

































 Биология
БиологияПохожие презентации:









")
Биофизика мышечного сокращения
1.
Биофизика мышечного сокращенияВнешние механические проявления движения мышц –
телодвижения, кровообращение, дыхание
и прочие макродвижения
Сокращение мышц (изменение размера) обусловлено
наличием в них
сократительного комплекса – актомиозина
2.
Проблемы:Микропроявления? – активный транспорт
веществ через клеточные мембраны
Преобразование химической энергии в механическую
Инженерная схема: ХЭ → ТЭ → ПромежМЭ → ЭЭ → МЭ
Схема организма: ХЭ → МЭ
3.
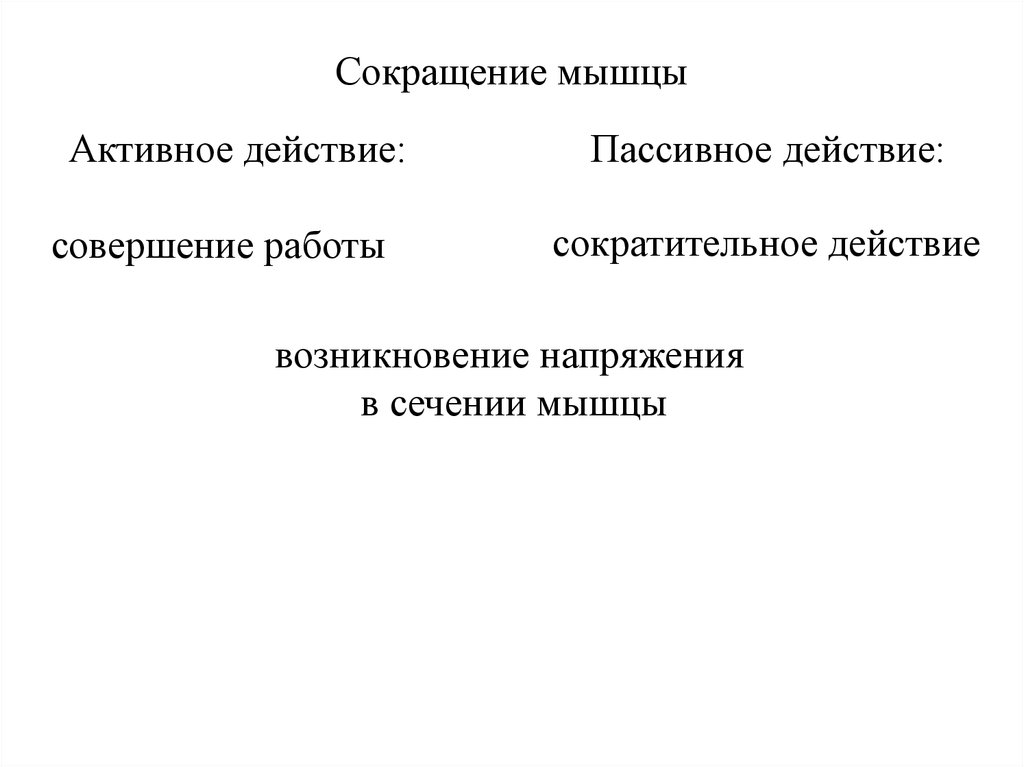
Сокращение мышцыАктивное действие:
совершение работы
Пассивное действие:
сократительное действие
возникновение напряжения
в сечении мышцы
4.
Крайние случаи сокращения мышц:Изометрические
неизменная длина мышц
Изотонические
неизменное напряжение
мышц
5.
Абсолютная мышечная сила:Отношение максимального веса груза, который может
поднять мышца к ее физиологическому сечению
Скелетная мышца – 106 Па, гладкая – 105 Па
6.
Зависимость между силой, развиваемой мышцейпри сокращении, от скорости сокращения (укорочения)
мышцы:
(основное уравнение сокращения мышцы)
7.
Максимальная скорость сокращениямышцы (МССМ)
Примерный вид зависимости
МССМ от величины нагрузки,
которую преодолевает мышца
(эксперимент)
Тип кривой - гипербола
Сила, преодолеваемая мышцей
8.
Основное уравнение сокращения мышцы Хилла:F a v b const
Интерпретация:
Байер:
F v const
P0 Fv – общая мощность мышцы при сокращении
P Fv – реальная мощность мышцы
P
P0
9.
Интерпретация:энергия на сокращение:
внешняя работа
внутренняя работа
(тепловое рассеяние)
10.
Энергетический баланс:Сокращение мышцы = механическое (внешнее)
сокращение + теплообразование
Преобразование энергии в организме =
= механическая + тепловая
Организм = теплоэнергоцентраль
11.
Разумная энергетическая теория:Хилл:
при каждом раздражении выделяется
постоянная теплота QA активации
(не зависит от нагрузки?):
QA const
Теплота сокращения QC:
QC k l
Зависит от сокращения мышц Δl и не зависит
от нагрузки
12.
К.п.д. мышечного сокращения:F l
полезное
η
затраченное
QA QC k l F l
F l
η
QA k l F l
13.
Данные Хилла:η Мех.Энергия 0,4
η( Пит.Веществ ) 0,5
Суммарный к.п.д. превращения энергии ПП→М :
η η( ПB ) η( МЭ) 0,2
14.
