































































































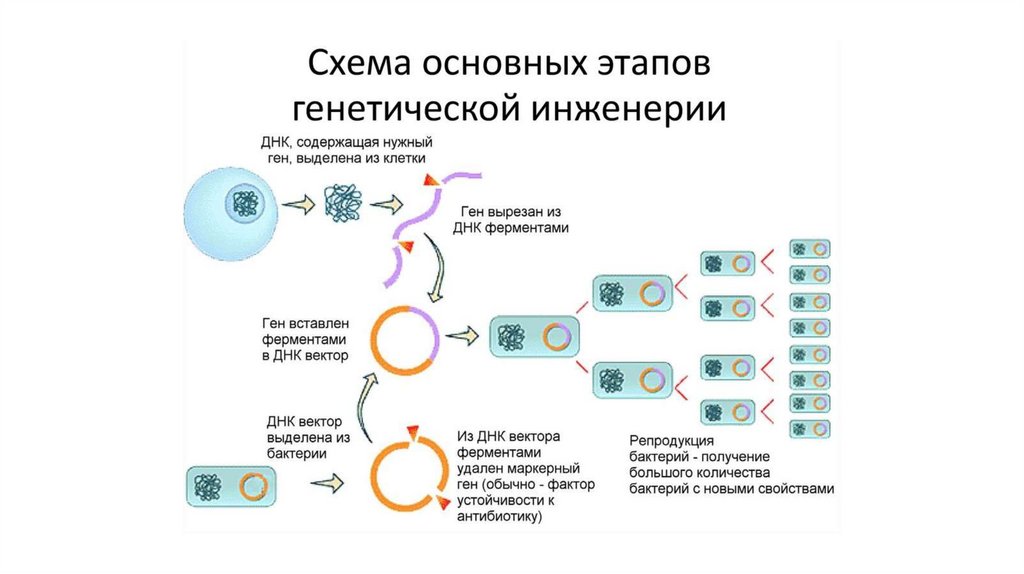













 Биология
БиологияПохожие презентации:










Особенности организации клеток эукариот, бактерий и архей
1.
Особенности организацииклеток эукариот, бактерий и
архей.
2.
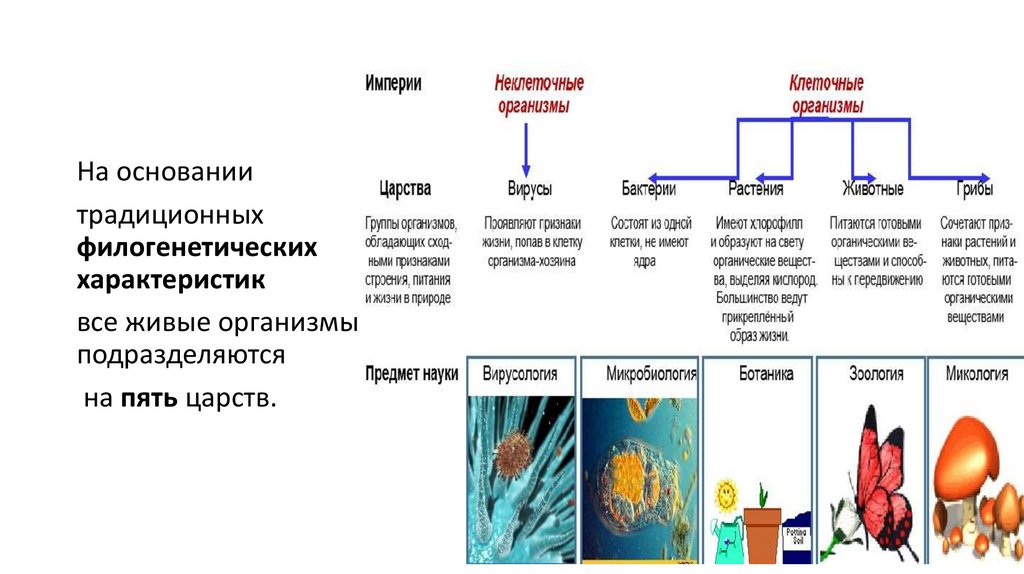
На основаниитрадиционных
филогенетических
характеристик
все живые организмы
подразделяются
на пять царств.
3.
4.
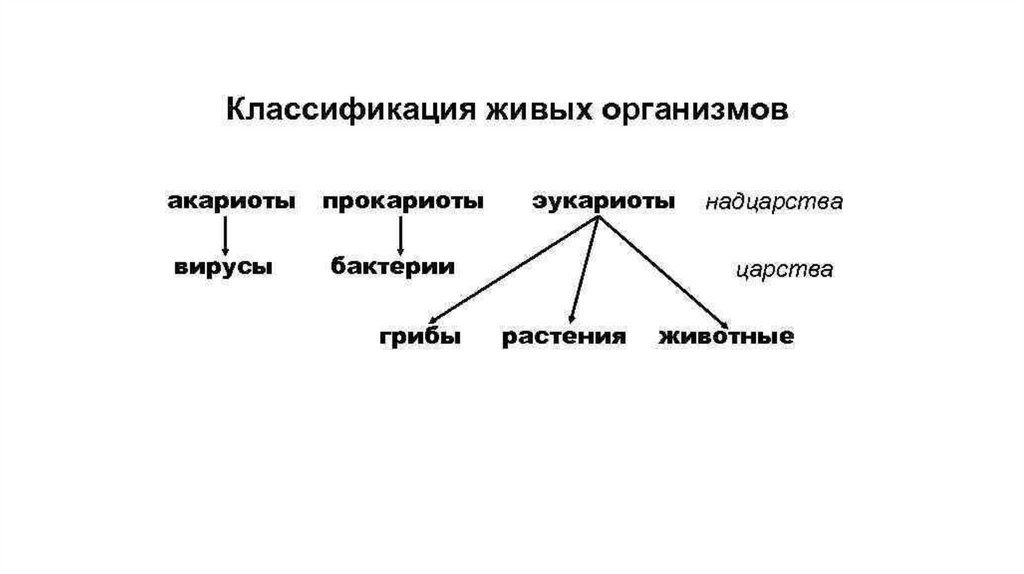
• Современные молекулярногенетические методыподразделяют клеточные
формы жизни лишь на 3
домена – Бактерии (или
Эубактерии), Археи( или
Архебактерии) и Эукариоты.
(Карл Везе и сотр.)
• После́дний универса́льный
о́бщий пре́док (англ. last
universal common ancestor,
LUCA
5.
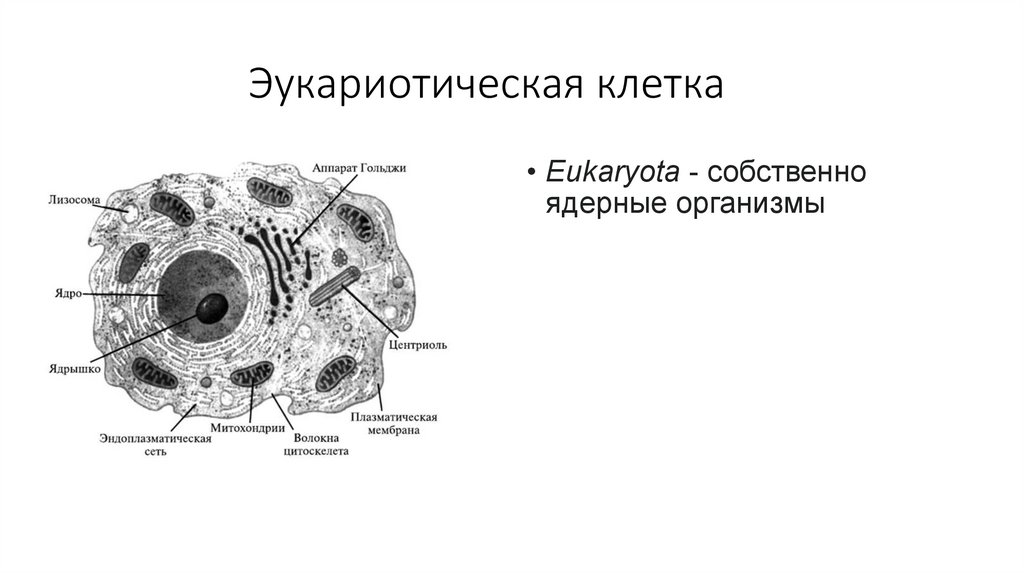
Эукариотическая клетка• Eukaryota - собственно
ядерные организмы
6.
