










")











")
















")






 и ингибирующими (Gi)")





")
.")














 Биология
Биология Химия
ХимияПохожие презентации:
")









Сигнальные молекулы. Химическая сигнализация
1. Сигнальные молекулы
2. Химическая сигнализация обеспечивает взаимодействие между клетками в многоклеточных организмах
3. СИГНАЛЬНЫЕ МОЛЕКУЛЫ
• Сигнальные молекулы • «КЛАССИЧЕСКИЕ»– это вещества,
ГОРМОНЫ
участвующие в
• ГОРМОНЫ МЕСТНОГО
химических
ДЕЙСТВИЯ и
механизмах передачи
функционально им
сигналов между
родственные:
клетками в
• Цитокины
многоклеточном
организме
• НЕЙРОМЕДИАТОРЫ
4. Общие свойства сигнальных молекул
• Выделяются во внеклеточное пространство (илиэкспрессируются на поверхности плазматической
мембраны);
• Не используются клетками в качестве пластического или
энергетического материала;
• Способны направленно взаимодействовать с клетками,
имеющими рецепторы для данного вида молекул;
• Обладают очень высокой биологической активностью и
вызывают эффекты в клетках при очень низких
концентрациях (около 10-11 - 10-6 моль/л, а в некоторых
случаях даже при 10-15 моль/л).
5. Варианты реализации нейро-гуморальных механизмов
Варианты реализации нейрогуморальных механизмовАутокринные
Паракринные
Эндокринные
6. Цитокины
• Цитокины — низкомолекулярные (8–50 kDa) белки и пептиды(в основном гликопротеины), которые продуцируются
различными типами клеток (моноцитами, тканевыми
макрофагами, лимфоцитами, клетками
ретикулоэндотелиальной системы, эндотелием,
нейтрофилами).
• Биологическая функция цитокинов заключается в регуляции
защитных реакций организма на местном и системном уровнях,
а также в регуляции процессов регенерации тканей после
повреждения.
• Определенный цитокин может связываться с рецепторами на мембране той
же самой клетки, которая его секретировала, оказывая аутокринное
действие; он может связываться с рецепторами на клетке-мишени в
непосредственной близости от клетки-продуцента, оказывая паракринное
действие; в некоторых случаях он может связываться с клетками-мишенями
в отдаленных частях тела, оказывая эндокринное действие
7.
Группы цитокинов:- провоспалительные (интерлейкины 1,2,6,8,
ФНОα, интерферон γ);
- противовоспалительные (интерлейкины 4,10,
TGFβ);
- регуляторы клеточного и гуморального
иммунитета — (естественного или
специфического), обладающие собственными
эффекторными функциями (противовирусными,
цитотоксическими).
8. Семейства цитокинов
Наиболее часто выделяют следующие семейства цитокинов:• Интерлейкины (регулируют межклеточные взаимодействия между
лейкоцитами);
• Факторы некроза опухолей (TNFα, который продуцируется макрофагами,
моноцитами и другими клетками, и TNFβ (лимфотоксины), продуцируемые
T- и B-лимфоцитами);
• Хемокины (Хемокины играют важную роль в координации движения Т- и Влимфоцитов, дендритных и других клеток при их дифференцировке, участии
в иммунном ответе. Хемокины инициируют локальное воспаление в
результате вовлечения клеток в процесс хемотаксиса )
• Интерфероны (блокируют репликацию вируса в соседних клетках и
участвуют во взаимодействии между клетками иммунной системы) ;
• Колониестимулирующие факторы (регулируют деление и дифференцировку
костномозговых стволовых клеток и клеток кроветворного ростка, могут
стимулировать дифференцировку и функциональную активность некоторых
клеток вне костного мозга) ;
• Ростовые факторы ( играют ключевую роль в клеточной пролиферации,
дифференцировке, апоптозе и миграции)
9. НЕЙРОМЕДИАТОРЫ
• Сигнальные молекулы, обеспечивающиепередачу сигналов от нейрона к нейрону
или от нейрона к другой клетке
10. Классические нейромедиаторы должны отвечать следующим критериям:
• Образовываться в нейронах и/или накапливаться впресинаптических пузырьках механизмом активного
транспорта.
• Выделяться через пресинаптическую мембрану в
синаптическую щель в ответ на повышение концентрации
Ca2+ в нейроне.
• При достижении постсинаптической мембраны
взаимодействовать со специфическими рецепторами
постсинаптической мембраны с последующим
изменением мембранного потенциала.
• Разрушаться и/или подвергаться удалению из синапса
после диссоциации из комплекса с рецептором
11. По химическому строению нейромедиаторы могут быть разделены на три группы:
1) низкомолекулярные органические вещества (глицин иL-глутамат, ГАМК, гистамин, дофамин, цетилхолин,
АТФ и др.);
2) газообразные нейромедиаторы (СО, NO, H2S);
3) нейромедиаторы пептидной природы (опиоиды:
эндорфины, энкефалины, динорфины; гипофизарные:
вазопрессин, окситоцин; гипоталамические:
тиреолиберин, соматостатин, кортистатин, а также
нейропептид Y и др.)
