




















")

























 Химия
ХимияПохожие презентации:










Депонирование и мобилизация жиров
1. Депонирование и мобилизация жиров
2. Обмен триацилглицеринов
С пищей в сутки поступает 70 г ТГ.Эндогенный синтез ТГ идёт в:
• печени,
• жировой ткани,
• стенке кишечника.
В плазме крови содержится 1-2,3 ммоль/л ТГ.
ТГ – резервное топливо, которое накапливается
в цитоплазме жировых клеток.
В состав мембран ТГ не входят.
3. Функции резервных жиров
• резервуар энергии,• теплоизоляционная,
• защита от механических травм.
4. Транспорт ТГ осуществляют
• ХМ,• ЛПОНП.
5. Липопротеидлипаза
• осуществляет гидролиз ТГ.В жировой ткани жир накапливается за счёт
• поступления из ЛП,
• образования из глюкозы в жировых клетках.
В жировых депо гидролиз осуществляют
тканевые липазы.
6. Тканевые липазы активируются
• адреналином,• глюкагоном,
• АКТГ.
7. Мобилизация депонированных жиров
• происходит путём их гидролиза до жирныхкислот и глицерина липазами жировых клеток.
• Жирные кислоты поступают в кровь и
транспортируются в соединении с альбумином
к разным органам.
• Глицерин поступает в кровь и там
превращается в глицерофосфат, который
используется в гликолизе или глюконеогенезе.
8. Окисление глицерина
CH2OHCH2OH
CHOH
CH2OH
Глицерин
АТФ
Mg
2+ АДФ
Глицеролкиназа
CHOH
CH2OPO3H2
Глицерол-3-фосфат
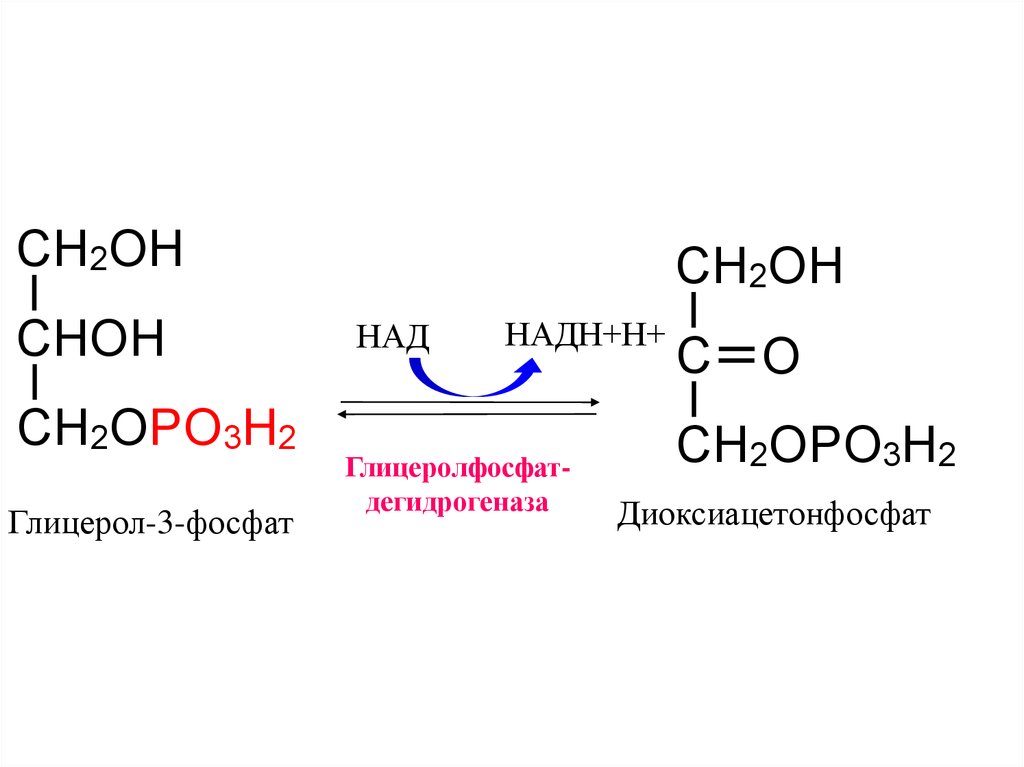
9.
CH2OHCHOH
CH2OPO3H2
Глицерол-3-фосфат
CH2OH
НАД
НАДН+Н+
Глицеролфосфатдегидрогеназа
C O
CH2OPO3H2
Диоксиацетонфосфат
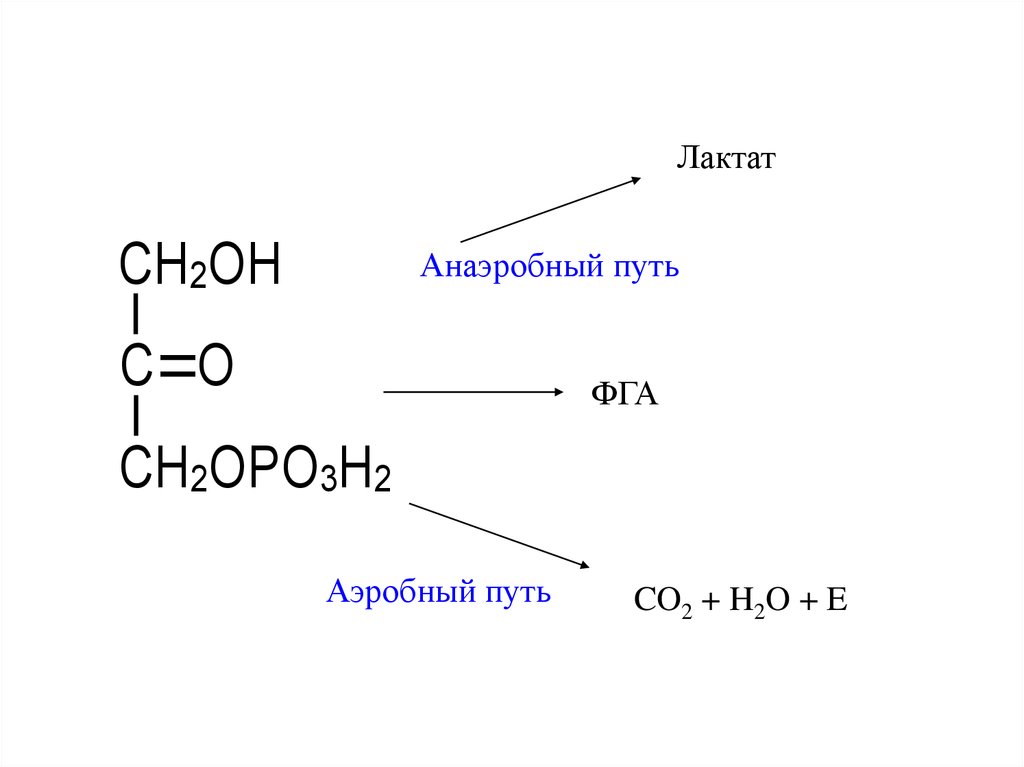
10.
ЛактатCH2OH
Анаэробный путь
C O
ФГА
CH2OPO3H2
Аэробный путь
CO2 + H2O + E
11.
CH2OHC O
ТриозофосфатCH2OPO3H2 изомераза
Диоксиацетон
O
C
H
CHOH
CH2OPO3H2
Фосфоглицериновый
альдегид
12.