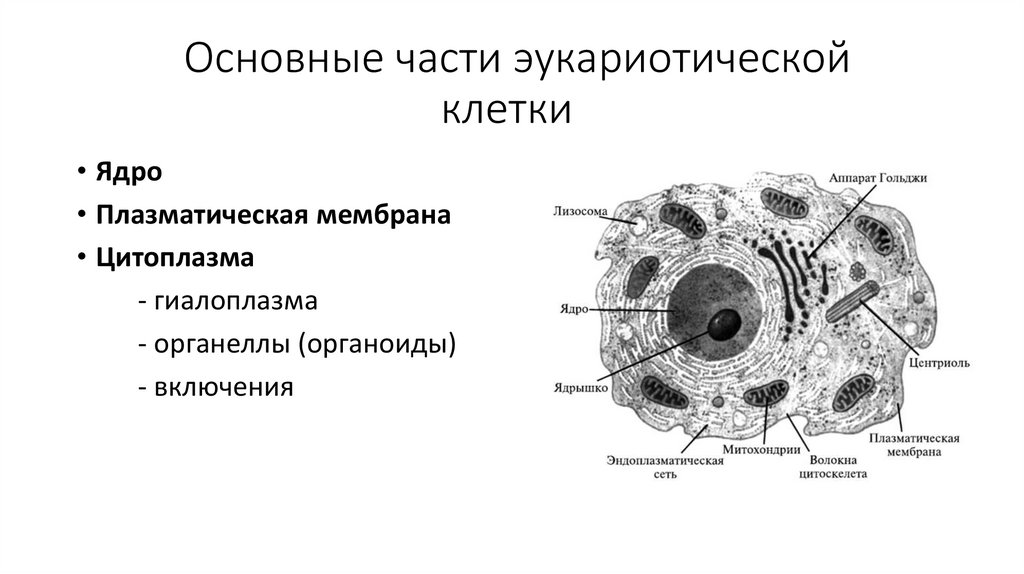
Строение клетки7.
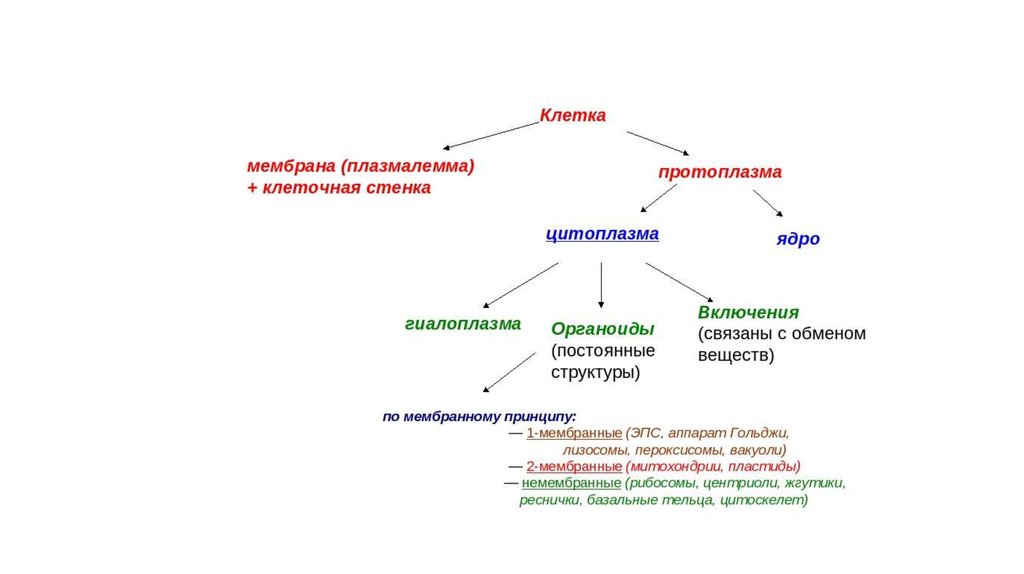
Основные части эукариотическойклетки
• Ядро
• Плазматическая мембрана
• Цитоплазма
- гиалоплазма
- органеллы (органоиды)
- включения
8.
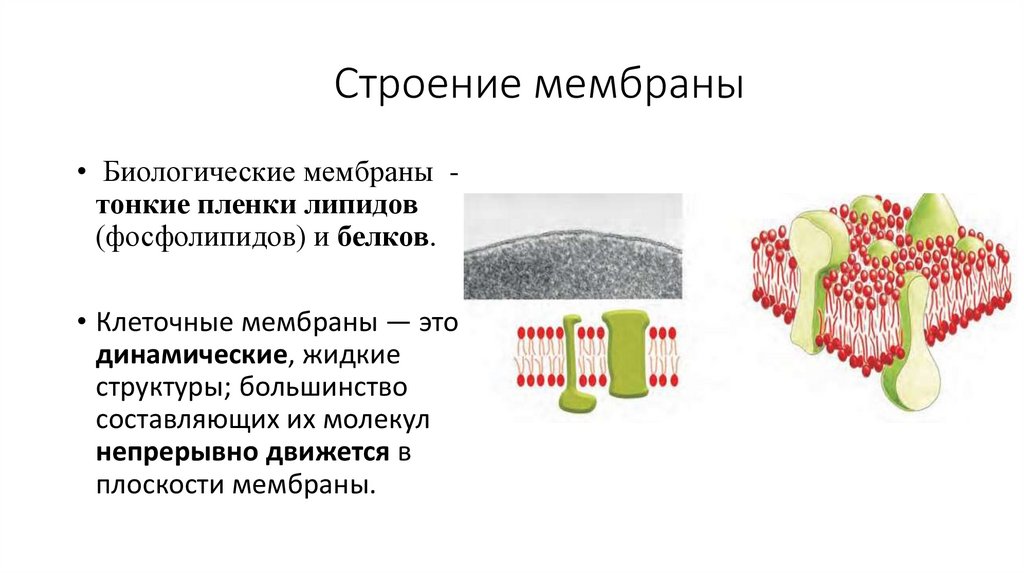
Строение мембраны• Биологические мембраны тонкие пленки липидов
(фосфолипидов) и белков.
• Клеточные мембраны — это
динамические, жидкие
структуры; большинство
составляющих их молекул
непрерывно движется в
плоскости мембраны.
9.
• Молекулы липидов организованы в непрерывный двойной слойтолщиной около 5 нм.
• Липидный бислой служит относительно непроницаемым
барьером для большинства растворимых в воде молекул.
Свойство полупроницаемости мембраны
10.
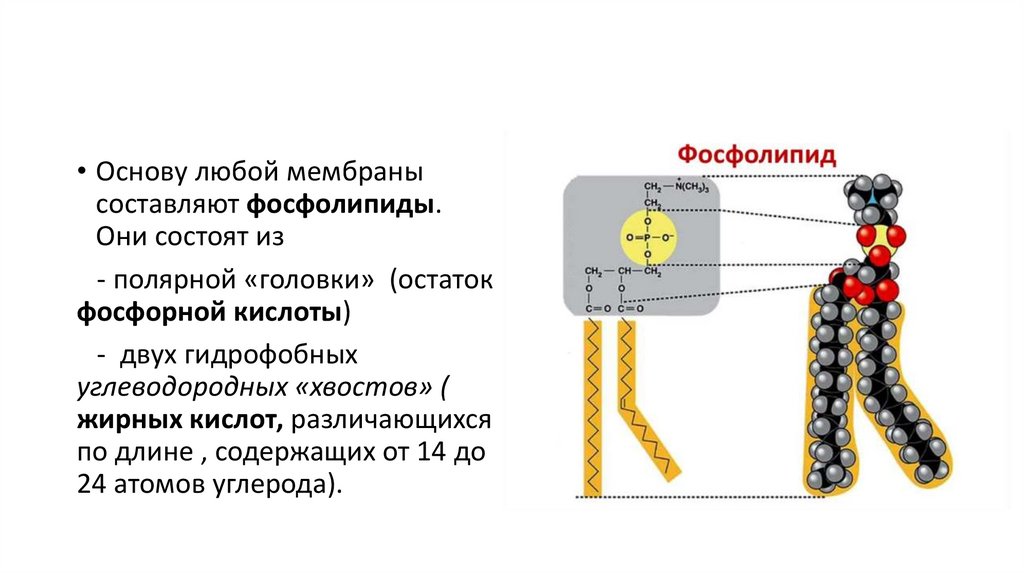
• Основу любой мембранысоставляют фосфолипиды.
