


































")
























 при рН 7,0 можливих акцепторів електронів при диханні")
")


")
")

















 Биология
БиологияПохожие презентации:


")







Метаболізм мікроорганізмів
1.
2.
Метаболізм – це сукупність хімічних реакцій, яківідбуваються за участі специфічних ферментних
систем і забезпечують процеси життєдіяльності,
розвитку та розмноження організму.
Метаболізм є сумарним результатом двох процесів:
катаболізму та анаболізму.
Катаболізм – це сукупність ферментативних реакцій,
пов’язаних із розщепленням (окисненням) субстрату
до простих кінцевих продуктів, що супроводжуються
утворенням відновлених еквівалентів та виділенням
енергії, яка запасається у АТФ.
Основна мета реакцій катаболізму – це забезпечення
клітини енергією.
Анаболізм – це сукупність енергозалежних біохімічних
процесів, спрямованих на синтез простих молекул,
полімерів та структурних компонентів клітини.
Процеси анаболізму супроводжуються окисненням
відновлених еквівалентів та гідролізом АТФ.
Основна мета реакцій анаболізму – це біосинтез.
3.
Для мікроорганізмів характерна різноманітність типівметаболізму.
Завдяки різноманітним типам метаболізму
мікроорганізми:
• виживають у різних умовах довкілля,
• легко пристосовуються до зміни зовнішніх чинників
середовища,
• мають надзвичайно широкий ареал розповсюдження.
Тип метаболізму мікроорганізмів визначається трьома
показниками:
енергією, необхідною для активного біосинтезу;
електронами, необхідними для функціонування
електрон-транспортних ланцюгів та утворення
відновлених еквівалентів (НАДН2);
вуглецем - основним компонентом клітини.
4.
За джерелом енергії мікроорганізми поділяють на:• хемотрофи (одержують енергію з хімічних реакцій)
• фототрофи (одержують енергію при перетворення
енергії світла).
За походженням джерела (донора) електронів
мікроорганізми поділяють на:
• органотрофи (одержують електрони з органічних
сполук: цукрів, орг. кислот, спиртів, амінокислот).
• літотрофи – одержують електрони з неорганічних
сполук: амонію, нітриту, сульфіду, молекулярного
водню, заліза-ІІ.
За джерелом вуглецю мікроорганізми поділяють на:
• автотрофи (споживають неорганічний вуглець:
діоксид вуглецю, карбонати.
• гетеротрофи – споживають органічний вуглець:
цукри, орг.кислоти, спирти, амінокислоти.
5. Типи метаболізму
Загальна назва типу метаболізму мікроорганізмів,складається з 4 коренів
1. перший корінь указує на джерело енергії, яку
використовує мікроорганізм;
2. другий корінь – на походження електронів;
3. третій корінь – на джерело вуглецю
4. четвертий корінь походить від грецького trophe –
живлення.
Наприклад, хемо-літо-авто-трофи
6. Типи метаболізму мікроорганізмів
Джерела енергіїЕнергія хімічних реакцій
Джерела
(донори)
електронів
Органічні
речовини
Джерела вуглецю
Органічні
речовини
Хемоорганогетеротрофи
Неорганічні
речовини
Хемоорганоавтотрофи
Енергія
світла
Неорганічні Хемоліторечовини
гетеротрофи
Хемолітоавтотрофи
Органічні
речовини
Фотоорганоавтотрофи
Фотоорганогетеротрофи
Неорганічні Фотоліторечовини
гетеротрофи
Фотолітоавтотрофи
7. Для здійснення метаболічних реакцій клітині потрібний ще й акцептор електронів
Термінальні акцептори електроніву мікроорганізмів
Молекулярний
кисень
(використовують аероби)
Інші окиснені сполуки
(використовують анаероби)
Неорганічні
Органічні
8. Катаболізм аеробних хемоорганотрофів
9.
Аеробні хемоорганотрофивикористовують:
• Джерело енергії – хімічні реакції
• Донор електронів – органічні речовини
• Термінальний акцептор електронів –
молекулярний кисень
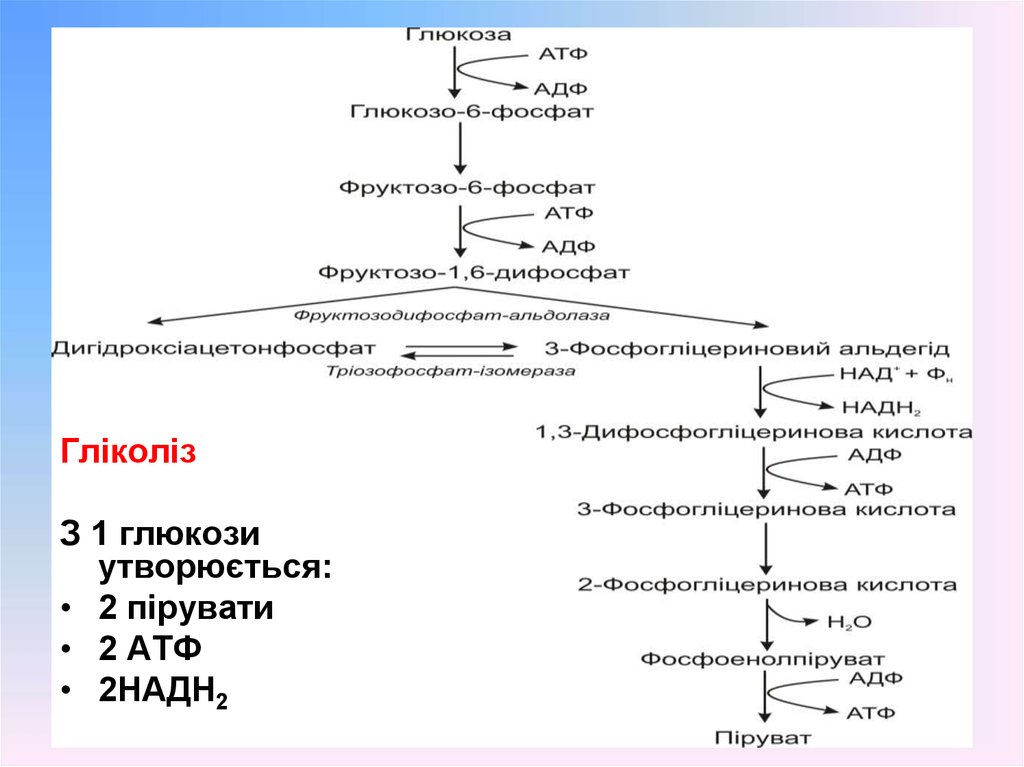
10. Етапи катаболічного перетворення гексоз у мікробній клітині
11. Етапи катаболічного перетворення гексоз у мікробній клітині
Етап А: активація молекули глюкози фосфатноюгрупою. Цей етап пов’язаний із затратою енергії. У
бактерій фосфорилювання глюкози відбувається при
її транспорті у клітину.
