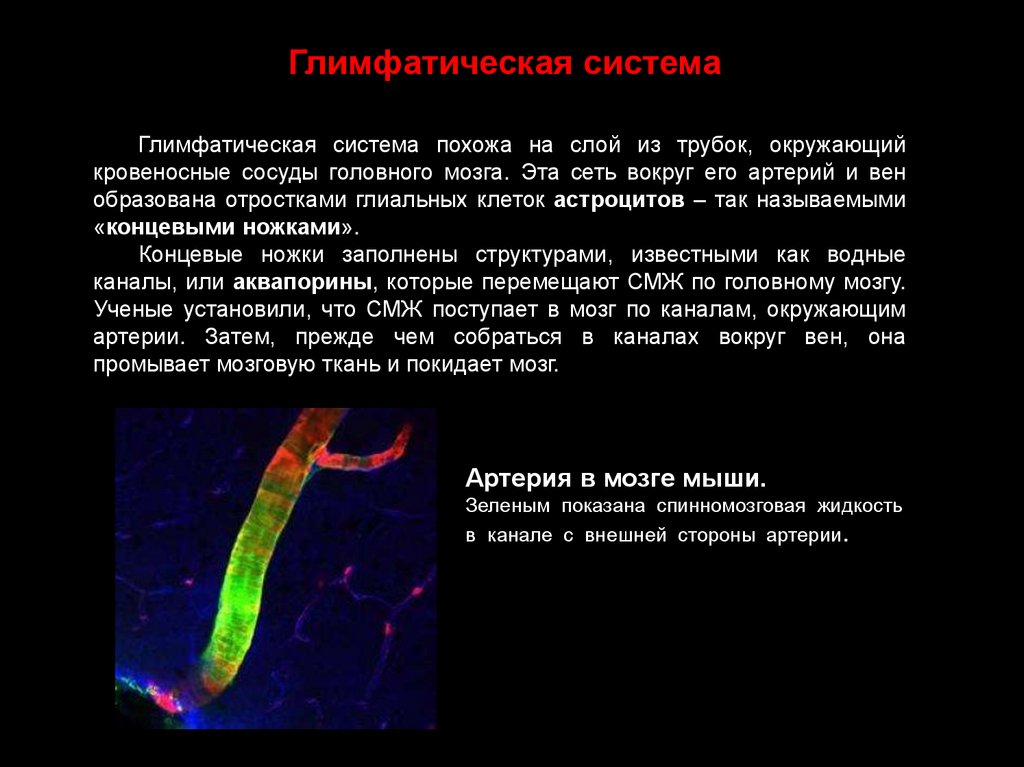
Глимфатическая системаГлимфатическая система похожа на слой из трубок, окружающий
кровеносные сосуды головного мозга. Эта сеть вокруг его артерий и вен
образована отростками глиальных клеток астроцитов – так называемыми
«концевыми ножками».
Концевые ножки заполнены структурами, известными как водные
каналы, или аквапорины, которые перемещают СМЖ по головному мозгу.
Ученые установили, что СМЖ поступает в мозг по каналам, окружающим
артерии. Затем, прежде чем собраться в каналах вокруг вен, она
промывает мозговую ткань и покидает мозг.
Артерия в мозге мыши.
Зеленым показана спинномозговая жидкость
в канале с внешней стороны артерии.
44. Рефлекторная теория
1637 — Р. Декарт (R. Descartes, Франция) дал первое описание двигательного рефлекса.
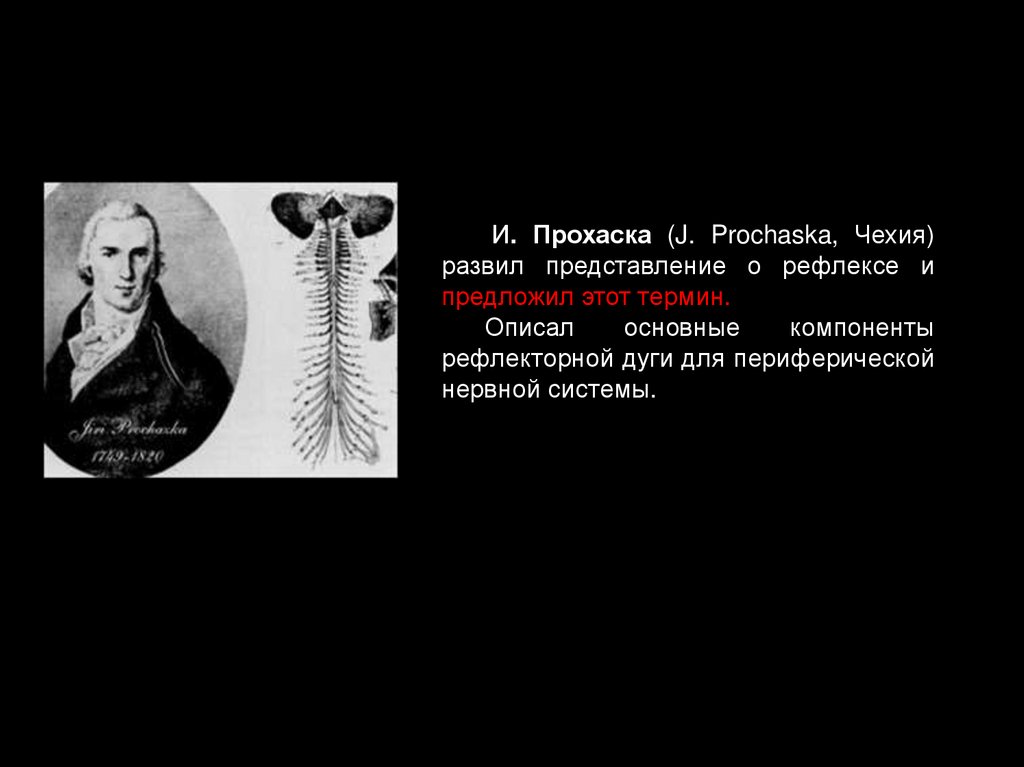
1784 — И. Прохаска (J. Prochaska, Чехия) развил представление о рефлексе и предложил
этот термин. Описал основные компоненты рефлекторной дуги. Дал биологическое
обоснование рефлекса.