Они состоят из
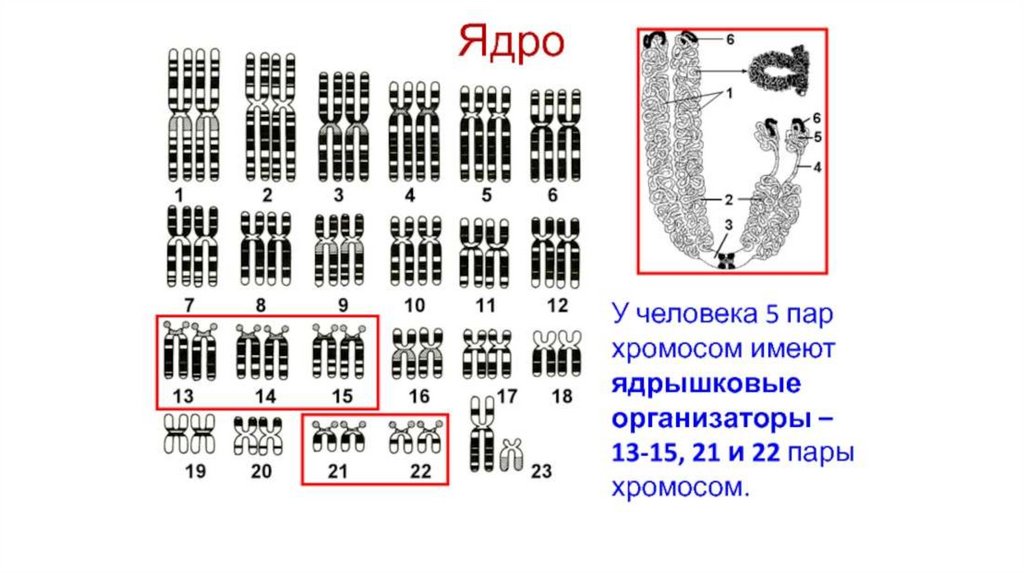
- полярной «головки» (остаток
фосфорной кислоты)
- двух гидрофобных
углеводородных «хвостов» (
жирных кислот, различающихся
по длине , содержащих от 14 до
24 атомов углерода).
11.
• Липидные молекулы самопроизвольноагрегируют в водном растворе, пряча
свои гидрофобные углеводородные
«хвосты» внутрь и выставляя
гидрофильные «головки» наружу. При
этом возможны 2 варианта
1) формируются сферические
мицеллы, в которых «хвосты»
расположены внутри шара
2) образуются двухслойные
пленки, или бислои, в которых
гидрофобные «хвосты» расположены
между гидрофильными «головками»
12.
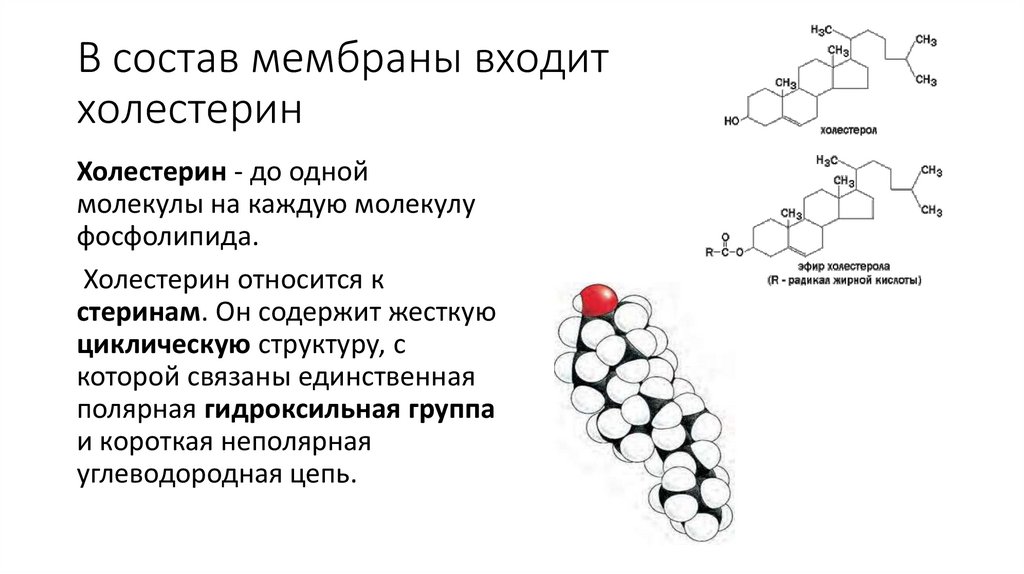
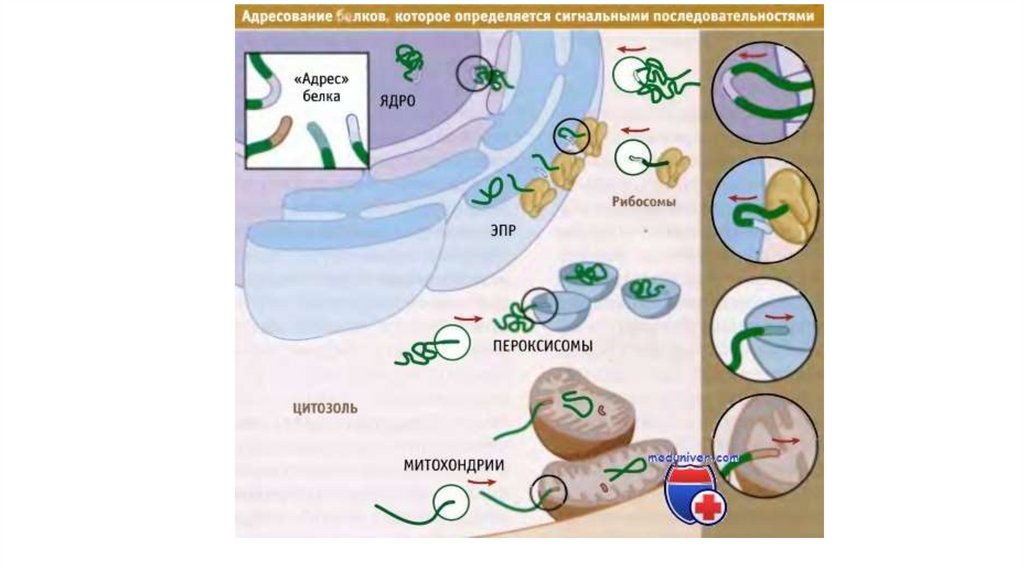
В состав мембраны входитхолестерин
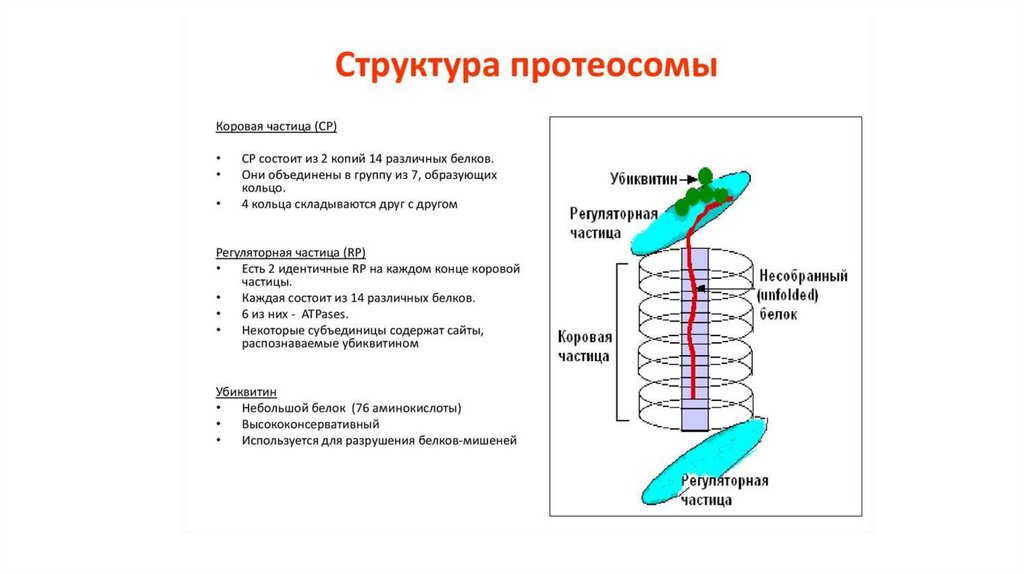
Холестерин - до одной
молекулы на каждую молекулу
фосфолипида.
