


























 Биология
БиологияПохожие презентации:










Гистология, цитология и эмбриология: предмет, задачи, основные теоретические положения, методы исследования
1.
Тема лекции: Гистология, цитология и эмбриология: предмет, задачи, основныетеоретические положения, методы исследования. Элементы тканей. Цитология.
Гистология (histos, греч. – ткань, logos – учение) – наука, изучающая
закономерности развития, строения и жизнедеятельности тканей
в ходе их исторического и
индивидуального развития.
Ткань – это
филогенетически
обусловленная система
гистологических элементов
(клеток и межклеточного
вещества), объединенных
общей структурой, функцией
и происхождением.
Заварзин Алексей Алексеевич
(1886-1845)
Ткань – фило- и онтогенетически обусловленная система
клеток и их производных, объединенных строением, особенностями
жизнедеятельности (функция, обновление) и происхождением
2.
Классификация тканейЭпителиальные ткани – занимают
пограничное положение в организме и
обеспечивают обмен со средой.
Ткани внутренней среды – формируют
опорные структуры и поддерживают
гомеостазис.
Мышечные ткани – обеспечивают
сокращение и движение органов и всего
организма.
Ткани нервной системы – получают
информацию и управляют работой мышц
и внутренних органов
Франц Лейдиг “Анатомические и гистологические
исследования над рыбами и рептилиями” (1853)
Алберт фон Келикер
3.
4 типа тканей многоклеточных организмов4.
Основные задачи гистологии, как науки:1. изучение закономерностей цито- и гистогенеза, строения и
функций клеток и тканей;
2. изучение закономерностей дифференциации и регенерации
тканей;
3. выяснение роли нервной, эндокринной и иммунной систем
организма, в регуляции процессов онтогенеза клеток, тканей и
органов;
4. исследование возрастных изменений клеток, тканей, органов;
5. исследование адаптации клеток, тканей, органов к действию
различных биологических, физических, химических и других
факторов;
6. изучение процессов морфогенеза в системе мать-плод;
7. исследование особенностей эмбриогенеза человека.
5.
Цитология (kytos, греч. – клетка; logos – учение) – наука о клетке.Она включает рассмотрение вопросов о развитии, строении и функциях клеток
и их производных, а также механизмов воспроизведения и взаимодействия.
Эмбриология (embryon, греч. – зародыш; logos – учение) – наука о
закономерностях развития зародыша.
Гистология, цитология и эмбриология, объединенные в одну дисциплину, решает
главную задачу – выяснение источников и закономерностей развития тканей,
их реактивности и регенерации, и в связи с этим– возможность целенаправленного
воздействия на ткани.
Основные теоретические положения гистологии, цитологии и эмбриологии
1.Клеточная теория Маттиас Шлейден и Теодор Шванн (1838), Рудольф Вирхов (1855)
* клетка — элементарная, структурно-функциональная единица всего живого.
* единая система, она включает множество закономерно связанных между собой
элементов – органелл.
* Клетки всех организмов гомологичны.
* Клетка происходит только путём деления материнской клетки.
Современные добавления к положениям клеточной теории:
* Клетки прокариот и эукариот не полностью гомологичны друг другу.
* В основе деления клетки заложено копирование нуклеиновых кислот.
* Клетки многоклеточных тотипотентны, но отличаются экспрессией различных генов,
что приводит к их морфологическому и функциональному разнообразию —
к дифференцировке.
6.
2. Теория гистогенезаДетерминация – необратимый процесс, определяющий направление развития клеток тканей из
эмбриональных зачатков
Ступенчатое, согласованное с программой развития организма, ограничение возможных путей
развития вследствие детерминации называется коммитированием
Дифференцировка направлена на создание в многоклеточном организме (человека) более 206
структурно-функциональных видов клеток
7.
3. Теория зародышевых листковПервые сведения о зародышевых листках – Карл Вольф, Максим Бер.
Образование зародышевых листков стали рассматривать как первый признак
дифференцировки зародыша.
В 1901г. А.О. Ковалевским была сформулирована теория зародышевых листков:
· зародышевые листки имеются у всех многоклеточных;
· ткани и органы образуются из зародышевых листков;
· одни и те же листки у животных дают начало одним и тем же структурам;
· для всех многоклеточных характерны одни закономерности развития.
После образования зародышевых листков их клетки специализируются в направлении
определенных эмбриональных зачатков, а далее – определенных тканей
8.
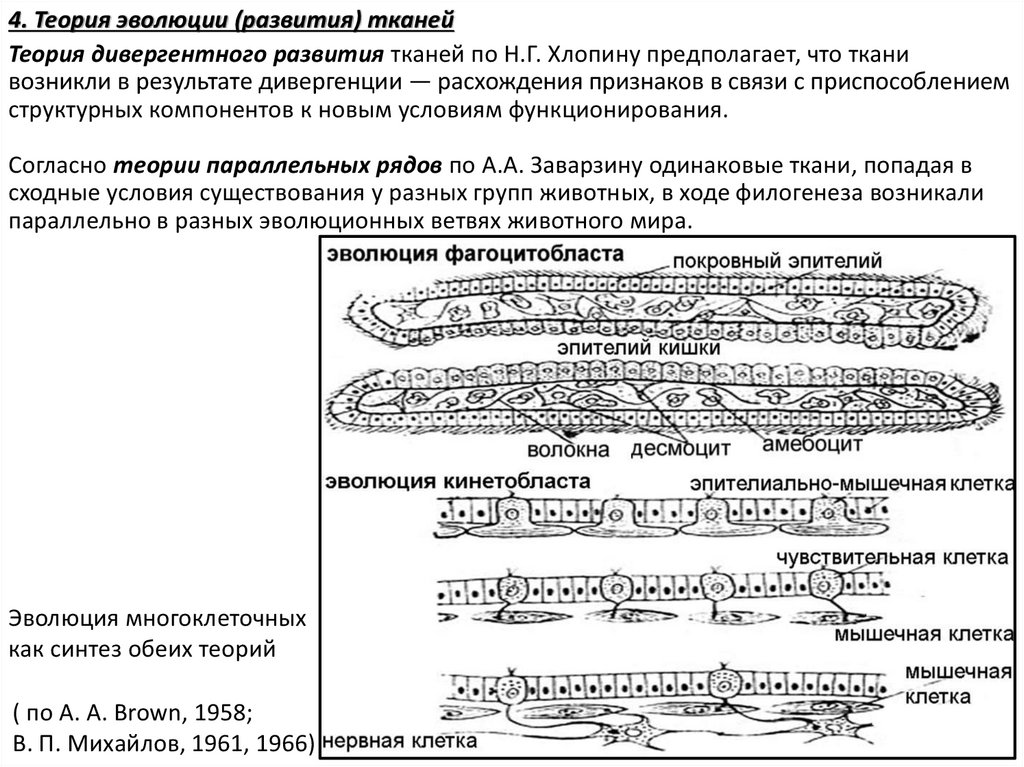
4. Теория эволюции (развития) тканейТеория дивергентного развития тканей по Н.Г. Хлопину предполагает, что ткани
возникли в результате дивергенции — расхождения признаков в связи с приспособлением
структурных компонентов к новым условиям функционирования.