1811 и 1822 — Ч. Белл (Ch. Bell, Великобритания) и Ф. Мажанди (F. Magendie, Франция)
открыли порядок распределения чувствительных и двигательных нервных волокон между
дорсальными и вентральными корешками спинного мозга (закон Белла—Мажанди).
Маршалл Холл предложил термин рефлектрная дуга и описал 3 ее компонента.
1832 — М. Голл и И. Мюллер (М. Goll, J. Muller, Германия) изложили учение о рефлексе.
1862 – И.П. Сеченов открыл явление торможения в ЦНС, что послужило толчком к
развитию учения о координации рефлекторных актов.
1902 — в докладе из лаборатории И. П. Павлова впервые заявлено об открытии условных
рефлексов.
1923 — А. А. Ухтомский (СССР) создал учение о доминанте.
1932 — Ч. Шеррингтон и Э. Эдриан (Е. Adrian, Великобритания) — Нобелевская премия за
открытие роли рецепторов нервов, передачи информации в виде электрических импульсов.
1930—е — П. К. Анохин (СССР) высказал идею об «акцепторе результата действия» и,
таким образом, открыл существование обратной связи в физиологических процессах (но не
использовал этого термина). Позднее создал учение о «функциональных системах».
1941 — Н. А. Бернштейн (СССР) в трудах по физиологии движений создал схему
рефлекторного кольца.
45. Французский философ, математик, анатом
Под рефлексом понималдвижение «животных духов от
мозга к мышцам, по типу
отражения светового луча:
-Внешние предметы действуют
на переферические окончания,
расположенных внутри «нервной
трубки»
нервных нитей. Они,
натягиваясь, открывают клапаны
отверстий, ведущих из мозга в
нервы. По каналам этих нервов
«животные духи» перемещаются
в соответствующие
мышцы,
которые
в
результате
раздуваются, и, таким образом, происходит движение.
Сами же «животные духи»
находились в центре мозга – в
эпифизе.
Таким
образом,
теория
Декарта была дуалистичной.
46.
47.
И. Прохаска (J. Prochaska, Чехия)развил представление о рефлексе и
предложил этот термин.
Описал
основные
компоненты
рефлекторной дуги для периферической
нервной системы.
48.
• 1811 и 1822 — Ч. Белл (Ch. Bell, Великобритания) и Ф.Мажанди
(F.
Magendie,
Франция)
открыли
порядок
распределения чувствительных и двигательных нервных
волокон между дорсальными и вентральными корешками
спинного мозга (закон Белла—Мажанди).
49. Отец русской школы физиологии
Открылявление
торможения в ЦНС, что
послужило
толчком
к
развитию
учения
о
координации рефлекторных
актов.
Впервые констатировал,
что все сознательное и бессознательное
совершенствуется по типу рефлекса.
50.
ШЕ́ РРИНГТОН (Sherrington) Чарлз Скотт.Автор фундаментальных открытий в области нейрофизиологии (главным
образом рефлекторной деятельности). Создал учение о рецептивных полях.
Разработал концепцию интегративной деятельности нервной системы.
Труды Шеррингтона сыграли важную роль в развитии экспериментальной и
клинической неврологии. Ввел представление о синапсах и определил их
значение в механизмах нервно-мышечной передачи.
Нобелевская премия (1932, совместно с Э. Д. Эдрианом)
51. Основоположник советской школы физиологии, лауреат Нобелевской премии.
Развилрефлекторную
теорию И.М. Сеченова и
выделил
особый
класс
рефлексов
–
«условные
рефлексы».
Сформулировал основные
принципы
рефлекторной
деятельности.
Заложил основы учения о
ВНД,
типах
нервной
системы, анализаторах.
52. Принципы рефлекторной теории (по И.П. Павлову)
1. Принцип детерменизма (причинности) –
всякий нервный акт вызван определенным воздействием.
Стимул → рецептор → ощущение
2. Принцип стрктурности –
в мозге нет процессов, которые не имели бы материальной основы.
физиологический акт нервной деятельности приурочен к определенной структуре.
Каждый
3. Принцип единства анализа и синтеза –
аналитическая деятельность заключается в избирательном реагировании на составляющие
внешнего воздействия. С другой стороны, нервной системой обеспечивается целостное
восприятие всей совокупности сигналов – т.е. синтез.
53.
Создал учение о ДОМИНАНТЕ.54. Функциональная система
Пётр Кузьмич Анохин. Основные работыпосвящены
изучению
деятельности
целого организма и особенно головного
мозга
55.
56.