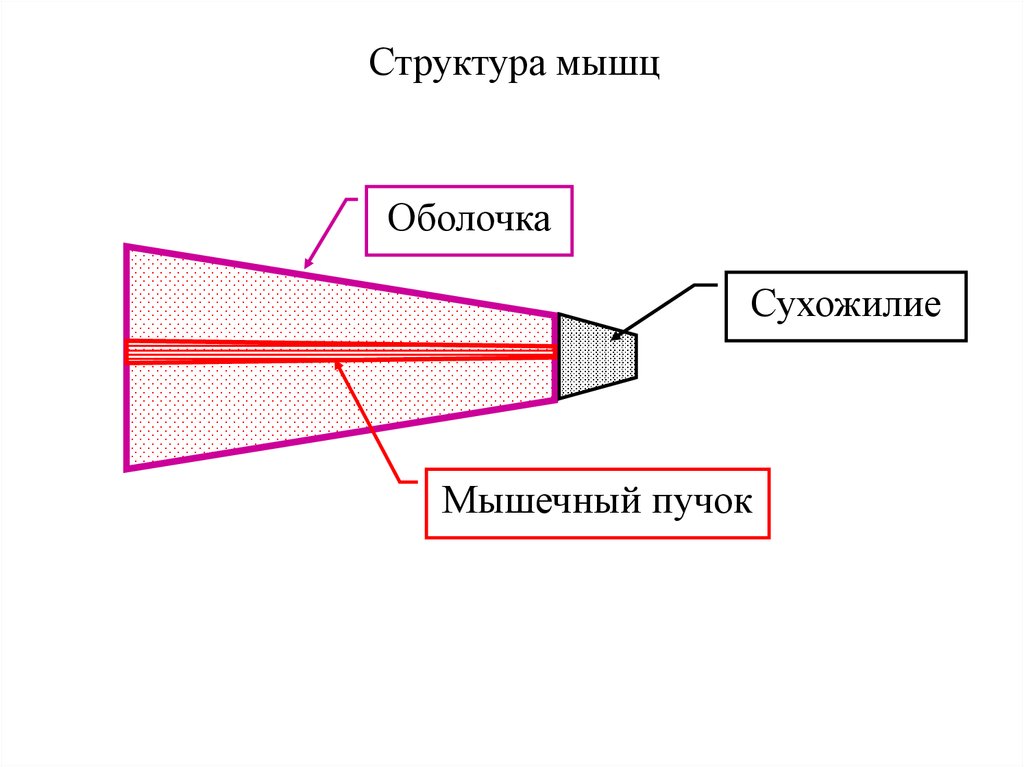
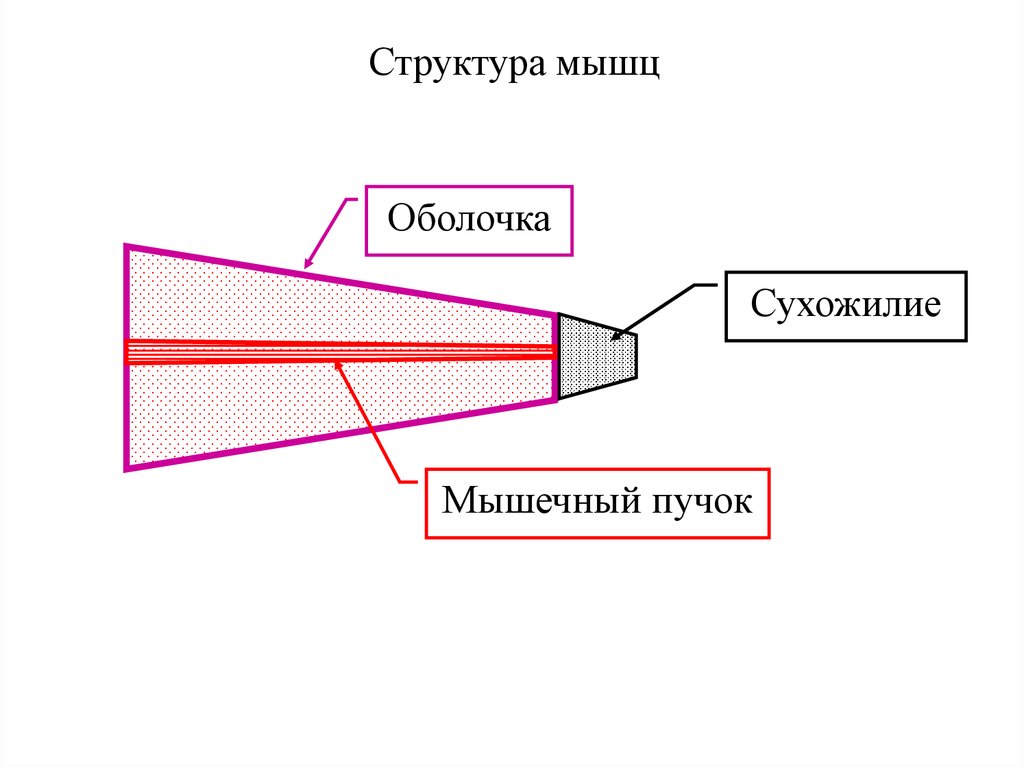
Структура мышцОболочка
Сухожилие
Мышечный пучок
15.
