






























































































 Биология
БиологияПохожие презентации:










Анатомия и физиология ЦНС
1.
Анатомия и физиологияЦНС
2.
Предмет и задачи анатомии ЦНСОтделы нервной системы
Анатомия человека — это наука о происхождении и развитии, формах и
строении человеческого организма. Анатомия изучает внешние формы
и пропорции тела человека и его частей, отдельные органы, их
конструкцию, микроскопическое строение.
Задачи анатомии:
• исследование основных этапов развития человека в процессе
эволюции;
• изучение особенностей строения тела и отдельных органов в
различные возрастные периоды;
• исследование формирования человеческого организма в условиях
внешней среды.
3.
Предмет и задачи анатомии ЦНСОтделы нервной системы (продолжение)
Всю НС можно разделить по топографическому (в зависимости от места
расположения той или иной ее части) и функциональному (по
выполняемым функциям) принципам.
По топографическому принципу НС делят на центральную и
периферическую.
Центральная нервная система (ЦНС) включает головной и спинной
мозг, защищенные мозговыми оболочками.
Периферическая нервная система — это нервы, нервные узлы
(ганглии), нервные сплетения и нервные окончания — включает 12 пар
черепных нервов, 31 пару спинномозговых нервов, сенсорные
(чувствительные) и вегетативные ганглии, нервные сплетения.
4.
Предмет и задачи анатомии ЦНСОтделы нервной системы (продолжение)
По функциональному признаку НС подразделяется на
соматическую и вегетативную нервную систему. У каждой из них
есть центральная (т. е. находящаяся в ЦНС) и периферическая
(находящаяся за пределами ЦНС) части.
5.
Предмет и задачи анатомии ЦНСОтделы нервной системы (продолжение)
Соматическая НС — отдел НС, который регулирует работу
скелетных мышц, запуская поведенческие реакции и осуществляя
связь организма с внешней средой. Человек может произвольно,
по собственному желанию управлять деятельностью скелетной
мускулатуры.
6.
Предмет и задачи анатомии ЦНСОтделы нервной системы (продолжение)
Вегетативная (автономная) НС (ВНС) — отдел НС, регулирующий
работу внутренних органов. ВНС управляет деятельностью гладкой
и сердечной мускулатуры и желез, регулируя (усиливая или
ослабляя) и координируя деятельность внутренних органов.
Человек без специальной тренировки не может сознательно
управлять деятельностью этой системы, т. е. она непроизвольная. В
ВНС выделяют симпатический, парасимпатический и
метасимпатический отделы.
7.
Основные функции нервной системы- регулирует работу организма;
- обеспечивает связь с окружающей средой;
- координирует работу всех систем органов;
- обеспечивает протекание психических процессов.
8.
Нервная тканьНервная ткань — это основная ткань, из которой построена
нервная система. Она состоит из нервных клеток — нейронов,
которые выполняют основные, специфические функции, и
глиальных клеток — нейроглии, выполняющие вспомогательные
функции.
9.
Нервная ткань (продолжение)Источники развития — нервная трубка, нервный гребень (ганглиозные
пластинки) и плакоды. Нервная трубка образуется в результате
смыкания краёв нервного желобка, развивающегося из эктодермы.
Нервный гребень расположен между нервной трубкой и эктодермой. Он
образуется в результате выселения клеток из утолщенных краев
нервного желобка — нервных валиков. Плакоды представляют собой
утолщения эктодермы по бокам нервной трубки на головном конце
зародыша. Нейробласты нервной трубки дают начало нервным клеткам,
а глиобласты — глиальным клеткам головного и спинного мозга. Из
клеток нервного гребня происходят нейроны и нейроглия всех нервных
ганглиев, а из плакод — рецепторные (нейросенсорные) клетки органа
обоняния, нейроны слухового и вестибулярного ганглиев. Клетки
микроглии образуются из промоноцитов красного костного мозга.
10.
Нервная ткань (продолжение)В ходе эмбриогенеза до 85 % образующихся нейронов гибнет в
результате апоптоза (генетически запрограммированной смерти).
Погибают лишние, дефектные нейроны, которые не нашли свои
«клетки-мишени» или оказались избыточными, «лишними».
В развитии нейронов выделяют две стадии: домедиаторную и
медиаторную. Во второй стадии в нейронах происходит синтез
нейромедиатора, его накопление и образование синапсов.
11.
Нервные клетки (нейроциты, нейроны)Нейроциты (нейроны) способны воспринимать, анализировать
раздражение, приходить в состояние возбуждения, генерировать
нервные импульсы, передавать их другим нейронам, либо рабочим
органам. Число нейронов в нервной ткани человека достигает одного
триллиона.
Как и другие клетки, нейроны состоят из цитоплазмы и ядра. В нейроне
выделяют перикарион или тело клетки (часть цитоплазмы вокруг ядра),
отростки и нервные окончания (концевые ветвления). Размеры
перикарионов варьируют от 4 мкм у клеток-зёрен мозжечка до 130 мкм
у ганглиозных нейронов коры головного мозга. Длина отростков может
достигать 1 м (например, отростки нейронов спинного мозга и
спинномозговых узлов достигают кончиков пальцев рук и ног).
12.
Общие принципы строения нейрона1. Тело нейрона.
2. Аксон.
3. Дендриты.
4. Перехват Ранвье.
5. Нервное окончание. (По Stevens, 1979).
13.
Нервные клетки (нейроциты, нейроны)Отростки нейронов делятся на два вида: аксоны (нейриты) и
дендриты. Аксон в нервной клетке всегда один, он отводит
нервный импульс от тела нейрона и передаёт его на другие
нейроны или клетки рабочих органов (мышцы, железы).
Дендритов (от греч. «dendron» — дерево) в нервной клетке один
или несколько, они приносят импульсы к телу нейрона. Дендриты в
тысячи раз увеличивают рецепторную, воспринимающую
поверхность нейрона.
14.
Нервные клетки (нейроциты, нейроны)(продолжение)
Нейрон является самостоятельной структурно-функциональной
единицей, но с помощью своих отростков взаимодействует с
другими нейронами, образуя рефлекторные дуги — нейронные
цепи, из которых построена нервная система.
В организме человека нервный импульс передаётся от одного
нейрона к другому, либо на рабочий орган не напрямую, а через
химический посредник — медиатор.
В нервной системе животных и человека обнаружено около сотни
разных медиаторов, а соответственно и нейронов различной
медиаторной природы.
15.
Классификации нейроновОна осуществляется по трём основным группам признаков:
морфологическим, функциональным и биохимическим.
1. Морфологическая классификация нейронов (по особенностям
строения). По количеству отростков нейроны делятся на:
• униполярные (с одним отростком);
• биполярные (с двумя отростками);
• псевдоуниполярные (ложно униполярные);
• мультиполярные (имеют три и более отростков).
Последних в нервной системе больше всего.
16.
Типы нервных клеток1. Униполярный нейрон.
2. Псевдоуниполярный нейрон.
3. Биполярный нейрон.
4. Мультиполярный нейрон.
В цитоплазме нейронов видны нейрофибриллы.
(По Ю. А. Афанасьеву и др.).
17.
Классификации нейронов (продолжение)Псевдоуниполярными нейроны называют потому, что отходя от
тела, аксон и дендрит вначале плотно прилегают друг к другу,
создавая впечатление одного отростка, и лишь потом Т-образно
расходятся (к ним относятся все рецепторные нейроны спинальных
и краниальных ганглиев).