OC
H
CHOH
O
+ НАД+ + H3PO4
CH2OPO3H2
Фосфоглицериновый
альдегид
C
O PO3H2
CHOH
+ НАДН+Н+
Глицеральдегидфосфат
дегидрогеназа
CH2OPO3H2
1,3-дифосфоглицерат
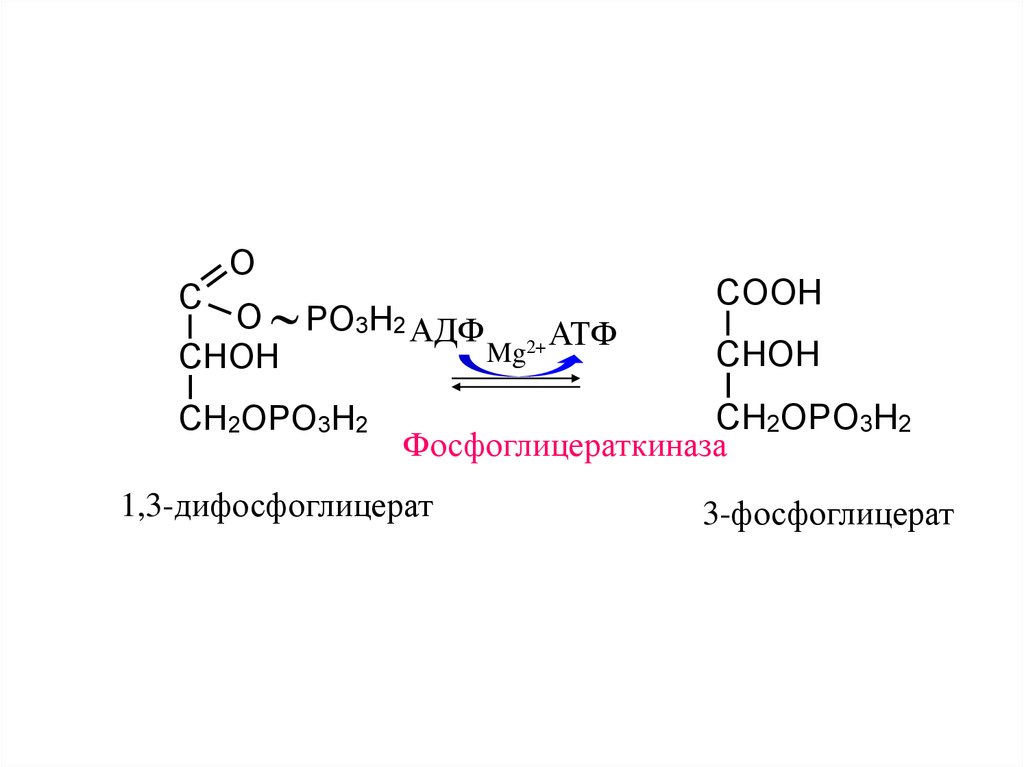
13.
OC
O PO3H2 АДФ
2+ АТФ
Mg
CHOH
CH2OPO3H2
COOH
CHOH
CH2OPO3H2
Фосфоглицераткиназа
1,3-дифосфоглицерат
3-фосфоглицерат
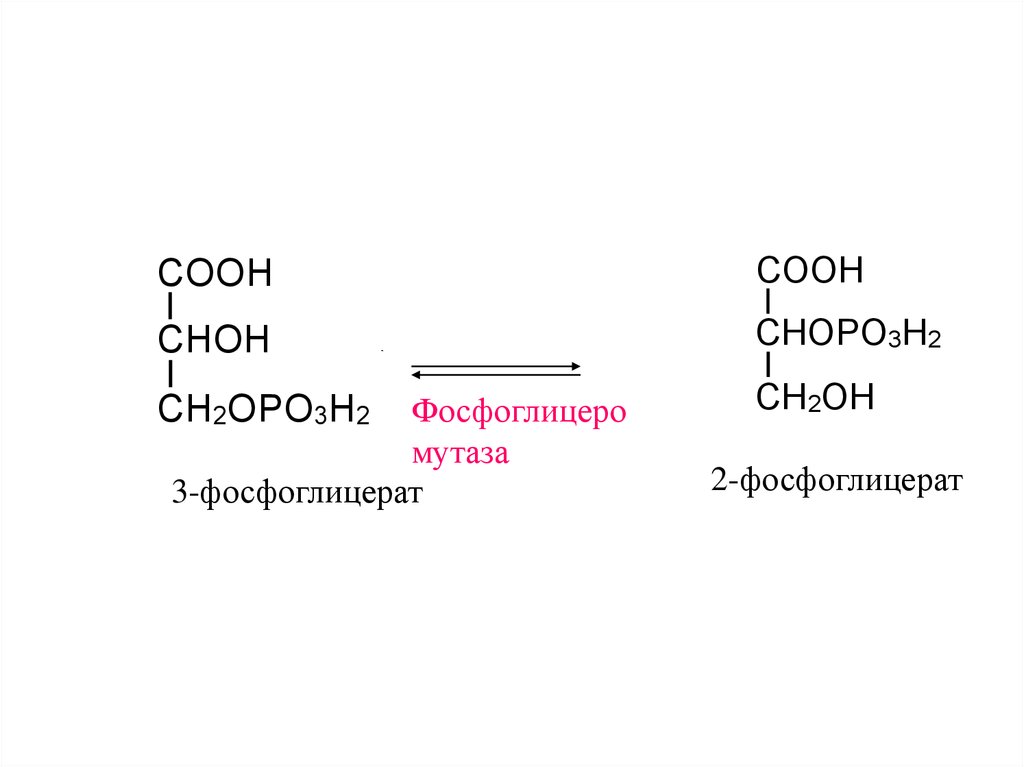
14.
COOHCOOH
CHOH
CHOPO3H2
Фосфоглицеро
мутаза
3-фосфоглицерат
CH2OPO3H2
CH2OH
2-фосфоглицерат
15.
COOHCHOPO3H2
CH2OH
2-фосфоглицерат
COOH
Mg2+ Н2О CO
Енолаза
PO3H2
CH2
Фосфоенолпируват
16.
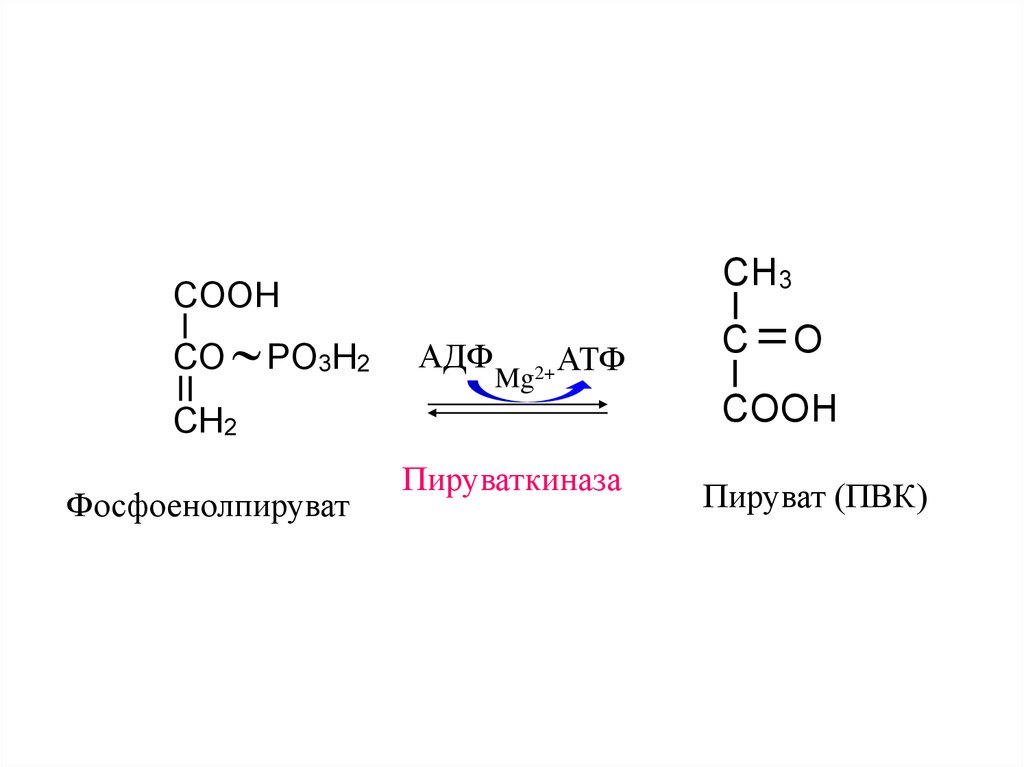
CH3COOH
CO
PO3H2
АДФ
Mg
2+ АТФ
CH2
Фосфоенолпируват
Пируваткиназа
C
O
COOH
Пируват (ПВК)
17.