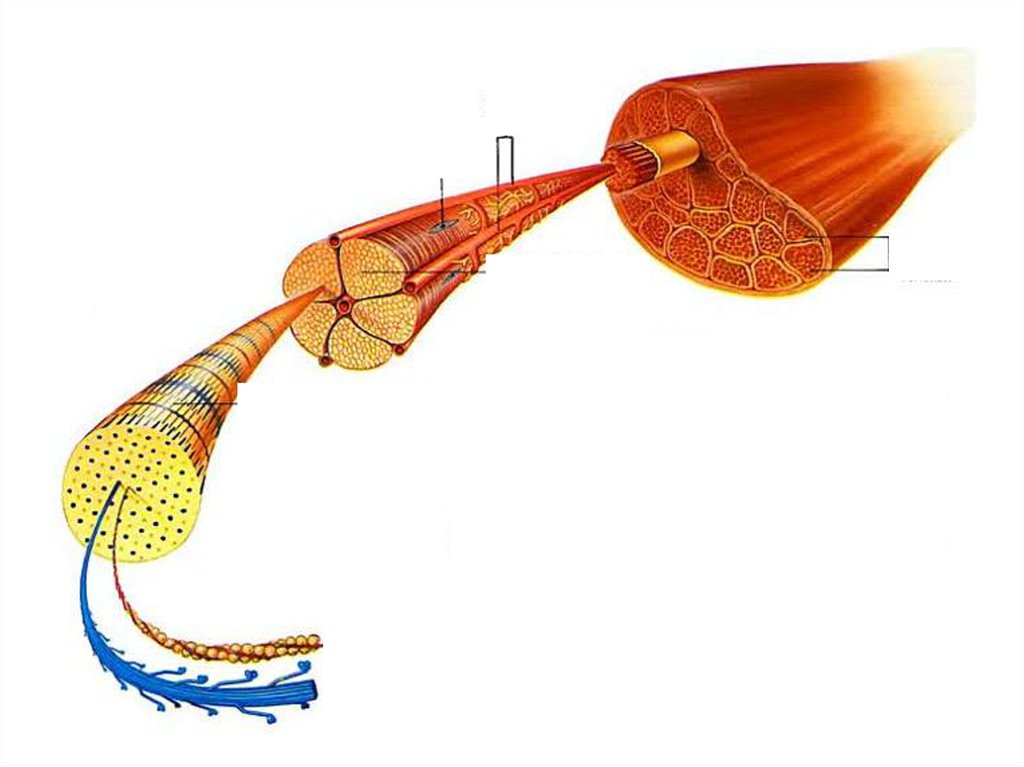
Мышечный пучокНесколько тысяч параллельных мышечных
волокон d = 10 – 100 мкм
16.
Отдельное мышечное волокноНерв
17.
Δφ ≈ 100 мВМембрана волокна –
плазмалемма h ≈ 10 нм
Ядро
МХ
Саркоплазма
Слой коллагеновых нитей
1000 – 2000 || миофибрилл d ≈ 1 мкм
18.
19.
Оптическая микроскопия:поперечная полосатость волокон
Саркоплазма
Обусловлена поперечной полосатостью миофибрилл,
образующих внутри волокна упорядоченную структуру
20.
Электронная микроскопия и РСА:отдельная миофибрилла
Тонкие актиновые протофибриллы
lА ≈ 1 мкм, d ≈ 5 – 7 нм
Толстые миозиновые протофибриллы
lM ≈ 1,5 мкм, d ≈ 16 нм
21.
I – диск (И)I – диск
А – диск (А)
Z – мембрана
Саркомер
(участок мяса)
22.
Каждая миозиновая нить соединена мостикамис 6-тью соседними актиновыми нитями:
≈ 40 нм
≈ 6,7 нм
23.
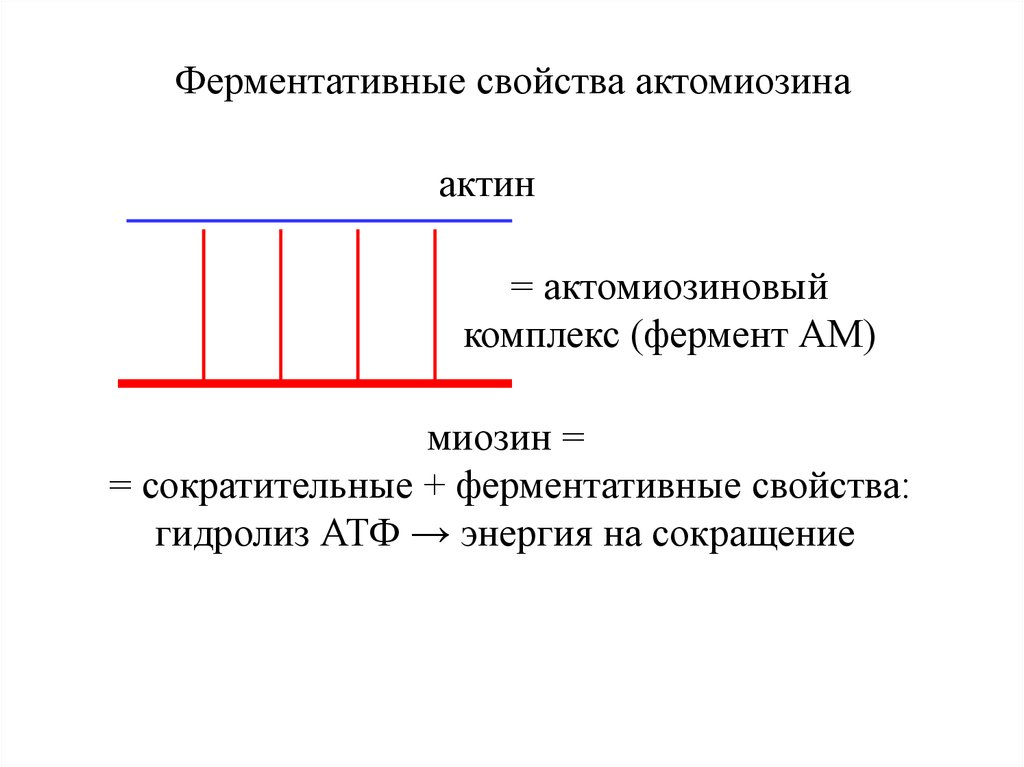
Ферментативные свойства актомиозинаактин
= актомиозиновый
комплекс (фермент АМ)
миозин =
= сократительные + ферментативные свойства:
гидролиз АТФ → энергия на сокращение
24.
Ферментативная активность (ФА)определяет способность к сокращению миофибрилл:
AAM
10
AM
AAM f (CCa 2 )
25.