Согласно теории параллельных рядов по А.А. Заварзину одинаковые ткани, попадая в
сходные условия существования у разных групп животных, в ходе филогенеза возникали
параллельно в разных эволюционных ветвях животного мира.
Эволюция многоклеточных
как синтез обеих теорий
( по А. А. Brown, 1958;
В. П. Михайлов, 1961, 1966)
9.
5. Теория регенерации тканей.Регенерация тканей – процесс, обеспечивающий её обновление в ходе нормальной
жизнедеятельности (физиологическая регенерация) или восстановление после
повреждения (репаративная регенерация).
Регенерация может происходить на уровне клетки или на внутриклеточном уровне.
Клеточные популяции – это совокупность клеток организма или ткани, сходных между
собой по какому-либо признаку.
По способности к самообновлению путём деления клеток
выделяют 4 категории клеточных популяций (по Леблону):
- Эмбриональная.
Клетки активно делятся, специализированные отсутствуют.
- Стабильная.
Клетки долгоживущие, активно функционирующие, вследствие
крайней специализации утратили способность к делению.
Например, нейроны, кардиомиоциты.
- Растущая (лабильная). Клетки специализированные,
Способны делиться в определённых условиях.
Например, эпителии почки, печени.
- Обновляющаяся.
Клетки постоянно и быстро делятся, вблизи расположены
специализированные и функционирующие потомки,
продолжительность жизни которых ограничена.
Например, клетки эпителия кожи, кишок, кроветворные клетки.
10.
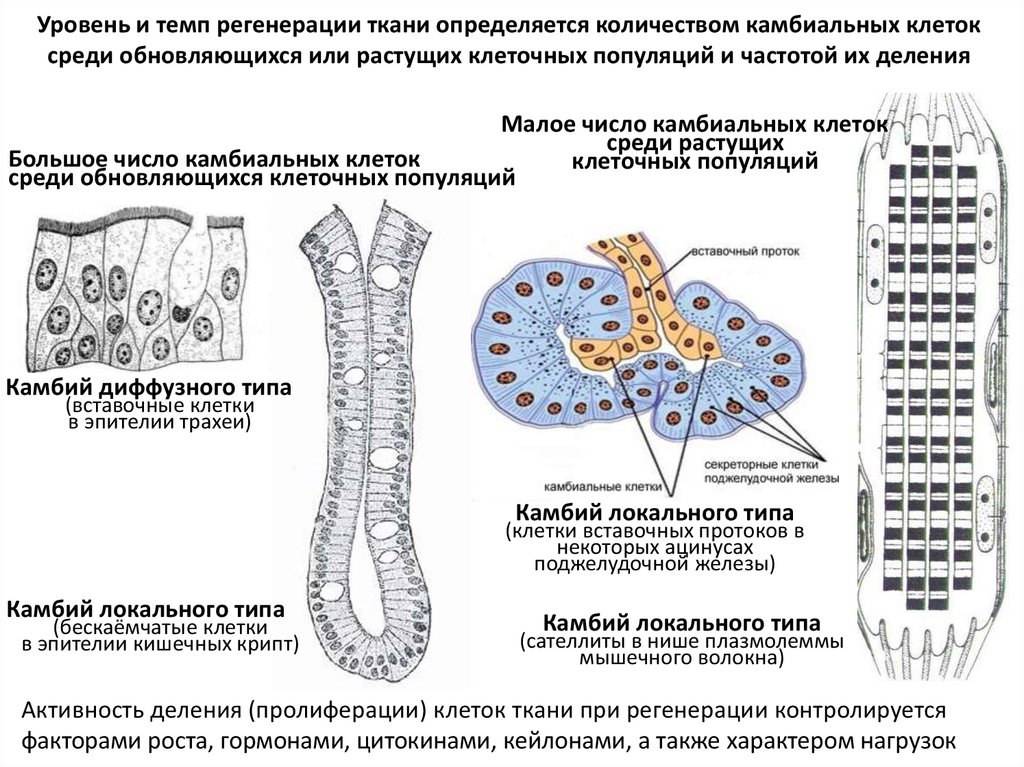
Уровень и темп регенерации ткани определяется количеством камбиальных клетоксреди обновляющихся или растущих клеточных популяций и частотой их деления
Малое число камбиальных клеток
среди растущих
Большое число камбиальных клеток
клеточных популяций
среди обновляющихся клеточных популяций
Камбий диффузного типа
(вставочные клетки
в эпителии трахеи)
Камбий локального типа
(клетки вставочных протоков в
некоторых ацинусах
поджелудочной железы)
Камбий локального типа
(бескаёмчатые клетки
в эпителии кишечных крипт)
Камбий локального типа
(сателлиты в нише плазмолеммы
мышечного волокна)
Активность деления (пролиферации) клеток ткани при регенерации контролируется
факторами роста, гормонами, цитокинами, кейлонами, а также характером нагрузок
11.