Униполярные нейроны встречаются только в эмбриогенезе.
Биполярными нейронами являются биполярные клетки сетчатки
глаза, спирального и вестибулярного ганглиев. По форме описано
до 80 вариантов нейронов: звёздчатые, пирамидальные,
грушевидные, веретеновидные, паукообразные и др.
18.
Классификации нейронов (продолжение)2. Функциональная (в зависимости от выполняемой функции и
места в рефлекторной дуге): рецепторные, эффекторные,
вставочные и секреторные.
19.
Классификации нейронов (продолжение)Рецепторные (чувствительные, афферентные) нейроны с помощью
дендритов воспринимают воздействия внешней или внутренней
среды, генерируют нервный импульс и передают его другим типам
нейронов. Они встречаются только в спинальных ганглиях и
чувствительных ядрах черепномозговых нервов.
Эффекторные (эфферентные) нейроны, передают возбуждение на
рабочие органы (мышцы или железы). Они располагаются в
передних рогах спинного мозга и вегетативных нервных ганглиях.
20.
Классификации нейронов (продолжение)Вставочные (ассоциативные) нейроны располагаются между
рецепторными и эффекторными нейронами; по количеству их
больше всего, особенно в ЦНС.
Секреторные нейроны (нейросекреторные клетки) — это
специализированные нейроны, по своей функции напоминающие
эндокринные клетки. Они синтезируют и выделяют в кровь
нейрогормоны, расположены в гипоталамической области
головного мозга. Они регулируют деятельность гипофиза, а через
него и многие периферические эндокринные железы.
21.
Классификации нейронов (продолжение)3. Медиаторная (по химической природе выделяемого медиатора):
- холинергические нейроны (медиатор ацетилхолин);
- аминергические (медиаторы — биогенные амины, например,
норадреналин, серотонин, гистамин);
- ГАМКергические (медиатор — гаммааминомасляная кислота);
- аминокислотергические (медиаторы — аминокислоты, такие как
глютамин, глицин, аспартат);
- пептидергические (медиаторы — пептиды, например, опиоидные
пептиды, субстанция Р, холецистокинин, и др.);
- пуринергические (медиаторы — пуриновые нуклеотиды, например,
аденин и др.).
22.
Пресинаптические нейроны, образующие синапсы исинтезирующие и выделяющие их медиаторы, называются
соответственно холинергическими, аминергическими, ГАМКергическими и др. Постсинаптические нейроны с рецепторами к
этим медиаторам называются соответственно холино-, амино- и
ГАМКреактивными.
23.
Внутреннее строение нейроновЯдро нейрона обычно крупное, округлое с мелкодисперсным
хроматином, 1-3 крупными ядрышками. Это отражает высокую
интенсивность процессов транскрипции в ядре нейрона.
Клеточная оболочка нейрона способна генерировать и проводить
электрические импульсы. Это достигается изменением локальной
проницаемости её ионных каналов для Na+ и К+, изменением
электрического потенциала и быстрым перемещением его по
цитолемме (волна деполяризации, нервный импульс).
24.
Внутреннее строение нейронов(продолжение)
В цитоплазме нейронов хорошо развиты все органоиды общего
назначения. Митохондрии многочисленны и обеспечивают высокие
энергетические потребности нейрона, связанные со значительной
активностью синтетических процессов, проведением нервных
импульсов, работой ионных насосов. Они характеризуются быстрым
изнашиванием и обновлением.
Комплекс Гольджи очень хорошо развит. Неслучайно эта органелла
впервые была описана и демонстрируется в курсе цитологии именно в
нейронах. При световой микроскопии он выявляется в виде колечек,
нитей, зёрнышек, расположенных вокруг ядра (диктиосомы).
Многочисленные лизосомы обеспечивают постоянное интенсивное
разрушение изнашиваемых компонентов цитоплазмы нейрона
(аутофагия).
25.
Ультрастуктурная организация теланейрона
Д — дендриты, А — аксон.
1. Ядро (ядрышко показано стрелкой).
2. Митохондрии.
3. Комплекс Гольджи.
4. Хроматофильная субстанция (участки гранулярной
цитоплазмотической сети).
5. Лизосомы.
6. Аксонный холмик.
7. Нейротрубочки, нейрофиламенты.
(По В. Л. Быкову).
26.
Внутреннее строение нейронов(продолжение)
Для нормального функционирования и обновления структур нейрона в
них должен быть хорошо развит белоксинтезирующий аппарат.
Гранулярная цитоплазматическая сеть в цитоплазме нейронов образует
скопления, которые хорошо окрашиваются основными красителями и
видны при световой микроскопии в виде глыбок хроматофильного
вещества (базофильное или тигровое вещество, субстанция Ниссля).
Термин «субстанция Ниссля» сохранился в честь учёного Франца Ниссля,
впервые ее описавшего. Глыбки хроматофильного вещества
расположены в перикарионах нейронов и дендритах, но никогда не
встречаются в аксонах, где белоксинтезирующий аппарат развит слабо.
При длительном раздражении или повреждении нейрона эти скопления
гранулярной цитоплазматической сети распадаются на отдельные
элементы, что на светооптическом уровне проявляется исчезновением
субстанции Ниссля (хроматолиз, тигролиз).
27.
Внутреннее строение нейронов(продолжение)
Цитоскелет нейронов хорошо развит, образует трёхмерную сеть,
представленную нейрофиламентами (толщиной 6-10 нм) и
нейротрубочками (диаметром 20-30 нм).
Нейрофиламенты и нейротрубочки связаны друг с другом поперечными
мостиками, при фиксации они склеиваются в пучки толщиной 0,5-0,3
мкм, которые окрашиваются солями серебра. На светооптическом
уровне они описаны под названием нейрофибрилл. Они образуют сеть в
перикарионах нейроцитов, а в отростках лежат параллельно.
Цитоскелет поддерживает форму клеток, а также обеспечивает
транспортную функцию — участвует в транспорте веществ из
перикариона в отростки (аксональный транспорт).
28.
Внутреннее строение нейронов(продолжение)
Включения в цитоплазме нейрона представлены липидными
каплями, гранулами липофусцина — «пигмента старения» —
жёлто-бурого цвета липопротеидной природы. Они представляют
собой остаточные тельца (телолизосомы) с продуктами
непереваренных структур нейрона.
По-видимому, липофусцин может накапливаться и в молодом
возрасте при интенсивном функционировании и повреждении
нейронов. Кроме того, в цитоплазме нейронов черной субстанции
и голубого пятна ствола мозга имеются пигментные включения
меланина. Во многих нейронах головного мозга встречаются
включения гликогена.
29.
Внутреннее строение нейронов(продолжение)
Нейроны не способны к делению, и с возрастом их число
постепенно уменьшается вследствие естественной гибели. При
дегенеративных заболеваниях (болезнь Альцгеймера, Гентингтона,
паркинсонизм) интенсивность апоптоза возрастает и количество
нейронов в определённых участках нервной системы резко
уменьшается.
30.
Особенности внутреннего строения- много митохондрий (синтезируют АТФ — запасают энергию);
- большое количество рибосом (находятся на шерох. плазматич. сети, в
цитоплазме нейрона участвуют в синтезе белка);
- сильно развитая гранулярная эндоплазматическая сеть (образует
тигроид или вещество Ниссля);
- развит цитоскелет (важен для транспортировки веществ);
- неспособность к делению;
- способность получать, генерировать и перерабатывать нервные
сигналы (потенциалы действия).