12. Гормоны местного действия (тканевые гормоны)
• образуются не в эндокринных железах, а в тканях разныхорганов
• обладают аутокринным и паракринным действием
К ним относятся тканевые гормоны пищеварительного тракта, а также
гормоны, регулирующие тонус кровеносных сосудов и возбудимость
нервной системы.
По химической природе тканевые гормоны подразделяются
на три группы:
1) эйкозаноиды (производные арахидоновой кислоты:
простагландины, простациклины, тромбоксаны, лейкотриены );
2) тканевые гормоны пептидной природы (гастрин, секретин,
холецистокинин, ангиотензин II, эритропоэтин, кинины) ;
3) биогенные амины (производные аминокислот: гистамин ,
серотонин, мелатонин ).
13. ГОРМОНЫ
• Гормоны (классические) – этобиологически активные вещества,
которые синтезируются в малых
количествах клетками эндокринной
системы и через циркулирующие
жидкости доставляются к клеткаммишеням, где оказывают свое
регулирующее действие (телекринный
эффект). В некоторых случаях могут
действовать и по паракринному
механизму
• В настоящее время у человека описано порядка 70
различных гормонов.
14. Классификация гормонов
По химическому строению:• Белково-пептидные гормоны
• Гормоны – производные аминокислот
• Стероидные гормоны
По физико-химическим свойствам выделяют:
гидрофильные (гормоны белково-пептидной
природы и биогенные амины) и липофильные
гормоны (тиреоидные и стероидные гормоны).
.
15. Белково-пептидные гормоны синтезируются
• В гипоталамусе (либерины, статины)• В гипофизе (АКТГ, ТТГ, ФСГ, ЛГ, гормон роста,
пролактин, вазопрессин, окситоцин)
• В поджелудочной железе (инсулин, глюкагон,
соматостатин)
• В щитовидной железе (кальцитонин )
• В паращитовидных железах (паратгормон )
• Ряд гормонов синтезируется неэндокринными
органами (лептин, гастрин, холецистокинин,
остеокальцин, эритропоэтин и пр.)
16. Синтез и созревание белково-пептидных гормонов
1) синтез на рибосомах гранулярного ЭПС длинногопредшественника гормона называемого препрогормоном;
2) отщепление от препрогормона сигнального пептида с
участием сигнальных пептидаз внутри цистерн грЭПС с
образованием прогормона;
3) дальнейшая фрагментация прогормона в комплексе
Гольджи с участием трипсиноподобных
прогормонконвертаз;
4) отщепление у некоторых гормонов С-концевых остатков
лизина и аргинина с участием карбоксипептидазы Е.
Часть гормонов подвергается дальнейшему С-концевому амидированию,
N-концевому ацетилированию, циклизации N-концевого глутамина,
N-, О-гликозилированию, реже ацилированию (грелин), либо
фосфорилированию (АКТГ) по остаткам серина.
17. Прекращение действия белково-пептидных гормонов
После взаимодействия гормонов с рецепторами , Гормонрецепторные комплексы интернализуются, после чегогормоны подвергаются тотальному протеолизу в
лизосомах. Рецепторы при этом рециркулируют из
эндосом обратно на клеточную мембрану и тем самым
повторно используются клеткой. Часть гормонов
разрушается в печени и почках. Некоторые гормоны
белково-пептидной природы подвергаются инактивации
по механизму ограниченного протеолиза с участием
дипептидил-пептидазы-4 (ДПП-4), которая отщепляет Хпролин дипептиды с N-конца различных гормонов
белково-пептидной природы, факторов роста, хемокинов.
18. Гормоны – производные аминокислот синтезируются
• В щитовидной железе (тиреоидныегормоны тироксин, или Т4, и
трийодтиронин, или Т3)
• В мозговом веществе надпочечников
(норадреналин, адреналин)
• В эпифизе (мелатонин)
19. Биосинтез тиреоидных гормонов
20.
Т3 и Т4 в последствии подвергаютсядейодированию с участием йодотирониндегалогеназ клеток-мишеней. Данные ферменты
содержат в активном центре остаток
селеноцистеина, принимающий участие в
дейодировании гормонов. Йодотирониндегалогеназа 2-го типа (5’-дейодиназа)
катализирует дейодирование наружного кольца
битирозинов, превращая малоактивный Т4 в
активный Т3. Фермент располагается в мембране
ЭПС рядом с ядром, в итоге образовавшийся Т3
сразу же попадает в ядро, где оказывает свое
действие.