Основной метод исследования в гистологии –световая микроскопия гистологических срезов
Схема микроскопа (середина ХХ века):
A) Окуляр, B) Объектив, C) Исследуемый
объект, D) Конденсор, E) Предметный
столик, F) Осветительное зеркало
Основные узлы микроскопа (1990-е гг.)
1 окуляр, 2 турель для объективов,
3 объектив, 4 макровинт, 5 микровинт,
6 предметный столик, 7 зеркало и/или
осветитель, 8 диафрагма и конденсор,
чаще всего в одном блоке
12.
Источник света- лампа
Источник электронов – катод
анод
конденсор
срез
конденсор
Детекторы отраженного
потока электронов
срез
Линза
окуляра
Проекционная
катушка
вакуум
Детектор электронов
и CD – камера
Изображение
на сетчатке глаза
Световая
микроскопия
Сканирующая
электронная микроскопия
13.
14.
15.
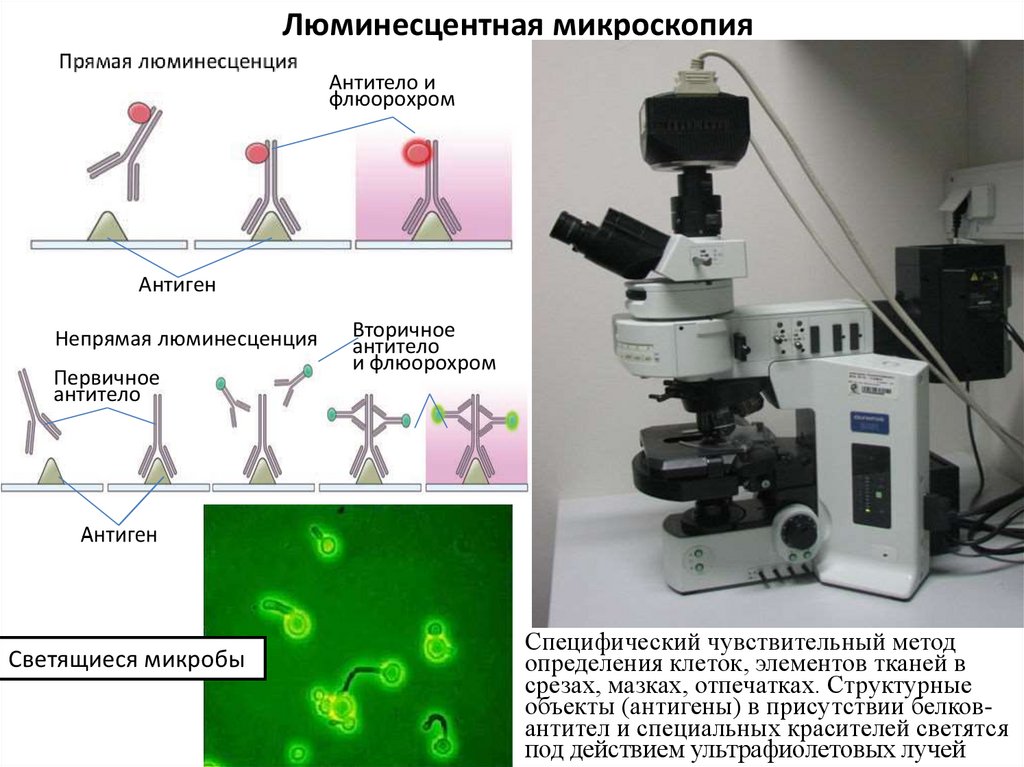
Люминесцентная микроскопияАнтитело и
флюорохром
Антиген
Непрямая люминесценция
Первичное
антитело
Светящиеся микробы
Вторичное
антитело
и флюорохром
Специфический чувствительный метод
определения клеток, элементов тканей в
срезах, мазках, отпечатках. Структурные
объекты (антигены) в присутствии белковантител и специальных красителей светятся
под действием ультрафиолетовых лучей
16.
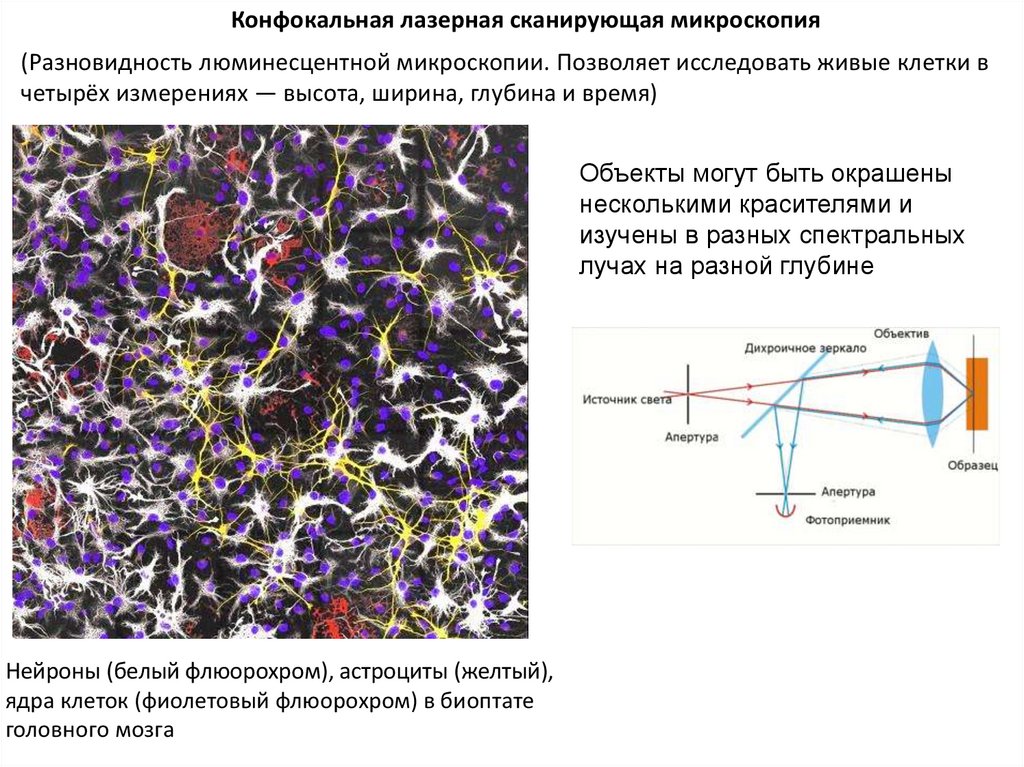
Конфокальная лазерная сканирующая микроскопия(Разновидность люминесцентной микроскопии. Позволяет исследовать живые клетки в
четырёх измерениях — высота, ширина, глубина и время)
Объекты могут быть окрашены
несколькими красителями и
изучены в разных спектральных
лучах на разной глубине
Нейроны (белый флюорохром), астроциты (желтый),
ядра клеток (фиолетовый флюорохром) в биоптате
головного мозга
17.
