





























 Биология
БиологияПохожие презентации:
")

")






")
Лекция 7. Активный обмен. Энергобаланс
1. Лекция 7
2.
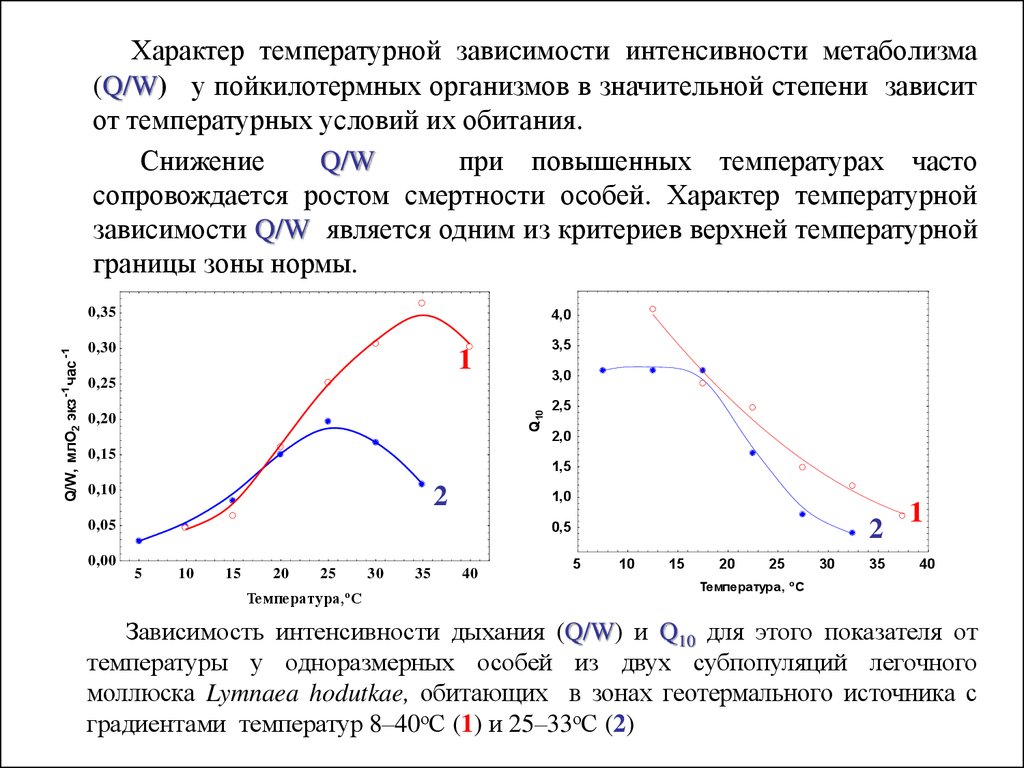
Характер температурной зависимости интенсивности метаболизма(Q/W) у пойкилотермных организмов в значительной степени зависит
от температурных условий их обитания.
Снижение
Q/W
при повышенных температурах часто
сопровождается ростом смертности особей. Характер температурной
зависимости Q/W является одним из критериев верхней температурной
границы зоны нормы.
4,0
0,30
3,5
1
3,0
0,25
Q10
Q/W, млО2 экз -1 час -1
0,35
0,20
0,15
2,5
2,0
1,5
0,10
2
1,0
0,05
2
0,5
0,00
5
10
15
20
25
Те мпе ратура,о С
30
35
40
5
10
15
20
25
30
35
1
40
Температура, о С
Зависимость интенсивности дыхания (Q/W) и Q10 для этого показателя от
температуры у одноразмерных особей из двух субпопуляций легочного
моллюска Lymnaea hodutkae, обитающих в зонах геотермального источника с
градиентами температур 8–40оС (1) и 25–33оС (2)
3.