21. Инактивация тиреоидных гормонов
осуществляется путем дейодирования, с участиемдегалогеназы 3-го типа (5-дейодиназы). Данный
фермент катализирует дейодирование
внутреннего кольца Т4 и Т3, что приводит к
образованию лишенных биологической
активности йодотиронинов, реверсивного Т3, rT3,
(в отличие от активного Т3 у rT3 остаток йода
отсутствует во внутреннем кольце) и
дийодтиронина, Т2. Поступившие в печень
йодотиронины претерпевают гидрофилизацию
путем сульфатирования, либо глюкуронирования
по свободному фенольному гидроксилу, после
чего удаляются в желчь.
22. Синтез катехоламинов
23. Инактивация катехоламинов с помощью моноаминооксидаз А и В:
Определение концентрации в суточной моче ванилилминдальнойкислоты, конечного продукта деградации адреналина и норадреналина,
применяется в диагностике феохромоцитомы, опухоли мозгового
вещества надпочечников.
24. Инактивация катехоламинов с помощью катехол-О-метилтрансферазы (КОМТ)
25. Мелатонин – гормон эпифиза. Синтезируется ночью из серотонина путем N-ацетилирования и метилирования 5- ОН группы. На свету
снижается образование. Мелатонин вклетках не хранится, а сразу по мере синтеза
высвобождается в кровь. Является главным регулятором
суточных ритмов. Обладает нейроэндокринными
эффектами, в частности тормозит секрецию гонадотропинов
и других гормонов гипофиза.
26. Деградация мелатонина
Происходит в клетках-мишенях путемпоследовательных реакций деацетилирования и
дезаминирования под действием
моноаминоксидазы А. Часть мелатонина
обезвреживается в печени посредством
гидроксилирования и сульфатирования в 6-м
положении индольного кольца (участвуют
цитохром P450-зависимые монооксигеназы и Осульфотрансферазы); далее неактивное
сульфатированное производное экскретируется в
желчь
27. Стероидные гормоны синтезируются
• В коре надпочечников• В семенниках
• В яичниках
28. Стероидные гормоны – производные холестерина
• С21 – гормоны коры надпочечников и прогестерон• С19 – мужские половые гормоны
(андрогены, тестостерон)
• С18 – женские половые гормоны (эстрогены)
29. Схема синтеза стероидных гормонов
30. Инактивация стероидов
Протекает в печени, почках, кишечнике ислюнных железах путем окисления
гидроксила в 21-м положении с
образованием кето-группы. Далее в печени
идут реакции гидроксилирования и
глюкуронирования. При инактивации
стероидных гормонов образуются
производные с более низкой активностью,
которые выводятся из организма с мочой и
частично с желчью.
31. Иерархические взаимосвязи в эндокринной системе
32. Пути регуляции синтеза и секреции гормонов
1) отрицательная обратная связь:• ↑ концентрации глюкозы → ↑продукции инсулина → ↓
концентрации глюкозы;
• ↑ продукции паратгормона → ↑ концентрации кальция
→ ↓ продукции паратгормона;
• ↑ продукции АКТГ → ↑ продукции кортизола → ↓
продукции АКТГ.
2) положительная обратная связь (редко):
• ↑ продукции ЛГ → ↑ продукции экстрогенов → ↑
продукции ЛГ.
3) изменение количества рецепторов к гормону:
• ↑ продукции кортизола → ↑ количества
адренорецепторов → ↑ эффектов адреналина
33. Транспорт гормонов
Гидрофильные гормоны транспортируются кровью всвободном виде, липофильные с помощью
специальных транспортных белков.
Транспортные белки создают резервуар гормонов в крови, поскольку гормоны
в связанном виде не подвергаются расщеплению и выведению. При этом
активен только несвязанный с белками (свободный) гормон. При
разрушении свободных форм таких гормонов будет происходить
диссоциация связанной формы. Тем самым концентрация гормона в крови
будет поддерживаться на относительно постоянном уровне. В связи с этим
стероидные и тиреоидные гормоны имеют длительный период жизни в
русле крови (часы). Поскольку белково-пептидные гормоны и биогенные
амины не имеют специальных транспортных белков, они оказывают более
быстрый эффект, но при этом период их жизни меньше (минуты).
34. КЛЕТКИ – МИШЕНИ
• Специфически взаимодействуют ссигнальными молекулами с помощью
специальных белков-рецепторов.
Белки-рецепторы могут располагаться на
наружной мембране клетки, в цитоплазме,
или на ядерной мембране и на других
органеллах клетки.