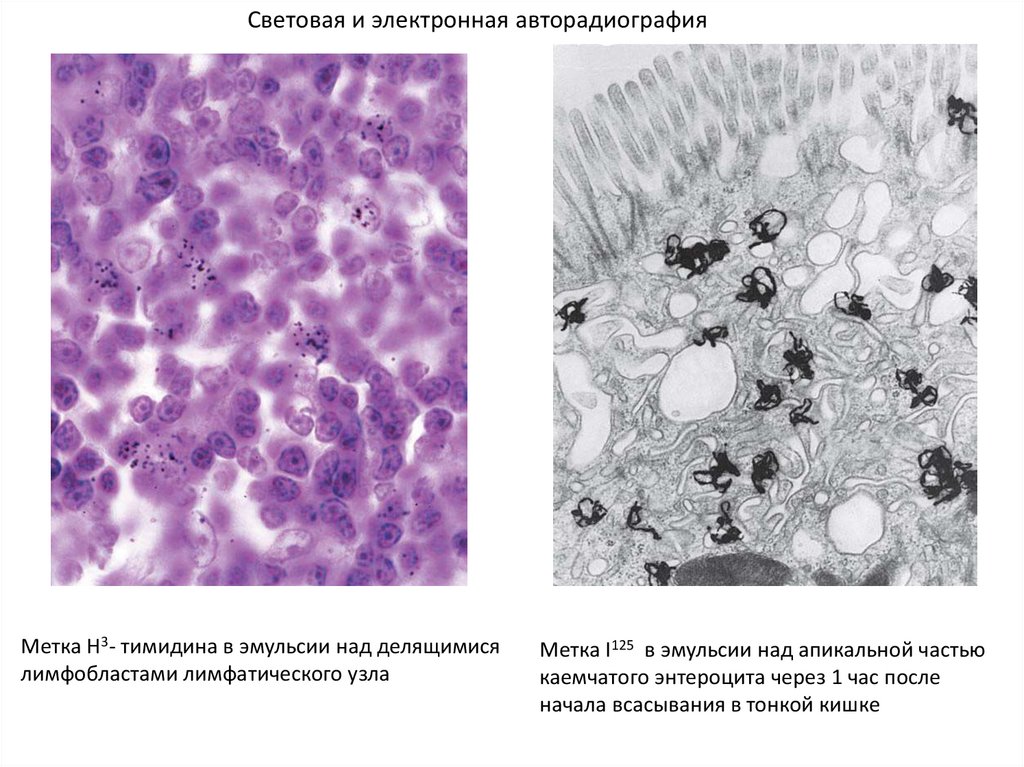
Световая и электронная авторадиографияМетка Н3- тимидина в эмульсии над делящимися
лимфобластами лимфатического узла
Метка I125 в эмульсии над апикальной частью
каемчатого энтероцита через 1 час после
начала всасывания в тонкой кишке
18.
ли19.
Основные этапы приготовления гистологических препаратов:1. взятие материала; 2. фиксация; 3. промывка в воде; 4. обезвоживание и уплотнение;
5. заливка; 6. приготовление срезов; 7.окрашивание; 8. заключение срезов.
20.
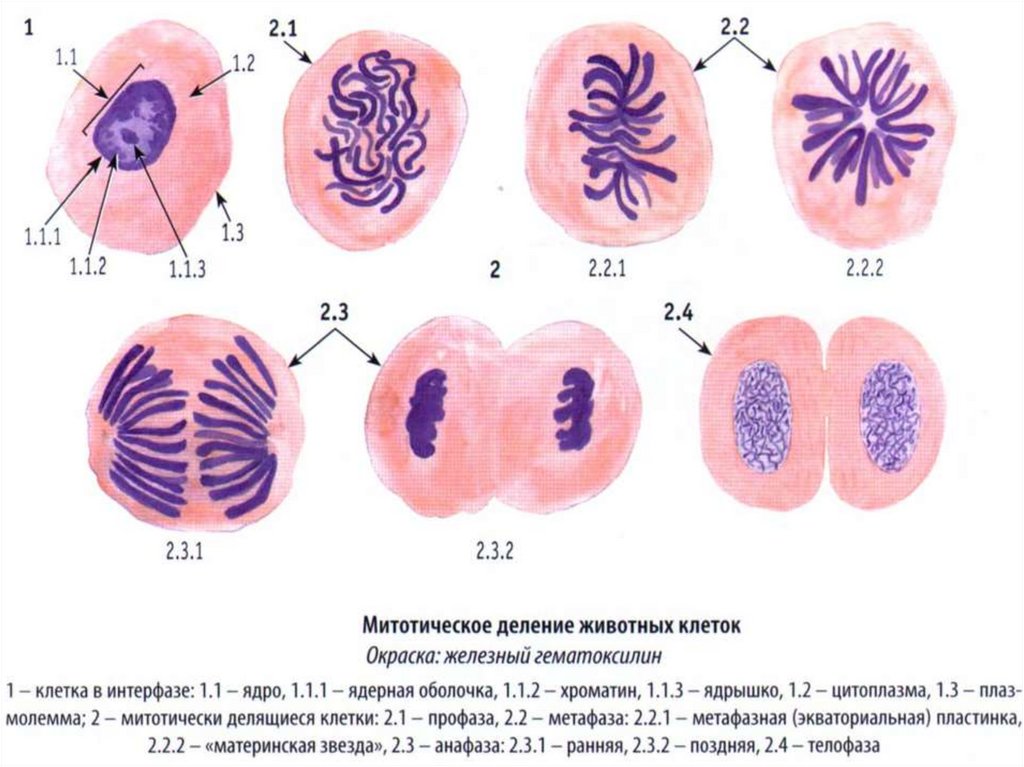
Гематоксилин и эозин – обзорные гистологические красители, взаимодополняющиетинкториальные свойства друг друга
1
3
Срезы поджелудочной железы, окрашенные: а – только гематоксилином, b – только эозином,
c – гематоксилином и эозином
Клетки срезов, окрашенные разными способами:
1 – неокрашенный срез,
2 – срез, окрашенный гематоксилином,
2.1 – ядро, 2.2 – 2итоплазма,
3 – срез, окрашенный эозином,
3.1 – ядро, 3.2 – цитоплазма,
4 – срез, окрашенный гематоксилином и эозином,
4.1 – ядро, 4.2 – цитоплазма
21.