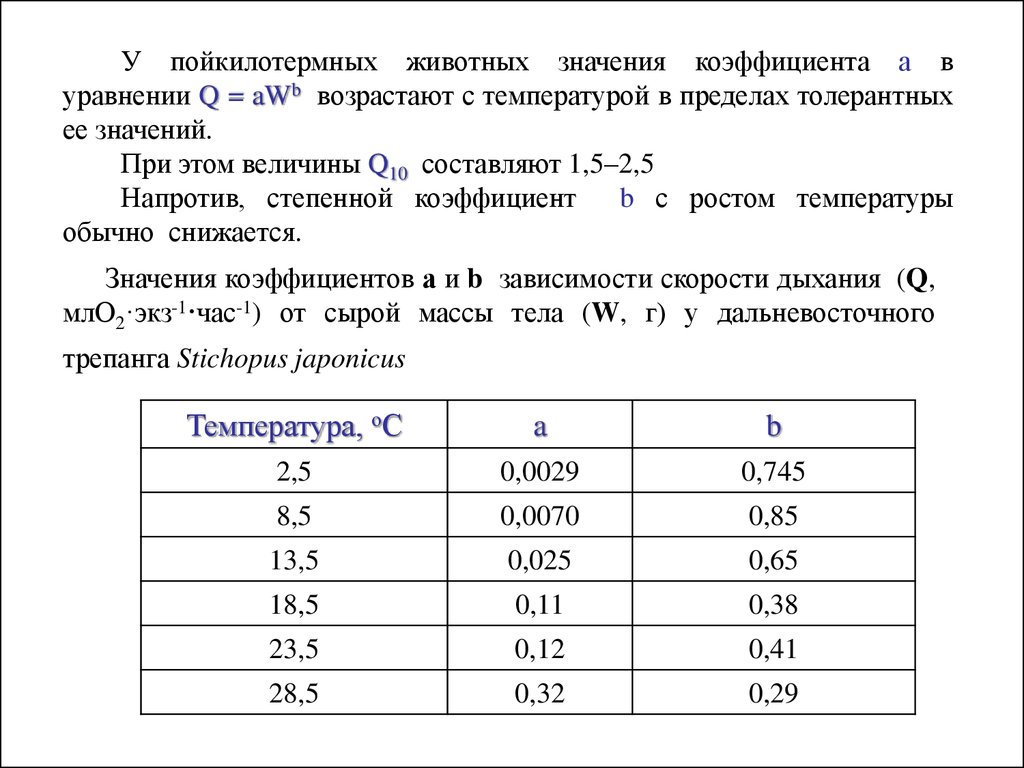
У пойкилотермных животных значения коэффициента а вуравнении Q = aWb возрастают с температурой в пределах толерантных
ее значений.
При этом величины Q10 составляют 1,5–2,5
Напротив, степенной коэффициент
b с ростом температуры
обычно снижается.
Значения коэффициентов а и b зависимости скорости дыхания (Q,
млО2·экз-1·час-1) от сырой массы тела (W, г) у дальневосточного
трепанга Stichopus japonicus
Температура, оС
a
b
2,5
0,0029
0,745
8,5
0,0070
0,85
13,5
0,025
0,65
18,5
0,11
0,38
23,5
0,12
0,41
28,5
0,32
0,29
4.
Wn > W4 > W3 > W2 > W1Tn > T4 > T3 > T2 > T1
«Веерная модель метаболизма». Линии регрессии уравнений связи
скорости метаболизма и массы тела при разных температурах на двойном
логарифмическом графике сходятся в одной точке.
Отсюда следует:
Термолабильность у более мелких особей выражена сильнее, чем у более
крупных.
У организмов существует некоторый предельный уровень скорости
метаболизма (Rk), который не может быть превышен.
5. Активный обмен у животных
Интенсивность потребления кислорода у всехживых организмов возрастает при любых формах
их
физической
активности,
включая
передвижение (плавание, бег, машущий полет),
добывание пищи, заботу о потомстве и пр.
Выделяют ряд уровней энергетического обмена
особей в зависимости от степени их двигательной
активности.
6.
Основной обмен характеризует минимально возможныйуровень обмена у особей с данной массой тела.