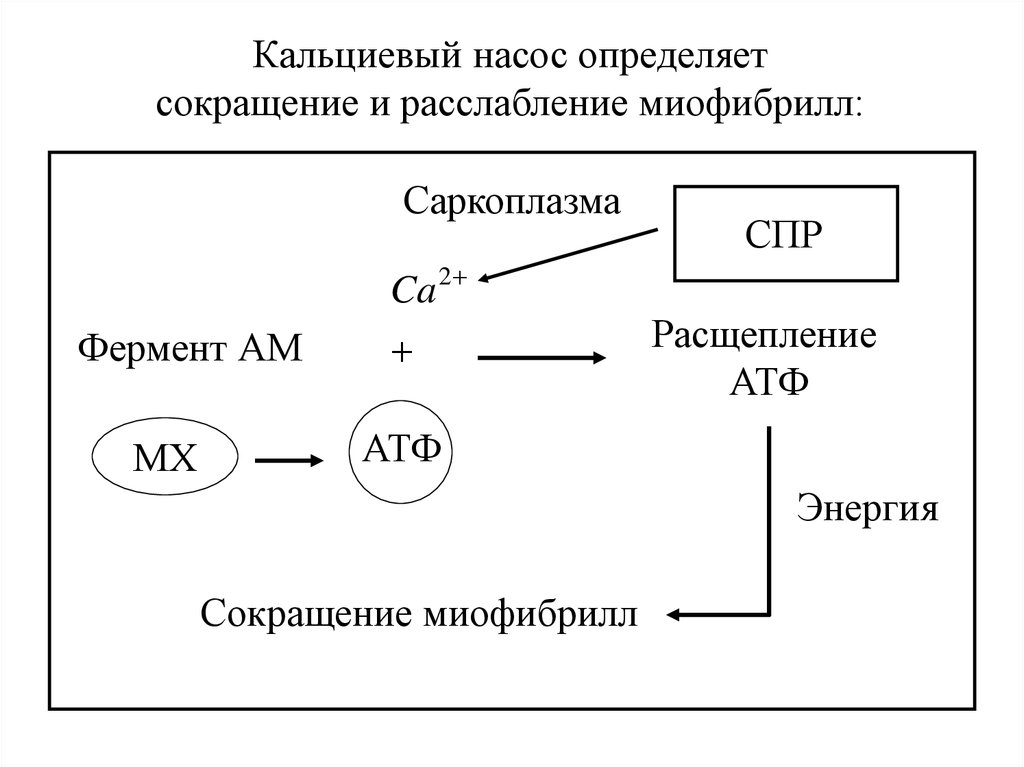
Кальциевый насос определяетсокращение и расслабление миофибрилл:
Саркоплазма
Фермент АМ
МХ
Ca 2
СПР
Расщепление
АТФ
АТФ
Энергия
Сокращение миофибрилл
26.
Локализация насоса – саркоплазматическийретикулум (СПР)
Деполяризация мембран СПР
Освобождение Са 2+ (СПР → саркоплазма)
↑ ФААМ
Расщепление АТФ
Сокращение миофибрилл
27.
Из саркоплазмы Са 2+СПР
Расслабление миофибрилл
28.
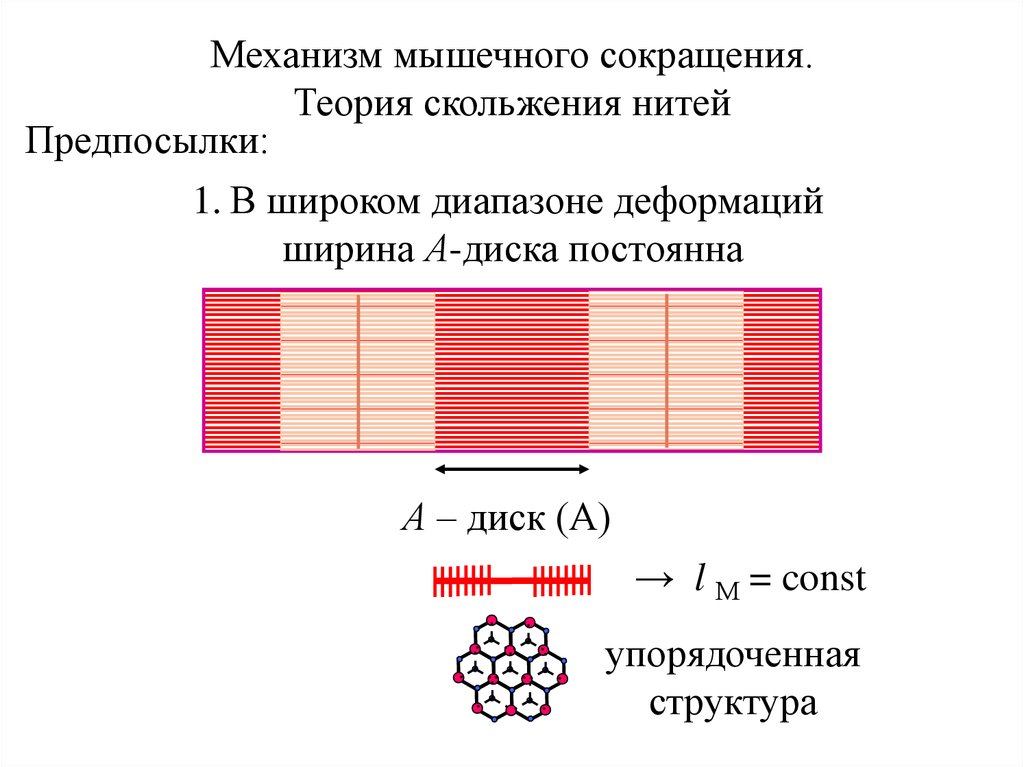
Механизм мышечного сокращения.Теория скольжения нитей
Предпосылки:
1. В широком диапазоне деформаций
ширина А-диска постоянна
А – диск (А)
→ l М = const
упорядоченная
структура
29.
2. При изменении длины саркомера изменяетсяширина I-диска (актиновые нити)
I – диск
I – диск
lC
lC1
30.
3. В большой области деформаций длинымиозиновых и актиновых нитей постоянны
4. При сокращении мышцы нити скользят друг
относительно друга без изменения длины
31.