Для чего нужна микроскопия серийных срезов?Варианты срезов почечного тельца:
размер и внутреннее строение тельца зависит от плоскости среза
22.
23.
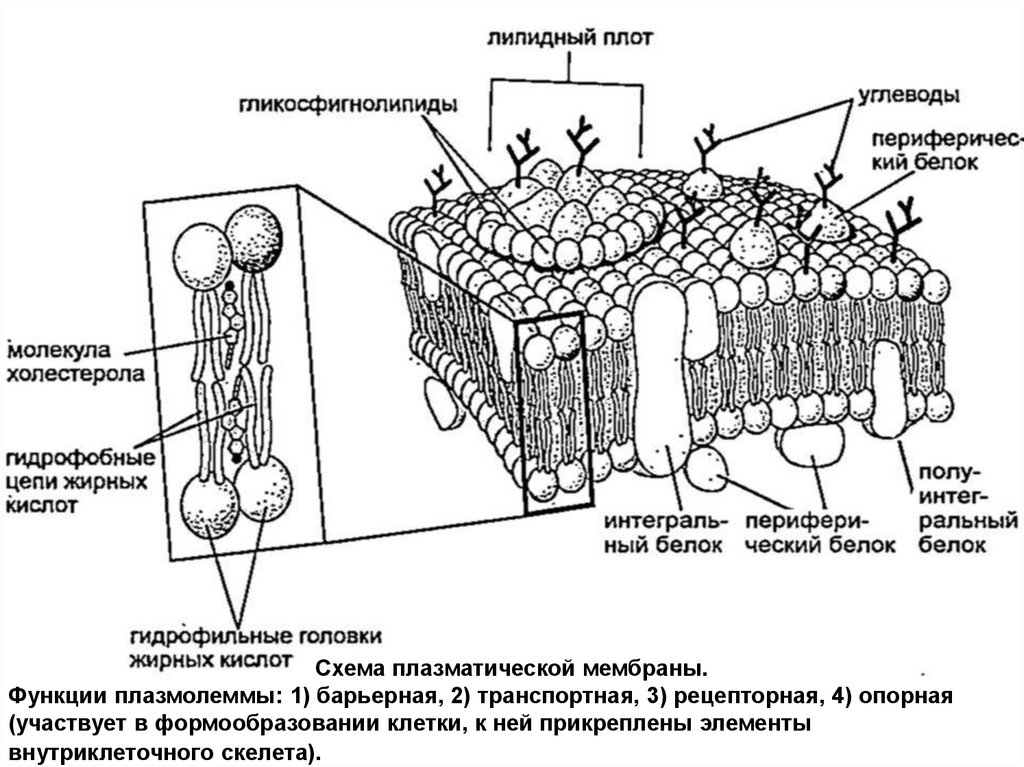
Схема плазматической мембраны.Функции плазмолеммы: 1) барьерная, 2) транспортная, 3) рецепторная, 4) опорная
(участвует в формообразовании клетки, к ней прикреплены элементы
внутриклеточного скелета).
24.
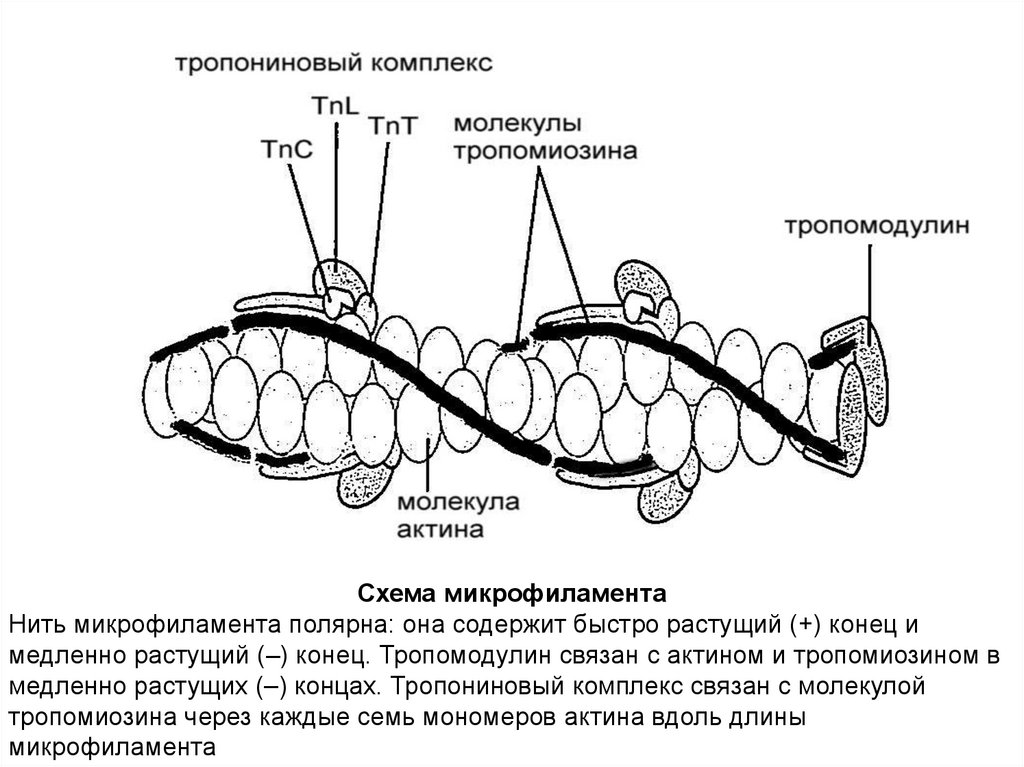
Схема микрофиламентаНить микрофиламента полярна: она содержит быстро растущий (+) конец и
медленно растущий (–) конец. Тропомодулин связан с актином и тропомиозином в
медленно растущих (–) концах. Тропониновый комплекс связан с молекулой
тропомиозина через каждые семь мономеров актина вдоль длины
микрофиламента
25.