Етап В:. Розщеплення глюкозо-6-фосфату до пірувату
у мікроорганізмів може відбуватися трьома
способами:
• гліколітичним шляхом
• шляхом Ентнера-Дудорова
• пентозофосфатним шляхом
12.
ГліколізЗ 1 глюкози
утворюється:
• 2 пірувати
• 2 АТФ
• 2НАДН2
13. Етапи катаболічного перетворення гексоз у мікробній клітині
Етап С: Окиснення пірувату до ацетил-КоА.Піруват + КоА + НАД+ → ацетил-КоА + НАДН2 + СО2
Етап D: Окиснення ацетильної групи ацетил-КоА до
СО2 у циклі Кребса
Призначення циклу Кребса – розщеплення ацетил-КоА
до 2 СО2.
Крім того при окисненні 1 ацетил-КоА утворюється: 3
НАДН2, 1 ФАДН2, 1 ГТФ (фактично 1 АТФ).
Такі проміжні продукти як α-кетоглутарат та
оксалоацетат використовуються у анаболізмі.
14. Цикл Кребса
ЩУКа съела ацетат, получается цитратчерез цисаконитат будет он изоцитрат
водороды отдав НАД, он теряет СО2
этому безмерно рад альфа-кетоглутарат
окисление грядет — НАД похитил водород
ТДФ, коэнзимА забирают СО2
а энергия едва в сукциниле появилась
сразу АТФ родилась и остался сукцинат
вот добрался он до ФАДа — водороды тому
надо
фумарат воды напился, и в малат он
превратился
тут к малату НАД пришел, водороды
приобрел
ЩУКа снова объявилась и тихонько затаилась
15. Етап Е: дихальний ланцюг
Дихальний ланцюг – цесукупність великої кількості
переносників і ферментів, які
фізично зв’язані з матрексом
мембран і занурені в їх
ліпідний шар.
За хімічною природою
переносники поділяються на:
• Флавопротеїни
• Залізо-сірчані білки
• Хінони
• Цитохроми
у прокаріот дихальний ланцюг
розміщений на ПМ та
мезосомах.
Місця винесення протонів через
мембрану називають
пунктами
фосфорилювання.
Їх максимальна кількість - 3.
16. Хеміосматична гіпотеза Мітчела
• На дихальний ланцюг надходять атоми водню з НАДН2 таФАДН2;
• Переносники у дихальному ланцюзі розміщені по мірі
зростання їх окисно-відновного потенціалу від -320 мВ (ОВП
системи НАДН2/НАД+) до +810 мВ (ОВП системи ½ О2/Н2О);
• Чергування переносників призводить до транслокації протонів
через мембрану з цитоплазми у периплазматичний простір (у
прокаріот);
• На мембранах виникає градієнт електрохімічного
потенціалу іонів водню (ΔμН+), який є рушійною силою
процесу утворення АТФ;
• Протони повертаються назад через мембрану лише у місцях
локалізації АТФ-синтетази (АТФ-ази). За рахунок цього
відбувається реакція фосфорилювання АДФ до АТФ: АДФ +
Фн + Н+ = АТФ + Н2О
17. Два способи утворення АТФ
Таким чином у аеробних хемоорганогетеротрофів існуєдва способи синтезу АТФ:
1. Субстратне фосфорилювання;
2. Окисне фосфорилювання.
Субстратне фосфорилювання – це процес синтезу
АТФ в результаті перетворення органічної
речовини з однією форми в іншу, наприклад, у
гліколізі та циклі Кребса.
Окисне фосфорилювання – це синтез АТФ за рахунок
градієнту електрохімічного потенціалу іонів водню
на АТФ-синтетазі.
18. Неповне окиснення органічних сполук аеробними хемоорганогетеротрофами
Типовими бактеріями, що здійснюютьнеповне окиснення є оцтовокислі
бактерії родів Acetobacter та
Gluconobacter, які окиснюють спирт та
цукор з утворенням оцтової кислоти.
У Gluconobacter окиснення ацетил-КоА у
циклі Кребса неможливе через
відсутність ферментів: α-кетоглутаратдегідрогенази, ізоцитратліази та
малатсинтетази.
Бактерії роду Acetobacter мають повний
цикл Кребса. Однак, спочатку вони
окиснюють весь етанол до ацетату,
який накопичується у середовищі. Після
повного використання етанолу бактерії
починають окиснювати ацетат.