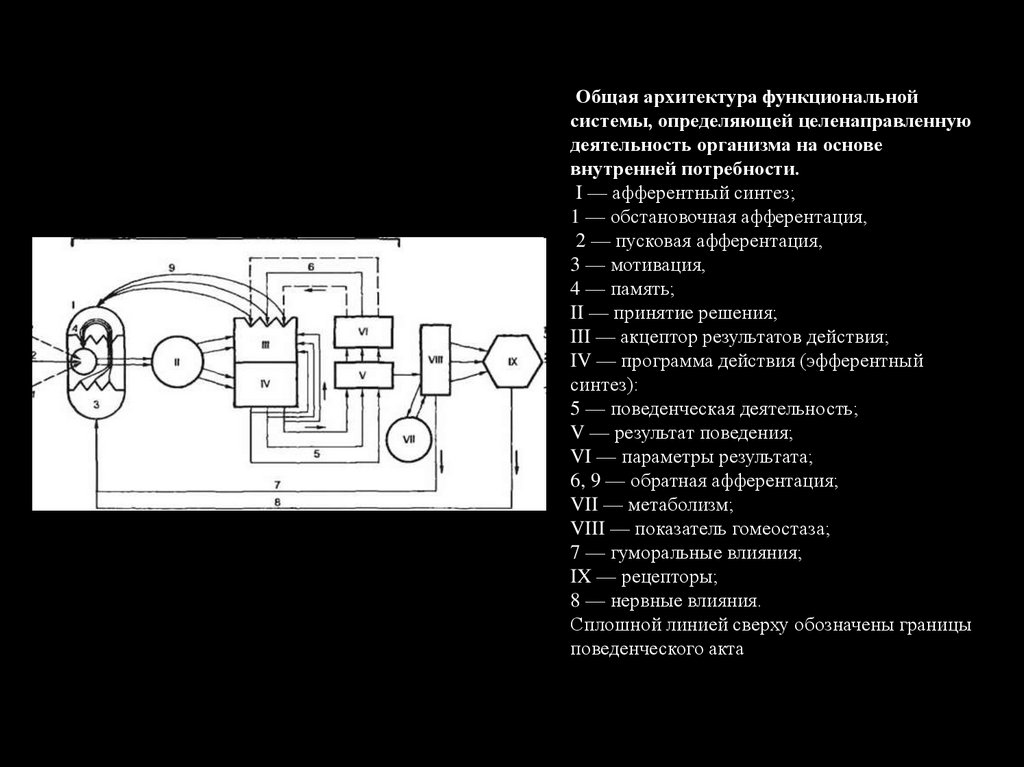
Общая архитектура функциональнойсистемы, определяющей целенаправленную
деятельность организма на основе
внутренней потребности.
I — афферентный синтез;
1 — обстановочная афферентация,
2 — пусковая афферентация,
3 — мотивация,
4 — память;
II — принятие решения;
III — акцептор результатов действия;
IV — программа действия (эфферентный
синтез):
5 — поведенческая деятельность;
V — результат поведения;
VI — параметры результата;
6, 9 — обратная афферентация;
VII — метаболизм;
VIII — показатель гомеостаза;
7 — гуморальные влияния;
IX — рецепторы;
8 — нервные влияния.
Сплошной линией сверху обозначены границы
поведенческого акта
57.
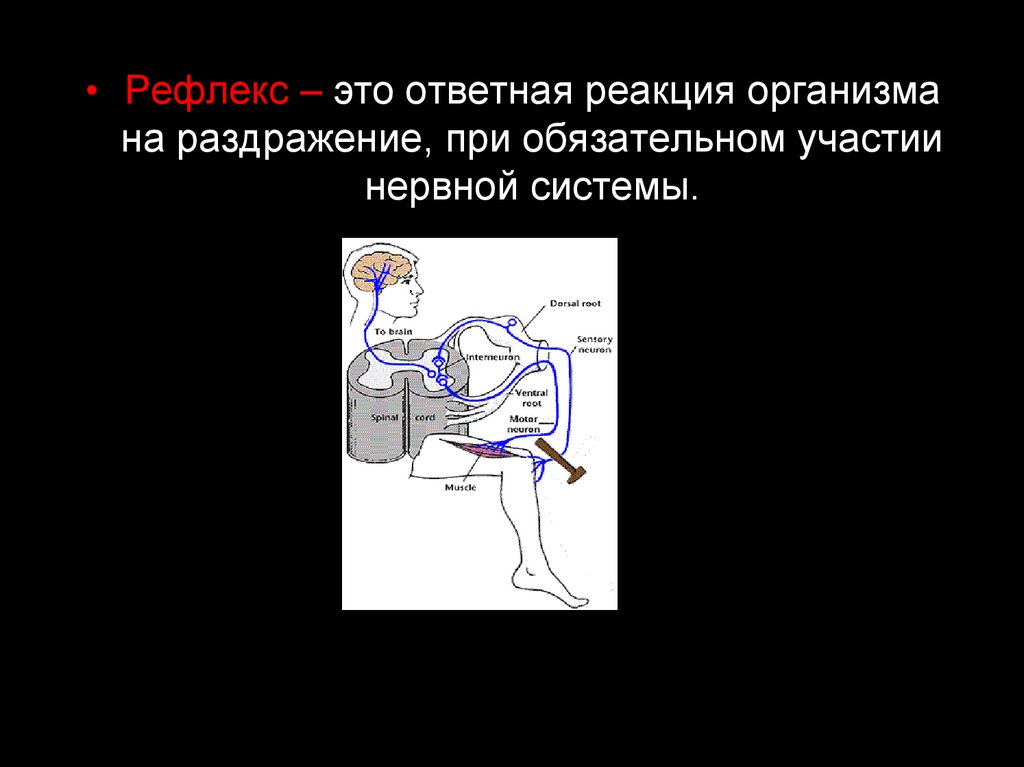
• Рефлекс – это ответная реакция организмана раздражение, при обязательном участии
нервной системы.