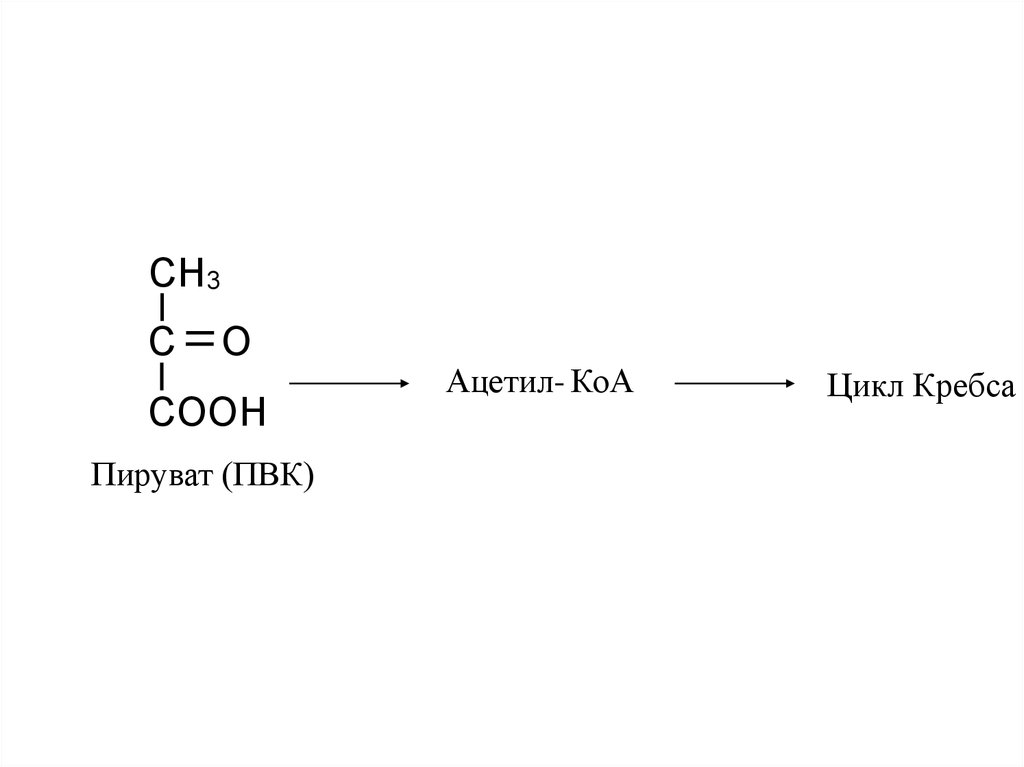
CH3C
O
COOH
Пируват (ПВК)
Ацетил- КоА
Цикл Кребса
18.
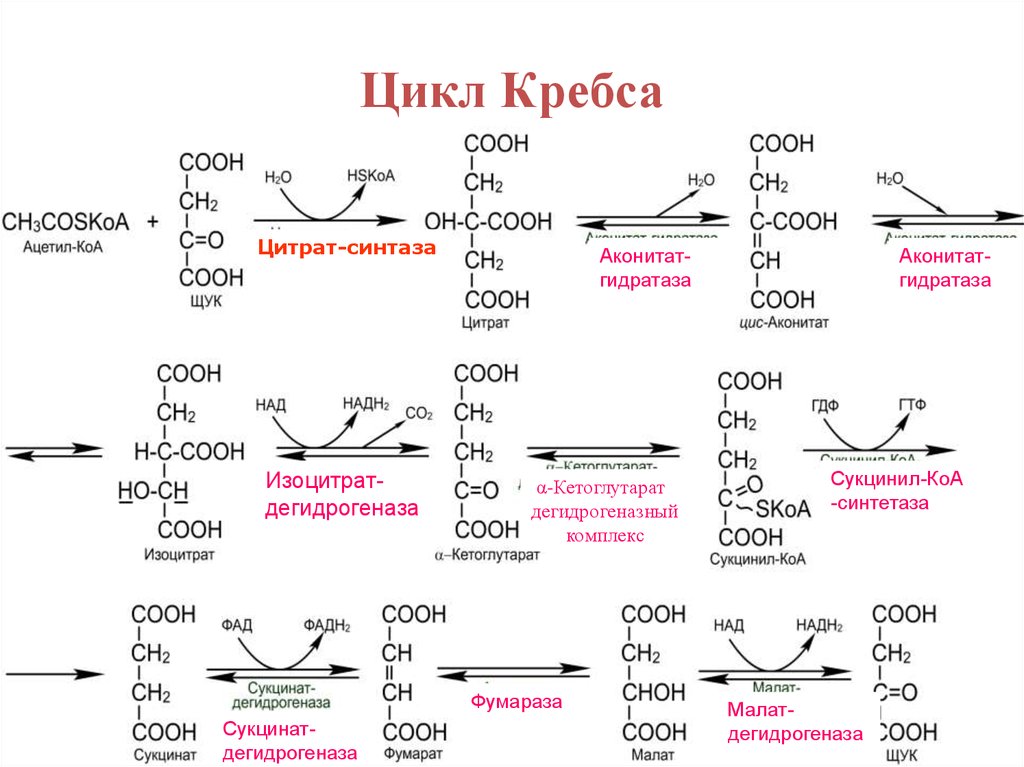
Цикл КребсаЦитрат-синтаза
Изоцитратдегидрогеназа
Аконитатгидратаза
α-Кетоглутарат
дегидрогеназный
комплекс
Фумараза
Сукцинатдегидрогеназа
Аконитатгидратаза
Сукцинил-КоА
-синтетаза
Малатдегидрогеназа
19.
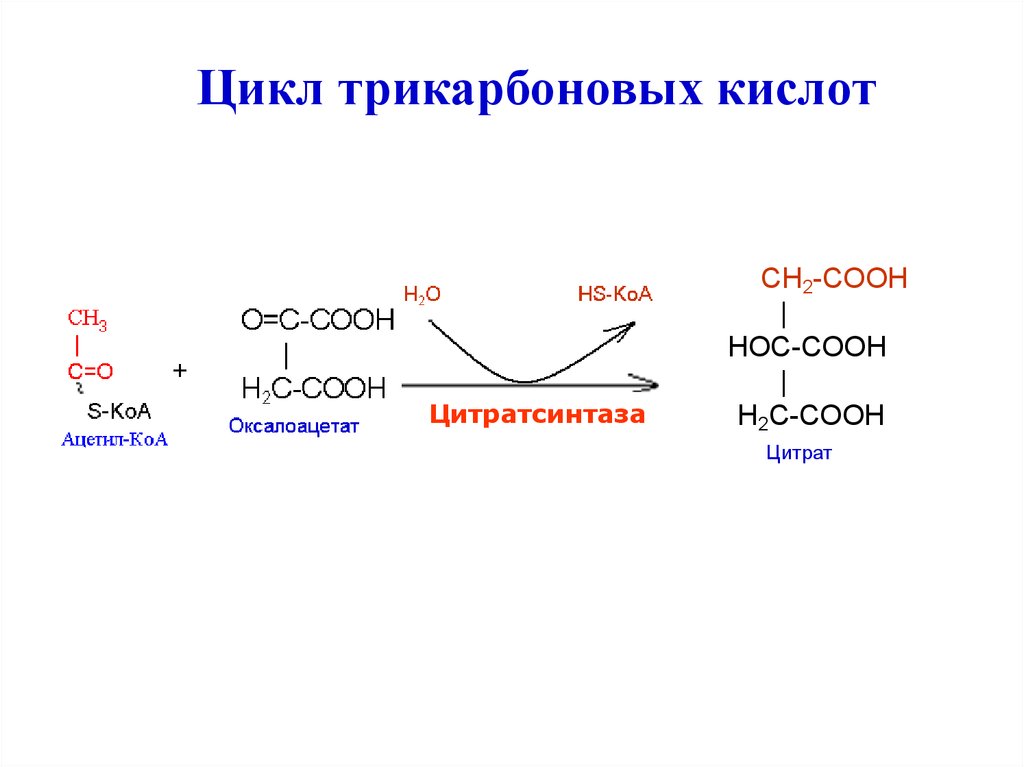
Цикл трикарбоновых кислотЦитратсинтаза
CH2-COOH
|
HOC-COOH
|
H2C-COOH
Цитрат
20.
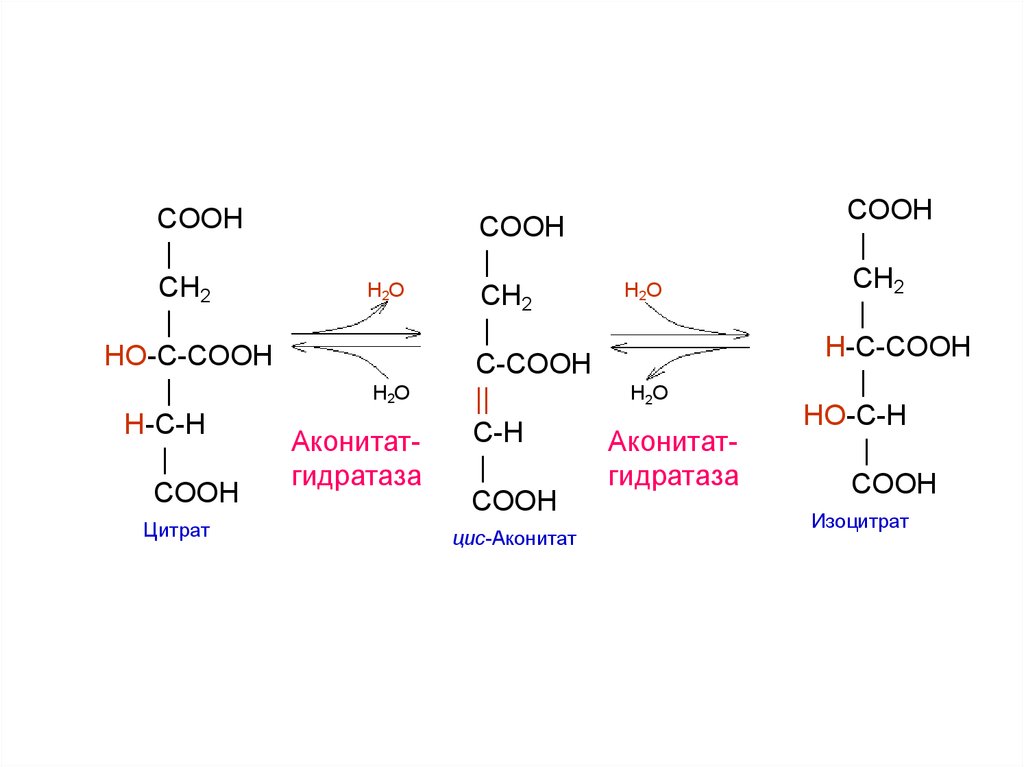
COOH|
CH2
H2O
|
HO-C-COOH
|
H2O
H-C-H
Аконитат|
гидратаза
COOH
Цитрат
COOH
|
H2O
CH2
|
C-COOH
H2O
||
C-H
Аконитат|
гидратаза
COOH
цис-Аконитат
COOH
|
CH2
|
H-C-COOH
|
HO-C-H
|
COOH
Изоцитрат
21.