Acetobacter використовують для
виробництва оцту.
Gluconobacter
Acetobacter aceti
19. Катаболізм аеробних хемолітотрофів
20.
Літотрофнімікроорганізми
використовують як
джерело (донор)
електронів неорганічні
сполуки у
відновленому стані:
Н2,
NH4+,
NO2-,
S0,
S2-,
CO,
Fe2+
21.
В залежності від донора електронів серед аеробниххемолітотрофів розрізняють:
водневі бактерії ( окиснюють молекулярний водень)
нітрифікуючі бактерії (окиснюють амоній та нітрит)
безколірні сіркобактерії (окиснюють сульфід та сірку)
тіонові бактерії (окиснюють сульфід)
карбоксидобактерії (окиснюють монооксид вуглецю)
залізобактерії (окиснюють залізо-ІІ)
22. Загальний принцип катаболізму хемолітотрофів
Метаболізм аеробних хемолітотрофів характеризуєтьсячотирма особливостями:
1. АТФ утворюється лише в процесі окисного
фосфорилювання, тобто на дихальному ланцюзі;
2. Реакцій субстратного фосфорилювання зазвичай
відсутні;
3. Електрони від донорів надходять на дихальний
ланцюг не на початок (флавопротеїни), а на рівні
цитохромів (виключення водневі бактерії)
4. Транспорт електронів по дихальному ланцюгу
відбувається у двох напрямках:
• прямому ( до О2, для отримання АТФ);
• зворотньому ( із затратою енергії, для відновлення
НАД+ і отримання НАДН2 для біосинтезу).
23. Загальний принцип катаболізму хемолітотрофів
24. Водневі бактерії
Водневі бактерії це аеробні хемолітоавтотрофніпрокаріоти, які здатні:
• отримувати енергію та електрони, шляхом окиснення
молекулярного водню;
• всі речовини клітини будувати з вуглецю СО2.
Рівняння метаболізму водневих бактерій виглядає
наступним чином:
6Н2 + 2О2 + СО2 → <СН2О> + 5Н2О
Водневі бактерії не є таксономічною групою. До них
належать представники 20 родів, які мають різну
морфологію, різне забарвлення за Грамом, рухливі і
нерухливі, спорові і неспорові форми.
Вони можуть використовувати як джерело вуглецю та
електронів також і різноманітні органічні сполуки.
25. Нітрифікатори
Нітрифікація – це процесмікробного аеробного окиснення
амонію до нітрату.
На першій фазі нітрифікації
відбувається окиснення амонію
(NH4+) до нітриту (NO2-), тобто
N3- перетворюється на N3+.
Представники: Nitrosomonas,
Nitrosococcus, Nitrosolobus,
Nitrosospira, Nitrosovibrio.
На другій фазі нітрифікації
відбувається окиснення нітриту
(NО2-) до нітрату (NO3-), тобто
N3+ перетворюється на N5+.
Представники: Nitrobacter,
Nitrospina, Nitrococcus, Nitrospira.
1 – Nitrosomonas sp.;
2 – Nitrosomonas europeae ;
3 – Nitrobacter sp.;
4 – Nitrobacter winogradskyi.
26. Аеробні бактерії, що окиснюють сульфід S2-
Аеробні бактерії, щоокиснюють сульфід S2можна поділити на 2 групи:
1. Тіонові бактерії
(хемолітотрофний метаболізм,
не накопичують S0)
Представники:Thiobacillus (4-6),
Thiomicrospira (7-8),
Paracoccus (9), Thiodendron
2. Безколірні сіркобактерії
(хемоорганотрофний
метаболізм, накопичують S0
в периплазмі)
Представники: Achromatium (1),
Macromonas,Thiothrix (2),
Thioploca (3), Thiospirillopsis,
Beggiatoa (10)
27. Розповсюдження бактерій, що окиснюють сполуки сірки
У гарячих джерелах термофільнітіобацили окиснюють сульфіди та
залізо-ІІ (національний парк
Yellowstone, США)
28. Катаболізм фототрофів
29.
Фотосинтез (фотофосфорилювання) – цепроцес одержання організмами енергії за
рахунок світла, або це перетворення енергії
світла у енергію хімічних зв’язків.
Для фотосинтезу найбільше значення має
видимий спектр сонячного світла, а саме
діапазон хвиль від 400 до 700 нм.
Ультрафіолетові промені несуть у собі
занадто багато енергії, тому вони руйнують
органічні молекули (на цьому грунтується
використання УФ-опроміненння для
дезинфекції), а інфрачервоні – мають
недостатньо енергії для генерації хімічних
реакцій і практично одразу ж перетворюються
на тепло.
30. Оксигенний та аноксигенний фотосинтези
За механізмом фотосинтез розподіляють на два типи:• Аноксигенний фотосинтез не супроводжується
виділенням молекулярного кисню. Цей більш древній
процес характерний для анаеробних прокаріот –
пурпурних бактерій, зелених бактерій, геліобактерій
та аеробних аноксигенних фотосинтетиків.
Особливим типом фотосинтезу, пов’язаним з
каротиноїдами, характеризуються представники
архебактерій – галобактерії.
• Оксигенний фотосинтез, при якому молекулярний
кисень виділяється в атмосферу, характерний для
двох груп прокаріотичних організмів – ціанобактерій
та прохлорофітів, а також для всіх еукаріот –
водоростей та вищих рослин.
31. Пігменти
Здатність організмів існувати зарахунок енергії світла
пов’язана з наявністю у них
специфічних фоторецепторних
молекул – пігментів.