58. Классификации рефлексов
1.2.
3.
4.
5.
6.
7.
По происхождению – безусловные и условные.
По биологическому значению:
оборонительные, пищевые, половые, ориентировочные,
позднотонические, локомоторные.
По расположению рецептора: интеро-, экстро-, проприорецепторы
По месту замыкания рефлектороной дуги: спинальные,
бульбарные, мезенцефальные, диэнцефальные, кортикальные.
По характеру рефлекторного действия: моторные, секреторные,
сосудодвигательные.
По количеству нейронов в рефлекторной дуге:
моносинаптические и полисинаптические.
Соматические и вегетативные.
59. Рефлекторная дуга
- это комплекс специфическиорганизованных нервных клеток,
взаимодействие которых необходимо для
осуществления рефлекторного акта.
Рефлекс проявляется только при
сохранении целостности всех элементов
рефлекторной дуги.
60. Рефлекторная дуга соматического и вегетативного рефлекса
вегетативнаясоматическая
61. Звенья рефлекторной дуги:
1.рецептор (восприятие стимула икодирование его характеристик),
2.афферент (кодирование параметров
стимула и передача его в центральное
звено),
3.центральное звено (анализ и синтез
полученной информации),
4.эфферент (передача команды к рабочему
органу),
5.рабочий орган (реализация ответной
реакции на действие стимула)
62. Время рефлекса
подразделяют на:1. общее время рефлекса – то время,
которое проходит от момента
раздражения до ответной реакции.
2. центральное время рефлекса составляет
80 % от общего времени и зависит от
количества синапсов между нейронами в
рефлекторной дуге.
63. Время рефлекса уменьшается при увеличении силы раздражителя
При этом возможно 2 варианта сокращениявремени рефлекса:
1. расширение рецептивного поля и
пространственная суммация на
центральном нейроне деполяризущих
потенциалов от нескольких афферентов.
2. возрастание частоты импульсации по
афферентам вызывает временную
суммацию и происходит гомосинаптическая
модуляция.
64. Рефлекторное кольцо
Рефлекторная реакция предполагаетвзаимодействие ЦНС с периферическими
органами по принципу обратной связи для
более точной координации и
приспособления.
ОБРАТНАЯ
АФФЕРЕНТАЦИЯ
ДВИГАТЕЛЬНАЯ
КОМАНДА
65. Физиология рецепторов
Рецепторы
являются
функциональными
и
структурными образованиями, обеспечивающими
восприятие информации о факторах внешней и
внутренней среды.
Морфологически рецепторы приспособлены к
восприятию
адекватного
специализированного
раздражения, порог к которому низок.
66. Классификация рецепторов
По модальности раздражения:Фоторецепторы,
терморецепторы,
механорецепторы, хеморцепторы, ноцицепторы.
По местоположению:
Экстрорецепторы
дистантные
экстрорецепторы
(зрительные,
слуховые, обонятельные)
- контактные экстрорецепторы (механо, вкусовые и
др.).
Интерорецепторы
Проприорецептор
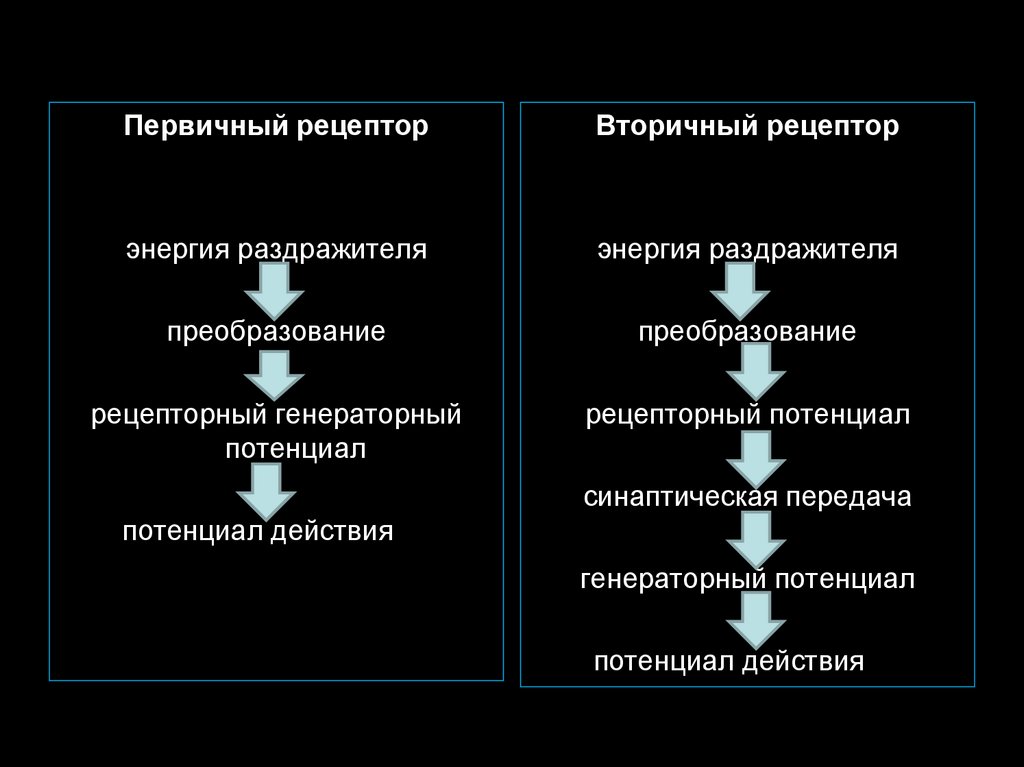
67. Первичные и вторичные рецепторы
Первичная реакция любого рецептора состоит в генерации рецепторного
потенциала. Он возникает в ответ на действие раздражителя. При этом
рецептором является или нервное окончание – первичный рецептор, или
сенсорный эпителий – вторичный рецептор.