У гомойотермных животных он измеряется в условиях
полного покоя, например, во время сна при температуре
критической точки или в термонейтральной зоне.
Энергия, выделяющаяся при дыхании, в этом случае
расходуется только на функционирование внутренних
органов – дыхательной, пищеварительной, нервной и
других систем организма.
Активный обмен – это уровень обмена особи при
более-менее существенном уровне ее двигательной
активности.
Он является суммой стандартного обмена особи и
затрат ее энергии на передвижение.
7.
Стандартныйобмен
имеет
место
при
минимальной двигательной активности организма.
Поэтому его часто называют обменом покоя.
Большинство данных по скорости обмена у
животных получены при их содержании в различных
экспериментальных установках, т. е. в условиях весьма
ограниченной двигательной активности.
Определение стандартного
обмена у ящериц
8.
Установка для определения активного обмена у рыб9.
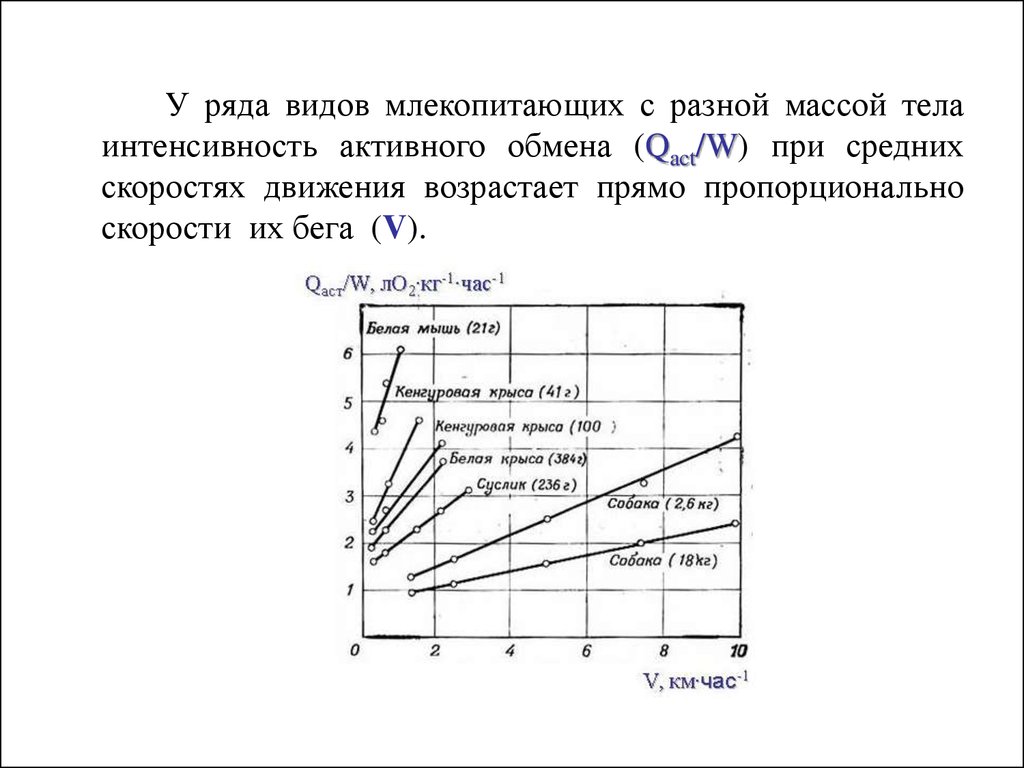
У ряда видов млекопитающих с разной массой телаинтенсивность активного обмена (Qact/W) при средних
скоростях движения возрастает прямо пропорционально
скорости их бега (V).
10.
Зависимость Qact/W от V у млекопитающих вупрощенном виде может быть выражена
уравнением:
Qact/W = f + gV.
Коэффициент f соответствует интенсивности
дыхания особи при V = 0, или интенсивность
стандартного обмена особи с массой W:
f = aWb-1.
Эмпирический коэффициент g равен тангенсу
угла наклона линии регрессии Qact/W по V.