Всі фотосинтезуючі пігменти
належать до сполук двох
хімічних класів:
пігменти, в основі яких лежить
тетрапірольна структура
(хлорофіли, бактеріохлорофіли
та фікобіліпротеїни) (Рис.вгорі)
пігменти, основу яких
складають довгі
поліізопреноїдні ланцюги
(каротиноїди) (Рис. внизу)
Пігменти
32. Хлорофіли та бактеріохлорофіли
• Хлорофіли (Хл) – це магній-порфіринові пігменти.Вони складаються з чотирьох, з’єднаних між собою
пірольних кілець. Утворюють закриту циліндричну
структуру. Містять метал (магній), що з’єдується з
атомами азоту пірольних кілець за допомогою
ковалентних та координаційних зв’язків. Ці пігменти
характерні для організмів, що здійснюють оксигенний
фотосинтез.
• Хлорофіли прокаріот, що здійснюють аноксигенний
фотосинтез, звуться бактеріохлорофілами (Бхл).
Ідентифіковано 6 типів бактеріохлорофілів ( a, b, c, d,
e, g) та два типи хлорофілів (a та b). Хлорофіли та
бактеріохлорофіли забезпечують поглинання світла в
області 650-1035 нм.
33. Фікобіліпротеїни та каротиноїди
• Фікобіліпротеїни – червоні і сині пігменти, Щоскладаються з білкової субодиниці та простетичної
групи – фікобілінів. Фікобіліни мають чотири
пірольних кільця, але утворюють назамкнену
структуру і не містять метал. Їх називають
тетрапіролами з відкритими ланцюгами.
Фікобіліпротеїни поглинають фотони у діапазоні 480650 нм і виконують функцію світлозбиральних
молекул.
• Каротиноїди – це велика група хімічних сполук, що є
продуктом конденсації, як правило, 8 залишків
ізопрена. Каротиноїди забезпечують поглинання
світла в області 400-550 нм та запобігають шкідливим
для клітини реакціям фотоокиснення.
34. Локалізація фотосинтетичного апарату прокаріот
Прокаріоти, на відміну відеукаріот, не мають
спеціальних органел
для фотосинтезу
(хлоропластів).
У пурпурних бактерій та
геліобактерій
фотосинтетичний
апарат локалізований у
ПМ та
внутрішньоцитоплазм
атичних мембранах.
У зелених бактерій - в ПМ
та особливих
структурах хлоросомах.
У ціанобактерій - у ПМ та
фікобілісомах, подібних
за будовою до
хлоросом.
Хлоросома.
35. Будова фотосинтетичного апарату
Фотосинтетичний апарат мікроорганізмівскладається з трьох копмонентів:
1. Світлозбиральні антени, що поглинають
енергію світла і передають її до фотохімічних
реакційних центрів. Їх ще називають
фоторецепторними, світлозбиральними або
антенними молекулами.
2. Фотохімічні реакційні центри (РЦ), де
відбувається трансформація
електромагнітної форми енергії у хімічну.
3. Фотосинтетичні електронтранспортні
системи, що забезпечують перенесення
електронів, пов’язане із синтезом АТФ.
36. Світлозбиральні антени
Пігменти, що входять до складу антен, поглинаютьсвітлову енергію і переходять у збуджений
синглетний стан.
Енергія цього стану передається на інші молекули
пігментів і досягає реакційного центру.
Частина енергії губиться у вигляді тепла чи
флуоресценції.
До світлозбиральних антен входять:
• значна частина хлорофілів (бактеріохлорофілів);
• каротиноїди;
• фікобіліпротеїни (тільки у бактерій, що здійснюють
оксигенний фотосинтез).
37. Фотохімічні реакційні центри (РЦ)
До складу фотохімічного реакційного центру входить:• Частина хлорофілу a (оксигенний фотосинтез)
• Частина бактеріохлорофілу a, b, g (аноксигенний
фотосинтез).
Це незначна частина від загальної кількості пігментів у
клітині:
• у аноксигенних фотосинтетиків це - 1/25-1/50 частина
всього бактеріохлорофілу;
• у оксигенних фотосинтетиків (ціанобактерії) це – 1/301/100 частини хлорофілу.
Під дією кванта світла хлорофіл збуджується (електрон
переходить на вищий синглетний рівень), а ОВП РЦ
зменшується з +500 до -700 мВ. Хлорофіл РЦ легко
віддає електрон.
38. Фотосинтетичні електронтранспортні системи
Здійснюють перенесення електрону, що відщепився відзбудженого хлорофілу (бактеріохлорофілу) у РЦ,
через систему переносників у двох напрямках:
1. Циклічний транспорт електронів - назад на
хлорофіл (бактеріохлорофіл) з утворенням градієнту
електрохімічного потенціалу іонів водню (ΔμН+) та
подальшим синтезом АТФ (фотофосфорилювання).
2. Нециклічний транспорт електронів – на НАД+ з
метою отримання відновлених еквівалентів для
біосинтезу
Електронтранспортна система містить комплекс
цитохромів bc1, цитохром c2, хінони,залізо-сірчані
білки.
39. Екзогенні донори електронів
Незалежно від напрямку руху електрона, що відірвався відбактеріохлорофілу, клітина відчуває недостачу електронів:
• при циклічному транспорті не вистачає електронів для
утворення відновлених еквівалентів,
• при нециклічному транспорті – для заповнення електронної
“вакансії” бактеріохлорофілу.
Тому фототрофні організми потребують додаткових джерел
електронів – екзогенних донорів електронів.
Екзогенними донорами електронів можуть бути:
• органічні сполуки (у фотоорганотрофів),
• нерганічні сполуки (у фотолітотрофів).
Неорганічними екзогенними донорами електронів
• при аноксигенному фотосинтезі є відновлені сполуки сірки
(S2-, SO32-, тіосульфат, тетратіонат, тіогліколят), а також
молекулярний водень.