Холестерин относится к
стеринам. Он содержит жесткую
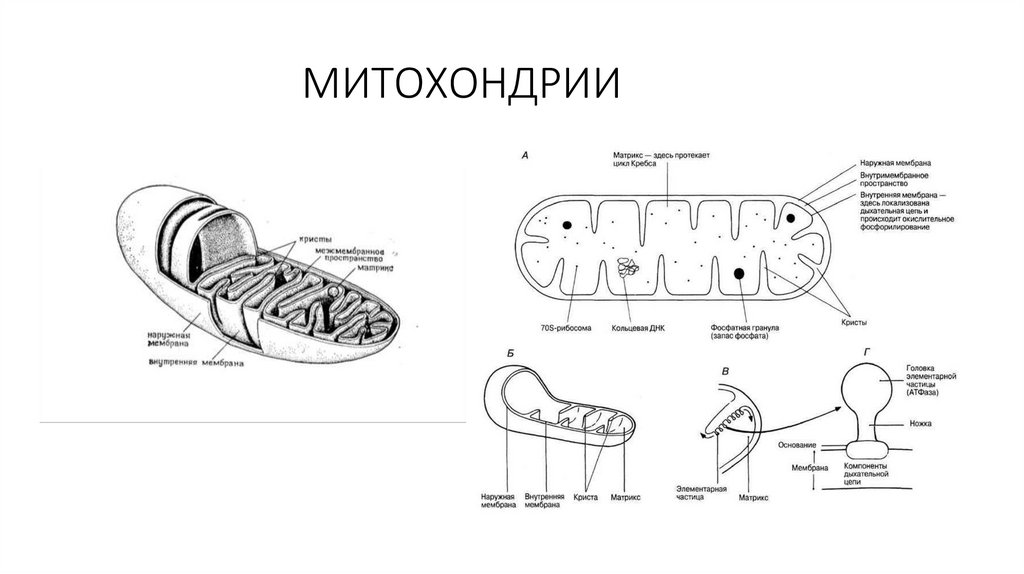
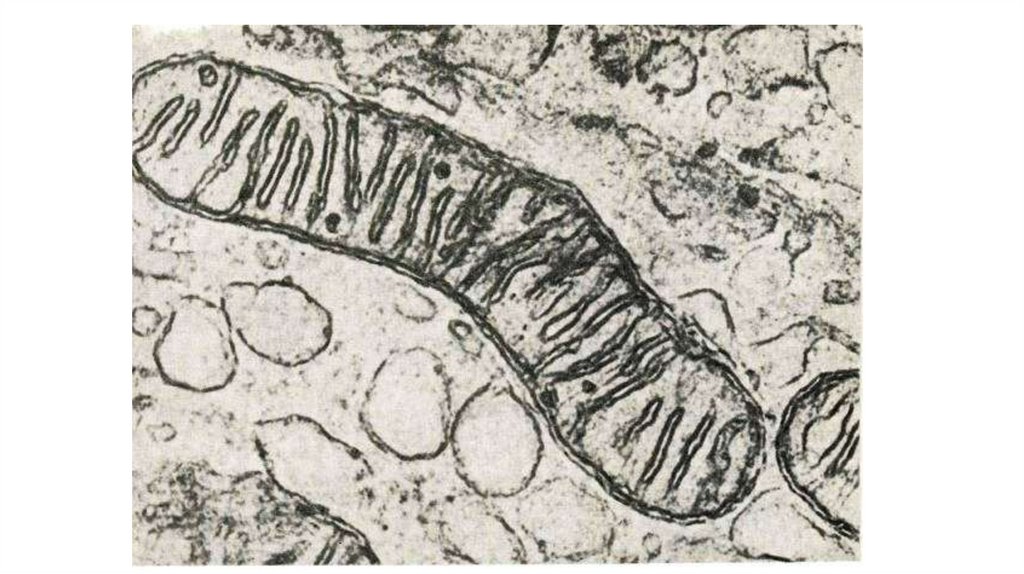
циклическую структуру, с
которой связаны единственная
полярная гидроксильная группа
и короткая неполярная
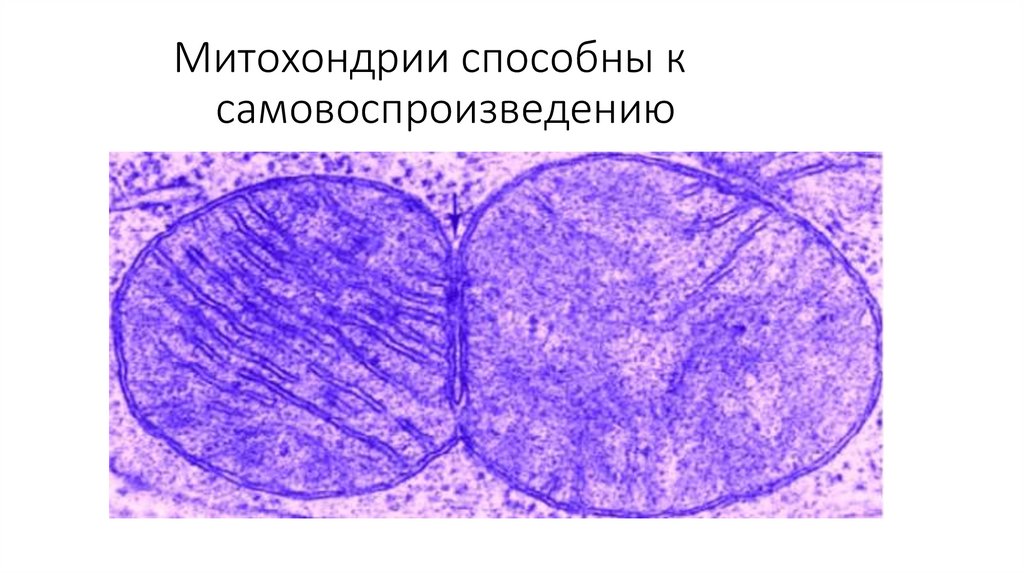
углеводородная цепь.
13.
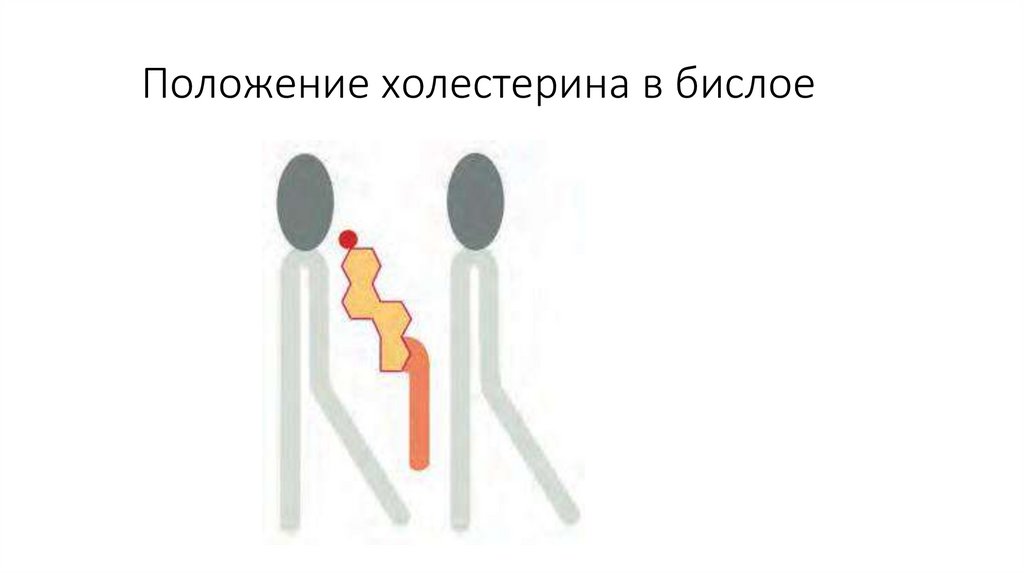
Положение холестерина в бислое14.
Функции холестерина• Холестерин регулирует свойства липидных бислоев:
- Снижает подвижность углеводородных цепей молекул
фосфолипидов,
- делает липидный бислой в этой области менее деформируемым
- уменьшает проницаемость бислоя для малых растворимых в воде
молекул.
Холестерин уплотняет упаковку липидов в бислое.
15.