35. Белки-рецепторы
ФУНКЦИИ РЕЦЕПТОРАСТРОЕНИЕ РЕЦЕПТОРА
• «Узнавание» гормона
• Гормон-связывающий
участок
• «Передающий»
участок
• Преобразование и
передача сигнала
36. Кинетика связывания гормона и рецептора
37. Рецепторы сигнальных молекул
• Гормоны белково-пептидной природы и биогенныеамины вследствие своей гидрофильности не
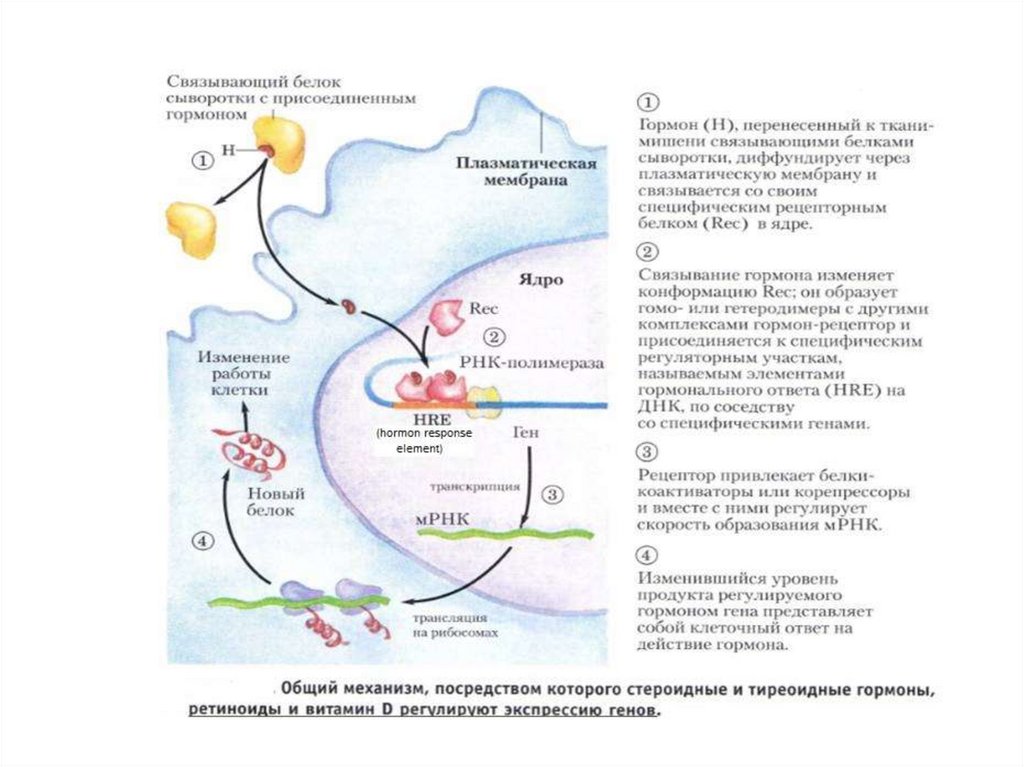
проникают через клеточные мембраны и воздействуют
на мембранные рецепторы.
• Стероидные и тиреоидные гормоны, обладающие
липофильностью, наоборот, проникают в клетку и
взаимодействуют с цитоплазматическими и ядерными
рецепторами.
Таким образом, выделяют гормоны с мембранным и
внутриклеточным механизмом действия.
38. МЕХАНИЗМЫ ВЗАИМОДЕЙСТВИЯ ГОРМОНОВ И КЛЕТОК-МИШЕНЕЙ
• Мембранный- Аденилатциклазная система
- Гуанилатциклазная система
- Инозитолфосфатный механизм
- Тирозин-киназный механизм
• Внутриклеточный
39. Этапы передачи сигнала в клетку
1) связывание лиганда (сигнальной молекулы) с рецептором;2) активация рецептора;
3) синтез в клетке молекул, усиливающих сигнал в ответ на
активацию рецептора;
4) активация ферментативных реакций в клетке;
5) ослабление сигнализации.
На каждом этапе возможны нарушения передачи сигнала,
что является причиной развития ряда заболеваний.
40. Рецепторы, локализованные в мембране Ионотропные:1 - содержащие субъединицу, связывающую сигнальную молекулу и ионный канал,
например рецептор ацетилхолина напостсинаптической мембране; Метаботропные: 2 - проявляющие
каталитическую активность после присоединения сигнальной молекулы,
например рецептор инсулина, обладающий тирозинкиназной активностью или
рецептор НУФ, обладающий гуанилатцикдазной активностью; 3, 4 передающие сигнал на фермент аденилатциклазу (АЦ) или фосфолипазу С
(ФЛС) при участии мембранных G-белков. Например разные типы рецепторов
адреналина, ацетилхолина и других сигнальных молекул
41. Рецепторы класса I (ионотропные)
Это рецепторы быстрого ответа. Содержатся впостсинаптической , а иногда и пресинаптической
мембране. После взаимодействия с нейромедиатором
ответ акцепторной клетки достигается за несколько
миллисекунд. Такая быстрая реакция становится
возможной благодаря тому, что рецептор представляет
собой олигомерный белок, имеющий ион-селективную
пору. Взаимодействие рецептора с нейромедиатором
приводит к перераспределению ионов, вызывая
деполяризацию (возбуждающие нейромедиаторы) или
гиперполяризацию постсинаптической мембраны
(тормозные нейромедиаторы). Примерами рецепторов
класса I являются рецепторы к глутамату, ГАМК, глицину,
никотиновый рецептор ацетилхолина.
42.
43.