У исследованной группы млекопитающих g
снижается с увеличением массы их тела (W, кг)
согласно уравнению:
g = pW-q
11.
Отсюдаполучаем
обобщенное
уравнение
зависимости интенсивности обмена млекопитающих
от массы их тела и скорости бега:
Qact/W = аWb-1 + (pW-q)·V
12.
Измерение активного обмена у крупных иподвижных копытных млекопитающих и у
летающих птиц, представляет значительные
методические трудности, поскольку требует
сложной и громоздкой аппаратуры, а часто и
вообще невозможно.
В последние годы для этих целей предложен
принципиально новый «метод дважды помеченной
воды».
13.
В органические вещества пищи,потребляемой
животным, вводятся изотопы водорода (дейтерий, 2D) и
кислорода (18O).
В процессе дыхания из органических веществ в
результате их окисления кислородом
образуются
углекислый газ и вода, количество которых возрастает
прямо пропорционально величине энергетических трат
животного.
Доля «тяжелой воды» (2D218O) в общем количестве
«метаболической воды» остается постоянной.
По изменению содержания «тяжелой воды»
в
организме во времени можно рассчитать общее
количество «метаболической воды», а на этой основе и
его энергетические траты.
14.
Рассчитаны уравнения зависимости Qact (ккал·особь-1·сутки-1) отмассы тела (W, кг) для млекопитающих и птиц при среднем уровне их
суточной подвижности
Значения
Qact
включают стандартный обмен, затраты на
терморегуляцию, перемещение в поисках пищи, ее потребление и
переваривание.
Плацентарные млекопитающие:
Qact = 221W0,81.
Птицы из отряда воробьинообразных:
Qact = 376W0,75.
Птицы из остальных отрядов
Qact = 202W0,81,
Уровень активного обмена у гомойотермных животных приблизительно в
2,5–3 раза выше, чем уровень их стандартного обмена.
Это нужно учитывать при оценке функциональной роли гомойотермных
животных в трансформации энергии в биогеоценозах.
15.
Питание организмовРацион любого организма, или количество пищи,
потребленное за единицу времени, в значительной степени
определяется массой его тела.
Если количество корма в среде находится с избытком,
рацион животных возрастает с увеличением массы их тела.
Однако увеличение рациона, как и скорости дыхания
животных, происходит в значительно меньшей степени, чем
массы тела.
Рацион организмов (R) связан с массой их тела (W)
степенным уравнением:
R = pWq,
Значения степенного коэффициента q изменяются от 0,55
до 0,95, но обычно находятся в пределах 0,60 – 0,80.
16.
Примером этого является кубинскийкраб Menippe mercenaria. Этот важный
промысловый вид является падальщиком и
питается, преимущественно, остатками
погибших животных.
17.
1,08
7
5
lgR
Рацион, R
0,8
6
0,6
4
0,4
3
2
1
0,2
0
100
200
300
400
500
1,6
М асса те ла, W
1,8
2,0
2,2
2,4
2,6
2,8
lgW
Зависимость рациона (R, г сырой массы корма·особь-1·сутки-1)
от сырой массы тела (W, г) у Menippe mercenaria.
Уравнение данной зависимости имеет вид:
R = 0,0746W0,80
18.
Допустим, соотношение между сухой и сырой массой(y) у пищевых
объектов, потребляемых особями и
калорийность их сухой массы (c) достаточно постоянны.
Тогда уравнение связи между R и W может быть
представлено в виде:
R = ycpWq
Поскольку ycp = n = const, полученное уравнение
можно упростить:
R = nWq
Это уравнение
представляет
зависимость
энергетического эквивалента потребленной организмом
пищи от массы его тела.
19.
Зависимость рациона от концентрации корма в средеУ всех видов животных, особенно у водных
беспозвоночных с фильтрационным питанием (например, у
жаброногих и ветвистоусых раков), величина рациона в
значительной степени зависит от концентрации корма в среде.
Как правило, при низких концентрациях корма рацион
особей возрастает пропорционально их увеличению.