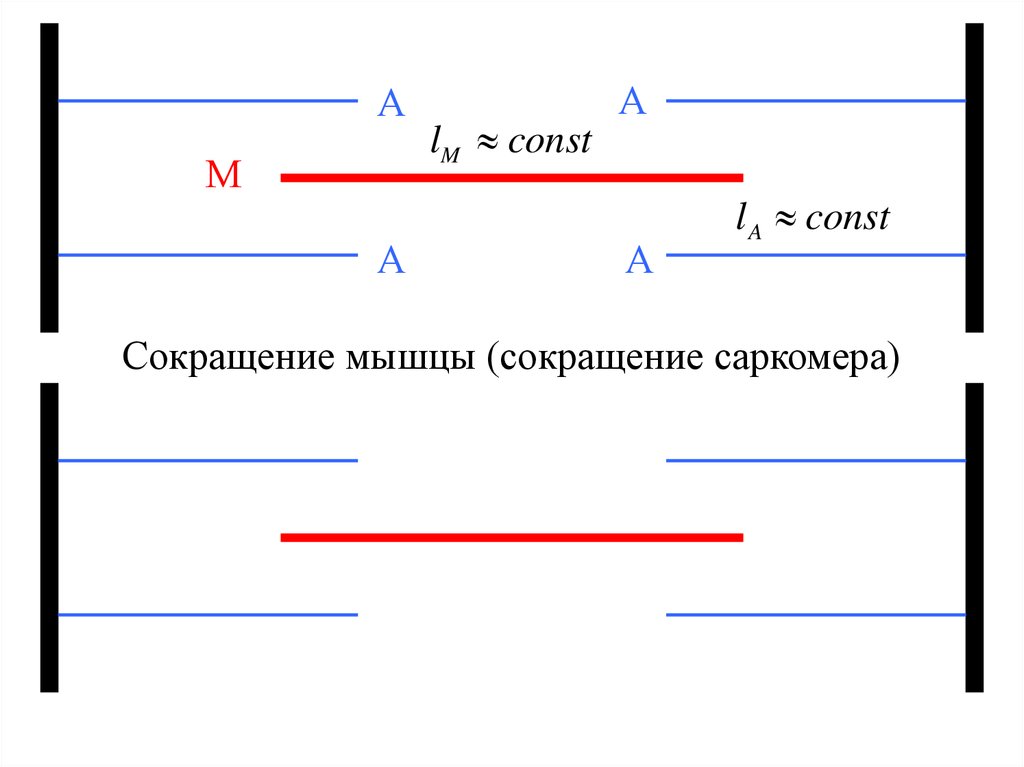
АМ
А
lM const
А
А
l A const
Сокращение мышцы (сокращение саркомера)
32.
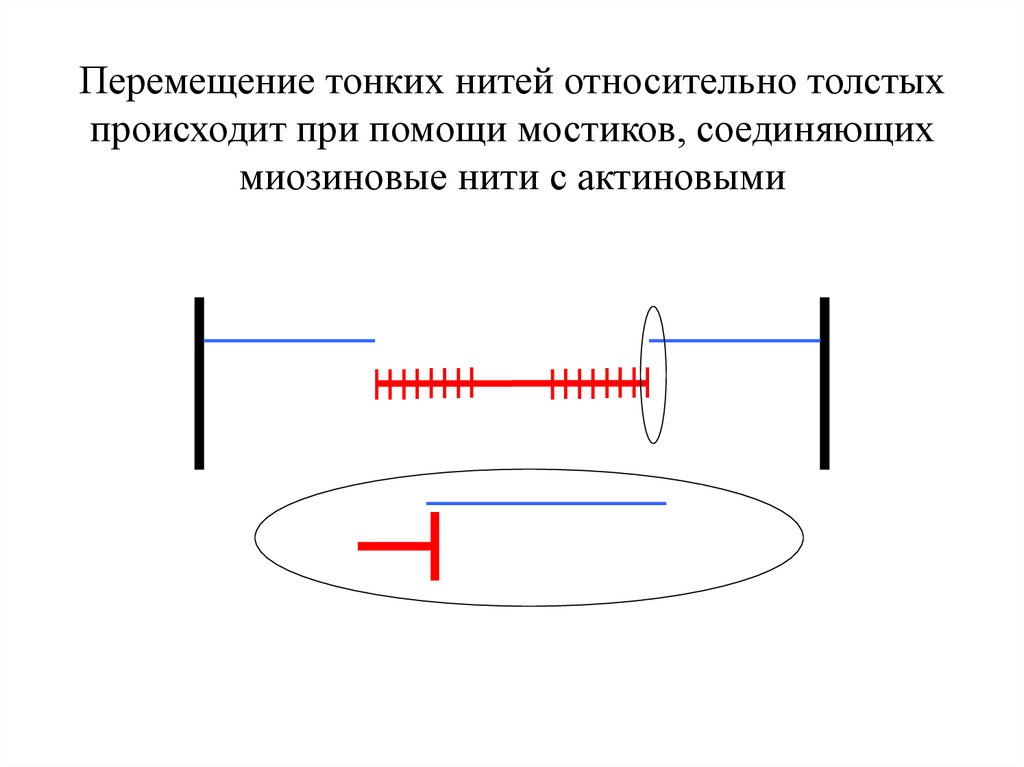
Перемещение тонких нитей относительно толстыхпроисходит при помощи мостиков, соединяющих
миозиновые нити с актиновыми
33.
Ca 2Гидролиз АТФ = АДФ + фосфатный остаток →
→ механическая энергия
34.
Перекрывание миозиновых и актиновых нитейпри различных длинах саркомера
N – количество мостиков в перекрытии нитей
N 0
≥ 3,65 мкм (1)
35.
N2,25 – 3,65 мкм
36.