Нейрон покрыт клеточной мембраной (передаёт нервные сигналы).
31.
Потенциал действияПотенциал действия («спайк») — волна возбуждения,
перемещающаяся по мембране живой клетки в виде
кратковременного изменения мембранного потенциала на
небольшом участке возбудимой клетки (нейрона или
кардиомиоцита), в результате которого наружная поверхность
этого участка становится отрицательно заряженной по отношению
к внутренней поверхности мембраны, в то время как в покое она
заряжена положительно. Потенциал действия является
физиологической основой нервного импульса.
32.
Потенциал действия (продолжение)Благодаря работе «натрий-калиевого насоса» концентрация ионов
натрия в цитоплазме клетки очень мала по сравнению с
окружающей средой. При проведении потенциала действия
открываются потенциал-зависимые натриевые каналы и
положительно заряженные ионы натрия поступают в цитоплазму
по градиенту концентрации, пока он не будет уравновешен
положительным электрическим зарядом. Вслед за этим потенциалзависимые каналы инактивируются и отрицательный потенциал
покоя восстанавливается за счёт диффузии из клетки положительно
заряженных ионов калия, концентрация которых в окружающей
среде также значительно ниже внутриклеточной.
33.
Фазы потенциала действияПредспайк — процесс медленной деполяризации мембраны до
критического уровня деполяризации (местное возбуждение, локальный
ответ).
Пиковый потенциал или спайк, состоящий из восходящей части
(деполяризация мембраны) и нисходящей части (реполяризация
мембраны).
Отрицательный следовой потенциал — от критического уровня
деполяризации до исходного уровня поляризации мембраны (следовая
деполяризация).
Положительный следовой потенциал — увеличение мембранного
потенциала и постепенное возвращение его к исходной величине
(следовая гиперполяризация).
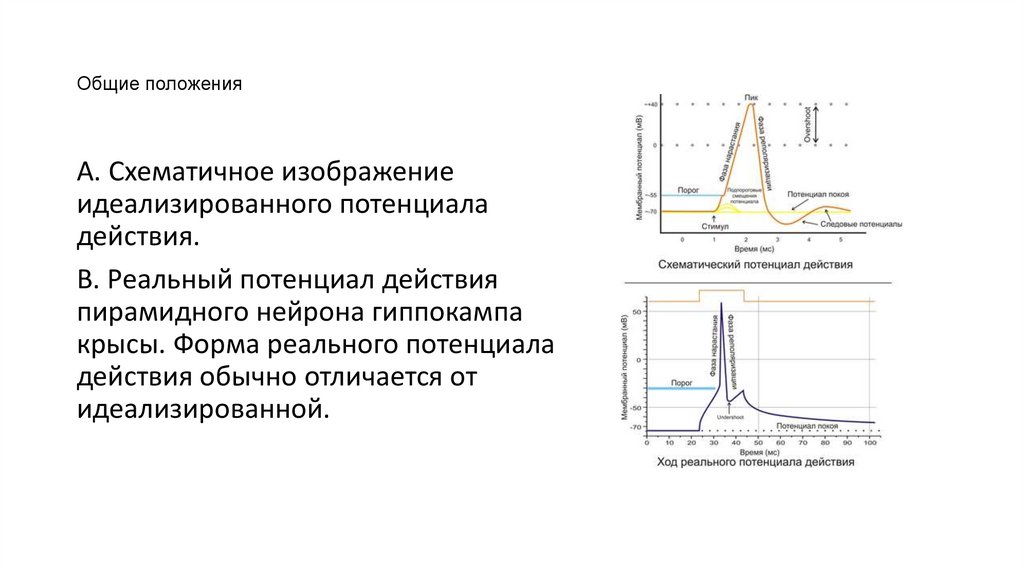
34.
Общие положенияA. Схематичное изображение
идеализированного потенциала
действия.
B. Реальный потенциал действия
пирамидного нейрона гиппокампа
крысы. Форма реального потенциала
действия обычно отличается от
идеализированной.
35.
Общие положения (продолжение)Поляризация мембраны живой клетки обусловлена отличием ионного состава
с её внутренней и наружной стороны. Когда клетка находится в спокойном
(невозбуждённом) состоянии, ионы по разные стороны мембраны создают
относительно стабильную разность потенциалов, называемую потенциалом
покоя. Если ввести внутрь живой клетки электрод и измерить мембранный
потенциал покоя, он будет иметь отрицательное значение (около −70 — −90
мВ).
Это объясняется тем, что суммарный заряд на внутренней стороне мембраны
существенно меньше, чем на внешней, хотя с обеих сторон содержатся и
катионы, и анионы. Снаружи — на порядок больше ионов натрия, кальция и
хлора, внутри — ионов калия и отрицательно заряженных белковых молекул,
аминокислот, органических кислот, фосфатов, сульфатов. Надо понимать, что
речь идёт именно о заряде поверхности мембраны — в целом среда и внутри,
и снаружи клетки заряжена нейтрально.
36.
Общие положения (продолжение)Потенциал мембраны может изменяться под действием различных
стимулов. Искусственным стимулом может служить электрический
ток, подаваемый на внешнюю или внутреннюю сторону мембраны
через электрод. В естественных условиях стимулом часто служит
химический сигнал от соседних клеток, поступающий через синапс
или путём диффузной передачи через межклеточную среду.
Смещение мембранного потенциала может происходить в
отрицательную (гиперполяризация) или положительную
(деполяризация) сторону.
37.
Общие положения (продолжение)В нервной ткани потенциал действия, как правило, возникает при
деполяризации — если деполяризация мембраны нейрона
достигает некоторого порогового уровня или превышает его,
клетка возбуждается, и от её тела к аксонам и дендритам
распространяется волна электрического сигнала.
В реальных условиях на теле нейрона обычно возникают
постсинаптические потенциалы, которые сильно отличаются от
потенциала действия по своей природе — например, они не
подчиняются принципу «всё или ничего». Эти потенциалы
преобразуются в потенциал действия на особом
немиелинезированном участке аксона — его начальном сегменте,
и затем обратно распространяется на сому нейрона и дендриты.
38.
Общие положения (продолжение)Простейшая схема, демонстрирующая мембрану
с двумя натриевыми каналами в открытом и
закрытом состоянии.
39.
Общие положения (продолжение)Это обусловлено тем, что на мембране клетки находятся ионные
каналы — белковые молекулы, образующие в мембране поры,
через которые ионы могут проходить с внутренней стороны
мембраны на наружную и наоборот.
Большинство каналов ионо-специфичны — натриевый канал
пропускает практически только ионы натрия и не пропускает
другие (это явление называют селективностью). Мембрана клеток
возбудимых тканей (нервной и мышечной) содержит большое
количество потенциал-зависимых ионных каналов, способных
быстро реагировать на смещение мембранного потенциала.
Деполяризация мембраны в первую очередь вызывает открытие
потенциал-зависимых натриевых каналов.
40.
Общие положения (продолжение)Когда одновременно открывается достаточно много натриевых
каналов, положительно заряженные ионы натрия устремляются
через них на внутреннюю сторону мембраны. Движущая сила в
данном случае обеспечивается градиентом концентрации (с
внешней стороны мембраны находится намного больше
положительно заряженных ионов натрия, чем внутри клетки) и
отрицательным зарядом внутренней стороны мембраны. Поток
ионов натрия вызывает ещё бо́льшее и очень быстрое изменение
мембранного потенциала, которое и называют потенциалом
действия (в специальной литературе обозначается ПД).
41.