Однако при дальнейшем повышении концентрации корма
увеличение рациона все более замедляется, а выше
определенного предела он стабилизируется на определенном
уровне.
Зависимость между концентрацией дрожжевых
клеток в воде (К, млн. клеток·мл-1) и рационом
(R, млн. клеток·особь-1час-1) у Artemia salina. 1 –
молодь; 2 – взрослые особи.
Рацион (R)
1,25
1,00
0,75
0,50
0,25
0,00
0,00
0,25
0,50
0,75
1,00
1,25
Концентрация корма (К)
1,50
1
2
20.
Полученнаязависимость
количества
потребленного корма (R) от его концентрации в
среде (К) может быть описана асимптотическим
уравнением В.С.Ивлева:
R = Rmax(1 – e-λK).
где:
Rmax – максимальный рацион особей, его определяют
эпирическим путем.
λ - константа
21.
Для молоди A.salina это уравнение имеет вид:R = 0,42(1 – e-1,566K),
а для взрослых рачков:
R = 1,24(1 – e-1,267K),
где:
R – рацион (млн. клеток·особь-1час-1),
К – концентрация корма (млн.клеток·мл-1).
По этим уравнениям можно рассчитать объем
воды
(V),
очищаемый
фильтраторами
от
водорослей:
V = R/K.
22.
Кривая уравнения зависимости R от K проходит черезначало координат, что предполагает, что животные
способны потреблять корм даже при самом низком его
содержании в среде.
Это в целом справедливо для
большинства
растительноядных видов животных, а также хищников,
падальщиков и детритофагов. Низкое количество корма в
среде такие виды компенсируют увеличением затрат
времени и энергии на его поиски, т.е., повышением
активного обмена.
Однако многие виды с фильтрационным типом
питания, например, ветвистоусые раки, если концентрации
корма (планктонные одноклеточные водоросли) падает
ниже определенного предела, вообще не способны его
улавливать.
Обычно чем грубее их фильтрационный аппарат, тем
при более высоких концентрациях водорослей они теряют
способность их потреблять.
23.
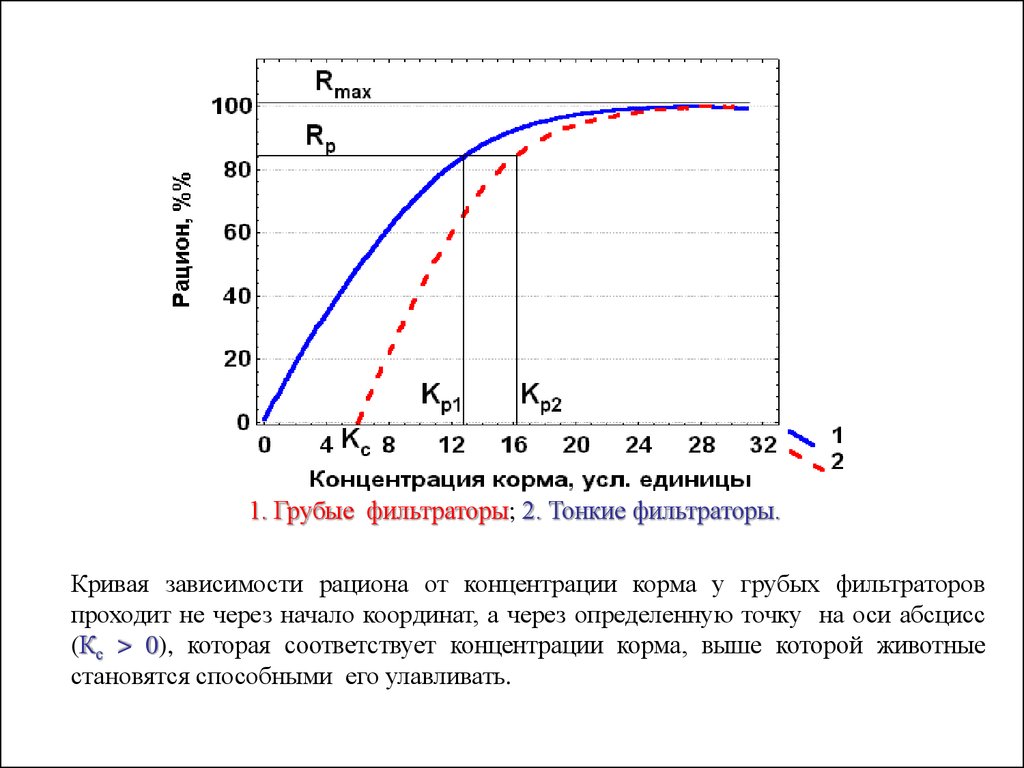
Исходя из строения фильрационного аппарата, ветвистоусые ракиподразделяются на «тонких» и «грубых» фильтраторов.