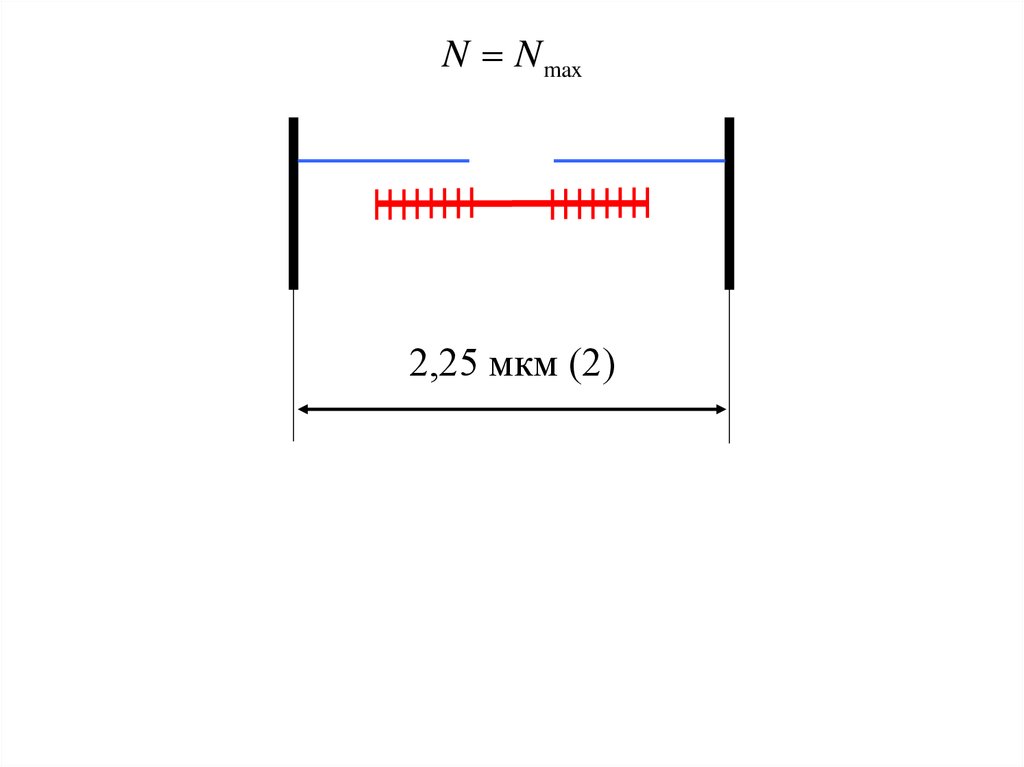
N N max2,25 мкм (2)
37.
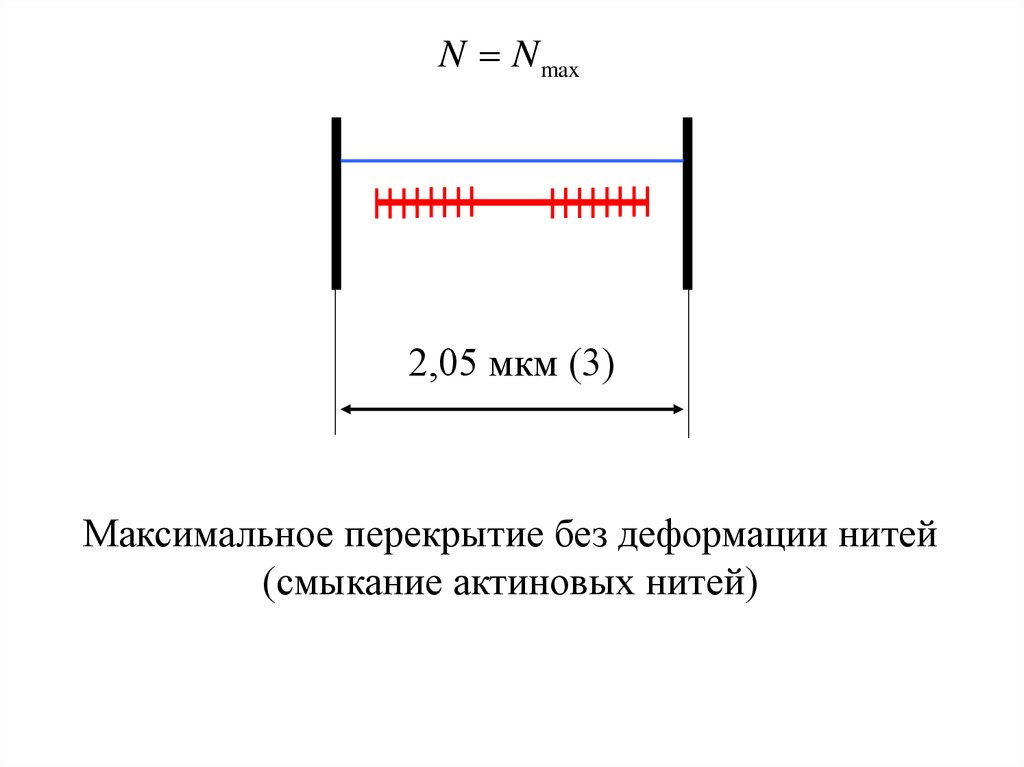
N N max2,05 мкм (3)
Максимальное перекрытие без деформации нитей
(смыкание актиновых нитей)
38.
Деформация актиновых нитей после их смыканияN
Начало
деформации
мостиков
1,65 мкм (4)
39.