44. Ионотропный рецептор N-холинорецептор
45. Внутриклеточные посредники передачи гормонального сигнала при мембранном механизме через метаботропные рецепторы
циклические нуклеотиды (цАМФ и цГМФ),
инозитолтрифосфат,
кальций-связывающий белок - кальмодулин,
ионы кальция,
ферменты синтеза циклических нуклеотидов,
Фосфолипаза С, фосфолипаза А2
протеинкиназы - ферменты фосфорилирования белков,
протеинфосфатазы - ферменты дефосфорилирования белков.
46. Концентрация цАМФ в клетке находится под контролем двух ферментов: аденилатциклазы и фосфодиэстеразы
47. Функционирование аденилатциклазной системы
(Gs-стимулирующий белок)(Gi- ингибирующий белок)
48. Трансмембранная передача сигналов через адренергические рецепторы, сопряженные с активирующими (Gs) и ингибирующими (Gi)
G-белками49. Аденилатциклазный путь, приводящий к экспрессии специфических генов Многие белковые гормоны: глюкагон, вазопрессин, паратгормон
и др., передающие свойсигнал посредством аденилатциклазной системы, могут не только вызвать изменение
скорости реакций путем фосфорилирования уже имеющихся в клетке ферментов, но и
увеличивать или уменьшать их количество, регулируя экспрессию генов посредством
фосфорилирования фактора транскрипции СRЕВ
CREB (cAMP response elementbinding protein)
CRE (cAMP response elements)
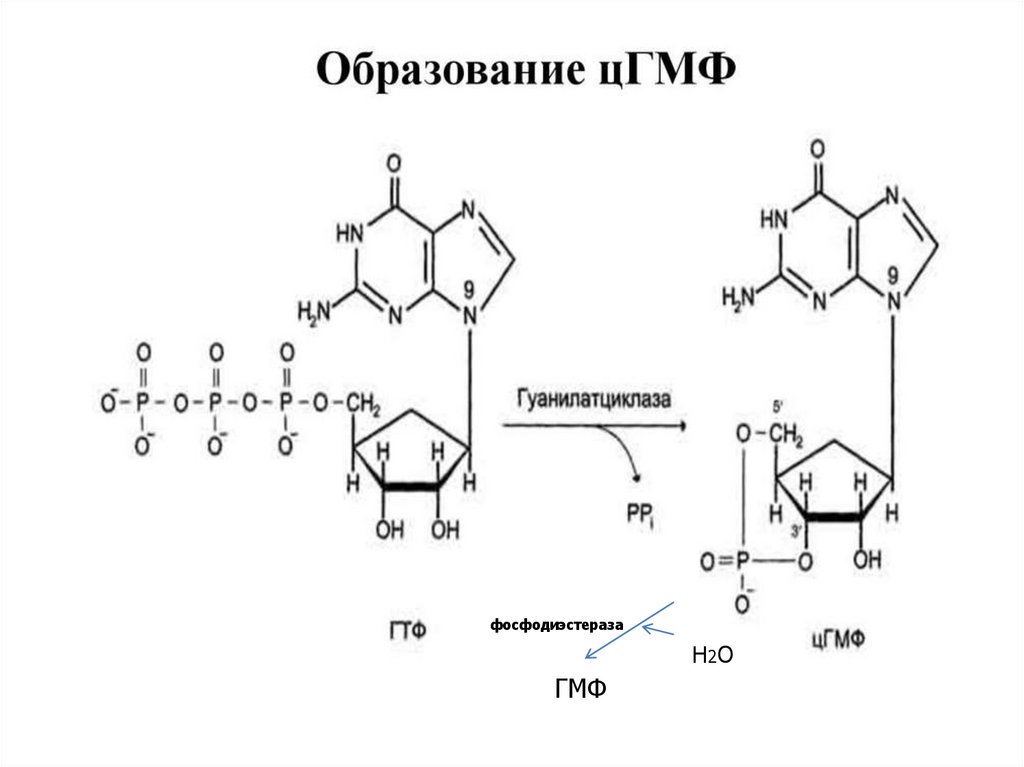
50.
фосфодиэстеразаН2 О
ГМФ
51.
52.
53.