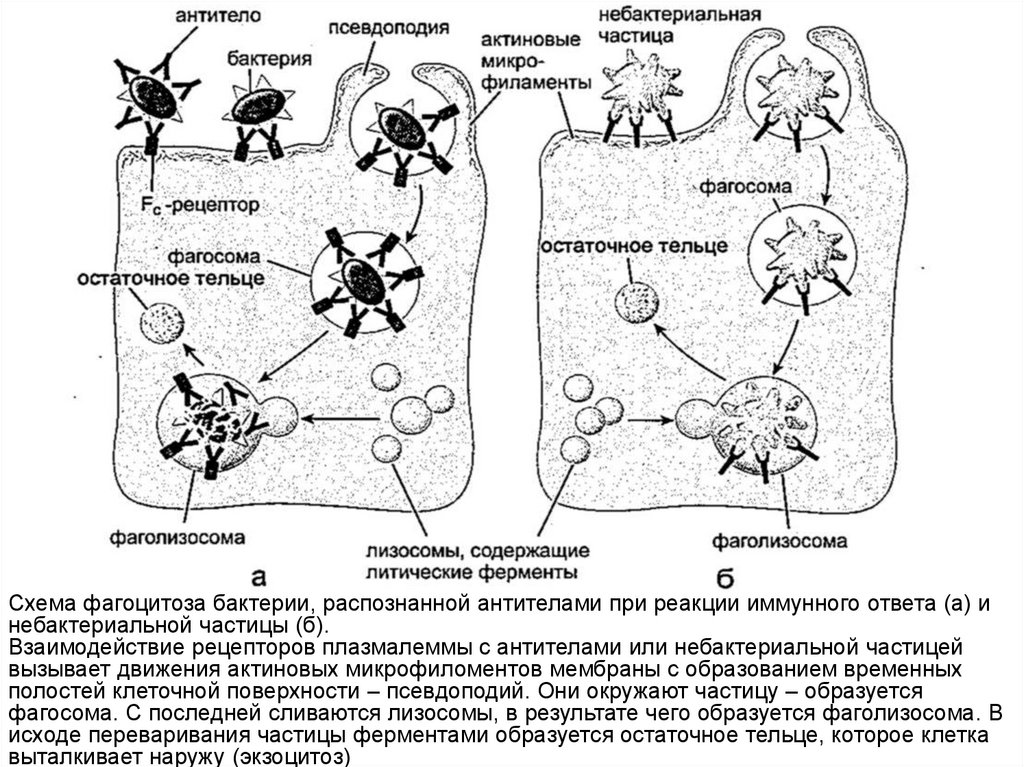
Схема фагоцитоза бактерии, распознанной антителами при реакции иммунного ответа (а) инебактериальной частицы (б).
Взаимодействие рецепторов плазмалеммы с антителами или небактериальной частицей
вызывает движения актиновых микрофиломентов мембраны с образованием временных
полостей клеточной поверхности – псевдоподий. Они окружают частицу – образуется
фагосома. С последней сливаются лизосомы, в результате чего образуется фаголизосома. В
исходе переваривания частицы ферментами образуется остаточное тельце, которое клетка
выталкивает наружу (экзоцитоз)
26.