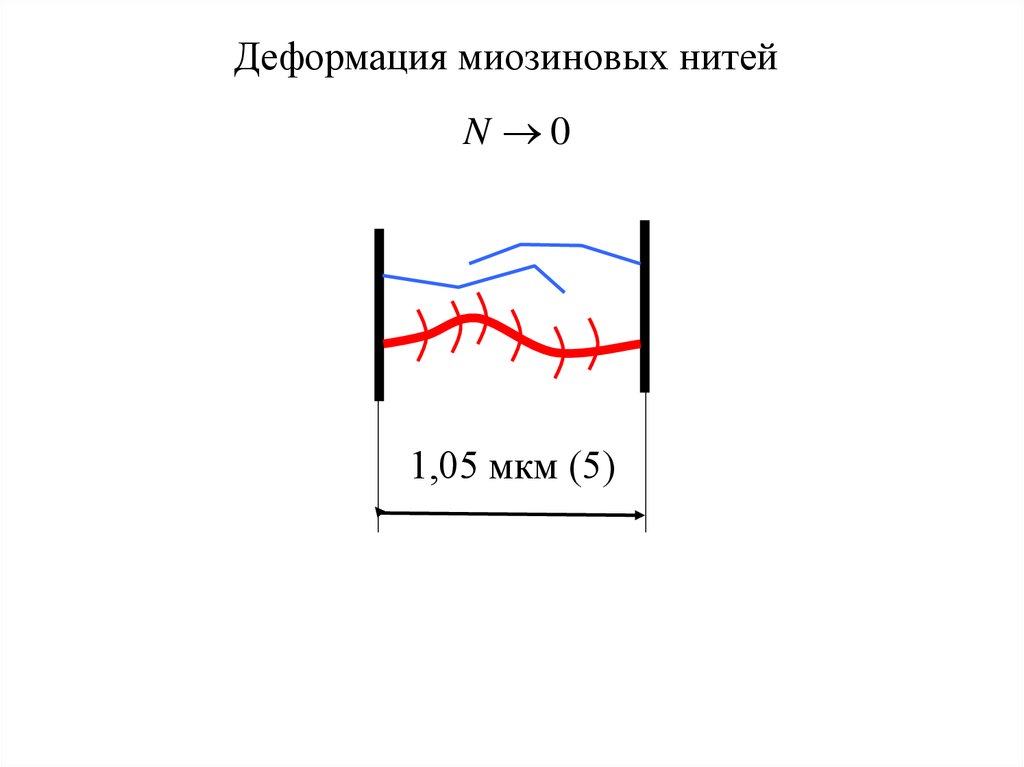
Деформация миозиновых нитейN 0
1,05 мкм (5)
40.
Разумное предположение: количество актино –миозиновых мостиков пропорционально
развиваемому мышечному усилию
FCOKP. f ( N )
41.
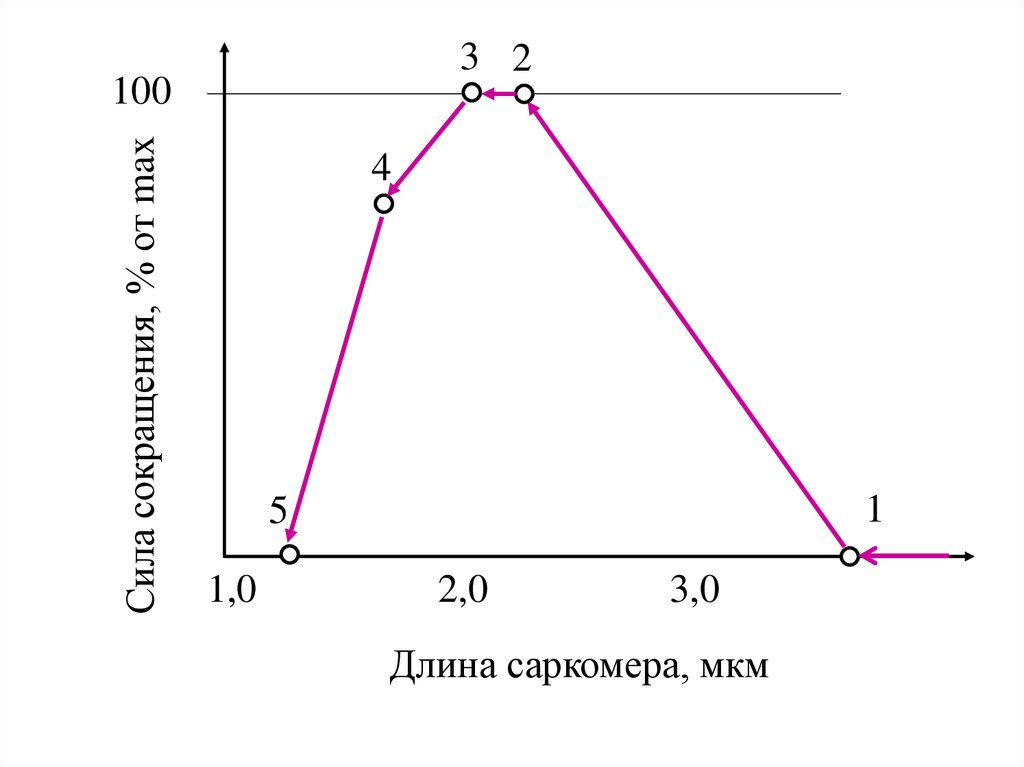
3 2Сила сокращения, % от max
100
4
1
5
1,0
2,0
3,0
Длина саркомера, мкм
42.
Самостоятельно на графике указать:1. Длины саркомера, соответствующие точкам 1,2,3,4,5
2. Тенденции изменения N на участках 1 – 2,
2 – 3, 3 – 4, 4 – 5
43.
Гипотеза Дэвиса о механизме взаимодействиямиозиновых и актиновых нитей
1. Состояние покоя:
АДФ
-
АТФ
-
АДФ
А
Электростатическое
отталкивание ↔ упругость
-
АТФ – аза
М
44.