• при оксигенному - вода
40. Загальна схема аноксигенного фотосинтезу
Позначення:РЦ- реакційний центр;
РЦ* - реакційний центр
зі збудженим
бактеріохлорофілом;
Бфф –
бактеріофеофітин;
Q – убіхінон;
bc1,c – цитохроми;
І-V – шляхи переносу
електронів від
бактеріохлорофілу та
екзогенних донорів.
41. Загальна схема оксигенного фотосинтезу
При оксигенномуфотосинтезі
екзогенним
донором
електронів
служить вода
Для здійснення
фотолізу води
необхідна
додаткова
фотосистема ІІ
При фотолізі води
виділяється
молекулярний
кисень
42.
Пурпурні сіркові1 – Chromatium vinosum;
2 – Thiospirillum jenense;
3 – Thiopedia rosea;
4 - Chromatium sp.;
5 – Thiocystis sp.;
6 – Thiocapsa liforalis;
Пурпурні несіркові
7 – Rhodospirillum
8 – Rhodobacter
9 – Rhodomicrobium
Зелені сіркові
10 – Chlorobium limicola;
11 – Prosthecochloris
12 – Pelodictyon
Геліобактерії
13 – Heliobacterium
Галобактерії
14 – Halobacterium
Аеробні аноксигенні
фотосинтетики
15 – Sandaracinobacter
16 –Roseococcus
43. Розповсюдження пурпурних сіркобактерій
44. Розповсюдження зелених сіркових та несіркових бактерій
45. Бактерії, що здійснюють оксигенний фотосинтез
1, 2 – Prochloron didenmiна тканинах осцидій;
3 – Prochloron didenmi;
4 – Nostoc sp.;
5 – Microcystis sp.;
6 – Spirulina sp.;
7 – Oscillatoria sp.;
8 – Anabaena sp.
46. Катаболізм анаеробів
47.
Для життєдіяльності багатьох мікроорганізмівмолекулярний кисень не є конче необхідним.
Роль термінального акцептора електронів у анаеробів
можуть виконувати ряд органічних та неорганічних
сполук.
За ставленням до кисню анаеробні мікроорганізми
розподіляють на :
• факультативно-анаеробні,
• облігатно анаеробні,
• аеротолерантні.
Анаероби
(за ставленням до кисню
Факультативноанаеробні
Облігатно
анаеробні
Аеротолерантні
48.
• Факультативно-анаеробні мікроорганізми живуть, якв атмосфері кисню, так і без нього. При наявності
молекулярного кисню у довкіллі, вони
використовують його як термінальний акцептор при
диханні. Якщо ж кисень відсутній, факультативні
анаероби легко переключаються на анаеробні
процеси - нітратне дихання або бродіння. Вони
містять супероксиддисмутазу та каталазу.
• Облігатно анаеробні мікроорганізми не
використовують кисень у своємо метаболізмі як
термінальний акцептор електронів при диханні. Їх
клітини позбавлені каталази, супероксиддисмутази та
пероксидази, тому вони гинуть в атмосфері кисню.
• Аеротолерантні анаероби – це мікроорганізми, які
не використовують кисень як термінальний акцептор
електронів, але мають деякі захисні механізми проти
пероксиду водню та супероксидного радикалу, тому
можуть існувати в аеробних умовах.
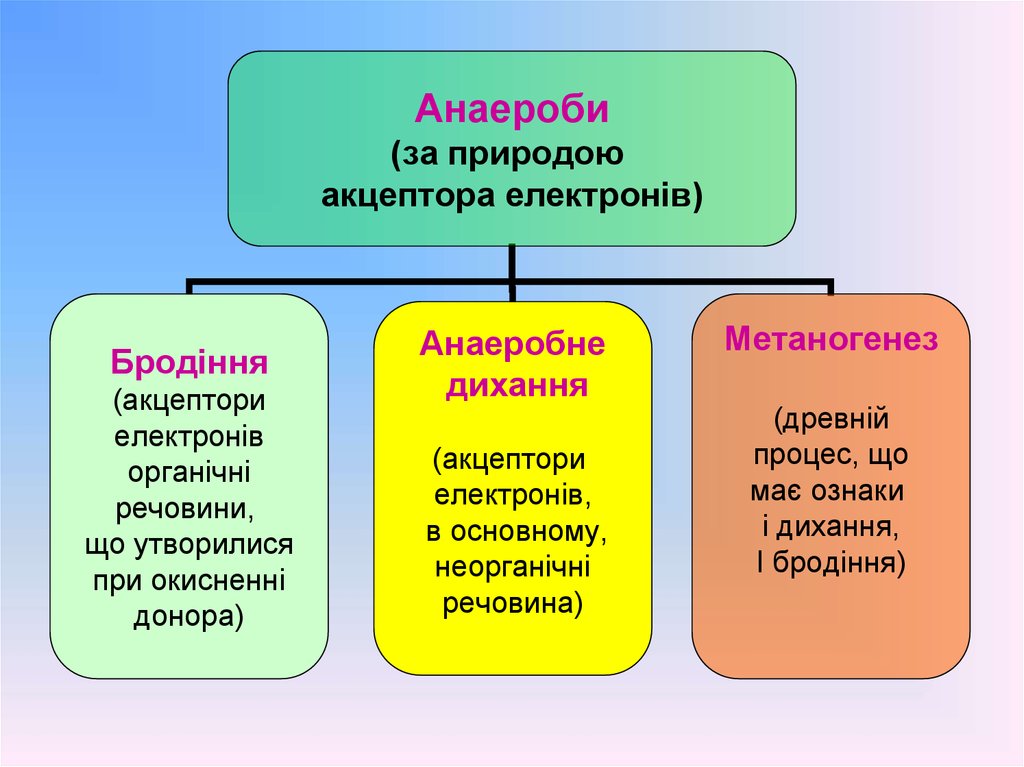
49.