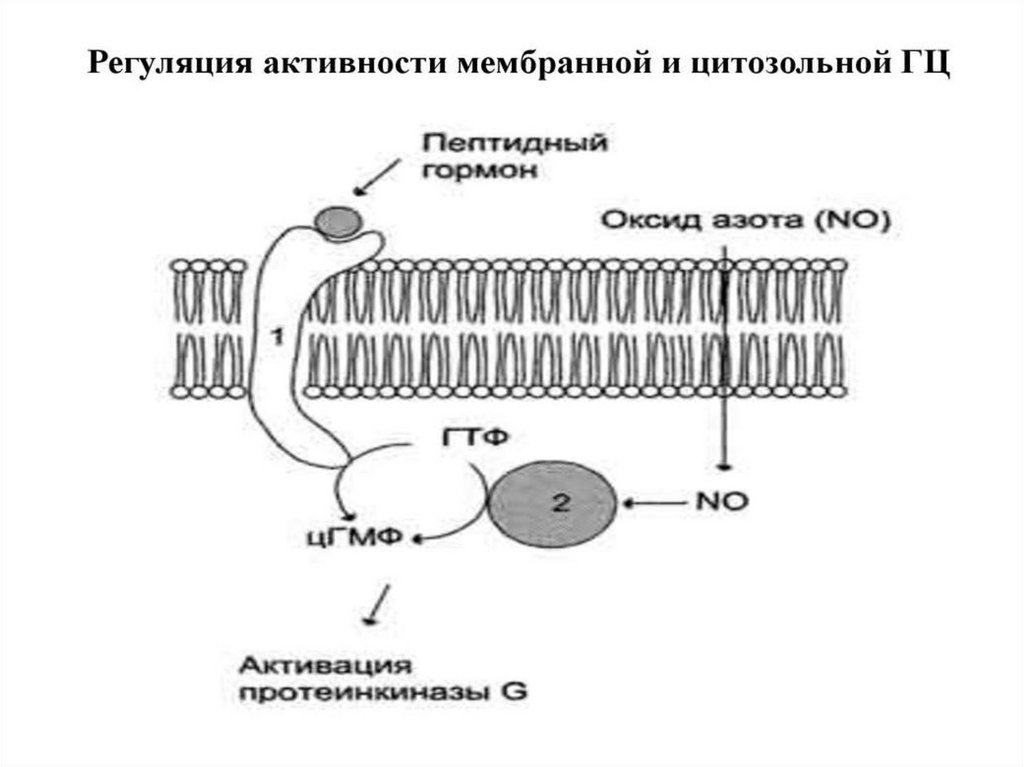
54. Регуляция активности мембранной гуанилатциклазы Одним из первичных мессенджеров протеинкиназы G является предсердный
натриуретический фактор (ПНФ), регулирующийгомеостаз жидкости в организме. ПНФ вызывает снижение
реабсорбции натрия в дистальном извитом канальце через цГМФ-зависимое
фосфорилирование эпителиальных натриевых каналов.
55. Через аденилатциклазу действуют
• Сигнальные молекулы, активирующие АЦкортиколиберин, соматолиберин, глюкагон, вазопрессин,
ЛГ, ФСГ, ТТГ, хорионический гонадотропин, АКТГ,
паратгормон, простагландины, бета-адренергические
катехоламины
• Сигнальные молекулы, угнетающие АЦ
соматостатин, ангиотензин II, ацетилхолин
(мускариновый эффект), дофамин, опиоиды, альфа-2адренергические катехоламины
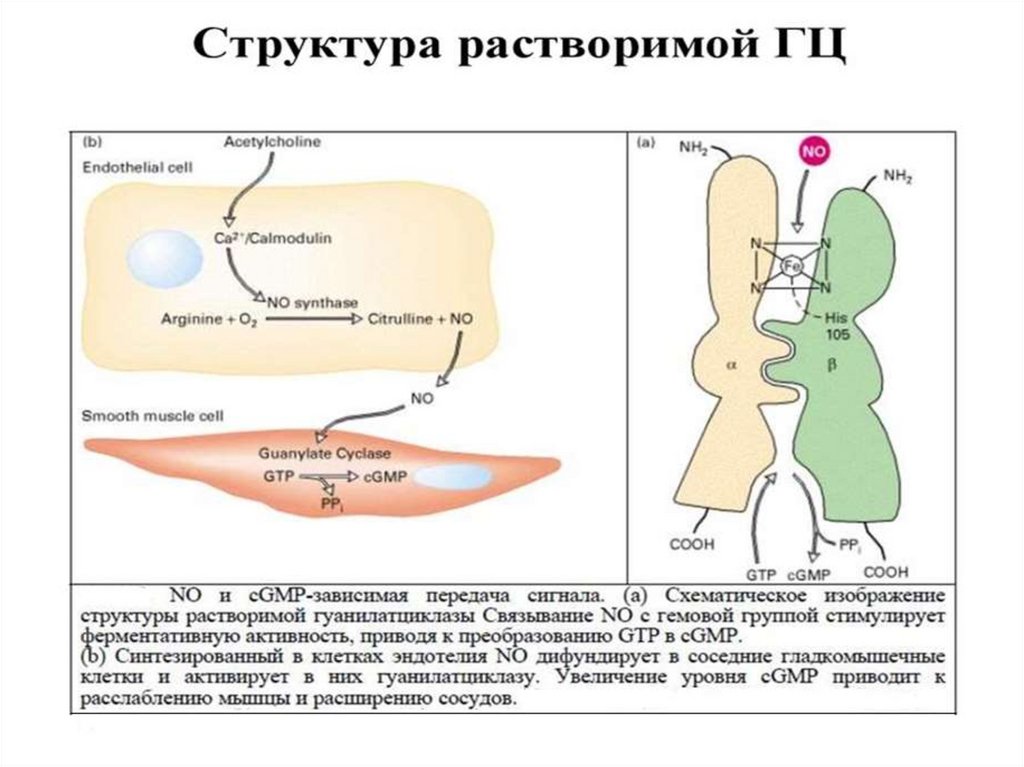
Действуют с участием гуанилатциклазы
предсердный натрийуретический гормон, гистамин,
ацетилхолин, брадикинин, эндотелин, NO
56. Инозитолфосфатная система
57. Гидролиз фосфатидилинозитол-4,5-бисфосфата (ФИФ2)
58. Са2+-канал мембраны эндоплазматического ретикулума, регулируемый инозитол-1,4,5-трифосфатом (ИФ3).