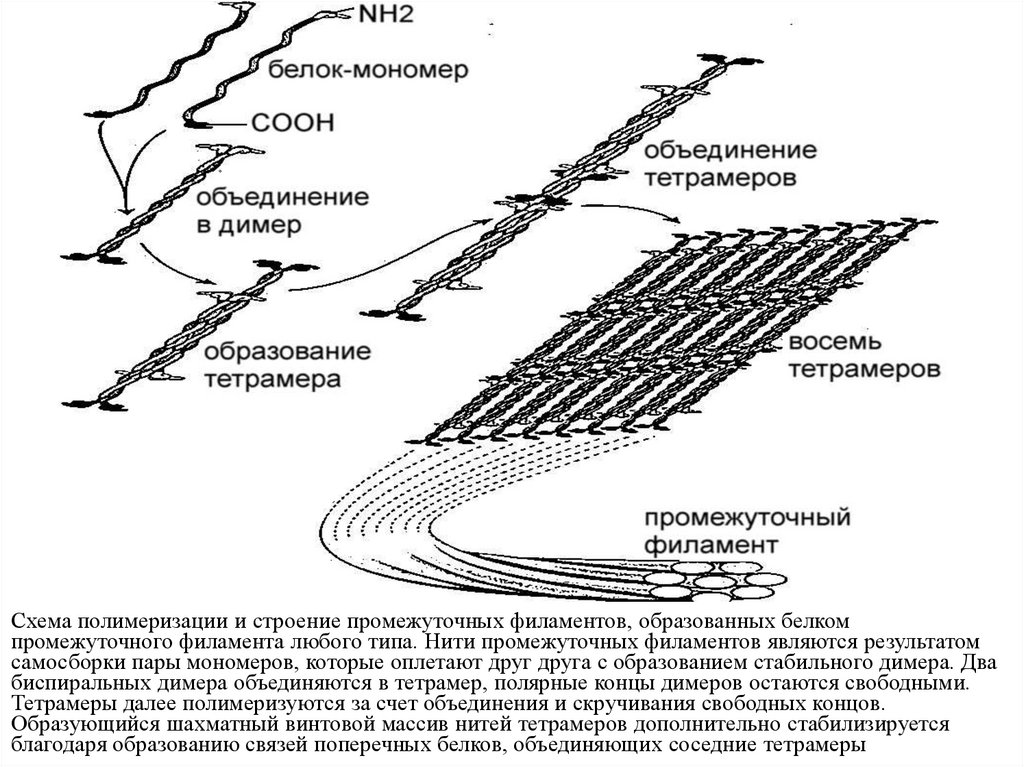
Схема полимеризации и строение промежуточных филаментов, образованных белкомпромежуточного филамента любого типа. Нити промежуточных филаментов являются результатом
самосборки пары мономеров, которые оплетают друг друга с образованием стабильного димера. Два
биспиральных димера объединяются в тетрамер, полярные концы димеров остаются свободными.
Тетрамеры далее полимеризуются за счет объединения и скручивания свободных концов.
Образующийся шахматный винтовой массив нитей тетрамеров дополнительно стабилизируется
благодаря образованию связей поперечных белков, объединяющих соседние тетрамеры
27.
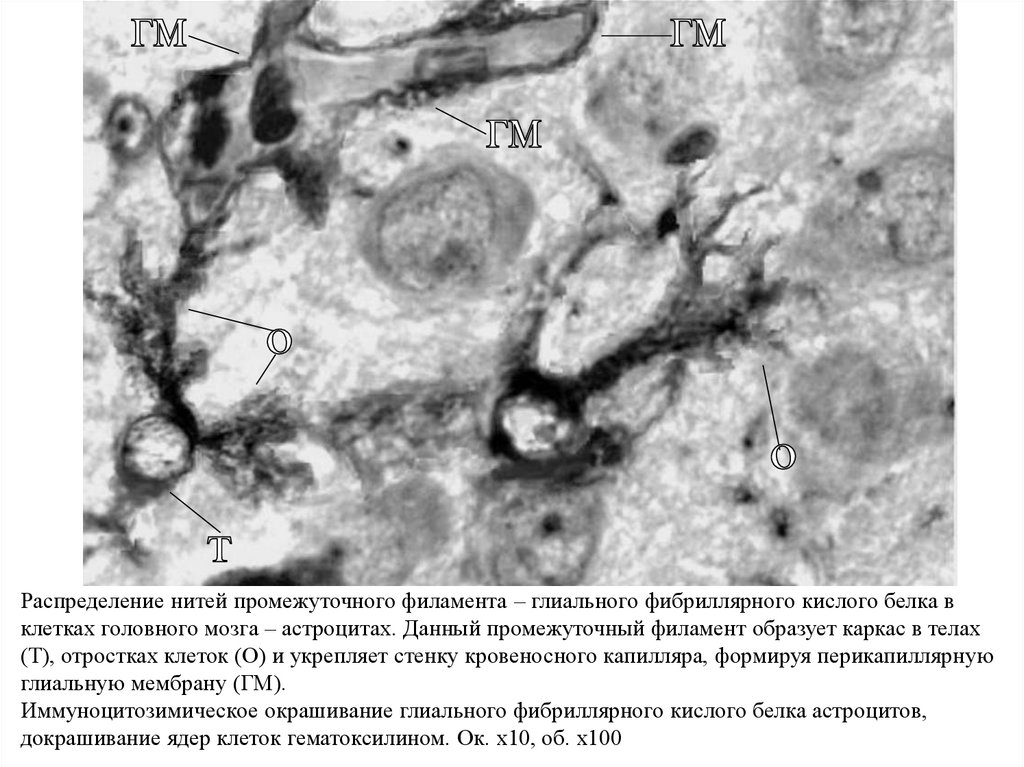
Распределение нитей промежуточного филамента – глиального фибриллярного кислого белка вклетках головного мозга – астроцитах. Данный промежуточный филамент образует каркас в телах
(Т), отростках клеток (О) и укрепляет стенку кровеносного капилляра, формируя перикапиллярную
глиальную мембрану (ГМ).
Иммуноцитозимическое окрашивание глиального фибриллярного кислого белка астроцитов,
докрашивание ядер клеток гематоксилином. Ок. х10, об. х100
28.
Схема обеспечения молекулярного транспорта, опосредованного микротрубочами.Микротрубочки направляют белки АТФ-зависимого транспорта макромолекул и
органелл. Движения вдоль микротрубочек осуществляют белки динеины и
кинезины. Динеины движутся к их минус концу (то есть, по направлению к центру
клетки), тогда как кинезины движутся в сторону плюс конца (т.е. к периферии)
29.
Комплекс Гольджи клетки междолькового желчного протока, секретирующейкомпоненты желчи. Трансмиссионная электронная микроскопия. Ув. 10000.
Транспортные пузырьки (Т) и элементы шероховатой эндоплазматической сети (шЭПС)
видны рядом с поверхностью пластинчатого комплекса (ПК). Везикулы разной величины и
формы (В) могут являться секреторными гранулами (с электронно-светлым содержимым) или
лизосомами. ПК расположен вблизи ядра (Я) и его оболочки (ЯО)
30.
Структура митохондрии в секреторной клетке печени (гепатоците).А –трансмиссионная электронная микроскопия (собственное наблюдение).
Ув. 30000. К –кристы, образованные внутренней мембраной митохондрии.