Общие положения (продолжение)Согласно закону «всё-или-ничего» мембрана клетки возбудимой
ткани либо не отвечает на стимул совсем, либо отвечает с
максимально возможной для неё на данный момент силой. То
есть, если стимул слишком слаб и порог не достигнут, потенциал
действия не возникает совсем.
В то же время пороговый стимул вызовет потенциал действия
такой же амплитуды, как и стимул, превышающий пороговый. Это
отнюдь не означает, что амплитуда потенциала действия всегда
одинакова — один и тот же участок мембраны, находясь в разных
состояниях, может генерировать потенциалы действия разной
амплитуды.
42.
Общие положения (продолжение)После возбуждения нейрон на некоторое время оказывается в
состоянии абсолютной рефрактерности, когда никакие сигналы не
могут его возбудить снова, затем входит в фазу относительной
рефрактерности, когда его могут возбудить исключительно
сильные сигналы (при этом амплитуда ПД будет ниже, чем
обычно). Рефрактерный период возникает из-за инактивации
быстрого натриевого тока, то есть инактивации натриевых каналов.
43.
Распространение потенциала действияПо немиелинизированным волокнам
По немиелинизированному волокну потенциал действия
распространяется непрерывно. Проведение нервного импульса
начинается с распространением электрического поля. Возникший
потенциал действия за счет электрического поля способен
деполяризовать мембрану соседнего участка до критического
уровня, в результате чего на соседнем участке генерируются новые
потенциалы. Сам потенциал действия не перемещается, он
исчезает там же, где возник. Главную роль в возникновении нового
потенциал действия играет предыдущий.
44.
Распространение потенциала действияПо немиелинизированным волокнам
(продолжение)
Если внутриклеточным электродом раздражать аксон посередине,
то потенциал действия будет распространяться в обоих
направлениях. Обычно же потенциал действия распространяется
по аксону в одном направлении (от тела нейрона к нервным
окончаниям), хотя деполяризация мембраны происходит по обе
стороны от участка, где в данный момент возник потенциал.
Одностороннее проведение потенциала действия обеспечивается
свойствами натриевых каналов — после открытия они на
некоторое время инактивируются и не могут открыться ни при
каких значениях мембранного потенциала (свойство
рефрактерности). Поэтому на ближнем к телу клетки участке, где
до этого уже «прошел» потенциал действия, он не возникает.
45.
Распространение потенциала действияПо немиелинизированным волокнам
(продолжение)
При прочих равных условиях распространение потенциала
действия по аксону происходит тем быстрее, чем больше диаметр
волокна. По гигантским аксонам кальмара потенциал действия
может распространяться почти с такой же скоростью, как и по
миелинизированным волокнам позвоночных (около 100 м/c).
46.
По миелинизированным волокнамПо миелинизированному волокну потенциал действия распространяется
скачкообразно (сальтаторное проведение). Для миелинизированных
волокон характерна концентрация потенциалзависимых ионных
каналов только в областях перехватов Ранвье; здесь их плотность в 100
раз больше, чем в мембранах безмиелиновых волокон. В области
миелиновых муфт потенциалзависимых каналов почти нет.
Потенциал действия, возникший в одном перехвате Ранвье, за счет
электрического поля деполяризует мембрану соседних перехватов до
критического уровня, что приводит к возникновению в них новых
потенциалов действия, то есть возбуждение переходит скачкообразно,
от одного перехвата к другому. В случае повреждения одного перехвата
Ранвье потенциал действия возбуждает 2-й, 3-й, 4-й и даже 5-й,
поскольку электроизоляция, создаваемая миелиновыми муфтами,
уменьшает рассеивание электрического поля.
47.
По миелинизированным волокнам(продолжение)
«Скачкообразное распространение» увеличивает скорость
распространения потенциала действия по миелинизированным
волокнам по сравнению с немиелинизированными. Кроме того,
миелинизированные волокна толще, а электрическое сопротивление
более толстых волокон меньше, что тоже увеличивает скорость
проведения импульса по миелинизированным волокнам. Другим
преимуществом сальтаторного проведения является его экономичность
в энергетическом плане, так как возбуждаются только перехваты Ранвье,
площадь которых меньше 1 % мембраны, и, следовательно, необходимо
значительно меньше энергии для восстановления трансмембранных
градиентов Na+ и K+, расходующихся в результате возникновения
потенциал действия, что может иметь значение при высокой частоте
разрядов, идущих по нервному волокну.
48.
По миелинизированным волокнам(продолжение)
Чтобы представить, насколько эффективно может быть увеличена скорость
проведения за счёт миелиновой оболочки, достаточно сравнить скорость
распространения импульса по немиелинизированным и миелинизированным
участкам нервной системы человека. При диаметре волокна около 2 µм и
отсутствии миелиновой оболочки скорость проведения будет составлять ~1
м/с, а при наличии даже слабой миелинизации при том же диаметре волокна
— 15-20 м/с. В волокнах большего диаметра, обладающих толстой
миелинововой оболочкой, скорость проведения может достигать 120 м/с.
Скорость распространения потенциала действия по мембране отдельно
взятого нервного волокна непостоянна — в зависимости от различных условий
эта скорость может очень значительно уменьшаться и, соответственно,
увеличиваться, возвращаясь к некоему исходному уровню.
49.
Активные свойства мембраныСхема строения мембраны клетки.
50.
Активные свойства мембраны(продолжение)
Активные свойства мембраны, обеспечивающие возникновение потенциала
действия, основываются главным образом на поведении потенциалзависимых
натриевых (Na+-) и калиевых (K+-) каналов. Начальная фаза ПД формируется
входящим натриевым током, позже открываются калиевые каналы и
выходящий K+-ток возвращает потенциал мембраны к исходному уровню.
Исходную концентрацию ионов затем восстанавливает натрий-калиевый
насос.
По ходу ПД каналы переходят из состояния в состояние: у Na+-каналов
основных состояний три — закрытое, открытое и инактивированное (в
реальности дело сложнее, но этих трёх достаточно для описания), у K+каналов два — закрытое и открытое.
Поведение каналов, участвующих в формировании ПД, описывается через
проводимость и вычисляется через коэффициенты переноса (трансфера).
Коэффициенты переноса были выведены Ходжкином и Хаксли.
51.
Аксональный транспортАксональный транспорт (аксоток) — это перемещение веществ от
тела нейрона в отростки (антероградный аксоток) и в обратном
направлении (ретроградный аксоток). Различают медленный
аксональный ток веществ (1-5 мм в сутки) и быстрый (до 1-5 м в
сутки). Обе транспортные системы присутствуют как в аксонах, так
и в дендритах.
Аксональный транспорт обеспечивает единство нейрона. Он
создаёт постоянную связь между телом нейрона (трофическим
центром) и отростками. Основные синтетические процессы идут в
перикарионе. Здесь сосредоточены необходимые для этого
органеллы. В отростках синтетические процессы протекают слабо.
52.
Аксональный транспорт (продолжение)Антероградная быстрая система транспортирует к нервным окончаниям
белки и органеллы, необходимые для синаптических функций
(митохондрии, фрагменты мембран, пузырьки, белки-ферменты,
участвующие в обмене нейромедиаторов, а также предшественники
нейромедиаторов). Ретроградная система возвращает в перикарион
использованные и поврежденные мембраны и белки для деградации в
лизосомах и обновления, приносит информацию о состоянии
периферии, факторы роста нервов.
Медленный транспорт — это антероградная система, проводящая белки
и другие вещества для обновления аксоплазмы зрелых нейронов и
обеспечения роста отростков при их развитии и регенерации.