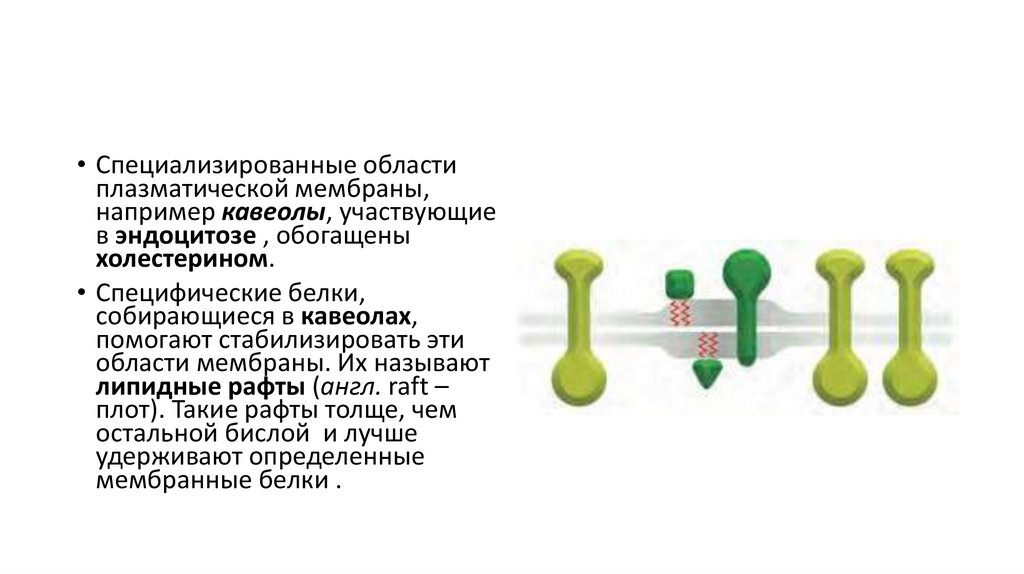
• Специализированные областиплазматической мембраны,
например кавеолы, участвующие
в эндоцитозе , обогащены
холестерином.
• Специфические белки,
собирающиеся в кавеолах,
помогают стабилизировать эти
области мембраны. Их называют
липидные рафты (англ. raft –
плот). Такие рафты толще, чем
остальной бислой и лучше
удерживают определенные
мембранные белки .
16.
• Для изучения мембраниспользовали два типа
препаратов :
• (1) бислои в форме
сферических везикул,
называющихся липосомами,
диаметр которых изменяется
от 25 нм до 1 мкм ;
17.
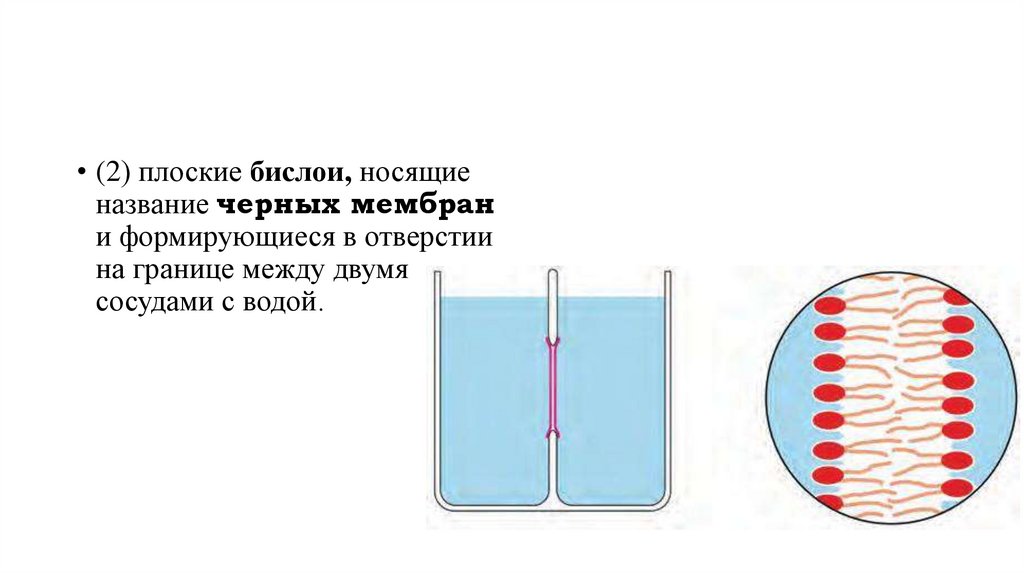
• (2) плоские бислои, носящиеназвание черных мембран
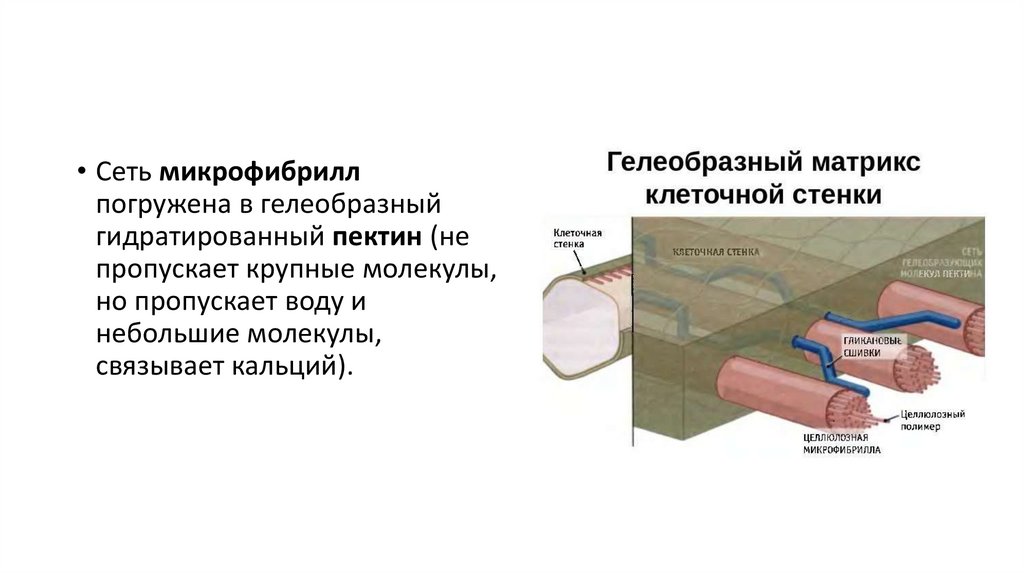
и формирующиеся в отверстии
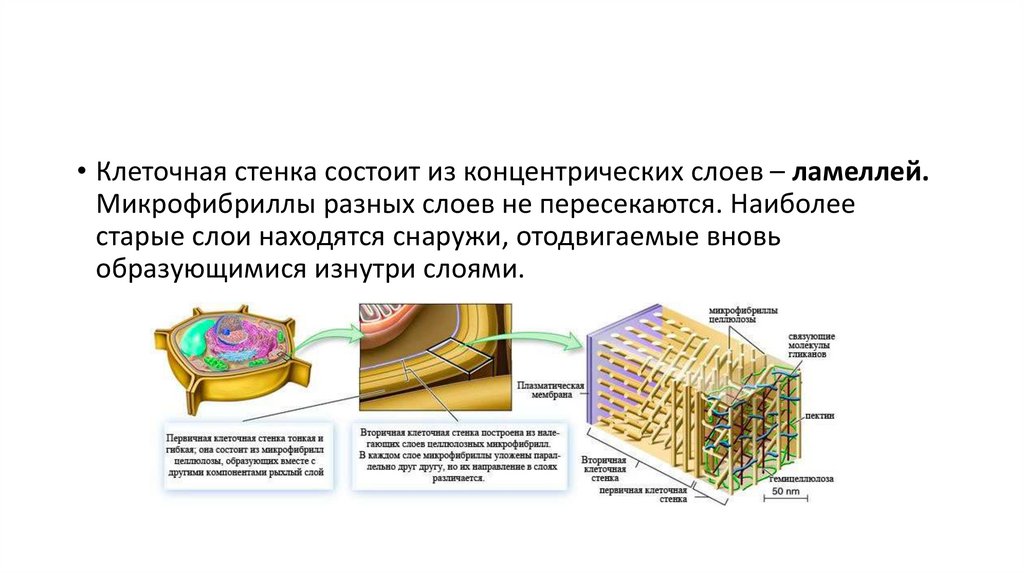
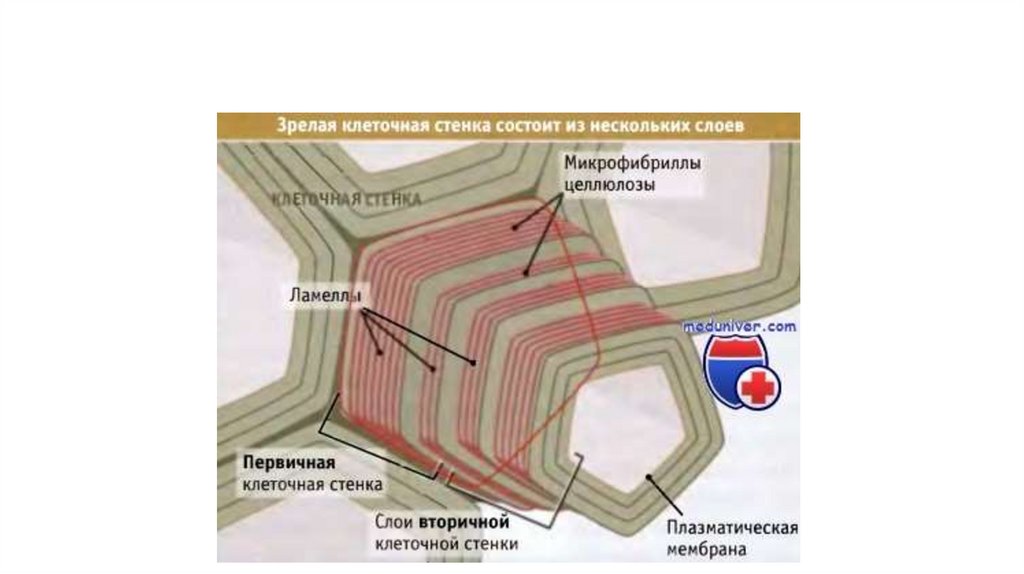
на границе между двумя
сосудами с водой.