Анаероби(за природою
акцептора електронів)
Бродіння
(акцептори
електронів
органічні
речовини,
що утворилися
при окисненні
донора)
Анаеробне
дихання
(акцептори
електронів,
в основному,
неорганічні
речовина)
Метаногенез
(древній
процес, що
має ознаки
і дихання,
І бродіння)
50. Сучасні методи роботи з облігатними анаеробами
Анаеробні боксиГерметичні пробірки
Анаеростати
51. Бродіння
52.
Бродіння – це спосіб отримання енергії, приякому АТФ утворюється у процесах
анаеробного окиснення органічних субстратів
в результаті субстратного фосфорилювання
без участі неорганічних акцепторів
електронів.
Це найдревніший і найбільш примітивний спосіб
отримання енергії.
Примітивність процесів бродіння полягає у
тому, що при розщепленні субстрату
вивільнюється лише незначна кількість
хімічної енергії, а більшість її залишається у
кінцевих продуктах
53. Бродіння – низько енергетичний процес
• Бактерії-бродильники отримують енергію лише зарахунок субстратного фосфорилювання.
• Цикл Кребса у них не бере участь у процесах
катаболізму, а дихальний ланцюг не функціонує, або
зовсім відсутній.
• Тому, при розщепленні 1 молекули глюкози,
синтезується, зазвичай, 2 молекули АТФ (для
порівняння, при аеробному диханні утворюється 38
АТФ на 1 глюкозу).
• Кількість реакцій, що призводить до безпосереднього
синтезу АТФ при бродінні досить обмежена:
54. Метаболіти, що утворюються при бродінні
Термінальними акцепторами водню у процесі бродіння, якправило, служать органічні метаболіти, що утворилися в
результаті розщеплення субстрату.
Відновлені сполуки, що приєднали до себе водень, виділяються з
клітини і накопичуються у середовищі у значних кількостях.
Це органічні кислоти, спирти та гази.
Назва типу бродіння визначається його кінцевими метаболітими:
• Молочнокисле (гомо- та гетероферментативне)
• Спиртове
• Гліцеринове
• Гліцеро-піровиноградне
• Бутандіолове
• Мурашинокисле
• Пропіоновокисле
• Маслянокисле
• Ацетон-бутанолове
Через низький енергетичний вихід процесів бродіння клітині
доводиться переробляти величезну кількість субстрату та
виділяти багато метаболітів.
55. Гомоферментативне молочнокисле бродіння
Глюкоза окиснюєтьсяшляхом гліколізу до:
2 молекул пірувату;
2 АТФ;
2 НАДН2.
Акцептором водню є
піруват.
Піруват приймає водень з
НАДН2 і відновлюється до
лактату.
Загальне рівняння:
Глюкоза → 2 Лактат + 2 АТФ
Гомоферментативне
молочнокисле бродіння
здійснюють:
Lactococcus lactis,
Pediococcus cerevisiae,
Lactobacillus acidophilus,
Lactobacillus bulgaricus
56. Гетероферментативне молочнокисле бродіння
Глюкоза окиснюється спочатку пентозофосфатним шляхом, а потім гліколізом.Акцепторами водню є піруват, ацетил-КоА та
оцтовий альдегід
Кінцеві продукти : лактат, ацетат, етанол, СО2.
Гетероферментативне молочнокисле бродіння
здійснюють:
Lactobacillus plantarum,
Lactobacillus brevis,
Leuconostoc mesenteroides.
57. Молочнокислі бактерії
1 – Lactobacillus brevis;2 – L. bulgaricus;
3 – L. casei;
4 – Bifidobacterium sp.;
5 – Pediococcus sp.;
6 – Leuconostoc sp.;
7 – Lactococcus lactis;
8 – Streptococcus faecalis
58. Спиртове бродіння
1. Глюкоза окиснюється шляхом гліколізу до 2 молекулпірувату; 2 АТФ; 2 НАДН2.
2. Піруват декарбоксилюється ( виділяється СО2) до
оцтового альдегіду, який і є акцептором водню.
3. Оцтовий альдегід приймає водень з НАДН2 і
відновлюється до етанолу.
Загальне рівняння: Глюкоза → 2 Етанол + 2 СО2+ 2 АТФ
59. Мікроорганізми, що здійснюють спиртове бродіння
1 – Saccharomyces cerevisiae; 2 – Zymomonas mobilis;3 – Sarcina ventriculi; 4 – Erwinia amylovora
60. Анаеробне дихання
61.
Анаеробне дихання – це процес окиснення органічнихта неорганічних субстратів при якому термінальними
акцепторами електронів є окиснені неорганічні
сполуки, наприклад, SO42-, SO32-, S0, NO3-, NO2-,
метали зі змінною валентністю (Fe3+, Mn4+), СО2;
металовмісні кисневі аніони (селенати, арсенати),
радіонукліди (U+6, Tc+7).
В залежності від природи термінального акцептора
електронів анаеробне дихання буває
• нітратним,
• сульфатним,
• сірковим,
• залізним,
• вуглецевим, тощо.
62. Дихальний ланцюг при анаеробному диханні короткий
Значення ОВП термінального акцептора електроніввизначає місце виходу електрона з дихального
ланцюга, а отже і кількість АТФ, що утворюється в
результаті окисного фосфорилювання.
Чим вище значення ОВП акцептора, тим більше енергії
отримує мікроорганізм за рахунок дихання.
Оскільки, ОВП термінальних акцепторів при
анаеробному диханні значно нижчі, ніж ОВП O2/H2O,
кількість АТФ тут утворюється менше, ніж при
аеробному диханні.