COOH|
CH2
|
H-C-COOH
|
HO-C-H
|
COOH
Изоцитрат
НАД+
НАДН+Н+
Изоцитратдегидрогеназа
СО2
COOH
|
CH2
|
CH2
|
C=O
|
COOH
L-кетоглутарат
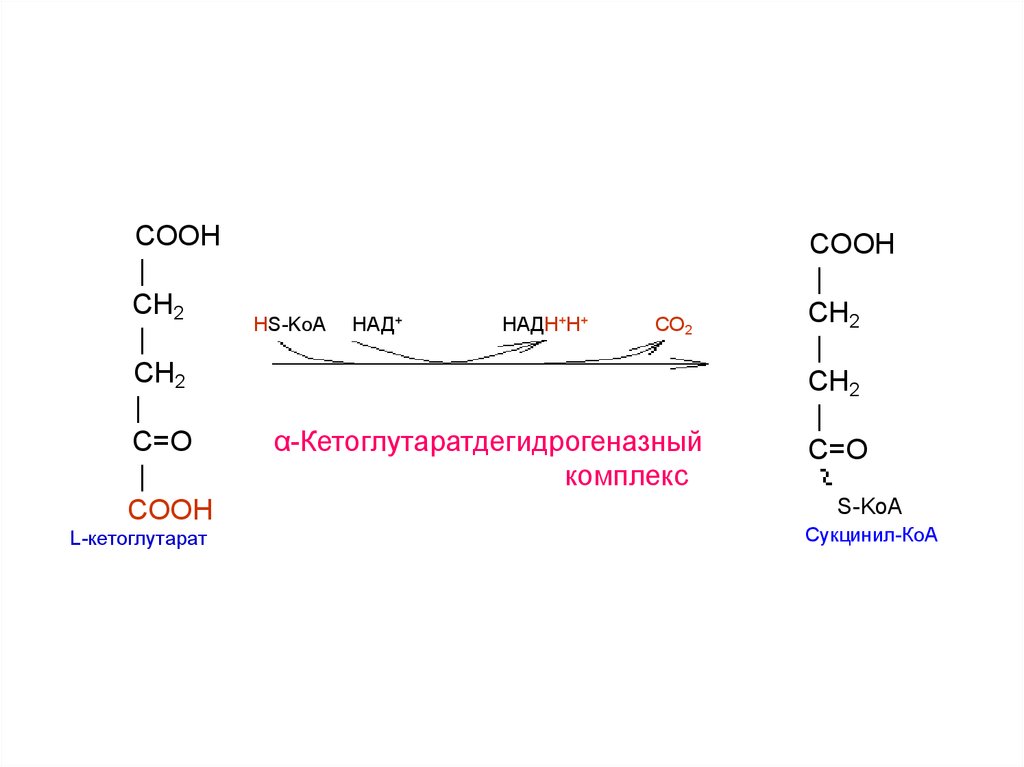
22.
COOH|
CH2
|
CH2
|
C=O
|
COOH
L-кетоглутарат
HS-KoA
НАД+
НАДН+Н+
СО2
α-Кетоглутаратдегидрогеназный
комплекс
COOH
|
CH2
|
СН2
|
C=O
S-KoA
Cукцинил-КоА
23.
COOH|
CH2
|
ГДФ
ГТФ
СН2
+ Фн
|
C=O
Сукцинил-КоА-синтетаза
COOH
|
CH2
|
СН2
|
COOH
S-KoA
Cукцинил-КоА
Сукцинат
+ HS-KoA
24.
COOH|
CH2
|
СН2
|
COOH
Сукцинат
ФАД
ФАДН2
Сукцинатдегидрогеназа
COOH
|
CH
||
HС
|
COOH
Фумарат
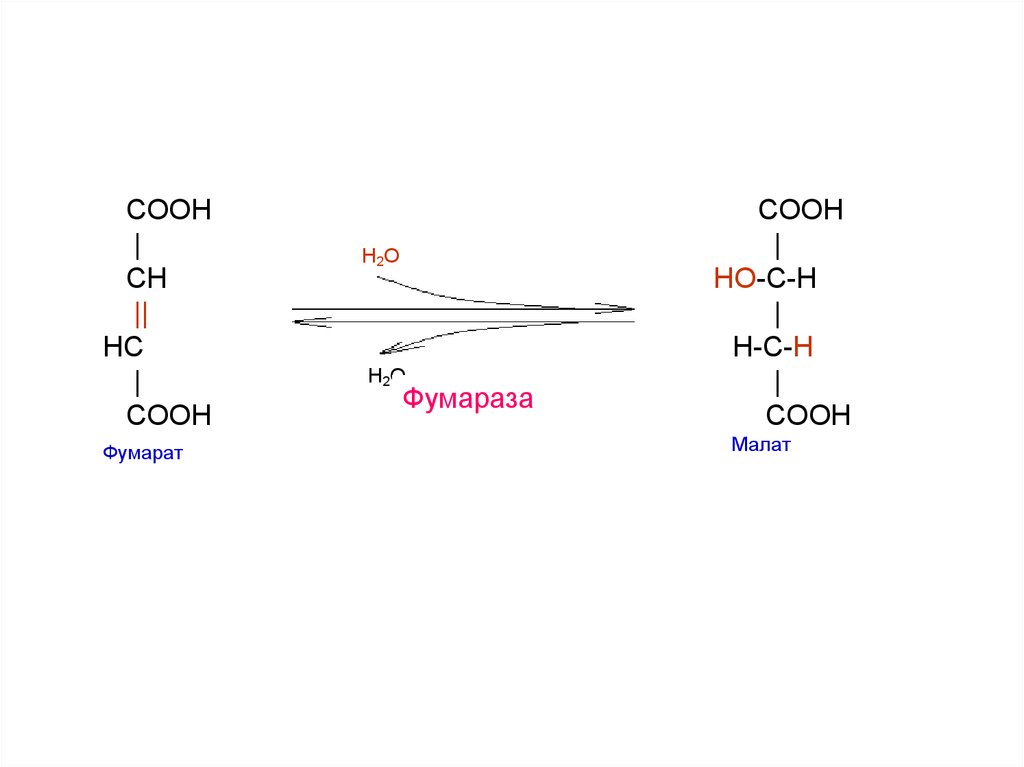
25.