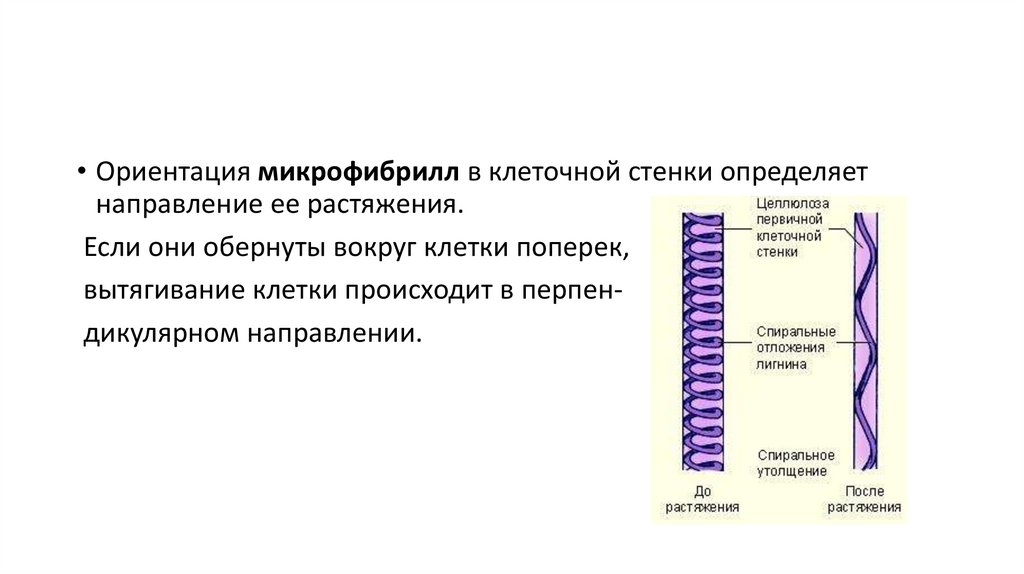
18.
ГликолипидыК липидам наружного монослоя в мембранах животных клеток
присоединены олигосахариды.
Углеводные группы к липидным молекулам добавляются в
аппарате Гольджи.
Они играют важную роль во взаимодействии клетки с ее
окружением.
19.
• Гликолипиды содержатся во всех плазматических мембранахживотных клеток (составляют примерно 5 % всех липидных
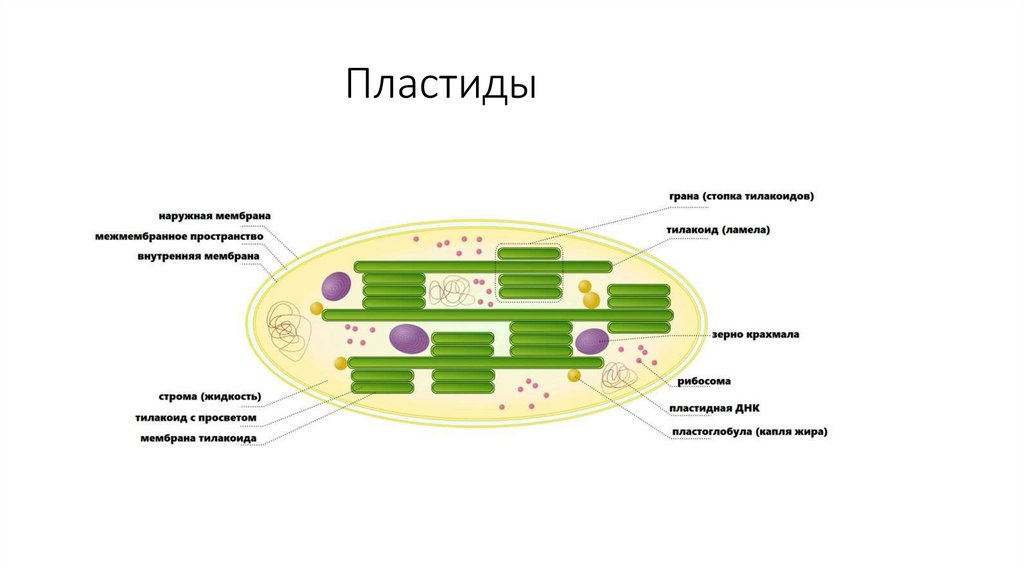
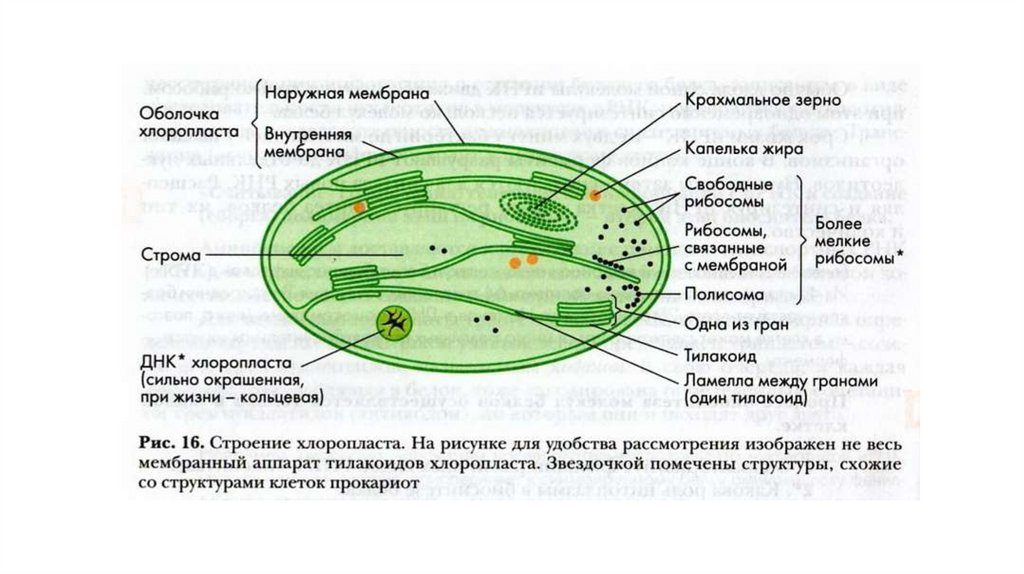
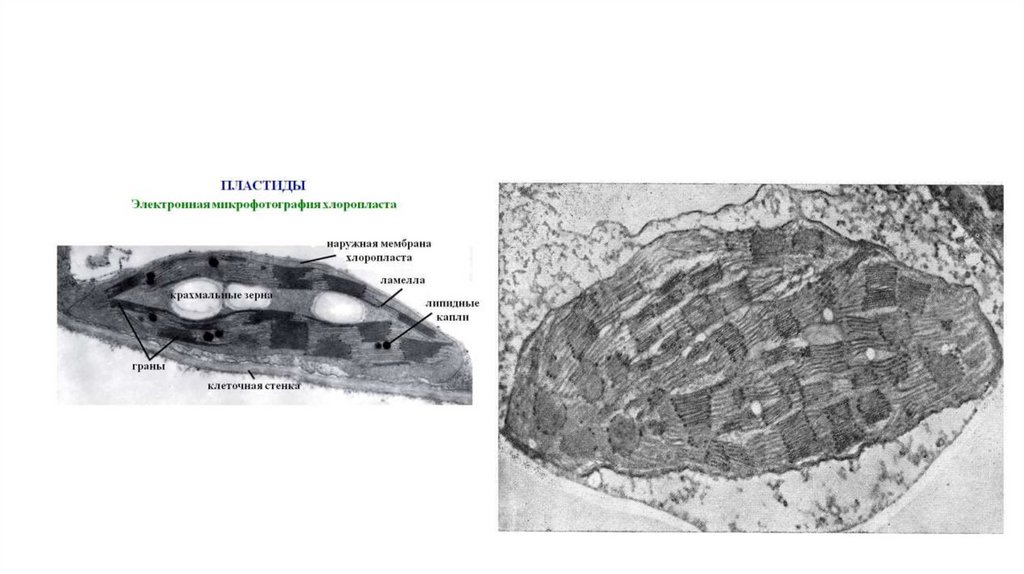
молекул внешнего монослоя).