Рецепторный потенциал в случае первичного рецептора является
одновременно и генераторным, от величины которого зависит частота и
продолжительность генерации ПД, поступающих в ЦНС.
В случае вторичного рецептора рецепторный потенциал через
синаптическую передачу вызывает образование генераторного потенциала на
сенсорном нейроне и последующую генерацию ПД (фоторецептор, рецепторы
кортиева органа).
68.
Первичный рецепторВторичный рецептор
энергия раздражителя
энергия раздражителя
преобразование
преобразование
рецепторный генераторный
потенциал
рецепторный потенциал
синаптическая передача
потенциал действия
генераторный потенциал
потенциал действия
69. ФУНКЦИОНАЛЬНЫЕ МЕЖНЕЙРОННЫЕ ОБРАЗОВАНИЯ В ЦНС
• НЕРВНЫЕ ЦЕПИ• НЕЙРОННЫЕ СЕТИ
• НЕЙРОННЫЕ АНСАМБЛИ (модули)
• НЕРВНЫЕ
ЦЕНТРЫ
70. Нейронные сети
В нейронную сеть, в отличие от рефлекторной дуги, включаютсякак последовательные, так и параллельные нервные цепи,
которые между собой взаимодействуют.
Нейронные сети выполняют сложные задачи по обработке
информации (сенсорные цепи).
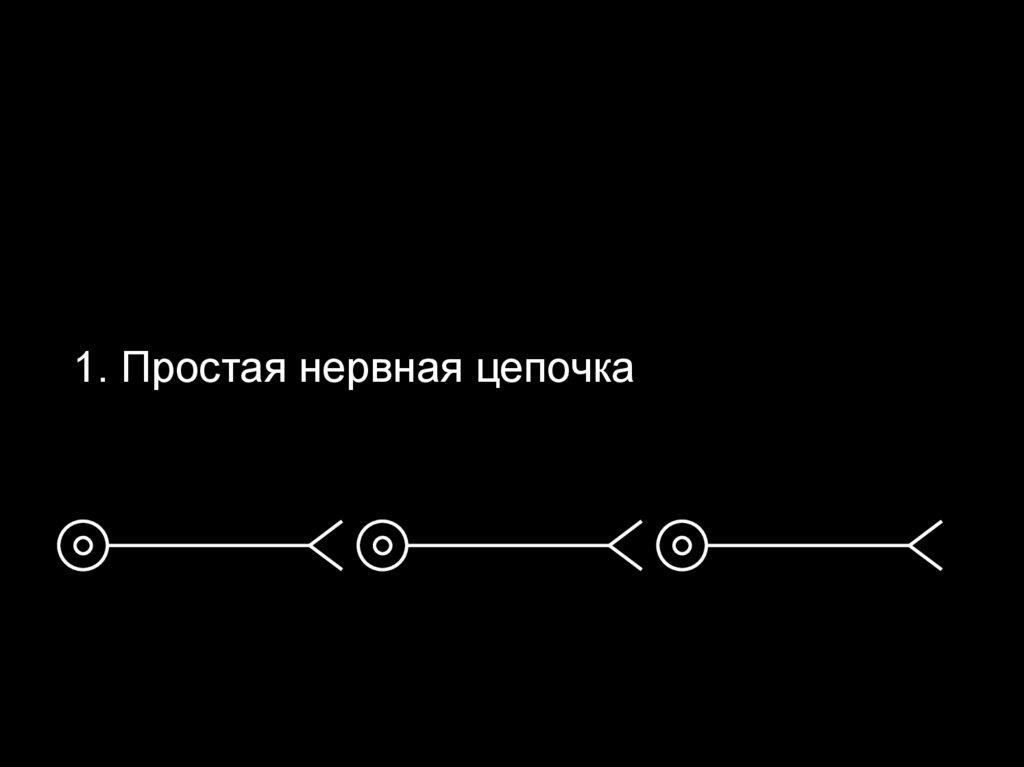
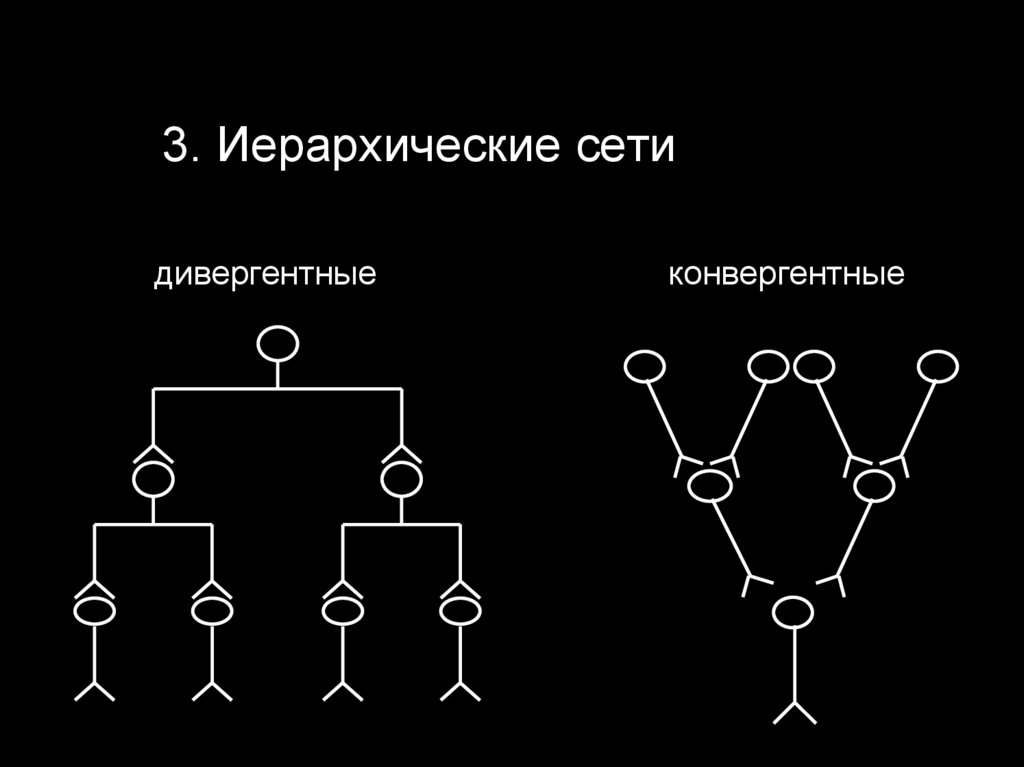
Различают 3 типа нейронных сетей:
1. Простые нервные цепочки
2. Локальные сети
3. Иерархические сети
71.
1. Простая нервная цепочка72.
2. Локальные сети (Леренто де Но).Реверберация возбуждения ограничивается
тормозным сигналом или попаданием в фазу
рефрактерности.
73.
3. Иерархические сетидивергентные
конвергентные
74. НЕЙРОННЫЕ АНСАМБЛИ
Нейронный ансамбль или модуль – представляет собой совокупность
локальных нейронных сетей, которая обрабатывает информацию,
подвергает трансформации, передает ее на выход.
Один модуль может входить в состав различных функциональных
образований.
Характерные признаки нейронного ансамбля:
1. Локальный синергизм реакции нейронов центральной зоны;
2. Наличие тормозной окантовки;
3. Наличие
определенного числа нейронов со стабильными
ответами
на
адекватное
афферентное
раздражение
(располагаются в центральной ядерной зоне ансамбля).
75. модуль
76. НЕРВНЫЙ ЦЕНТР
Это физиологическая структурная единица ЦНССуществует 2 понятия НЦ:
Анатомическое понятие:
НЦ – это группа нейронов, расположенная в определенном отделе
ЦНС и необходимая для осуществления рефлекса.
Физиологическое понятие:
НЦ – это совокупность нейронов, расположенных на различных
этажах ЦНС и принимающих участие в регуляции
определенной функции.
77. Разделяют нервные центры по функциям:
1.чувствительные (слуховой, зрительный,и т.п. )
2.вегетативные (сердечный,
дыхательный, пищеварительный и т.п.)
3.двигательные
4.центры психических функций (речи,
эмоций)
78. Нервные центры обладают рядом свойств и особенностей которые зависят от:
1. свойств нейронов,2. взаимосвязи нейронов,
3. свойств синапсов.
79. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
1. Одностороннее проведение возбуждения.2. Центральная задержка рефлекса.
3. Последействие (кратковременное и реверберация)
4. Суммация: пространственная и временная. Центральное
облегчение и окклюзия.
5. Трансформация ритма.
6. Фоновая электрическая активность.
7. Тонус нервного центра (открыт Бронжестом) .
8. Утомление нервных центров.
9. Высокая чувствительность к гипоксии и глюкозе .
10. Посттетаническое усиление.
11. Пластичность (пресинаптическая и постсинаптическая;
гомосинаптическая и гетеросинаптическая).
80. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
1. одностороннее проведение возбуждения(химические синапсы),
2. задержка проведения возбуждения
(химические синапсы),
81.
3. Последействие :1. кратковременное - связано с особенностью
синаптической передачи (медиатор не сразу
прекращает действовать, а лишь через время
(около 10 мс))
2. длительное - связано с циркуляцией
возбуждения по сетям
Лоренто де Но
82. 4. Временная и пространственная суммация
83. Центральное облегчение: при одновременном раздражении двух афферентов с перекрывающимися подпороговыми зонами количество
возбужденных нейронов больше, чем арифметическаясумма возбужденных нейронов при их раздельном
раздражении
А
1
При раздражении А
возбуждаются 2
нейрона (1,2)
2
3
При раздражении В
возбуждаются 2
нейрона (5, 6)
В
4
При раздражении А + В
возбуждаются 6 нейронов
(1, 2, 3, 4, 5, 6)
5
6
84.