63. Стандартні окисно-відновні потенціали (Е0’) при рН 7,0 можливих акцепторів електронів при диханні
Окисно-відновна параЕ0’, В
Н+ / Н2
НСО3- / ацетат
НСО3- / СН4
-0,42
-0,35
-0,33
S0 / HSHSO3- / HSNO2- / NH4+
-0,27
-0.12
+0.34
NO2- / NO
NO3- / NO2Fe3+ / Fe2+
+0.35
+0.43
+0.77
64. Нітратне дихання (дисиміляційна нітратредукція)
Нітратне дихання – це процес використанняфакультативно-анаеробними мікроорганізмами нітрату
(NO3-), як термінального акцептора електронів при
анаеробному диханні.
Оскільки нітрат в результаті відновлюється, нітратне
дихання називають дисиміляційною нітратредукцією,
а мікроорганізми, що здійснюють цей процес –
нітратредукторами.
65. Донори електронів при нітратному диханні
Донорами електронів при нітратному диханні є:• органічні речовини (глюкоза, етанол, сукцинат,
бензоат),
• неорганічні речовини (молекулярний водень).
Нітратредуктори можуть бути
• хемоорганотрофами (Pseudomonas aeruginosa),
• хемолітотрофами (Hydrogenomonas agilis,).
Механізм окиснення органічних субстратів, як правило,
не відрізняється від тих, що мають місце за аеробних
умов.
АТФ нітратредуктори отримують в результаті:
• субстратного фосфорилювання;
• окисного фосфорилювання.
66. Два шляхи відновлення нітрату
При нітратному диханні бактерії відновлюють нітратдвома шляхами:
1. Через нітрит до газоподібних продуктів NO, N2O, N2.
Оскільки середовище при цьому позбавляється
азоту, цей процес зветься денітрифікацією.
Його здійснюють: Pseudomonas (P.aeruginosa, P.
fluorescens, P.stutzeri), Alcaligenes, Paracoccus,
Hyphomicrobium, Thiobacillus denitrificans), Bacillus.
2. Через нітрит до амонію. Цей процес має назву
дисиміляційна (дихальна) амоніфікація. Він
характерний для ряду бацил, ентеробактерій та
стрептококів.
67. Два шляхи відновлення нітрату (продовження)
Окисне фосфорилювання при нітратному диханніможна розділити на кілька складових:
1. Відновлення нітрату до нітриту (цей етап спільний
для асиміляційної амоніфікації і денітрифікації).
2. Відновлення нітриту до амонію (асиміляційна
амоніфікація).
3. Відновлення нітриту до газоподібних продуктів
(денітрифікація).
68. Сульфатне дихання (дисиміляційна сульфатредукція)
Сульфатне дихання (дисиміляційна сульфатредукція)– це процес використання облігатними анаеробними
бактеріями сульфату (SO42-), як термінального
акцептора електронів при анаеробному диханні.
Сульфатредуктори бувають:
• хемоорганогетеротрофами та
• хемолітоавтотрофами.
Донорами електронів для них є молекулярний водень,
форміат, ацетат, лактат, фумарат, сукцинат, пропіонат,
жирні кислоти С4-С20, метанол, етанол, пропанол.
Донорами вуглецю – органічні сполуки (названі вище) та
СО2.
69. Деякі сульфатредуктори
А – Desulfivibrio desulfuricans;B – Desulfovibrio gigas;
C – Desulfovibrio piger;
D – Desulfovibrio salexigens;
E – Desulfovibrio sp.;
F – Desulfobulbus propionicus,
G – Desulfobulbus marinus;
H – Desulfobacter postgatei;
I – Desulfobacter hydrogenophilus;
J – Desulfobacter latus,
K – Desulfobacter curvatus,
L – Desulfobacterium
autotrophicum;
M – Desulfobacterium niacini;
N, O – Desulfobacterium
vacuolatum.
70. Сульфатредуктори по різному ставляться до ацетату
За вуглецевим обміном сульфатредукториподіляються на дві групи:
1. ті, що не здатні окислювати ацетил-КоА
(вони окиснюють органічні сполуки лише до
ацетату і не використовують ацетат як
джерело електронів та вуглецю – «неповні
окиснювачі»);
2. ті, що окиснюють ацетил-КоА,
тобто розкладають органічні сполуки
повністю, до СО2 (такі бактерії здатні рости на
ацетаті).
71. Розподіл сульфатредукторів за вуглецевим метаболізмом
Сульфатредуктори, щоокиснюють органічні сполуки
лише до ацетату
Desulfovibrio
Desulfomicrobium
Desulfomonile
Desulfobotulus
Desulfomonas
Desulfotomaculum (деякі)
Desulfobulbus
Thermodesulfobacterium
Archaeglobus
Сульфатредуктори, що
окиснюють органічні сполуки до
СО2
Desulfobacter
Desulfobacterium
Desulfococcus
Desulfonema
Desulfurella
Desulfosarcina
Desulfuromonas
Desulfoarculus
Desulfotomaculum (деякі)
72. Механізм окисного фосфорилювання у сульфатредукторів
Водень дифундує через ПМ упериплазму, де окислюється
комплексом гідрогенази та
цитохрома с3 згідно реакції :
4Н2 → 8Н+ + 8еНазад у клітину повертається
8е-, які відновлюють SO42доS2-.
Вісім протонів 8Н+
залишаються у периплазмі і
створюють градієнт
концентрації протонів.
АТФ синтезується на АТФсинтетазі згідно
хеміосматичної гіпотези
Мітчела.
Такий механізм утворення
градієнту протонів, зветься
векторним транспортом
електронів.