У первых щетинки на антеннах расположены достаточно густо, что
позволяет им улавливать пищевые частички при их низкой концентрации в
среде.
У вторых щетинки расположены разреженно, в результате чего многие
частички корма свободно проходят через них.
Тонкий фильтратор - дафния
Грубый фильтратор – сида
24.
Уравнение для «грубых фильтраторов» можетбыть описано уравнением Ивлева – Винберга –
Анисимова:
R = Rmax[1 – e-λ(K – Kс)].
где: Кс – минимальная концентрация корма в среде,
при которой животные способны питаться.
25.
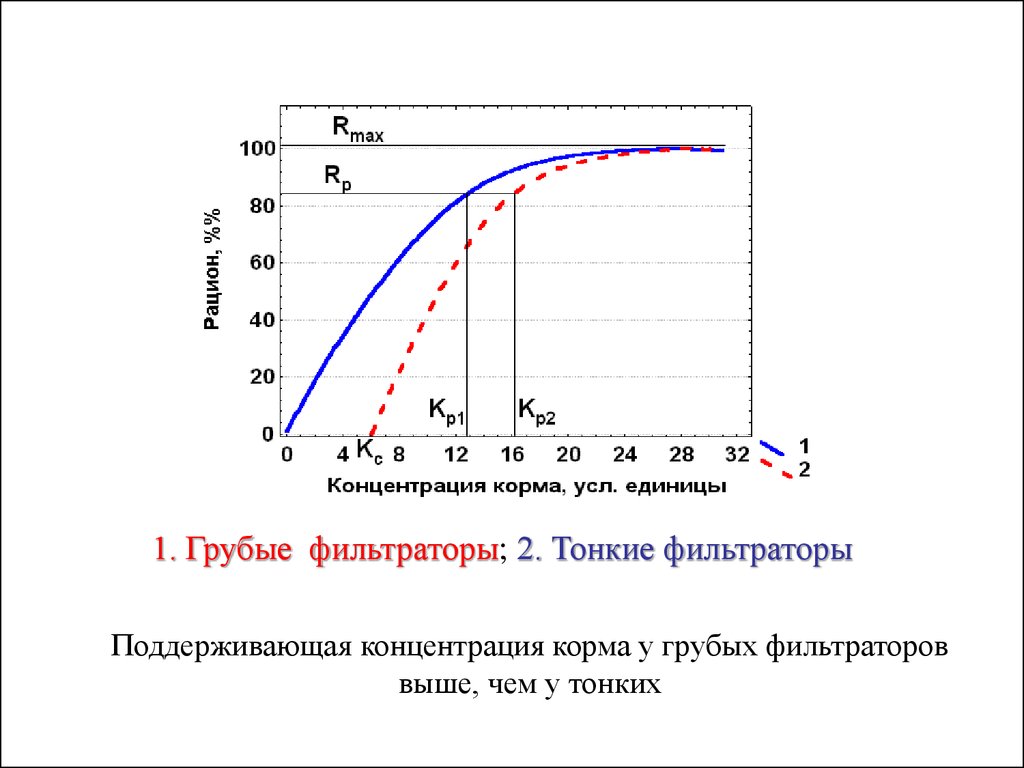
1. Грубые фильтраторы; 2. Тонкие фильтраторы.Кривая зависимости рациона от концентрации корма у грубых фильтраторов
проходит не через начало координат, а через определенную точку на оси абсцисс
(Кс > 0), которая соответствует концентрации корма, выше которой животные
становятся способными его улавливать.