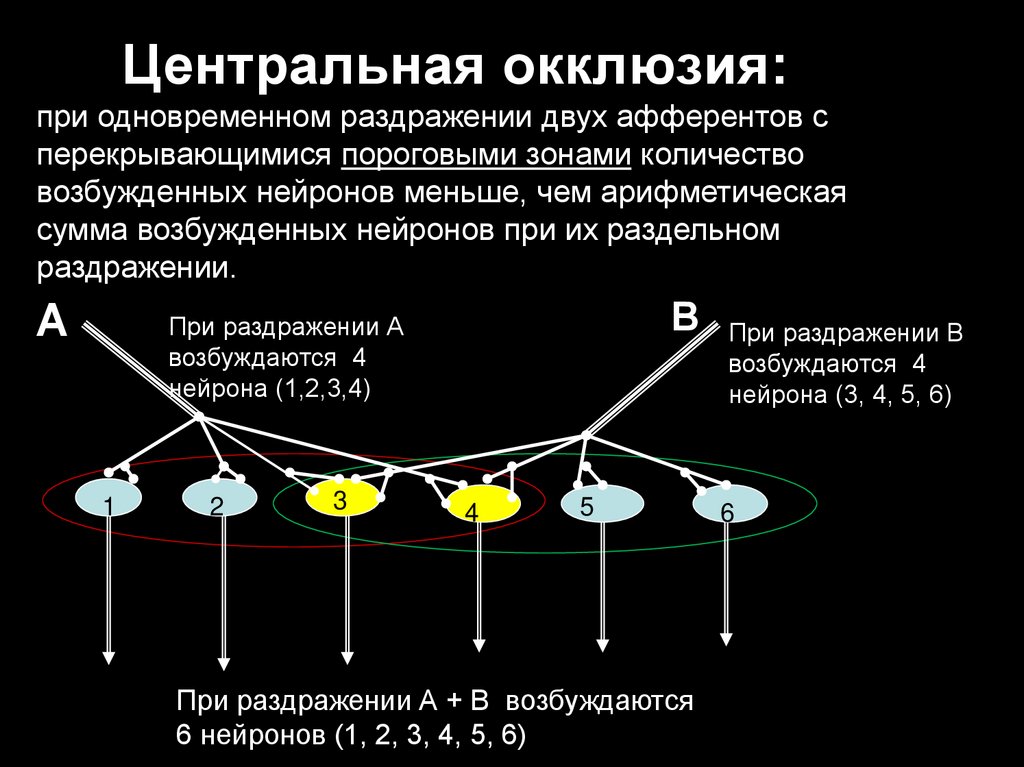
Центральная окклюзия:при одновременном раздражении двух афферентов с
перекрывающимися пороговыми зонами количество
возбужденных нейронов меньше, чем арифметическая
сумма возбужденных нейронов при их раздельном
раздражении.
А
В
При раздражении А
возбуждаются 4
нейрона (1,2,3,4)
1
2
3
4
5
При раздражении А + В возбуждаются
6 нейронов (1, 2, 3, 4, 5, 6)
При раздражении В
возбуждаются 4
нейрона (3, 4, 5, 6)
6
85.
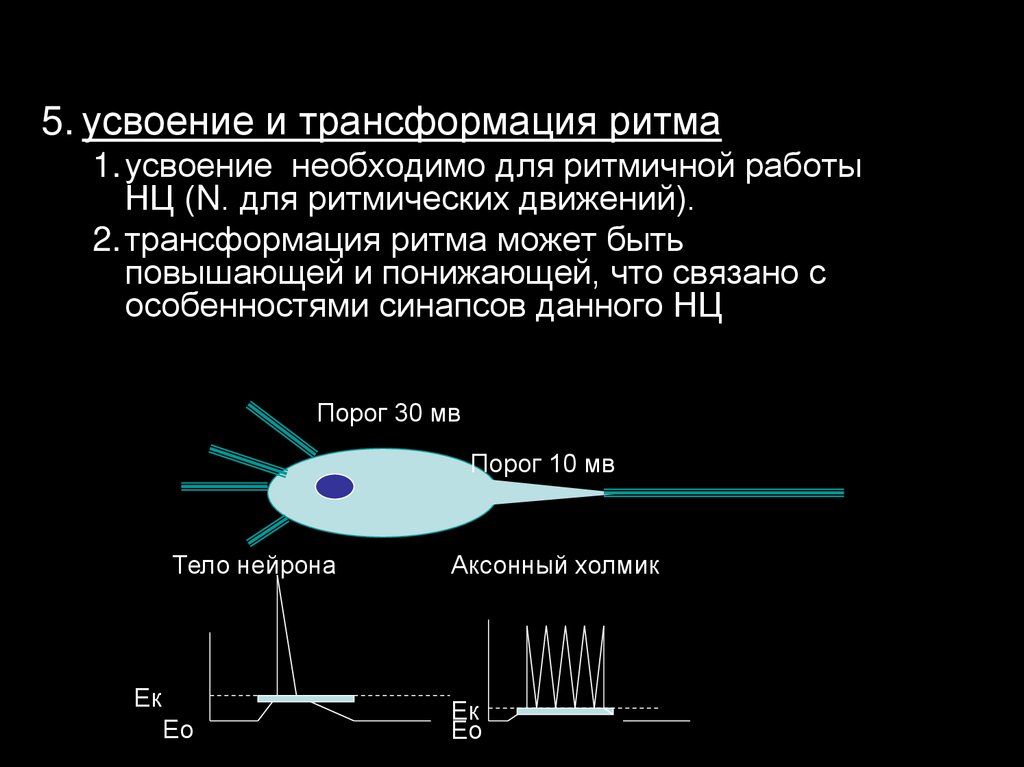
5. усвоение и трансформация ритма1. усвоение необходимо для ритмичной работы
НЦ (N. для ритмических движений).
