

























 Биология
БиологияПохожие презентации:









")
Энергетика мышечного сокращения
1.
Энергетика мышечного сокращенияАвтор-составитель :
Проф. Е.О. Данченко
ВГУ имени П.М. Машерова
2.
Сокращение мышечных волокон обусловленопродольным скольжением толстых миозиновых
и тонких актиновых филаментов относительно
друг друга
Осуществляется следующий цикл реакций:
1. Головка
молекулы
миозина
прочно
связывается с открытым участком актиновой
нити,
затем
Са2+
инициирует
взаимодействие головки с АТФ и головка
отделяется от актиновой нити (комплекс
миозин-АТФ обладает низким сродством к
актину
3.
2. Са2+-зависимая АТФ-аза головки миозинагидролизует АТФ, но оба продукта реакции
(АДФ и Рн) близко друг от друга.
3. Энергия, выделяющаяся при гидролизе АТФ,
вызывает аллостерические изменения в
миозиновой головке и головка образует
новый мостик с соседней молекулой актина.
4. Актин ускоряет выброс продуктов АТФ-азной
реакции из активного центра миозина. Это
приводит к изменению конформации головки
миозина, которая действует подобно «гребку
весла»
4.
5. Во время «гребка» миозиновые головкиотклоняются на определенный угол от оси и
перемещают миозиновый филамент вдоль
актинового филамента по направлению к Zдиску.
6. АДФ и Рн отсоединяются от миозина. Цикл
повторяется до тех пор, пока имеется АТФ.
7. В отсутствии последующего стимулирования
АТФ-зависимые кальциевые насосы
мембраны СПР быстро снижают
концентрацию ионов Са2+ до исходного
уровня.
5.
6.
Сокращение мышечных волокон обеспечивается за счет кооперативной работыбольшого количества молекул миозина, собранных в толстые нити. Движение пучка
молекул миозина вдоль нити актина можно сравнить с перетаскиванием бревна
работниками, из которых лишь небольшая часть тружеников (10–15%) опирается
ногами на землю. Подобно головкам миозина, работники периодически меняются
ролями, однако в каждый момент времени активно работает лишь небольшая часть
тружеников. Это биохимический механизм тренировочного процесса.
7.
Модель акто-миозинового комплекса:весла как головки миозина обеспечивают перемещение
лодки
8.
9.
Энергетическое обеспечение деятельностимышц
1) АТФ. АТФ → АДФ + энергия
Запас АТФ расходуется за 1-2 секунды.
2) Креатинфосфат (КФ)
Быстрая регенерация АТФ достигается за счет
переноса креатинкиназой фосфатной группы с КФ на
АДФ. Энергетический резерв креатинфосфата также
расходуется за несколько секунд.
креатинкиназа
Креатинфосфат + АДФ → креатин + АТФ
После
завершения
нагрузки
запасы
АТФ
и
креатинфосфата в мышцах восстанавливаются на 70%
через 30 секунд и полностью – через 3-5 минут
10.
3) Анаэробный гликолизВ
мышечной
ткани
относительно
долгосрочным
энергетическим
резервом
является гликоген
В покоящейся мышце содержание гликогена
составляет до 2% от мышечной массы
Гликоген расщепляется до глюкозо-6фосфата, который путем гликолиза
превращается в пируват
При недостатке кислорода пируват
восстанавливается в лактат, который
диффундирует в кровь (цикл Кори).
11.
Цикл Кори11
12.
4) Окислительное фосфорилированиеВ аэробных условиях пируват поступает в
митохондрии, окисляется в цикле Кребса и
дыхательной цепи.
Окислительное фосфорилирование –
постоянно действующий путь синтеза АТФ.
Это самый эффективный путь синтеза АТФ,
но он реализуется при условии хорошего
снабжения мышц кислородом.
Для синтез АТФ используются
«энергоносители» крови: глюкоза, жирные
кислоты и кетоновые тела.
13.