59. Через инозитолфосфатный механизм действуют
• гонадолиберин, тиролиберин, дофамин,тромбоксаны А2, лейкотриены,
ангиотензин II, эндотелин, паратгормон,
нейропептид Y,
aльфа-1адренергические катехоламины,
ацетилхолин, брадикинин, вазопрессин
60. Регуляция активности фосфолипазы А2
Основными факторами, регулирующими активность цитозольной фосфолипазы,
являются концентрация внутриклеточного Са2+ и активность
митогенактивируемых протеинкиназ (MAPK) .
• В неактивированных клетках концентрация внутриклеточного Са2+ обычно
колеблется в пределах 30-100 нмоль/л. При активации различных рецепторов
концентрация ионов Са2+ может достигать 1-3 мкмоль/л. Увеличение активности
цитозольной ФЛА2 происходит в диапазоне концентраций ионов Са2+ 150-800
нмоль/л. При увеличении концентрации ионов Са2+ происходит миграция
фосфолипазы к мембранам и её прикрепление к ним (подобно ПКС), после чего
начинается гидролиз фосфолипидов. Основными субстратами ФЛА2 становятся
фосфолипиды с арахидоновой кислотой в положении sn-2
• фосфорилирование митогенактивируемыми протеинкиназами (MAPK) по остатку
серина приводит к значительному увеличению активности фосфолипазы А2.
Т. о., цитозольная ФЛА2 принимает участие как в регуляции синтеза эйкозаноидов
при остром ответе клеток на различные провоспалительные стимулы, так и в ряде
случаев при отложенном ответе
61. Роль фосфолипазы А2 в синтезе эйкозаноидов
62.
ЭйкозаноидыКомплекс физиологически активных органических соединений
являющихся производными арахидоновой (С20:4;5,8,11,14) кислоты
Эйкозаноиды служат вторичными мессенджерами
гидрофильных гормонов , передающих сигналы при посредничестве G-белков
Эйкозаноиды являются паракринными регуляторами
Эффекты эйкозаноидов определяются свойствами клеток-мишеней
63.
Лейкотриены1) Активируют аллергические реакции
2) Активируют иммунные реакции
3) Активируют анафилактические реакции
4) Сокращение гладких мышц дыхательных путей
5) Сокращение гладких мышц пищеварительного тракта
6) Оказывают сосудосуживающие действие
64.
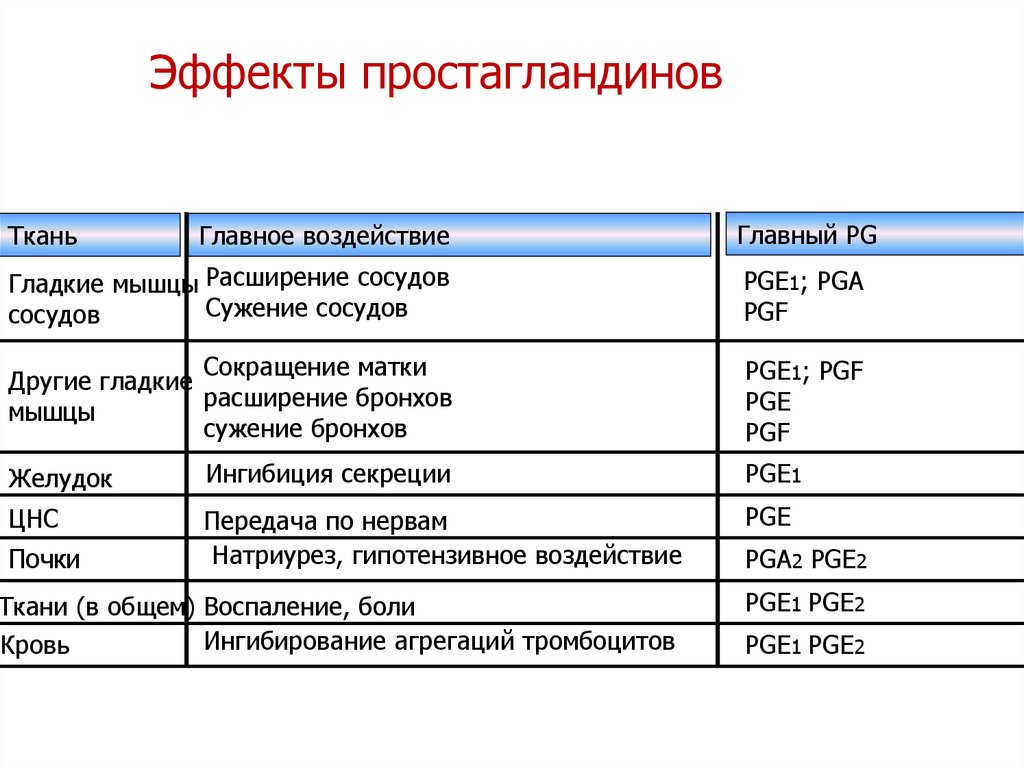
Эффекты простагландиновГлавное воздействие
Главный PG
Гладкие мышцы Расширение сосудов
Сужение сосудов
сосудов
PGЕ1; PGА
PGF
Сокращение матки
Другие гладкие
расширение бронхов
мышцы
сужение бронхов
PGЕ1; PGF
PGE
PGF
Ткань
Желудок
Ингибиция секреции
PGЕ1
ЦНС
Передача по нервам
Натриурез, гипотензивное воздействие
PGЕ
Почки
Ткани (в общем) Воспаление, боли
Ингибирование агрегаций тромбоцитов
Кровь
PGA2 PGE2
PGЕ1 PGE2
PGЕ1 PGE2
65. Тирозин-киназный каталитический рецептор
• Обладает сродством к гормону• Гормон-рецепторный комплекс проявляет
киназную активность – участвует в
фосфорилировании белков в цитоплазме
клетки-мишени
66. Тирозин-киназа принимает участие в реализации эффектов
• Инсулина• Макрофагального
колониестимулирующего фактора
• Фактора роста тромбоцитов
67. Активация рецептора инсулина - тирозиновой протеинкиназы.