• Наиболее сложные гликолипиды - ганглиозиды. Больше всего
ганглиозидов содержится в плазматических мембранах нервных
клеток, где они составляют 5–10% всей массы липидов;
20.
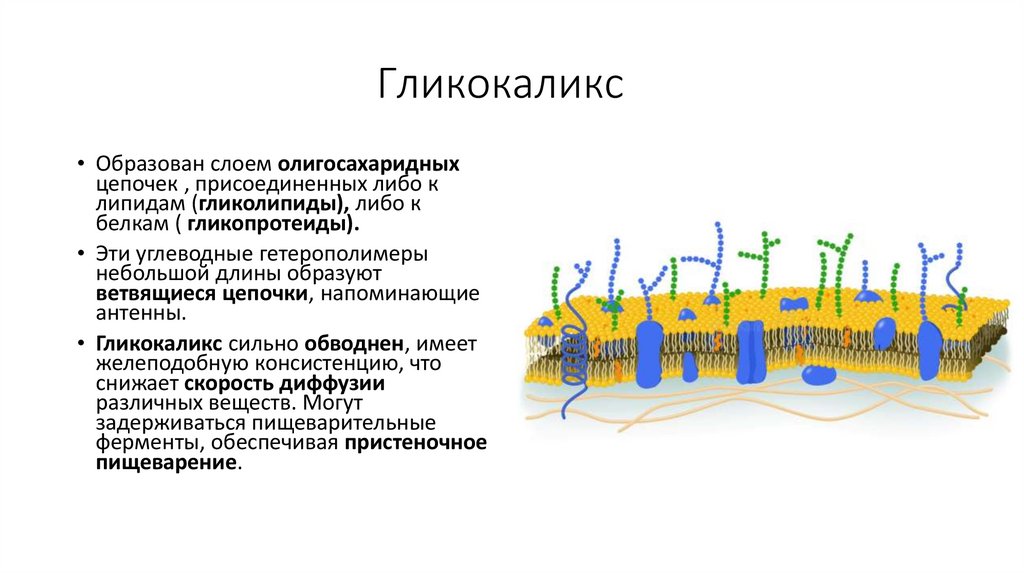
Гликокаликс• Образован слоем олигосахаридных
цепочек , присоединенных либо к
липидам (гликолипиды), либо к
белкам ( гликопротеиды).
• Эти углеводные гетерополимеры
небольшой длины образуют
ветвящиеся цепочки, напоминающие
антенны.
• Гликокаликс сильно обводнен, имеет
желеподобную консистенцию, что
снижает скорость диффузии
различных веществ. Могут
задерживаться пищеварительные
ферменты, обеспечивая пристеночное
пищеварение.
21.
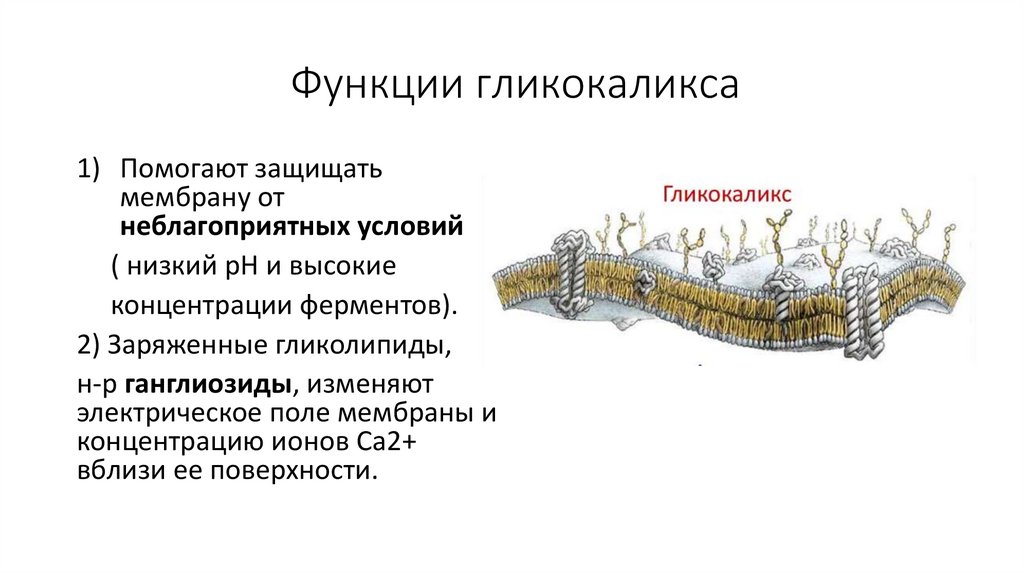
Функции гликокаликса1) Помогают защищать
мембрану от
неблагоприятных условий
( низкий pH и высокие
концентрации ферментов).
2) Заряженные гликолипиды,
н-р ганглиозиды, изменяют
электрическое поле мембраны и
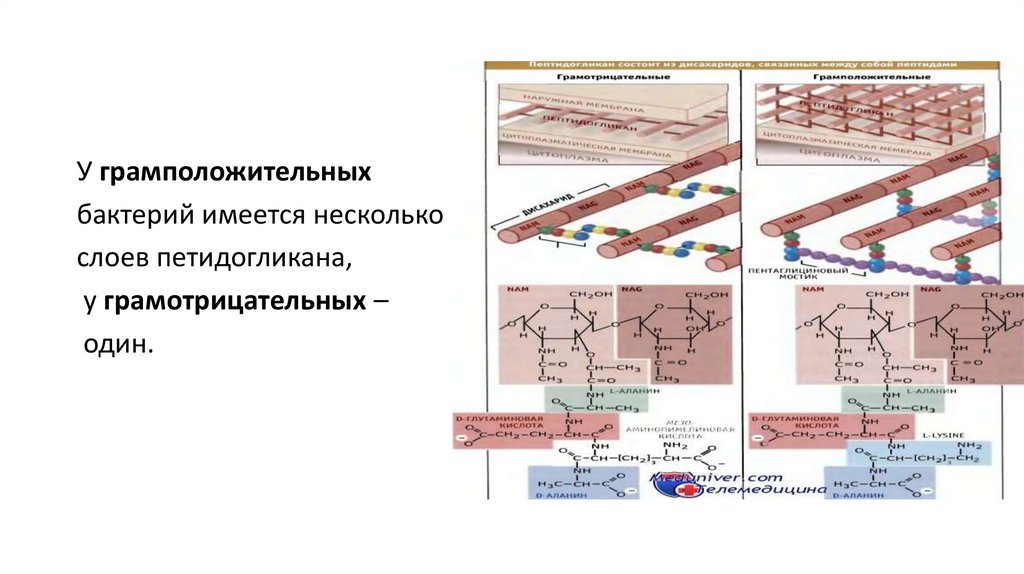
концентрацию ионов Ca2+
вблизи ее поверхности.
22.
3) гликолипиды участвуют в процессе клеточного узнавания приадгезии клеток .
4) Участвуют в обеспечении транспорта через мембрану клетки.
5) Некоторые гликолипиды позволяют бактериальным токсинам
проникнуть в клетку.