Окисление жиров происходит по уравнению:Жиры + кислород + АДФ → АТФ + углекислый газ
+ вода.
Распад глюкозы протекает в 2 этапа:
Глюкоза + АДФ → АТФ + молочная кислота.
Молочная кислота + кислород + АДФ → АТФ +
углекислый газ + вода.
Кислород требуется только на втором шаге:
если его достаточно, молочная кислота не
накапливается в мышцах.
14.
5) Образование инозинмонофосфата (ИМФ)Другим источником быстрого восстановления
АТФ является превращение АДФ в АТФ и
АМФ (2 АДФ ↔ АТФ +АМФ), катализируемая
миокиназой.
Образовавшийся АМФ за счет
дезаминирования частично превращается в
ИМФ, что сдвигает реакцию в сторону
образования АТФ
15.
Типы мышечных волоконВолокна скелетных мышц имеют разные
механические и метаболические свойства.
Различные типы мышечных волокон
определяют по максимальной скорости их
сокращения (быстрой и медленной) и
главного метаболического пути, который они
используют для образования
АТФ (окислительный и гликолитический).
16.
I тип: медленные окислительные (МО) (красныеволокна).
Медленные, тонкие, слабые, неутомляемые
мышечные волокна.
В обеспечении этих мышц кислородом
принимает участие миоглобин.
Они имеют хорошее кровоснабжение, много
митохондрий, высокую активность ферментов
окислительного фосфорилирования
Предназначены для работы в аэробном режиме
Например, такие мышцы служат для
поддержания тела в определенном положении
(поза, осанка)
17.
II тип: быстрые гликолитические волокна (белыеволокна)
Главный источник образования АТФ анаэробный гликолиз.
Толще, чем мышечные волокна I типа,
отличаются быстрыми сокращениями,
развивают большую силу и быстрее утомляются.
Такие мышцы обеспечивают кратковременную
работу максимальной мощности.
Эти волокна хуже кровоснабжаются и имеют
меньше митохондрий, липидов и миоглобина.
Мышцы истощаются в результате изменения рН
за счет повышения концентрации лактата
18.
IIа тип: быстрые окислительно-гликолитические(или просто быстрые окислительные)
Промежуточные волокна, средней толщины.
Более выносливы, чем волокна IIb типа, но
утомляются быстрее, чем волокна I типа.
Способны к выраженному сокращению, при
этом развивают среднюю силу.
Источники энергии: как окислительные,
так анаэробные механизмы (быстрые
окислительные волокна).
19.
IIb тип: быстрые гликолитические волокнаКрупные, быстрые, сильные,
быстроутомляемые мышечные волокна.
Активируются при кратковременных
нагрузках и развивают большую силу.
Получают энергию через процессы
анаэробного окисления, источником энергии
является гликоген.
В этих волокнах обнаруживают большое
количество гликогена и мало митохондрий.
20.
Тренировки не влияют на соотношение быстрых имедленных волокон.
У не тренирующихся людей обычно быстрые волокна
- гликолитические и промежуточные, а медленные –
окислительные.
Однако при правильных тренировках на увеличение
выносливости быстрые мышечные волокна
превращаются из гликолитических в промежуточные.
Также возможен переход промежуточных волокон в
окислительные.
При силовых тренировках промежуточные волокна
могут переходить в гликолитические.
При этом соотношение медленных и быстрых
волокон генетически предопределено практически не
меняется вне зависимости от тренировок (переход не
более 1-3%).
21.
У человека нет специализированных мышц,но есть специализированные волокна всех
видов: в мышцах разгибателях больше
«белых» волокон, в мышцах спины больше
«красных» волокон.
Процентное содержание тех или иных
волокон предопределяет специализацию
спортсмена
Как правило, обладатели преимущественно
красной мускулатуры достигают лучших
результатов в видах на выносливость
(плавание, велосипедный спорт, бег на
средние и длинные дистанции)
22.
Главным источником энергии для мышц всостоянии покоя служит β-окисление жирных
кислот, при интенсивной работе – метаболизм
углеводов.