73. Залізне та марганцеве дихання
Залізне дихання – цевикористання Fe3+ як
термінальний акцептор
електронів за анаеробних
умов.
В результаті нерозчинний
Fe3+ відновлюється до
розчинного Fe2+.
Марганове дихання – це
використання Mn4+як
термінального акцептора
електронів за анаеробних
умов.
В результаті нерозчиний
Бактерії, що здатні відновлювати
Mn4+ відновлюється до
залізо та марганець
розчинного Mn2+.
(1,2 – Shewanella sp.;
Донорами електронів є
3 – Geobacter sp.;
органічні кислоти ацетат, лактат, піруват, 4 – клітини Geobacter sp. на Fe(III)).
форміат:
74. Мультигемові цитохроми переносять е- через периплазму
Мультигемові цитохроми переносять ечерез периплазмуОсновна проблема у використанні Fe(III) для дихання
полягає у нерозчинності цього катіона, а отже у
неможливості його надходження всередину клітини.
Цю проблему грамнегативні бактерії, вирішують так:
Їх клітини містять велику кількість тетра- та декагемових
цитохромів типу с, які утворюють мультигемовий
«провідник» між внутрішньою і зовнішньою
мембранами.
По цьому «провіднику» електрони від реакцій
вуглецевого метаболізму, що відбуваються у
цитоплазмі, передаються через дві мембрани та
периплазму назовні клітини, де, власне, і
відбувається відновлення нерозчинних сполук заліза
(ІІІ).
75. Мультигемові цитохроми
76. Вуглецеве дихання
Вуглецеве диханняпритаманне
гомоацетогенним
бактеріям.
Гомоацетогени – це
облігатно анаеробні
бактерії, які
використовують СО2, як
термінальний акцептор
електронів, і утворюють
єдиний кінцевий продукт
– ацетат.
Джерелами вуглецю можуть
бути органічні речовини
та СО2.
(1,5,6,7 - Acetobacterium woodii;
2 - Acetobacterium wieringae;
3 - Acetobacterium carbinolicum;
4 - Acetobacterium malicum;
8 - Acetogenium kivui)
77. Метаногенез
78.
Метаногени – це групаархеїв, які в процесі
своєї життєдіяльності
утворюють метан (СН4).
Метаногенез – це єдиний
шлях за допомогою
якого метаногени
можуть отримувати
енергію для росту, і це
єдині відомі організми,
що здатні утворювати
1,5 – Methanococcus jannaschii;
2 – Methanopyrus kandleri;
метан, як продукт
3 – Methanosarcina acetivorans;
катаболізму.
4 – Methanoculleus submarines.
79. Особливості будови клітин метаногенів
Метаногени мають особливості:• У будові плазматичної мембрани;
• У будові клітинної стінки;
• Містять унікальні коферменти, що беруть участь у
перенесенні водню та С1-сполук при утворенні
метану , а саме:
кофермент F420,
метанофуран,
тетрагідрометаноптерін,
кофермент М,
фактор В,
кофермент F430,
метанофеназин.
80. Субстати метаногенів
Метаногени використовують як субстрати, аотже перетворюють на метан, такі сполуки:
1. Субстрати, що містять окиснений вуглець –
вуглекислий газ СО2, форміат НСООН,
монооксид вуглецю СО;
2. Субстрати, що містять метильну групу –
метанол СН3ОН, метиламін , диметиламін,
триметиламін, метилмеркаптан,
диметилсульфід;
3. Оцтову кислоту СН3СООН.
81. Утворення метану на різних субстратах
СО2 + 4Н2 → СН4 + 2Н2О∆G0’= -134 кДж /реакція
4НСООН → СН4 + 3СО2+2Н2О
∆G0’= -281 кДж / реакція
4СО + 2Н2О → СН4 + 3СО2
∆G0’= -209 кДж /реакція
4СН3ОН → 3СН4 + СО2+2Н2О
∆G0’= -323 кДж /реакція
СН3ОН + Н2 → СН4 + Н2О
∆G0’= -133 кДж /реакція
СН3СООН → СН4 + СО2
∆G0’= -130 кДж /реакція
82. Метаногенез – унікальний древній процес
Метаногенез часом розглядають як процес анаеробногодихання і називають його карбонатним диханням,
оскільки при рості метаногенів на суміші СО2 та Н2
молекулярний водень є донором електронів, а
вуглекислий газ – акцептором.
В літературі часто зустрічається термін «метанове
бродіння». При рості метаногенів на метанолі одна
молекула субстрату окиснюється, а друга –
відновлюється, що нагадує процес зародження
амінокислот.
Метаногенез не можна розглядати ні як анаеробне
дихання, ні як бродіння. Це унікальний специфічний
процес отримання енергії, характерний лише для
древніх форм живих організмів – археїв.
83. Анаеробні мікробні угрупування
• Мікробні угруповання (мікробні асоціації) – цесукупність популяцій різних видів
мікроорганізмів, які мешкають разом у просторі і
часі.
Вони об’єднані трофічними зв’язками, і одні види,
в результаті своєї життєдіяльності, створюють
сприятливі умови для існування інших.
• Метаногенним мікробним угрупованням
називають трофічну систему анаеробних
бактерій, останнім продуктом якої є метан.
У розщепленні субстрату метаногенним
угрупованням задіяно кілька фізіологічних груп
мікроорганізмів
84. Деструкція целюлози метаногенним мікробним угрупованням
1 - первинні бактеріїбродильники;2 – метаногени, що
окиснюють водень;
3 – метаногени, що
розщеплюють ацетат;
4 - вторинні бактеріїбродильники
(синтрофні бактерії);
5 – гомоацетогенні
бактерії
85. Розповсюдження метаногенних угрупувань
Рисові поляТермітники
Болота
Жуйні тварини