COOH|
CH
||
HС
|
COOH
Фумарат
H2O
H2O
Фумараза
COOH
|
HO-C-H
|
H-С-H
|
COOH
Малат
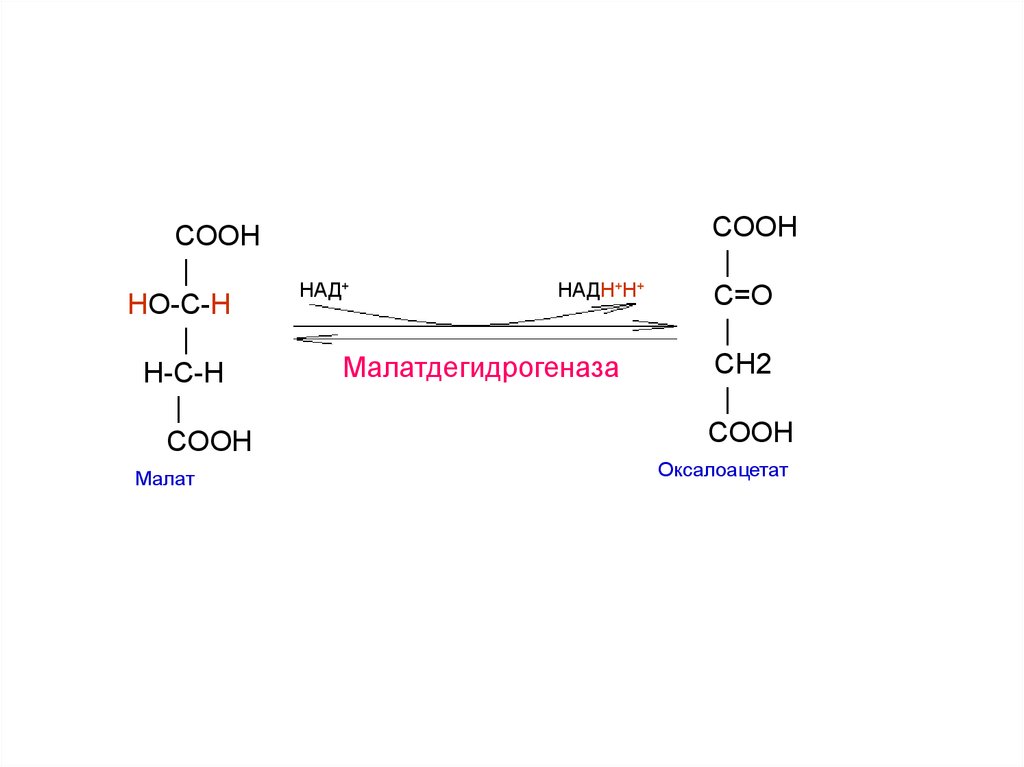
26.
COOH|
HO-C-H
|
H-С-H
|
COOH
Малат
НАД+
НАДН+Н+
Малатдегидрогеназа
COOH
|
C=O
|
СH2
|
COOH
Оксалоацетат
27. Баланс аэробного распада глицерина
• От глицерина до ФГАзатрата
– 1 АТФ
и получение + 3 АТФ (окислительное фосфорилирование).
• На втором этапе гликолиза при окислении
+
1 молекулы ФГА получаем 2 АТФ и 1 НАДН+Н,
то есть 2+3=5 АТФ.
Таким образом от глицерина до ПВК получаем
(3-1) + 5 = 7АТФ.
• Окислительное декарбоксилирование ПВК даёт 3АТФ.
• ЦТК даёт 12 АТФ.
ИТОГО: 7+3+12 = 22АТФ даёт окисление 1 молекулы
глицерина в аэробных условиях.
28.
ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ- аэробный процесс.
- В катаболизме жирных кислот выделяют 3 части:
1. β-окисление,
2. ЦТК,
3. дыхательная цепь.
Позвоночные половину энергии получают за счёт
окисления жирных кислот , особенно в спячке
и при голодании.
29.
• Кнооп установил, что окисление идёт вβ-положении.
• Ленинджер и Кеннеди установили, что
процесс протекает в митохондриях с
использованием АТФ.
• Линен, Грин, Очоа установили этапы
окисления, роль КоАSH
30. Активация жирных кислот происходит на наружной поверхности мембраны митохондрий.
R-COOH + HS-KoA +АТФR CO S-KoA + АМФ + ФФн
Ацил-КоАсинтетаза
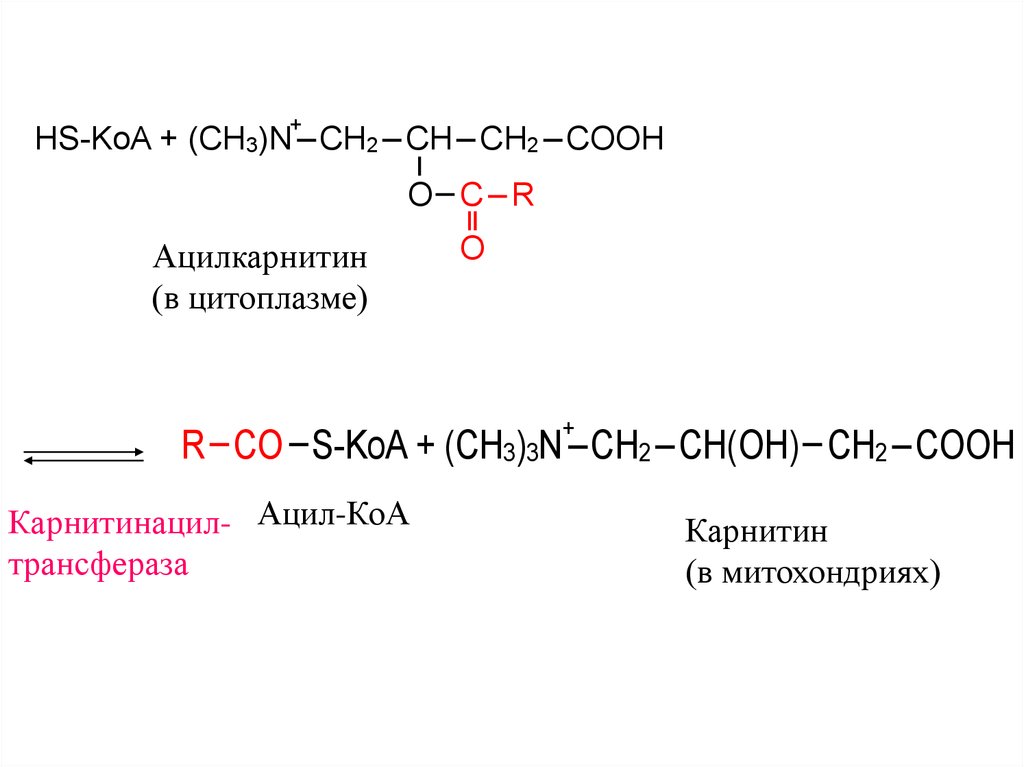
31. Транспорт жирных кислот в митохондрии из цитоплазмы осуществляет карнитин.
+R CO S-KoA + (CH3)3N CH2 CH(OH) CH2 COOH
Ацил-КоА
Ацилкарнитин
(в цитоплазме)
Карнитин
Карнитинацилтрансфераза
32.
+HS-KoA + (CH3)N CH2 CH CH2 COOH
O C R
Ацилкарнитин
(в цитоплазме)
O
+
R CO S-KoA + (CH3)3N CH2 CH(OH) CH2 COOH
Карнитинацил- Ацил-КоА
трансфераза
Карнитин
(в митохондриях)
33.
β-ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТв митохондриях включает стадии
- первая стадия дегидрирования,
- стадия гидратации,
- вторая стадия дегидрирования,
- тиолазная реакция.
34.
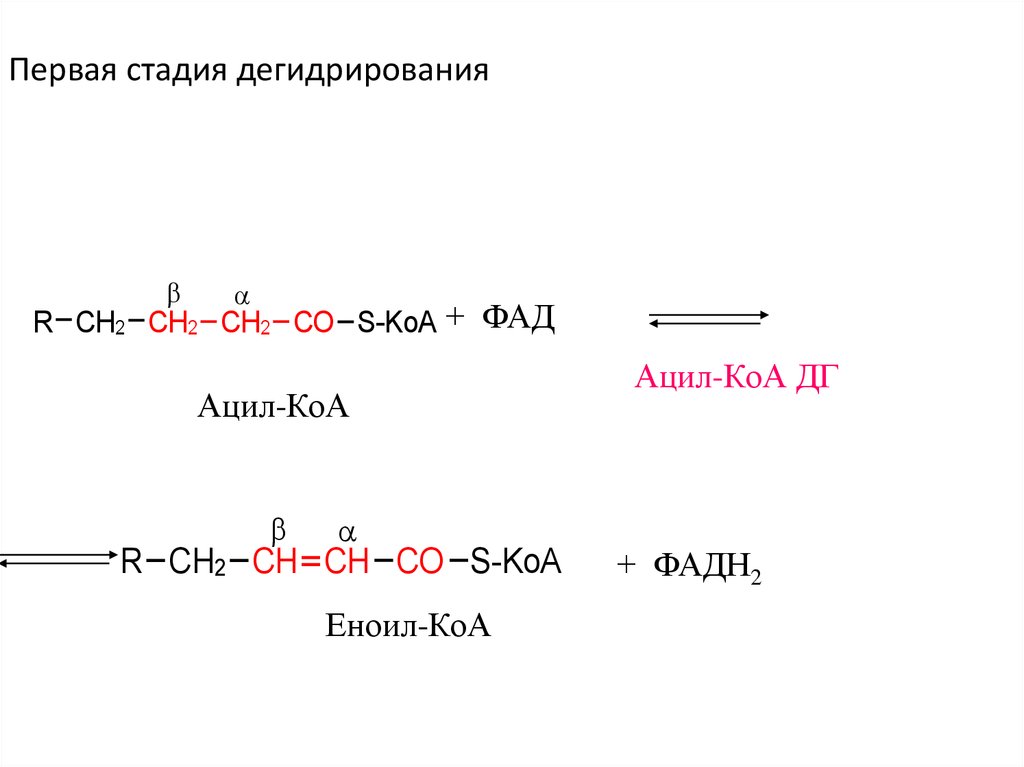
Первая стадия дегидрированияR CH2 CH2 CH2 CO S-KoA + ФАД
Ацил-КоА
R CH2 CH CH CO S-KoA
Еноил-КоА
Ацил-КоА ДГ
+ ФАДН2
35.