Ретроградный транспорт может иметь значение в патологии. За счёт
него нейротропные вирусы (герпеса, бешенства, полиомиелита) могут
перемещаться с периферии в центральную нервную систему.
53.
Нейроглия (глиоциты)Глиоциты выполняют в нервной ткани вспомогательные функции:
опорную, разграничительную, трофическую, секреторную и
защитную. Они поддерживают постоянство среды вокруг
нейронов. Клетки нейроглии делятся на две группы: макроглию и
микроглию. Клетки макроглии бывают трёх типов: эпендимоциты,
астроциты и олигодендроциты.
54.
Нейроглия (глиоциты) (продолжение)Эпендимоциты. Выстилают каналы и желудочки спинного и
головного мозга, по которым циркулирует спинномозговая
жидкость (ликвор). Эти клетки напоминают однослойный
призматический эпителий. На апикальных концах эпендимоцитов
расположены реснички, помогающие движению спинномозговой
жидкости. Через апикальные концы эпендимоциты могут выделять
биологически активные вещества, которые с ликвором разносятся
по всему мозгу. От базальных концов эпиндимоцитов отходят
отростки, которые могут идти через весь мозг и образуют на его
поверхности глиальную мембрану. В желудочках мозга находятся
сосудистые сплетения. Они покрыты специализированными
эпендимоцитами, участвующими в образовании ликвора.
55.
Нейроглия (глиоциты) (продолжение)Астроциты. Различают протоплазматические и волокнистые астроциты.
Протоплазматические астроциты имеют короткие толстые отростки. Они
расположены в сером веществе мозга, выполняют разграничительную и
трофическую функции. Волокнистые астроциты находятся в белом
веществе, имеют многочисленные тонкие длинные отростки, которые
оплетают кровеносные сосуды мозга, образуя периваскулярные
глиальные пограничные мембраны. Их отростки также изолируют
синапсы. Таким образом, они изолируют нейроны и кровеносные
сосуды и участвуют в образовании гемато-энцефалического барьера,
обеспечивают обмен веществ между кровью и нейронами. Они также
участвуют в образовании оболочек мозга и выполняют опорную
функцию (образуют каркас мозга).
56.
Нейроглия (глиоциты) (продолжение)Олигодендроциты имеют мало отростков, окружают нейроны,
выполняя трофическую (участие в питании нейронов) и
разграничительную функции. Олигодендроциты, расположенные вокруг
тел нейронов, называются мантийными глиоцитами. Олигодендроциты,
расположенные в периферической нервной системе и образующие
оболочки вокруг отростков нейронов, называют леммоцитами
(шванновскими клетками).
Микроглия (глиальные макрофаги). Образуются из костномозговых
предшественников моноцитов. Покоящиеся микроглиоциты имеют
короткие ветвящиеся отростки. Под действием микроорганизмов и
продуктов распада нервной ткани они активируются, теряют отростки,
округляются и превращаются в «зернистые шары» (реактивная
микроглия). При этом они, как макрофаги, уничтожают разрушенные
нервные и глиальные клетки.
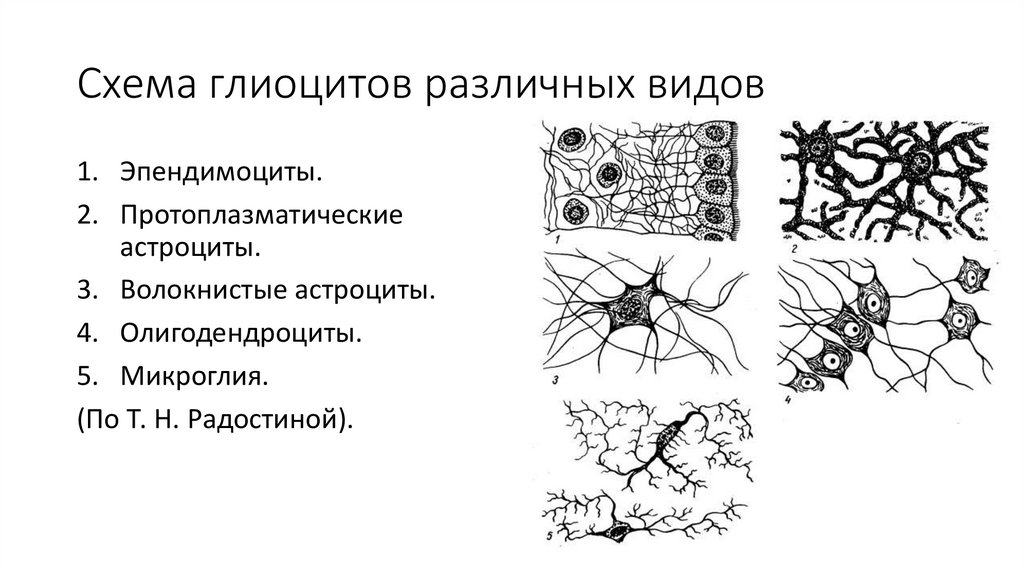
57.
Схема глиоцитов различных видов1. Эпендимоциты.
2. Протоплазматические
астроциты.
3. Волокнистые астроциты.
4. Олигодендроциты.
5. Микроглия.
(По Т. Н. Радостиной).
58.
Нервные волокнаЭто отростки нейронов, покрытые глиальными оболочками.
Отростки нейронов лежат внутри нервных волокон и называются
осевыми цилиндрами. Их окружают глиальные клетки —
олигодендроциты, которые здесь называются леммоцитами
(оболочечными клетками) или шванновскими клетками. Нервные
волокна бывают миелиновые и безмиелиновые.
59.
Нервные волокна (продолжение)Миелиновые нервные волокна. Они толще безмиелиновых, содержат только
по одному осевому цилиндру (2-20 мкм). Поскольку миелин представляет
собой обмотку из липидных мембран, на гистологических препаратах он
хорошо импрегнируется осмиевой кислотой. Остатки цитоплазмы леммоцитов
сохраняются между витками мезаксона, образуя насечки миелина. Они не
окрашиваются осмиевой кислотой и поэтому видны на фоне миелина в виде
косых светлых полос.
Поскольку в процессе образования миелиновой оболочки осевой цилиндр и
леммоциты продолжают расти, то каждый последующий, наружный виток
мезаксона шире предыдущего. Участки волокна, не покрытые миелином,
называются узловыми перехватами. Они находятся на границе между двумя
соседними леммоцитами. Соответственно, участок волокна образованный
одной глиальной клеткой именуется межузловым сегментом. Снаружи всё
волокно, включая узловые перехваты, покрыто базальной мембраной. По
миелиновым волокнам нервный импульс передается с большой скоростью, до
120 м/сек.
60.
Нервные волокна (продолжение)Безмиелиновые нервные волокна. Они состоят из тяжа
леммоцитов, которые содержат несколько (10-20) осевых
цилиндров, погруженных в леммоциты. Поэтому безмиелиновые
нервные волокна называют волокнами кабельного типа. Каждый
из них как бы подвешен на сдвоенной мембране (мезаксон), как на
брыжейке. Такие волокна чаще встречаются в вегетативной
нервной системе. Нервный импульс по ним проводится медленно
(1-2 м/сек).
61.
Строение нервных волокон на светооптическом(А, Б) и ультрамикроскопическом (а, б) уровнях
А,а. Миелиновое волокно.
Б,б. Безмиелиновое волокно.
1.
Осевые цилиндры.
2.
Миелиновый слой.
3.
Соединительная ткань.
4.
Насечка миелина.
5.
Ядро нейролеммоцита.
6.
Узловой перехват.
7.
Микротрубочки.
8.
Нейрофиламенты.
9.
Митохондрии.