После жира основным резервом запасенной
энергии служат белки скелетных мышц.
Это объясняет большую потерю мышечной
массы (особенно у взрослых людей),
наблюдающуюся при длительной
калорической недостаточности.
23.
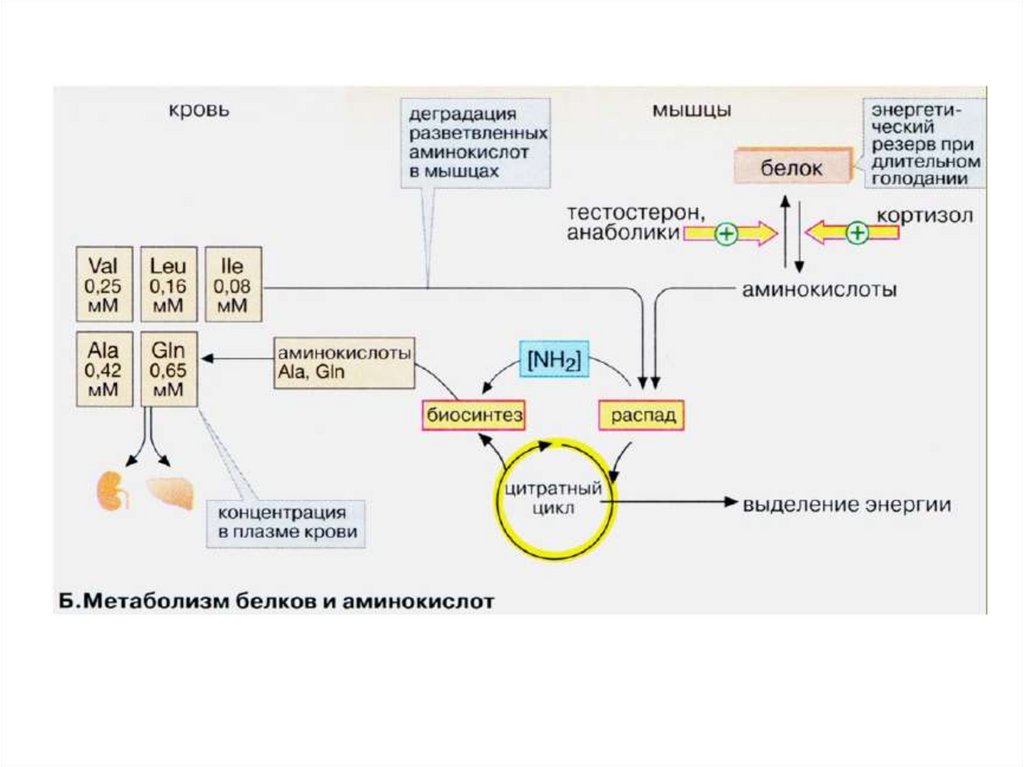
Аминокислоты, высвобождающиеся привнутриклеточном распаде белков,
реутилизируются для синтеза белка в клетке
или переносятся к другим органам для
обеспечения анаболических процессов.
Мышцы обладают высокой активностью в
отношении деградации одних и синтеза
других аминокислот.
Мышцы – главное место катаболизма
аминокислот с разветвленной цепью.
24.
Источником почти всего пирувата, идущего насинтез аланина, является гликолиз
экзогенной глюкозы.
Формируется глюкозо-аланиновый цикл, в
котором аланин мышц переносится в печень
и используется в процессе печеночного
глюконеогенеза и в то же время доставляет в
печень аминогруппы, удаляемые в виде
мочевины.
25.
Углеродный скелет аминокислот, подвергшихсядеградации и включившихся в цикл
трикарбоновых кислот в мышечной ткани,
превращается в глутамин и пируват.
ПВК далее окисляется в ЦТК или превращается
в лактат.
После всасывания большая часть аминокислот,
образующихся в процессе распада мышечного
белка, покидает мышцы.
Исключением являются изолейцин, валин,
глутамат, аспартат и аргинин. Они участвуют в
образовании глутамина, а он уже покидает
мышцы и используется другими тканями.