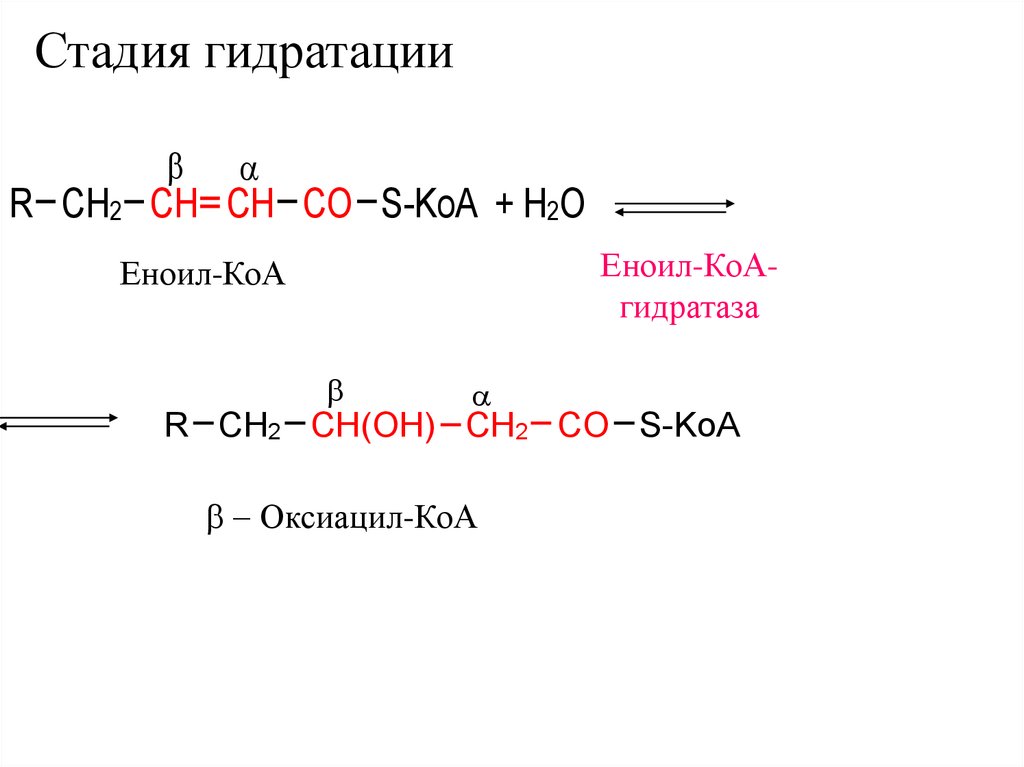
Стадия гидратацииR CH2 CH CH CO S-KoA + H2O
Еноил-КоАгидратаза
Еноил-КоА
R CH2 CH(OH) CH2 CO S-KoA
- Оксиацил-КоА
36.
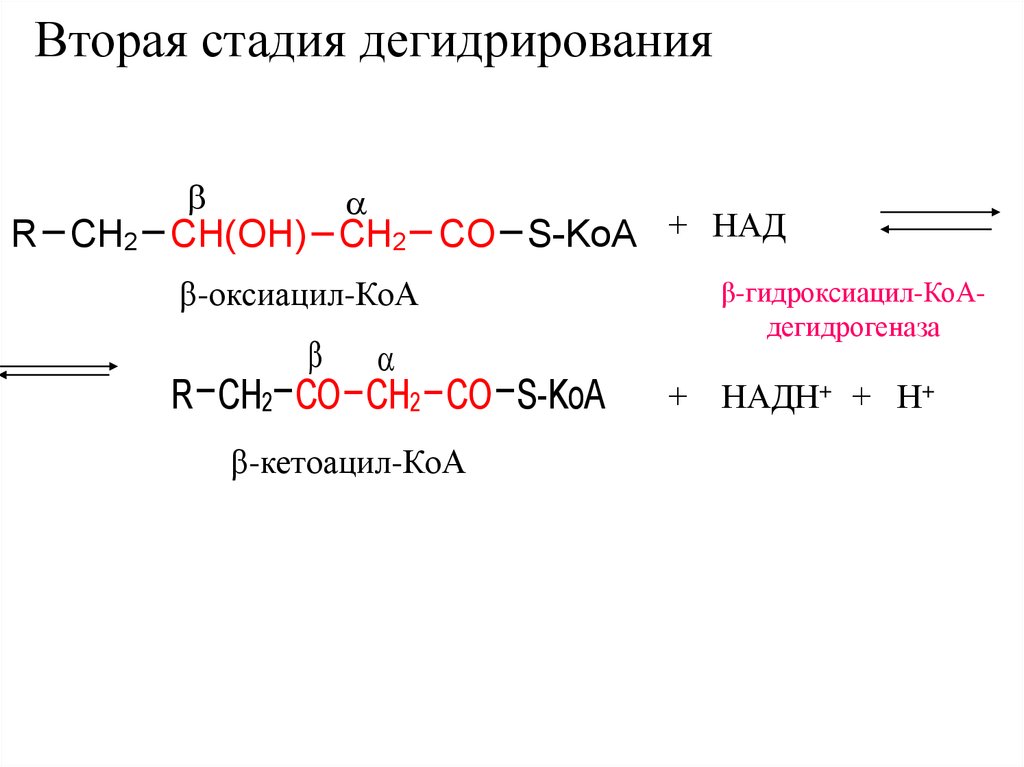
Вторая стадия дегидрированияR CH2 CH(OH) CH2 CO S-KoA + НАД
β-оксиацил-КоА
R CH2 CO CH2 CO S-KoA
β-кетоацил-КоА
β-гидроксиацил-КоАдегидрогеназа
+ НАДН+ + Н+
37.
Тиолазная реакцияR CH2 CO CH2 CO S-KoA
Тиолаза
β-кетоацил-КоА
R CO S-KoA
+ HS-KoA
+
Ацил-КоА вновь проходит путь β-окисления
вплоть до образования бутирил-КоА,
который окисляется до двух молекул ацетил-КоА.
38. Баланс β-окисления пальмитиновой кислоты (С16)
При окислении жирной кислоты, содержащей nуглеродных атомов
• получается n/2 ацетил-КоА,
• происходит (n/2 – 1) циклов β-окисления,
так как при окислении бутирил-КоА получаются сразу
2 молекулы ацетил-КоА.
Расчёт для пальмитиновой кислоты:
• 16/2 = 8 ацетил-КоА,
• 16/2 – 1 = 7 циклов β-окисления,
• 7*5 = 35
• 8*12 = 96
• 96 + 35-1=130 АТФ.
39. β-окисление жирных кислот с нечётным числом атомов углерода
В конечном итоге образуютсяацетил-КоА и пропионил-КоА.
АТФ, биотин-СО2
Пропионил-КоА
Метилмалонил-КоА
Карбоксилаза
Сукцинил-КоА
Цикл Кребса
мутаза
40. β-окисление ненасыщенных жирных кислот
• Наличие дополнительных ферментов изомеразыи эпимеразы обеспечивает возможность полного
окисления всех ненасыщенных жирных кислот.
• Осуществляется:
• перемещение двойной связи из положения 3-4 в
положение 2-3,
• изменение конфигурации двойной связи из цисв транс-положение при помощи фермента цистранс-еноил-КоА-изомеразы.
41. Окисление жирных кислот протекает в
• печени,• мышцах,
• жировой ткани.
Мышцы, миокард, печень активно
используют жирные кислоты как источники
энергии.
42. Регуляция b-окисления
Регуляция -окисления• Регуляторный фермент –
карнитинацилтрансфераза.
• Чем интенсивнее идёт распад АТФ, тем быстрее
окисляются жирные кислоты.
• Скорость -окисления зависит от доступности
субстрата ацил-КоА.
• -окисление активируется в постабсорбтивный
период или при длительной физической работе,
когда в результате распада жиров в жировой
ткани в крови увеличивается концентрация
жирных кислот
43. Биосинтез липидов идёт в
жировой ткани,
печени,
почках,
нервной ткани.