10.
Мезаксон.
11.
Базальная мембрана.
(По Т. Н. Радостиной).
62.
Регенерация нейронов и нервных волоконНейроны взрослого человека и животных не способны к делению,
клеточной регенерации. Однако у них хорошо развита
внутриклеточная регенерация: обновление макромолекул и
органелл. При гибели одних нейронов сохранившиеся нейроны
гипертрофируются и берут на себя функции погибших. Возможно
также восстановление повреждённых отростков нейронов и,
соответственно, регенерация периферических нервов.
63.
Регенерация нейронов и нервных волокон(продолжение)
После перерезки нервного волокна наступает дегенерация осевого
цилиндра дистальней места повреждения. Леммоциты и
макрофаги фагоцитируют продукты распада, очищают место
провреждения, а затем размножаются и образуют тяжи — ленты
Бюнгера. На проксимальном отрезке осевого цилиндра образуется
наплыв аксоплазмы — формируется колба роста (как в
эмбриогенезе).
Осевой цилиндр растёт по дорожке из леммоцитов со скоростью 24 мм в сутки до тех пор, пока не достигает иннервируемого органа.
После этого вокруг новообразованного осевого цилиндра
леммоциты образуют миелиновую оболочку, а в рабочем органе
вновь формируется (восстанавливается) нервное окончание.
64.
Регенерация нейронов и нервных волокон(продолжение)
Эти процессы завершаются в течение нескольких месяцев от
момента повреждения. Однако, если возникает препятствие на
пути роста осевых цилиндров, они начинают расти беспорядочно и
образуют клубок, называемый ампутационной невромой. При её
раздражении возникает сильная боль, которая воспринимается как
происходящая из первоначально иннервируемой области,
например как боль в удалённой конечности (фантомные боли).
65.
Нервные окончанияВсе нервные волокна заканчиваются концевыми аппаратами,
называемых нервными окончаниями. По функции они делятся на
эффекторные, рецепторные и межнейрональные синапсы.
Нервный импульс в организме человека обычно передаётся с
одной нервной клетки на другую или с нейрона на рабочий орган
через медиатор, химический посредник. Медиатор
взаимодействует со специфическими рецепторами другого
нейрона или клетками рабочего органа и через целый каскад
вторичных внутриклеточных посредников, меняет функцию
рабочего органа или другого нейрона.
66.
Нейроглия: классификация и функцииПространство между нейронами заполнено клетками, которые
называются нейроглией. Они происходят из общего
нейробластического зачатка. В отличие от нейронов клетки
нейроглии делятся в течение всей жизни человека. Количество
глиальных клеток превосходит количество нейронов в 10 раз.
Функции: опорная, трофическая. Разграничительная, поддержание
гомеостаза вокруг нейронов, защитная, секреторная.
Клетки нейроглии являются как бы упаковочным материалом для
тонкой сети нейронов, предохраняя ее от механических
воздействий.
67.
Нейроглия: классификация и функции(продолжение)
Клетки нейроглии подразделяются на макроглию, к которой
относятся:
• астроциты (имеют звездчатую форму. Питание, выведение
вредных веществ образуют непосредственный контакт между
нейроном и кровеносным сосудом. Некоторые из таких
отростков оканчиваются на кровеносных сосудах. Участвуют в
регуляции ионного состава нервной ткани, влияют на активность
нейронов, а также обеспечивают восстановление нервов после
повреждения);
• олигодендроциты (образование миелиновых оболочек, роль
биологического изолятора);
68.
Нейроглия: классификация и функции(продолжение)
• эпендимоциты (выстилают все внутренние полости в головном и
спинном мозге. Выполняют функцию барьера между тканью мозга и
омывающей его жидкостью — ликвором. Регулируют секрецию и
состав ликвора);
• микроглию (обеспечивают иммунитет в нервной системе. По
происхождению они является клетками крови, способными к
фагоцитозу. То есть микроглия отличается от макроглии, которая имеет
тоже происхождение, что и нейроны. Способны захватывать
чужеродных агентов (бактерий, вирусов), а также продукты распада
нервной ткани и обезвреживать их).
Эти клетки формирует гемато-энцефалический барьер. Наличие этого
барьера не позволяет токсическим веществам проникать в нервную
систему.
69.
Нейроглия: классификация и функции(продолжение)
70.
Строение нервного волокнаКлассификация нервных волокон
Нервные волокна — отростки нервных клеток (нейронов),
имеющие оболочку и способные проводить нервный импульс.
Миелиновое нервное волокно имеет в своём составе следующие
элементы (структуры):
1) осевой цилиндр, располагающийся в самом центре нервного
волокна;
2) миелиновую оболочку, покрывающую осевой цилиндр;
3) шванновскую оболочку.
71.
Строение нервного волокнаКлассификация нервных волокон
(продолжение)
Осевой цилиндр состоит из нейрофибрилл. Мякотная оболочка
содержит большое количество веществ липоидного характера,
известных под названием миелина. Миелин обеспечивает
быстроту проведение нервных импульсов. Миелиновая оболочка
покрывает осевой цилиндр не на всём промежутке, образуя
промежутки, получившие название перехваты Ранвье. В области
перехватов Ранвье осевой цилиндр нервного волокна примыкает к
верхней — шванновской оболочке.
72.
Строение нервного волокнаКлассификация нервных волокон
(продолжение)
Промежуток волокна, расположенный между двумя перехватами
Ранвье, называют сегментом волокна. В каждом таком сегменте на
окрашенных препаратах можно видеть ядро шванновской
оболочки. Оно лежит приблизительно посредине сегмента и
окружено протоплазмой шванновской клетки, в петлях которой и
содержится миелин.
Между перехватами Ранвье миелиновая оболочка также не
является сплошной. В толще ее обнаруживаются так называемые
насечки Шмидт-Лантермана, идущие в косом направлении.
73.
Строение нервного волокнаКлассификация нервных волокон
(продолжение)
Клетки шванновской оболочки также, как и нейроны с отростками,
развиваются из эктодермы. Они покрывают осевой цилиндр
нервного волокна периферической нервной системы аналогично
тому, как клетки глии покрывают нервное волокно в центральной
нервной системе. В результате этого они могут называться
периферическими глиальными клетками.
В центральной нервной системе нервные волокна не имеют
шванновских оболочек. Роль шванновских клеток здесь выполняют
элементы олигодендроглии. Безмиелиновое (безмякотное)
нервное волокно лишено миелиновой обкладки и состоит только
из осевого цилиндра и шванновской оболочки.
74.
Функция нервных волоконГлавная функция нервных волокон — передача нервного импульса. В
настоящее время изучено два типа нервной передачи: импульсная и
безимпульсная. Импульсная передача обеспечивается электролитными
и нейротрансмиттерными механизмами. Скорость передачи нервного
импульса в миелиновых волокнах значительно выше, чем в
безмякотных. В её осуществлении важнейшая роль принадлежит
миелину. Данное вещество способно изолировать нервный импульс, в
результате чего передача сигнала по нервному волокну происходит
скачкообразно, от одного перехвата Ранвье к другому. Безимпульсная
передача осуществляется током аксоплазмы по специальным
микротрубочкам аксона, содержащим трофогены — вещества,
оказывающие на иннервируемый орган трофическое влияние.
75.
Различают миелиновые и безмиелиновые волокна. Отросток —осевой цилиндр (аксон). В ЦНС оболочки отростков образуют
олигодендроциты, в ПНС — нейролеммоциты.