2. трансформация ритма может быть
повышающей и понижающей, что связано с
особенностями синапсов данного НЦ
Порог 30 мв
Порог 10 мв
Тело нейрона
Ек
Ео
Аксонный холмик
Ек
Ео
86.
6. фоновая активность обеспечивается:поступлением импульсов от соседних НЦ
(дивергенция, конвергенция)
суммацией миниатюрных потенциалов,
возникающих за счет утечки медиатора
7. тонус НЦ необходим для обеспечения
готовности к рефлекторной деятельности
и возникает:
1. за счет спонтанной активности нейронов НЦ,
2. из-за постоянного поступления импульсации от
рецепторов с переферии
87. Опыт Бронжеста
Тонус нервного центаОпыт Бронжеста
88.
8. утомление в НЦ возникает из-за:1.
2.
3.
4.
истощения медиатора,
истощения АТФ и глюкозы,
снижения чувствительности рецепторов,
метаболических сдвигов в нейронах.
9. Высокая чувствительность к О2
(необратимые последствия: кора – 5-6
минут, ствол – 15-20 минут, спинной мозг –
30 минут) , глюкозе.
89.
10. Посттетаническая потенциацияЕсли , после интенсивного раздражения, дать синапсу отдохнуть и
нанести одиночное раздражение, то медиатора выбросится
значительно больше и ответная реакция возрастёт.
Это явление называется посттетанической потенциацией.
Причины:
- Во время тетанического раздражения усиливаются синтетические
процессы.
- Увеличивается количество везикул, готовых к высвобождению.
- Небольшого Са2+-тока достаточно для массивного выброса
медиатора в синаптическую щель.
90.
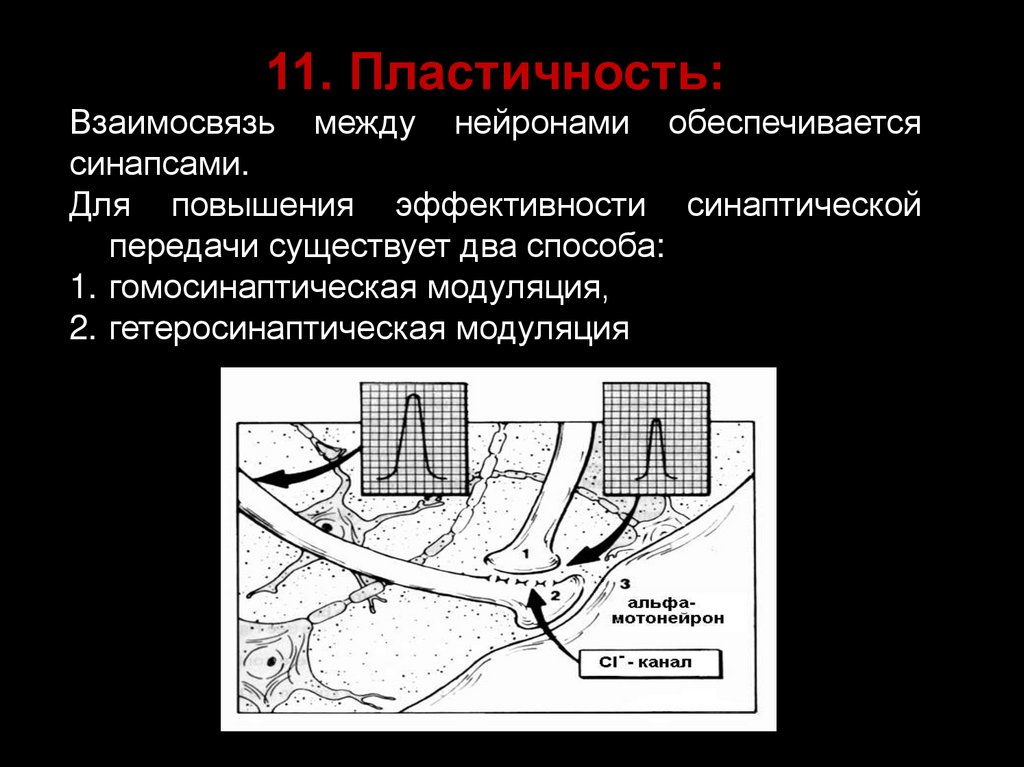
11. Пластичность:Взаимосвязь между нейронами обеспечивается
синапсами.
Для повышения эффективности синаптической
передачи существует два способа:
1. гомосинаптическая модуляция,
2. гетеросинаптическая модуляция
91. Гомосинаптическая модуляция
частое (тетаническое) раздражениепресинаптической мембраны
↓
повышение [Са2+ ] в синаптическом окончании
↓
увеличение выделения медиатора
↓
увеличение амплитуды и количества ПСП
Это явление тетанической потенциации.
92. Гетеросинаптическая модуляция
Этапы:Выделение медиатора
→↑ цАМФ
→ инактивация S- К+-каналов
→ удлинение фазы
деполяризации ПД
→ больше входящего Са2+
→ больше выделение
медиатора
→ больше амплитуда ПСП