44. Биосинтез липидов зависит от распада глюкозы
• АТФ,• НАДФН2,
• ацетил-КоА.
45. Пути образования и использования Ацетил-КоА
Стероидныегормоны.
46. Биосинтез жирных кислот
идёт в цитоплазме,
нужен ацетил-КоА из митохондрий,
участвует малонил-КоА,
происходит перенос ацетил-КоА в цитоплазму,
участвует мультиферментный комплекс синтетаза
жирных кислот,
• требуется биотин,
• нужен НАДФН2,
• требуется АПБ на всех этапах.
47. Перенос ацетил-КоА из митохондрий в цитоплазму
Ацетил-КоА не проникает через мембрану
митохондрий в цитоплазму.
Ацетил-КоА + ЩУК
цитрат + НSКоА
• Цитрат с помощью транслоказы переносится в
цитоплазму.
• В цитоплазме:
цитрат + НSКоА +АТФ
НАДН+Н+
ЩУК
НАД+
малат
Ацетил-КоА +АДФ+Фн+ЩУК
Цитратлиаза
НАДФН+Н+
НАДФ+
ПВК+ СО2
Малик-фермент
48. Реакции синтеза жирных кислот
СН3-CО-S-KoA + биотин-СО2 + АТФАцетил-КоА-карбоксилаза
(инсулинзависимая)
НООС-СН2-CО-S-KoA + АДФ + Фн малонилКоА
49. Ацетил-КоА-карбоксилаза
• аллостерический фермент, активатором являетсяцитрат,
• повышение содержания цитрата в митохондриях
приводит к тому, что при помощи челночного
механизма он поступает в цитоплазму.
• Появление цитрата в цитоплазме –сигнал того, что
ЦТК перегружен «топливом» и избыток ацетилКоА должен запасаться в виде жира.
50. Далее действует мультиферментный комплекс – синтетаза жирных кислот, который представляет собой 6 ферментов, связанных с АПБ.
SHАПБ
SH
51.
Роль простетической группы
в АПБ играет 4фосфопантетеин. Это
подвижная «рука»,
переносящая остатки жирных
кислот от активного центра
фермента к другому. Вторая
SH-группа в молекуле 3кетоацил-АПБ-синтазы от
цистеина.
• Сульфгидрильные группы
синтетазы жирных кислот
вначале взаимодействуют с
ацильными группами:
ацетильная группа
присоединяется к SH-группе
цистеина, а малонильная к
SH-группе фосфопантетеина.
52.
СН3-CО-S-KoA + HS-АПБАПБ-ацетилтрансфераза
HS-КоА + СН3-CО-S-АПБ
ацетил-АПБ
53.
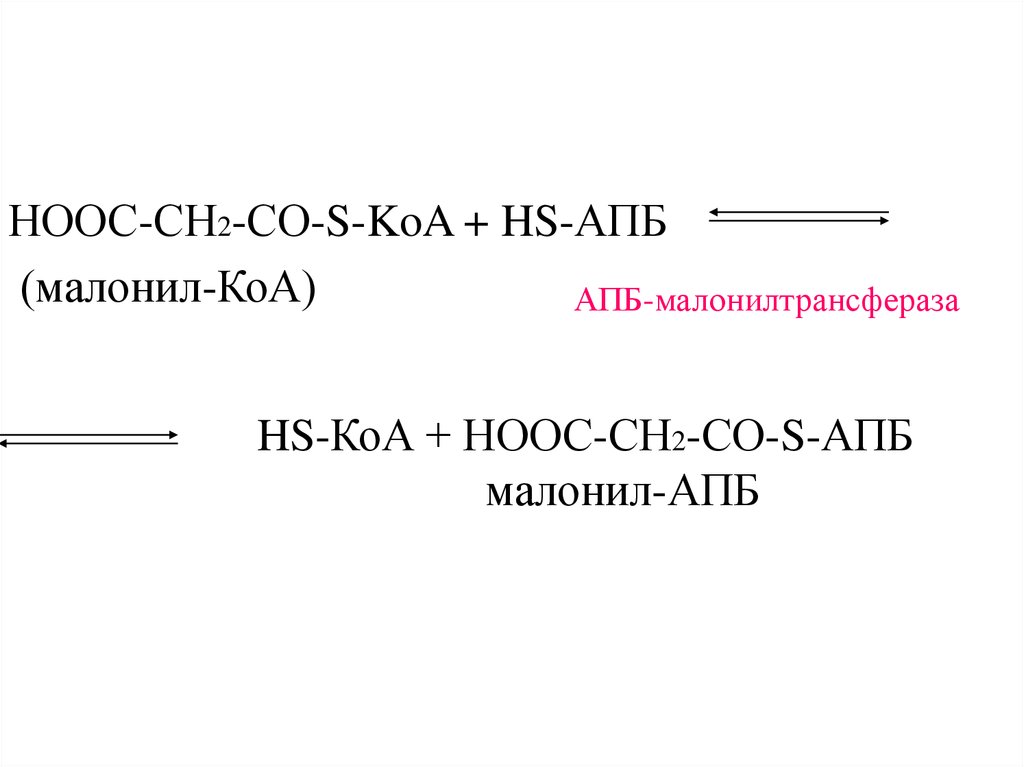
НООС-СН2-CО-S-KoA + HS-АПБ(малонил-КоА)
АПБ-малонилтрансфераза
HS-КоА + НООС-СН2-CО-S-АПБ
малонил-АПБ
54. Далее происходит конденсация малонил-АПБ и ацетил-АПБ.
СН3-CО-S-АПБ + НООС-СН2-CО-S-АПБацетил-АПБ
малонил-АПБ
СН3-CО-СН2-CО-S-АПБ + СO2
β-Кетоацилацетоацетил-АПБ
АПБсинтаза
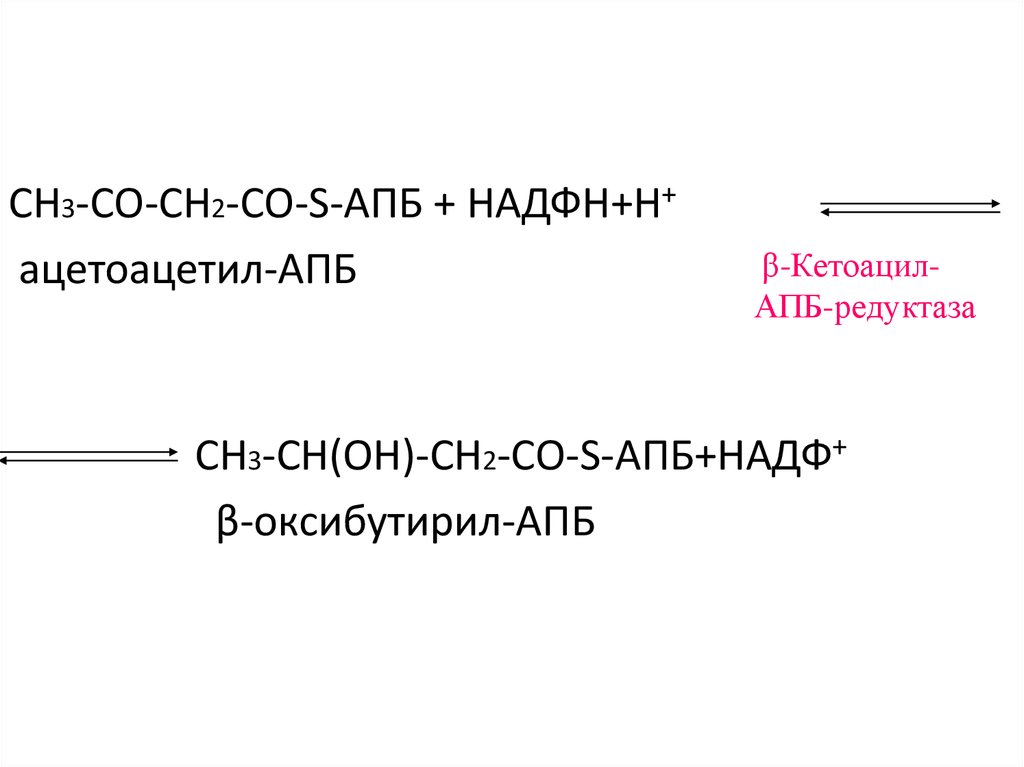
55.
СН3-CО-СН2-CО-S-АПБ + НАДФН+Н+ацетоацетил-АПБ
β-КетоацилАПБ-редуктаза
СН3-CH(ОH)-СН2-CО-S-АПБ+НАДФ+
β-оксибутирил-АПБ
56.