Активация рецептора инсулина тирозиновой протеинкиназы.Присоединение инсулина в центр связывания,
образованный а-субъединицами, вызывает
кооперативные конформационные изменения
рецептора. β-Субъединицы проявляют тирозинкиназную
активность и катализируют трансаутофосфорилирование
(первая β-субъединица фосфорилирует вторую βсубъединицу, и наоборот) по нескольким остаткам
тирозина. Фосфорилирование приводит к изменению
заряда, конформации и субстратной специфичности
фермента (Тир-ПК). Тирозиновая-ПК фосфорилирует
определенные клеточные белки, которые получили
название субстратов рецептора инсулина (IRS 1). В свою
очередь эти белки участвуют в активации каскада
реакций фосфорилирования:
• фосфопротеинфосфатазы (ФПФ), которая
дефосфорилирует специфические фосфопротеины;
• фосфодиэстеразы, которая превращает цАМФ в АМФ и
цГМФ в ГМФ;
• ГЛЮТ 4 - переносчиков глюкозы в инсулинзависимых
тканях, поэтому повышается поступление глюкозы в
клетки мышц и жировой ткани;
• тирозиновой протеинфосфатазы, которая
дефосфорилирует β-субъединицы рецептора инсулина;
• регуляторных белков ядра, факторов транскрипции,
повышающих или снижающих экспрессию генов
определенных ферментов.
IRS 1
IRS 1
68. Механизмы удаления вторичных посредников из цитозоля
• Действие фосфодиэстеразы (гидролиз ц-АМФили ц-ГМФ до АМФ или ГМФ соответственно)
• Удаление Ca++ из цитоплазмы в результате его
выхода из клеток или возврата в цистерны
саркоплазматического ретикулума
• Связывание кальция кальций-связывающими
белками.
69. Общие черты мембранных механизмов передачи сигналов
одним из этапов передачи сигнала
является фосфорилирование белков
прекращение активации происходит
благодаря механизмам, инициируемым
самими участниками процессов
70. ЭФФЕКТЫ, ВОЗНИКАЮЩИЕ В КЛЕТКАХ-МИШЕНЯХ
ЭФФЕКТЫ, ВОЗНИКАЮЩИЕ В КЛЕТКАХМИШЕНЯХ• Изменение скорости биосинтеза отдельных
белков (в том числе белков-ферментов);
• Изменение активности уже существующих
ферментов (например, в АЦ-системе и т.п.);
• Изменение проницаемости мембран в клеткахмишенях для отдельных веществ или ионов
(например, для Са+2).
71. Схема строения рецептора кортизола
• 1 – «Антигенный» участок (регулируетфункции промотора
• 2 – Участок для связывания с ДНК
• 3 – Участок связывания с гормоном
72.
73. Эффективность действия гормонов зависит от многих факторов
• Концентрация гормона• Количество (и качество) рецепторов на
мембране или в цитоплазме клеткимишени
• pH среды
• Концентрация ионов
• Присутствие лигандов,
взаимодействующих с рецепторами
74. Причины эндокринных нарушений
• Недостаточность гормона (гипосекреция)• Гиперсекреция гормона
• Нарушения, связанные с рецепторным аппаратом
клеток-мишеней
- Недостаточность синтеза белков-рецепторов
- Генетические дефекты белка-рецептора, приводящие к
снижению сродства к гормонам
- Блокирование белков-рецепторов лигандами (например,
антителами при аутоиммунных заболеваниях)