Безмиелиновые нервные волокна. В составе вегетативной нервной
системы. Волокна, содержащие несколько осевых цилиндров (1020 в НВ внутренних органов) — волокна кабельного типа. Оболочка
нейролеммоцита прогибается, его края над осевым цилиндром
сближаются и образуют сдвоенную мембрану — мезоаксон.
Передача импульса со скоростью 1-2 м/с.
76.
Миелиновые нервные волокна. В ЦНС и ПНС, диаметр 2-20 мкм.Состоят из осевого цилиндра, одетого оболочкой из шванновских
клеток. Различают 2 слоя: миелиновый внутренний и наружный,
состоящий из цитоплазмы, ядер нейроллеммоцитов и
нейролеммы.
Миелиновый слой содержит много липидов, встречаются насечки
миелина (Шмидта-Лантермана), через определенные интервалы
встречаются безмиелиновые участки — перехваты Ранвье.
77.
Периферическая нервная система: в процессе развития аксон погружается воболочку нейролеммоцита, края смыкаются — образуется мезоаксон, который
формирует миелиновый слой, ветвление аксонов происходит в области
перехватов. Межузловой сегмент — участок между перехватами.
Миелиновые волокна ЦНС — миелиновый слой формируется одним из
отростков олигодендроглиоцита. Не имеют насечек миелина, нервные
волокна не окружены БМ. Миелин содержит миелиновый щелочной белок и
протеолипидный белок. Передача импульса 5-120 м/с.
При травме распадается миелиновый слой и осевой цилиндр, продукты
распада нейтрализуются макрофагами за 1 неделю. В ЦНС не регенерируют, в
ПНС — хорошая регенерация. Ближайшие нейролеммоциты пролиферируют,
осевые цилиндры пускают множество отростков в нейролеммоциты, не
достигшие цели — погибают, иногда эти отростки сплетаются и образуют
ампутационную неврому.
78.
Межклеточные контакты: химические иэлектрические синапсы
Медиатор, находящийся в пузырьках, выделяется в синаптическую
щель с помощью экзоцитоза (пузырьки подходят к мембране,
сливаются с ней и разрываются, выпуская медиатор). Его
выделение происходит небольшими порциями — квантами.
Каждый квант содержит от 1.000 до 10.000 молекул
нейромедиатора. Небольшое количество квантов выходит из
окончания и в состоянии покоя.
Когда нервный импульс, т. е. ПД, достигает пресинаптического
окончания, происходит деполяризация его пресинаптической
мембраны. Открываются ее кальциевые каналы и ионы кальция
входят в синаптическую бляшку. Начинается выделение большого
количества квантов нейромедиатора.
79.
Межклеточные контакты: химические иэлектрические синапсы (продолжение)
Молекулы медиатора диффундируют через синаптическую щель к
постсинаптической мембране и взаимодействуют с ее
хеморецепторами. В результате образования комплексов
медиатор-рецептор, в субсинаптической мембране начинается
синтез так называемых вторичных посредников. В частности цАМФ.
Эти посредники активируют ионные каналы постсинаптической
мембраны. Поэтому такие каналы называют хемозависимыми или
рецепторуправляемыми. Т. е. они открываются при действии ФАВ
на хеморецепторы. В результате открывания каналов изменяется
потенциал субсинаптической мембраны. Такое изменение
называется постсинаптическим потенциалом.
80.
Электрические синапсыПредставляют собой щелевидное образование (размеры щели до 2 нм) с
ионными мостиками-каналами между двумя контактирующими клетками.
Передача импульса при помощи специализованных белков. Плотный
контакт между пост- и пресинаптическими мембранами.
Петли тока, в частности при наличии потенциала действия (ПД), почти
беспрепятственно перескакивают через такой щелевидный контакт и
возбуждают. В целом, такие синапсы обеспечивают очень быструю передачу
возбуждения. Но в то же время с помощью этих синапсов нельзя обеспечить
одностороннее проведение, т. к. большая часть таких синапсов обладает
двусторонней проводимостью. Кроме того, с их помощью нельзя заставить
эффекторную клетку (клетку, которая управляется через данный синапс)
тормозить свою активность. Аналогом электрического синапса в гладких
мышцах и в сердечной мышце являются щелевые контакты типа нексуса.
81.
Химические синапсыПо строению химические синапсы представляют собой окончания
аксона (терминальные синапсы) или его варикозную часть
(проходящие синапсы), которая заполнена химическим веществом
— медиатором. В синапсе различают пресинаптический элемент,
который ограничен пресинаптической мембраной,
постсинаптический элемент, который ограничен
постсипаптической мембраной, а также внесинаптическую область
и синаптическую щель, величина которой составляет в среднем 50
нм.
Односторонняя проводимость.
82.
Демиелинизирующие заболеванияДемиелинизирующие заболевания — это любые заболевания нервной
системы, при которых повреждается миелиновая оболочка нейронов. Это
повреждение нарушает проведение сигналов по пострадавшим нервам. В
свою очередь, снижение проводящей способности приводит к нарушениям
чувствительности, движений, когнитивных функций и других функций в
зависимости от того, какие затронуты нервы.
Демиелинизирующие заболевания могут вызываться генетическими
факторами, возбудителями инфекций, аутоиммунными реакциями, а также
другими неизвестными факторами. Отравление фосфорорганическими
соединениями из состава коммерческих инсектицидов, таких как препараты
для купания овец, гербицидами и препаратами против блох для домашних
животных может привести к демиелинизации нервов. Хронический приём
нейролептиков также может вызывать демиелинизацию. Дефицит витамина
B12 также может приводить с демиелинизации.
83.
Демиелинизирующие заболевания(продолжение)
Демиелинизирующие заболевания традиционно делятся на два типа:
демиелинизирующие миелинокластические заболевания и
демиелинизирующие лейкодистрофические заболевания. В первой
группе нормальный и здоровый миелин разрушается под воздействием
токсина либо аутоиммунной реакции. Во второй группе миелин
ненормален и происходит его дегенерация. Вторая группа названа
Позером дисмиелинизирующими заболеваниями.
Имеются доказательства того, что собственная иммунная система
организма как минимум частично ответственна за развитие наиболее
известного демиелинизирующего заболевания — рассеянного склероза.
Известно, что клетки приобретённого иммунитета, называемые Tлимфоцитами, присутствуют в очагах поражения. Другие клетки
иммунной системы, называемые макрофагами (и, возможно, тучными
клетками) также вносят вклад в повреждение.
84.
КлассификацияДемиелинизацию можно разделить на 2 типа:
миелинопатия — разрушение уже сформированного
миелина из-за причин, связанных с биохимическим дефектом
строения миелина, как правило, генетически обусловленным
(например, болезнь Канавана);
миелинокластия — разрушение нормально
синтезированного миелина под влиянием различных воздействий,
как внешних, так и внутренних (например, болезнь Бинсвангера).
85.
Классификация (продолжение)Подразделение на эти две группы весьма условно, так как первые
клинические проявления миелинопатий могут быть связаны с
воздействием различных внешних факторов, а миелинокластии
вероятнее всего развиваются у предрасположенных лиц.
Кроме того, демиелинизирующие заболевания можно разделить
на те, которые затрагивают центральную нервную систему, и те,
которые повреждают периферическую нервную систему.
86.
Нарушения, влияющие на ЦНС, включают всебя
рассеянный склероз, болезнь Девика, концентрический
склероз Бало, а также другие нарушения с участием иммунной
системы, называемые воспалительными демиелинизирующими
заболеваниями;
синдром осмотической демиелинизации;
миелопатии, например, спинная сухотка;
лейкоэнцефалопатии, например, ПМЛ;
лейкодистрофии и др.
87.