СН3-CH(ОH)-СН2-CО-S-АПБβ-оксибутирил-АПБ
β-оксиацилАПБ-дегидратаза
СН3-CH=СН-CО-S-АПБ + Н2О
кротонил-АПБ
57.
СН3-CH=СН-CО-S-АПБ + НАДФН+Н+кротонил-АПБ
еноилАПБ
-редуктаза
СН3-CH2-СН2-CО-S-АПБ +НАДФ+
бутирил-АПБ
58.
• Далее цикл повторяется.• Малонил-КоА переносится на SH-группу
фосфопантетеина АПБ.
Бутирил-АПБ + малонил-АПБ
кетокапронилАПБ + СО2
Пальмитиновая кислота – предшественник для
других жирных кислот.
59. Источники НАДФН+Н
• на 50% -пентозный цикл,• изоцитратдегидрогеназная реакция,
• малик-реакция.
НАДФН+Н+
Малат
НАДФ+
СО2 + ПВК
МДГ декарбоксилирующая
+
60. Полиненасыщенные жирные кислоты
• Линолевая, линоленовая жирные кислоты ворганизме не синтезируются.
• Арахидоновая кислота синтезируется из
линолевой, если последняя поступает в большом
количестве с пищей.
Мононенасыщенные жирные кислоты
• Олеиновая, пальмитоолеиновая жирные кислоты
синтезируются из пальмитиновой и стеариновой
кислот в микросомах клеток печени и жировой
ткани при участии оксигеназы и кислорода.
• Из олеиновой кислоты идёт синтез невроновой и
оксиневроновой кислот.
61. Биосинтез ТАГ
• В почках, стенке кишечника, печени высокаактивность глицеролкиназы.
+АТФ
Mg2+
+АДФ
глицеролкиназа
Глицерол-3-фосфат
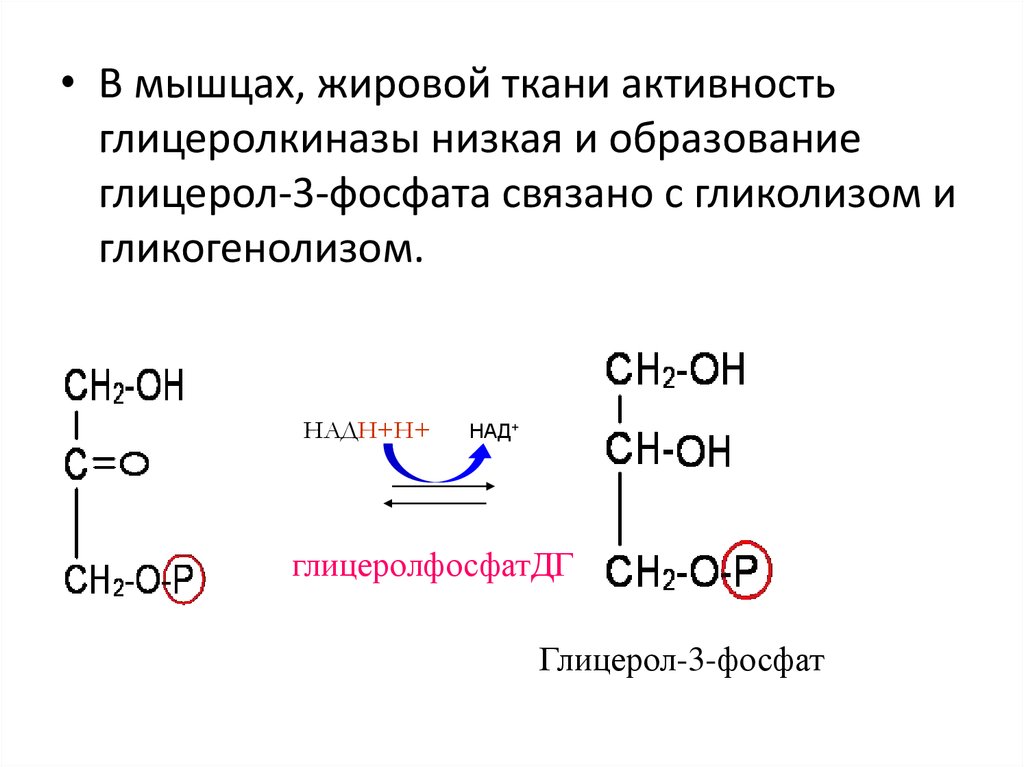
62.
• В мышцах, жировой ткани активностьглицеролкиназы низкая и образование
глицерол-3-фосфата связано с гликолизом и
гликогенолизом.
НАДН+Н+
НАД+
глицеролфосфатДГ
Глицерол-3-фосфат
63.
2Глицерол-3-фосфат
Фосфатидная
кислота
глицеролфосфатацил
трансфераза
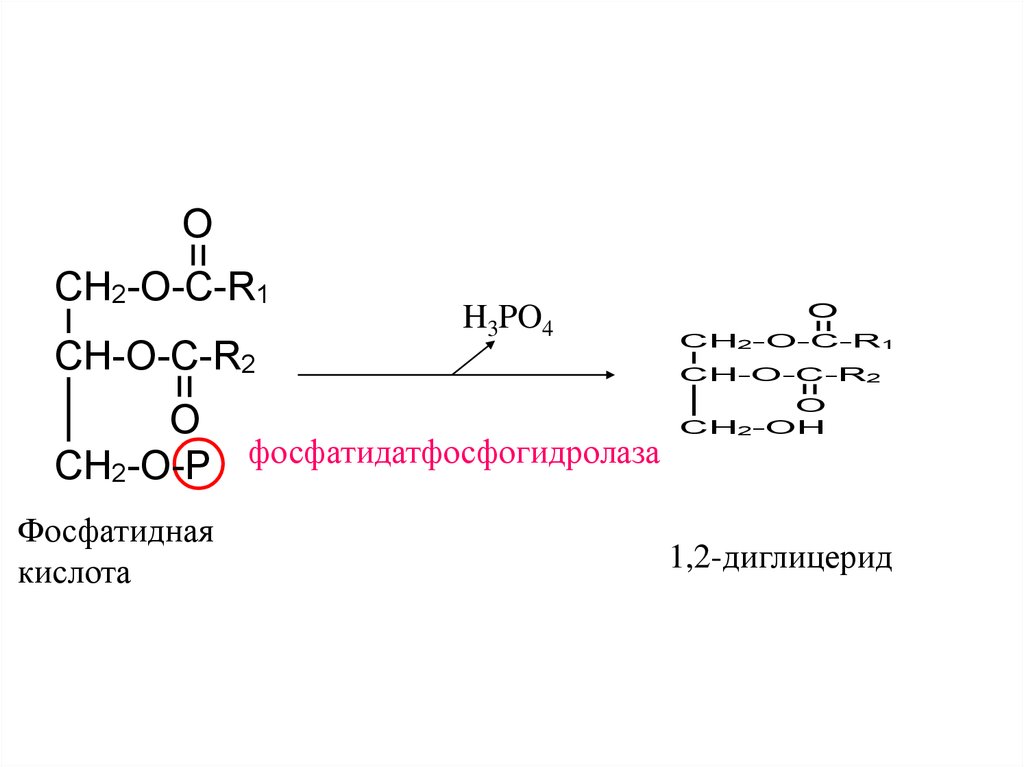
64.
CH2-O-C-R1CH-O-C-R2
H3PO4
O
CH2-O-P фосфатидатфосфогидролаза
Фосфатидная
кислота
CH2-O-C-R1
CH-O-C-R2
O
CH2-OH
1,2-диглицерид
+
R
65.
CH2-O-C-R1CH2-O-C-R1
CH-O-C-R2
O
CH2-OH
1,2-диглицерид
+ R3C
O
HSKoA
SKoA
диглицеридацилтрансфераза
CH-O-C-R2
O
CH2-O-C-R3
O
Триацилглицерин
Жиры, синтезированные в жировой ткани, там и
откладываются, а из печени жиры транспортируются
в составе липопротеинов
66. Показатели липидного обмена в крови здорового человека
Общие липиды
Триглицериды
Фосфолипиды
Холестерин
ЛПОНП
ЛПВП
ЛПНП
НЭЖК
4-10 г/л
1-1,5 г/л
2-2,5 г/л
1,5-2 г/л
1,2 г/л
3,5 г/л
4,5 г/л
0,1г/л
67. В плазме крови новорожденных
• содержание ЛП ниже, чем у взрослых,причём полностью отсутствуют ХМ, а
ЛПОНП резко снижены.
• Доля холестерина в ЛПВП новорожденных в
2 раза больше, чем у взрослых.
• С возрастом уровень ЛПВП снижается, а
ЛПНП - повышается.
• У новорожденных основным классом ЛП в
крови являются ЛПВП.