26.
Баланс энергии на организменном уровнеЭкологические системы существуют за счет
постоянного потока и трансформации вещества и энергии.
Основными элементами экологических систем
являются организмы.
Поэтому особенности трансформации вещества и
энергии в экологических системах в значительной
степени
определяются
закономерностями этих
процессов на организменном уровне.
Баланс энергии у гетеротрофных организмов можно
схематично представить следующим образом.
Потребленная пища, или рацион частично усваивается
организмом (усвоенная, или ассимилированная часть
рациона), а частично выделяется наружу
в виде
непереваренных остатков (неусвоенная часть рациона).
27.
Усвоение пищи заключается в разложении еесложных органических
веществ (белков,
углеводов, жиров) до сравнительно простых
мономеров - аминокислот, моносахаридов и т.д.
Одна их часть в процессе дыхания разлагается
далее – до углекислого газа и воды с
образованием АТФ.
Процессы обмена веществ, ведущие к
высвобождению энергии пищи в организме,
имеют общее название энергетический обмен.
28.
Другая часть мономеров используется дляобразования новых жиров, белков и углеводов,
типичных для данного организма.
Они
затем используются на построение
тканей и органов, т.е. на увеличение массы тела
(прирост).
Процессы образования в организме сложных
органических веществ из более простых
называется пластическим обменом.
29.
Процессы пластического обмена могутпроисходить только с участием АТФ. Поэтому оба
типа обмена в организме тесно взаимосвязаны.
При
синтезе белка в рибосомах на
присоединении одной молекулы аминокислоты к
строящейся полипептидной цепи затрачивается
энергия одной макроэргической связи АТФ.
При окислении одной молекулы глюкозы
образуется 38 молекул АТФ, этого количества
достаточно для построения полипептидной цепи,
состоящей из 39 аминокислот.
Поэтому интенсивно растущие организмы
потребляют много кислорода.
30.
Согласно первому закону термодинамики, в любойсистеме между потреблением и расходом энергии
существует равенство, т.е. соблюдается энергетический
баланс.
Энергетический баланс организма можно представить в
виде формулы:
R = P + T + H,
где:
R – энергия потребленной пищи;
Р – энергия, затраченная на прирост (увеличение)
массы тела особи;
Т – энергия, затраченная на дыхание;
Н – энергия неусвоенной части рациона. Она была
потреблена организмом с пищей, но не усвоена им и
выведена во внешнюю среду с продуктами распада.
31.
Соотношения между компонентами энергетического балансаА = P + T – усвоенная часть рациона.
U-1 = А/R – усвояемость пищи.
При потреблении животными растительной пищи U-1 в
среднем равно 0,5, а при потреблении ими животной пищи
– в среднем равно 0,8.
Значение U-1 у бактерий приближается к 1,00, поскольку
у них не образуются фекалии.
Если принять Р = 0, то Т = RU-1 = A.
Рацион организма, усвоенная часть которого равна его
затратам
энергии
на
дыхание,
называется
поддерживающим рационом (Rp).
Очевидно:
Rp = T/U-1.
32.
У животных с фильтрационным типомпитания концентрация корма, при которой
рацион
особей
становится
равным
поддерживающему
(Rp),
называется
поддерживающей концентрацией (Кс).
Если два вида-фильтратора
имеют
одинаковый поддерживающий рацион, то
поддерживающая концентрация корма (Кр)
будет ниже для вида с более совершенным
фильтрационным аппаратом
33.
1. Грубые фильтраторы; 2. Тонкие фильтраторыПоддерживающая концентрация корма у грубых фильтраторов
выше, чем у тонких
34.
К1 = P/R – валовая эффективностьроста,
или коэффициент экологической
эффективности роста первого порядка.