Демиелинизирующие заболеванияпериферической нервной системы включают
синдром Гийена-Барре;
невральная амиотрофия Шарко-Мари-Тута и др.
88.
Клиническая картинаКлинические картина складывается из нарастающих
неврологических симптомов. На ранних этапах, когда некоторые
нервные волокна уже поражены, объективные и субъективные
симптомы могут не проявляться. Это связано с компенсацией их
функций здоровыми клетками. Для разных демиелинизирующих
заболеваний симптомы могут существенно различаться и зависят
от локализации пораженных областей в центральной или
периферической нервной системе.
89.
Клиническая картина (продолжение)Обычными симптомами считаются:
Гемипарез
Клонические судороги
Гипертермия
Кома
Головокружение
Неврит зрительного нерва
Головная боль
Нарушения координации движений
Дизартрия
Нарушение равновесия
Диплопия
Парастезии
Депрессия
Парапарез
Нарушения зрения
Энурез
90.
ДиагностикаДиагностика демиелинизирующих заболеваний сложна и должна
основываться на всестороннем анализе симптоматики, результатах
клинических исследований:
магнитно-резонансная томография — метод визуализации мягких
тканей, основанный на явлении ядерного магнитного резонанса. Он позволяет
выявить очаги воспаления, но является ненадежным, потому что оценивает
воспаление по косвенному признаку — изменению концентрации протонов,
которая может быть вызвана другими причинами;
анализ вызванных потенциалов, регистрируемых с помощью
электроэнцефалографии или другими методами электрофизиологической
записи;
анализ спиномозговой жидкости — позволяет судить о воспалении и
иммунологических расстройствах на основании наличия олигоклональных
иммуноглобулинов IgG. Кроме того, позволяет выявить инфекционные
поражения ЦНС;
91.
Диагностика (продолжение)МР-спектроскопия и ЯМР-спектроскопия для выявления
специфических биомаркеров: CHI3L1, CXCL13, МикроРНК miR-223,
N-Ацетиласпартата, NEFL и других;
электромиография — для диагностики демиелинизирующих
заболеваний в периферической нервной системе.
Входящие в эту группу заболевания имеют собственные наборы
диагностических критериев, которые позволяют провести
дальнейшую дифференциацию и постановку конкретного
диагноза.
92.
У животныхДемиелинизирующие заболевания были найдены во всем мире у
различных животных. Это мыши, свиньи, крупный рогатый скот,
хомяки, крысы, овцы, сиамские кошки и ряд пород собак (в том
числе: чау-чау, спрингер-спаниель, далматин, самоедов,
золотистый ретривер, бернский зенненхунд, веймаранер,
австралийский шелковистый терьер и другие). Также данное
заболевание было обнаружено у северных морских котиков.
93.
Основные медиаторные системы нервнойсистемы
Нейромедиа́торы (нейротрансмиттеры, посредники) — биологически
активные химические вещества, посредством которых осуществляется
передача электрического импульса с нервной клетки через
синаптическое пространство между нейронами. Нервный импульс,
поступающий в пресинаптическое окончание, вызывает освобождение в
синаптическую щель медиатора.
Высвобожденный нейромедиатор затем диффундирует через щель и
связывается с рецепторами на постсинаптической мембране. Диффузия
является медленным процессом, но пересечение такой короткой
дистанции, которая разделяет пре- и постсинаптические мембраны (0,1
мкм или меньше), происходит достаточно быстро и позволяет
осуществлять быструю передачу сигнала между нейронами или между
нейроном и мышцей.
94.
Основные медиаторные системы нервнойсистемы (продолжение)
Синапс — это функциональный контакт специализированных
участков плазматических мембран двух возбудимых клеток. Синапс
состоит из пресинаптической мембраны, синаптической щели и
постинаптической мембраны. Мембраны клеток в месте контакта
имеют утолщения в виде бляшек — нервных окончаний. Нервный
импульс, достигший нервного окончания, не в состоянии
преодолеть возникшее перед ним препятствие — синаптическую
щель. После этого электрический сигнал преобразуется в
химический. Пресинаптическая мембрана содержит специальные
канальные белки, подобные белкам, формирующим натриевый
канал в мембране аксона.
95.
Основные медиаторные системы нервнойсистемы (продолжение)
Голубое пятно в сером веществе моста, ядро из скопления
норадренергических нейронов, имеет голубоватый оттенок за счет
меланина. Участвует в управлении общим уровнем активности НС,
в основном в сторону ее повышения, управляет эмоциональной
памятью и различными вегетативными функциями.
Межножковое ядро лежит над задним продырявленным
веществом в покрышке среднего мозга Это ядро содержит
норадренергические нейроны и работает в контакте с голубым
пятном, дополняя его эффекты.
96.
Основные медиаторные системы нервнойсистемы (продолжение)
Черная субстанция («substantia nigra») в покрышке среднего мозга называется
так потому, что часть ее клеток содержит темный пигмент меланин. Это ядро
также состоит из двух частей — медио-дорсальной компактной и
вентролатеральной ретикулярной. В компактной части находятся
дофаминергические нейроны, медиатором клеток ретикулярной части
является ГАМК (гамма-аминомасляная кислота). Эти две части отличаются и
своими многочисленными связями. Так компактная часть имеет двусторонние
связи с базальными ганглиями. Афферентами ретикулярной части служат
базальные ганглии, а также субталамус. Сама же она посылает эфференты в
средний мозг (к глазодвигательным ядрам) и таламус.
Черная субстанция также входит в экстрапирамидную систему. Это ядро
принимает участие в регуляции общего уровня двигательной активности
посредством влияния на структуры конечного мозга, хвостатое ядро и
скорлупу.
97.
Основные медиаторные системы нервнойсистемы (продолжение)
В срединной части ствола мозга расположена ретикулярная формация (РФ) —
скопление нейронов разных размеров и формы, разделенных множеством
проходящих в разных на правлениях волокон, напоминающих сеть (лат.
«reticulum»).
РФ играет ключевую роль в управлении общим уровнем активности нервной
системы, в частности в регуляции цикла «сон-бодрствование». Через пути,
связывающие РФ со спинным мозгом, она принимает участие в управлении
позой, локомоцией и целенаправленными движениями.
Ядра РФ участвуют также в регуляции, связанной с жизненно важными
рефлексами. Так, в РФ продолговатого мозга и моста находятся центры
дыхания (с подразделением на центр вдоха и центр выдоха),
сосудодвигательный центр (регулирующий тонус сосудов и работу сердца),
центр слюноотделения и выделения других пищеварительных соков, центр
глотания, а также центры таких защитных рефлексов, как кашель, чихание,
рвота.
98.
Основные медиаторные системы нервнойсистемы (продолжение)
Центральное серое вещество (ЦСВ) лежит в центре среднего мозга
вокруг мозгового водопровода, образуя слой около 2 мм. ЦСВ тесно
взаимодействует с ядрами шва, управляя работой их нейронов. Одна из
функций ЦСВ связана с регуляцией болевой чувствительности. При
раздражении его нейронов возможно обезболивание за счет влияний
на зоны спинного мозга, связанные с переключением болевых сигналов.
ЦСВ может оказывать целый ряд тормозных влияний на гипоталамус и
кору больших полушарий. Кроме того, центральное серое вещество
рассматривается как один из главных центров сна.
Ядра шва — структуры, входящие в Медианную зону РФ, протянутой от
продолговатого мозга до каудальных (задних) отделов среднего мозга.
Это Серотонинергические нейроны, они находятся в тесном
взаимодействии с центральным серым веществом покрышки среднего
мозга (участвует в регуляции уровней активации НС).