Наружная мембрана митохондрии формирует гладкую непрерывную наружную
оболочку, отдельную от внутренней оболочки. Б – схема митохондрии. Форма
элементарных частиц (вставка) отражает трехмерную структуру АТФ-синтетазы
31.
Схема строения реснички и базального тельца. А – продольный срез;Б – поперечный срез тела реснички; В, Г – срезы базального тельца.
1 – плазматическая мембрана,
2 – микротрубочки, 3 – дуплеты микротрубочек (а, в);
4 – триплеты микротрубочек (а, в, с)
32.
Рисунок тушью, выполнен с фотографии ядер двух различных типов клеток – нейрона (А) илимфоцита (Б), полученной с помощью трансмиссионной электронной микроскопии
(из кн.: Ross M., Wojciech P., 2011). х10.000
Функции ядра:
-хранение генетической информации (в молекулах ДНК, находящихся в хромосомах)
-реализация генетической информации, контролирующей осуществление разнообразных процессов в
клетке - от синтетических до запрограммированной гибели (апоптоз)
- воспроизведение и передачу генетической информации (при делении клетки)
33.
34.
35.
36.
Программированная клеточная гибель – апоптозФизиологическая и
патологическая
потеря клеток
Годом признания апоптоза как
физиологического явления считается
1972 год, когда английские
исследователи во главе с Дж. Керром
представили убедительные
морфологические доказательства
существования этого явления
Увеличение
числа клеток
В 2002 году Бреннер, Салстон и Хорвитц
получили Нобелевскую премию по
физиологии и медицине за исследования
генетической регуляции апоптоза.
Они открыли особые белки,
регулирующие этот процесс
(«генетическое регулирование развития
организма и запрограммированная смерть
клеток» - nobelprize.org)
37.
Морфогенетическийэмбриональный апоптоз
Апоптоз при удалении некоторых клеток
пролиферирующих клеточных популяций
Фрагмент стенки кишечной ворсинки.
АВ – апоптотические тельца
ВМ – базальная мембрана
Лапа эмбриона мыши (15-й день).
Между пальцами видны скопления
апоптотически гибнущих клеток
Wikimedia, the free media repository;
Michal Maňas
(Из кн.: Ross M., Wojciech P., 2011).
38.
Рецептор-зависимый сигнальный путьСхема передачи
сигналов апоптоза
при посредстве
рецепторов смерти
CD95
Эффекторная фаза –
каскад каспазных реакций
и образование апоптосомы
Индукционные каспазы: 9, 8, 10 активируют
эффекторные: 3, 6, 7.
Последние 1) осуществляют гидролиз белков,
2) инактивируют белки, блокирующие апоптоз,
3) разрушают ферменты репарации и репликации ДНК, образование иРНК
(Самуилов В. Д., Олескин А. В., Лагунова Е. М.;
МГУ им. М.В. Ломоносова)
39.
40.
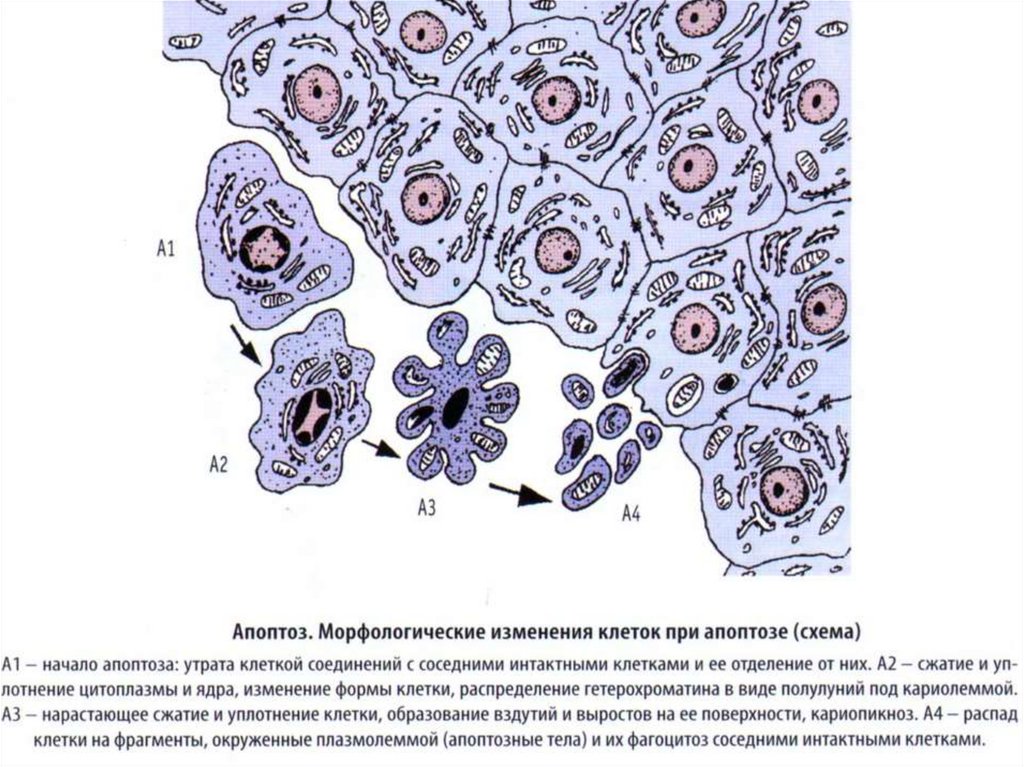
Морфологическая схема апоптоза (1 – 4)и некроза (5 – 6)
НИИ Эпидемиологии и Микробиологии им.
академика И.Н.Блохиной МЗ РФ
(Источник: http://immunelab. boom.ru/apoptos.html,
2006)
1 – нормальная клетка; 2 – начало апоптоза; 3 – фрагментация апоптотической клетки;
4 – фагоцитоз апоптотических телец окружающими клетками;
5 – гибель внутриклеточных структур при некрозе; 6 – разрушение клеточной мембраны