К2 = P/А – чистая эффективность
роста, или коэффициент эффективности
роста второго порядка.
К1 = К2 ·U-1
35.
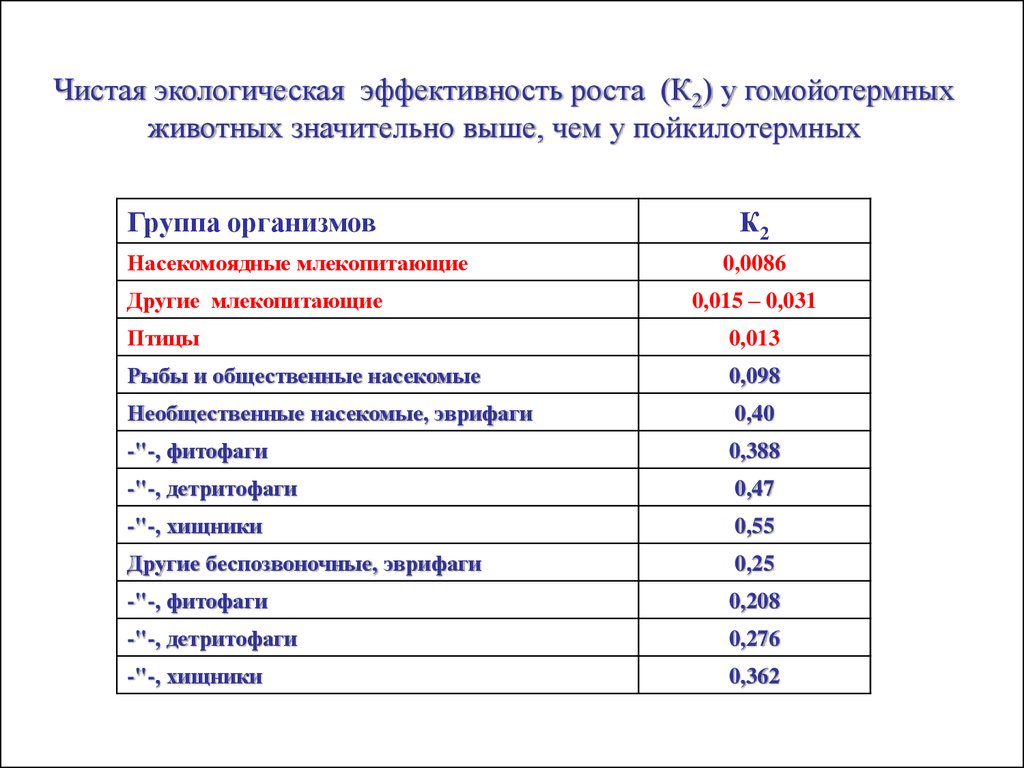
Чистая экологическая эффективность роста (К2) у гомойотермныхживотных значительно выше, чем у пойкилотермных
Группа организмов
Насекомоядные млекопитающие
Другие млекопитающие
К2
0,0086
0,015 – 0,031
Птицы
0,013
Рыбы и общественные насекомые
0,098
Необщественные насекомые, эврифаги
0,40
-"-, фитофаги
0,388
-"-, детритофаги
0,47
-"-, хищники
0,55
Другие беспозвоночные, эврифаги
0,25
-"-, фитофаги
0,208
-"-, детритофаги
0,276
-"-, хищники
0,362
36.
В практике рыбоводства и животноводства широкоиспользуется термин «кормовой коэффициент (КК)», или
отношение прироста массы тела организма к массе
потребленного им корма, т.е.:
КК = R/P = 1/K1.
Иногда рыбоводы утверждают, что у рыб, питающихся
комбикормом, КК может быть меньше единицы. Это
означает, что прирост массы их тела может превышать
массу съеденного ими корма, тогда К1 > 1,00.
Однако это обусловлено тем, что при расчетах прирост
рыб выражен в сырой массе, а комбикорм – в сухой.
При пересчете значений массы на их энергетические
единицы оказывается, что KK всегда больше единицы.