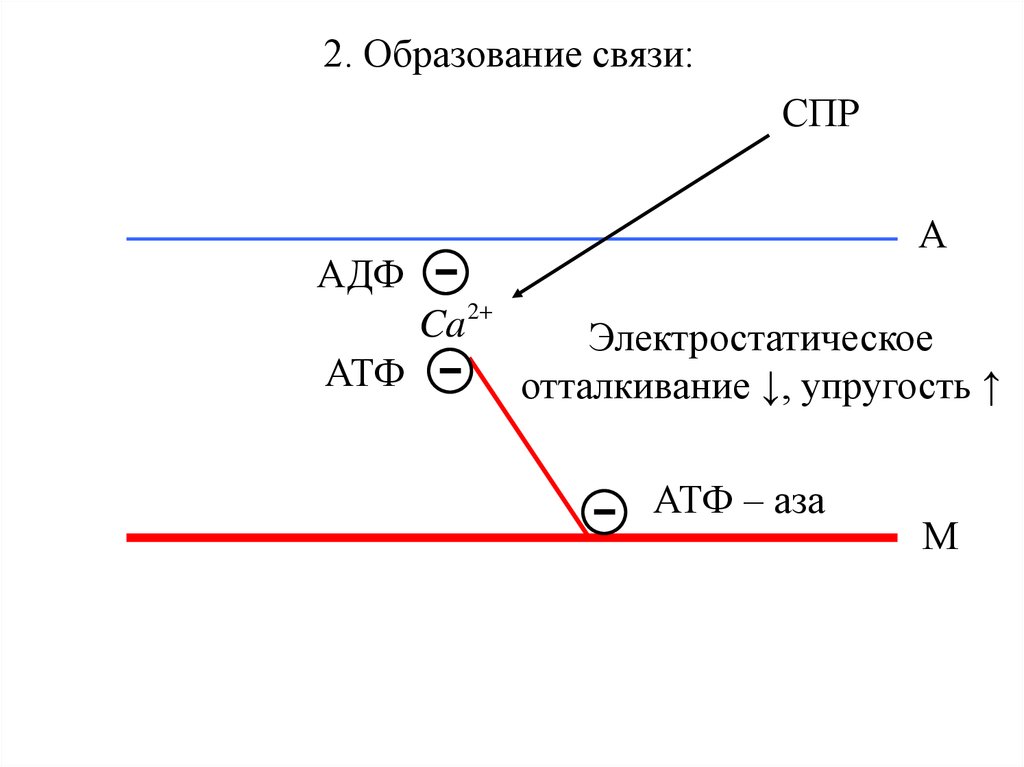
2. Образование связи:СПР
Ca
АТФ -
АДФ
А
2
Электростатическое
отталкивание ↓, упругость ↑
-
АТФ – аза
М
45.
3. Сокращение мостика:3.1. Высвобождение потенциальной энергии мостика
3.2. Перемещение актиновой нити на один шаг
3.3. Доставка АТФ к АТФ – азе.
А
Ca 2
АТФ
-
АТФ – аза
М
46.
4. Гидролиз АТФРазрыв связи между А и М
А
Д
Ca 2
--
АТФ – аза
М
47.
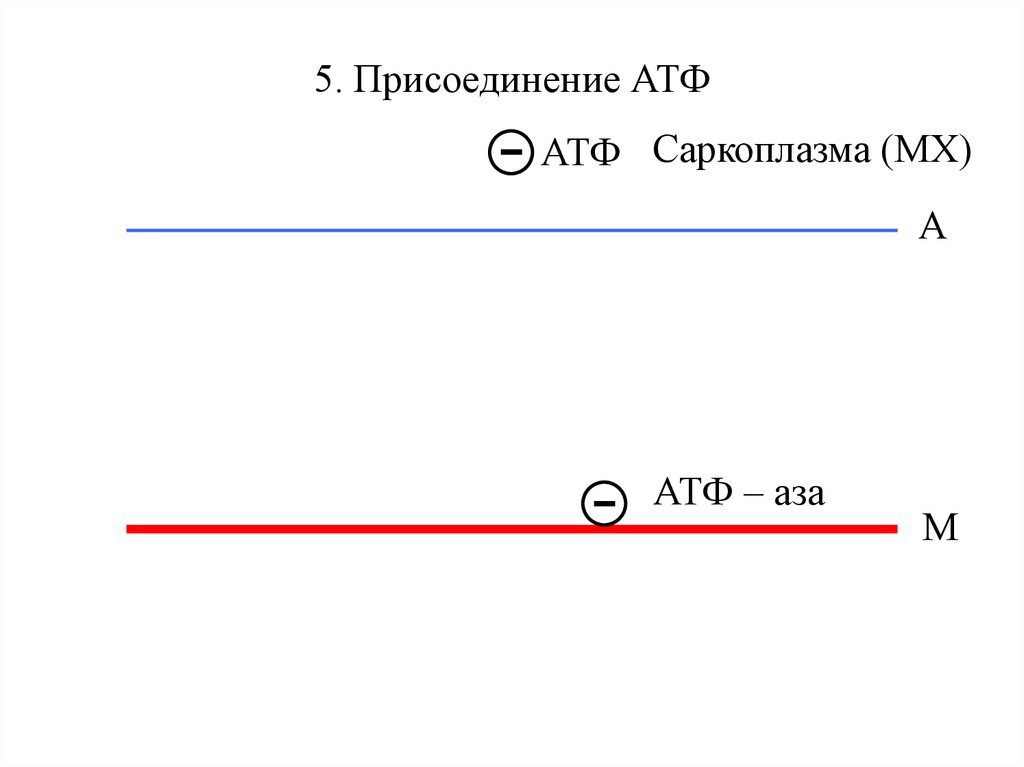
5. Присоединение АТФ- АТФ
Саркоплазма (МХ)
А
-
АТФ – аза
М
48.
6. Вытягивание мостикаА
Электростатическое
отталкивание ↑
- АТФАТФ – аза
-
М
49.
Выводы:1. Мышцы обладают как пассивными, так и активными
механическими свойствами
2. Активные механические свойства обусловлены
наличием в мышцах актино – миозинового комплекса
3. В мышцах осуществляется уникальное
преобразование химической энергии в механическую
4. Механизм мышечного сокращения объясняется
теорией скольжения нитей
50.
Структура мышцОболочка
Сухожилие
Мышечный пучок