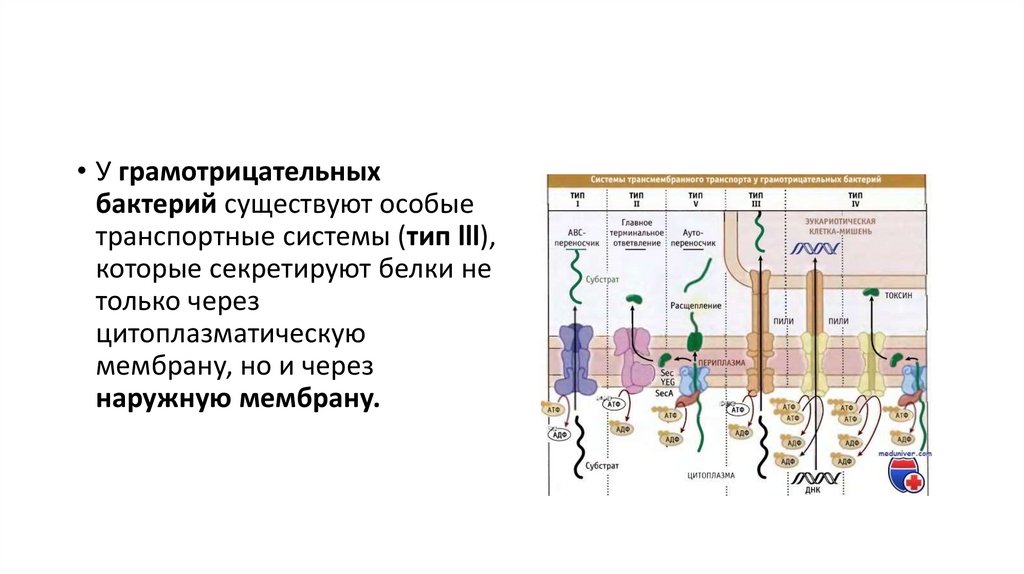
Н-р, холерный токсин связывается с теми клетками (и проникает
только в них), которые несут на своей поверхности определенные
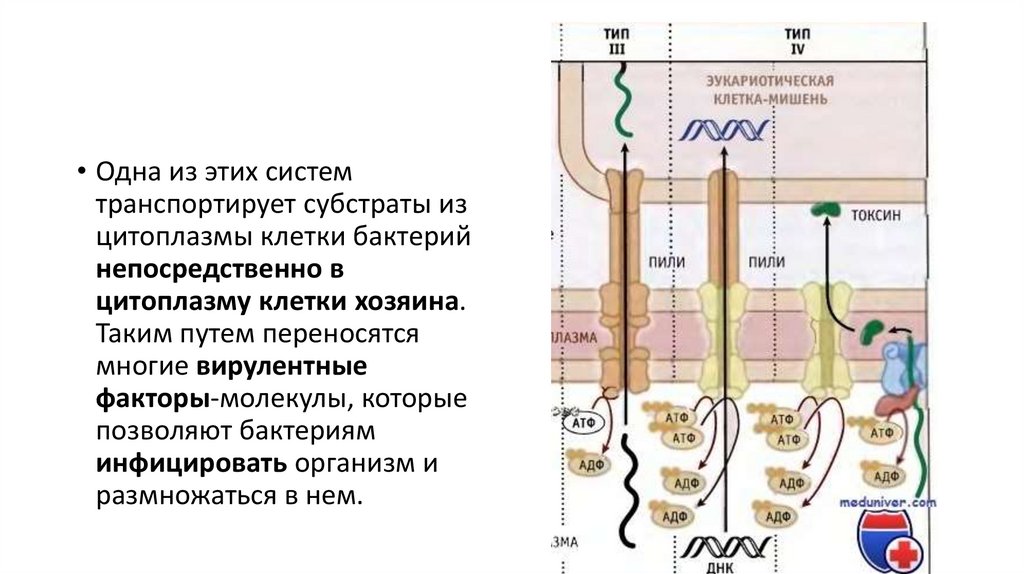
гликолипиды, н-р, с эпителиальными клетками желудочнокишечного тракта.
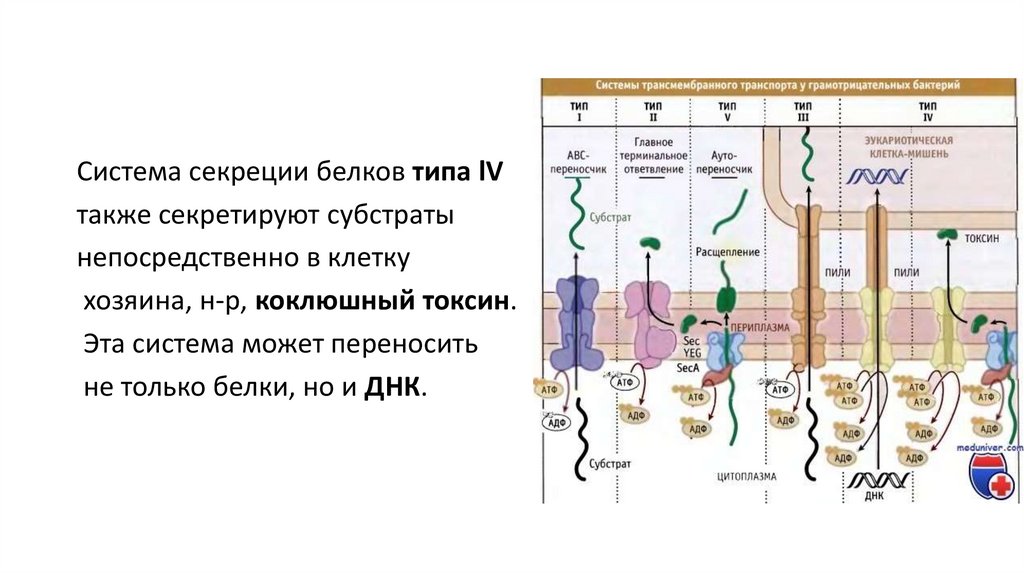
23.
Мембранные белки• Мембранные белки
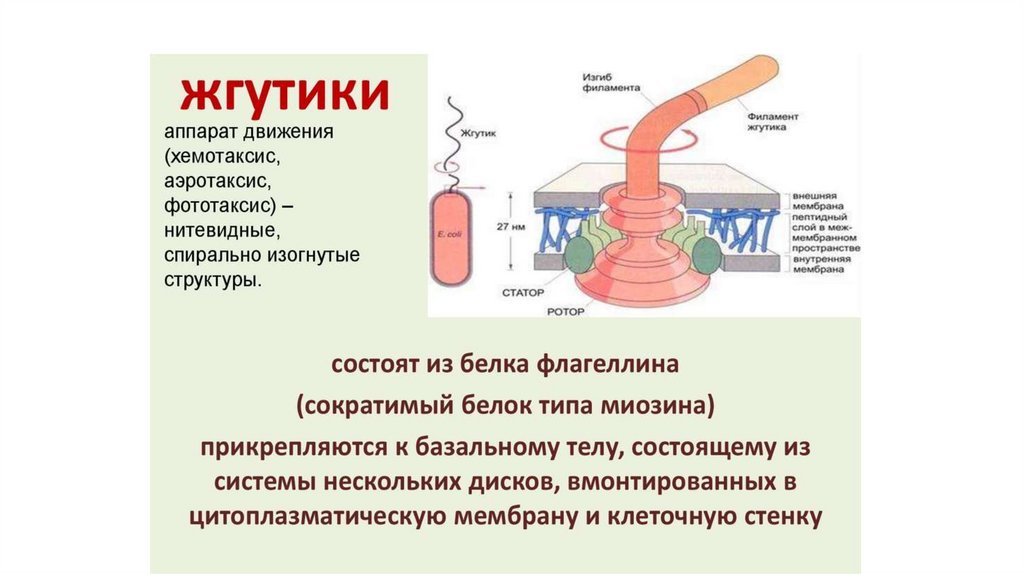
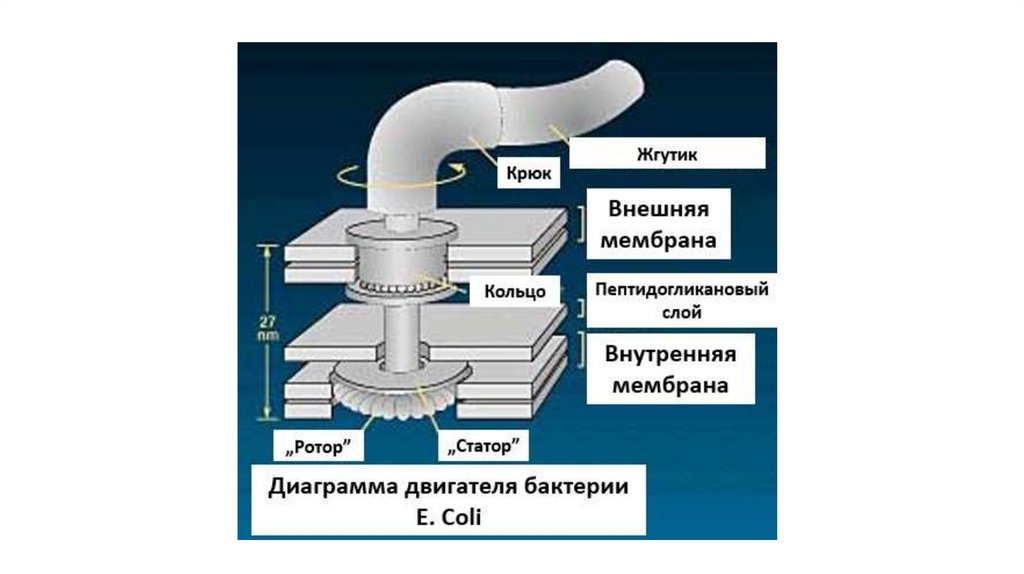
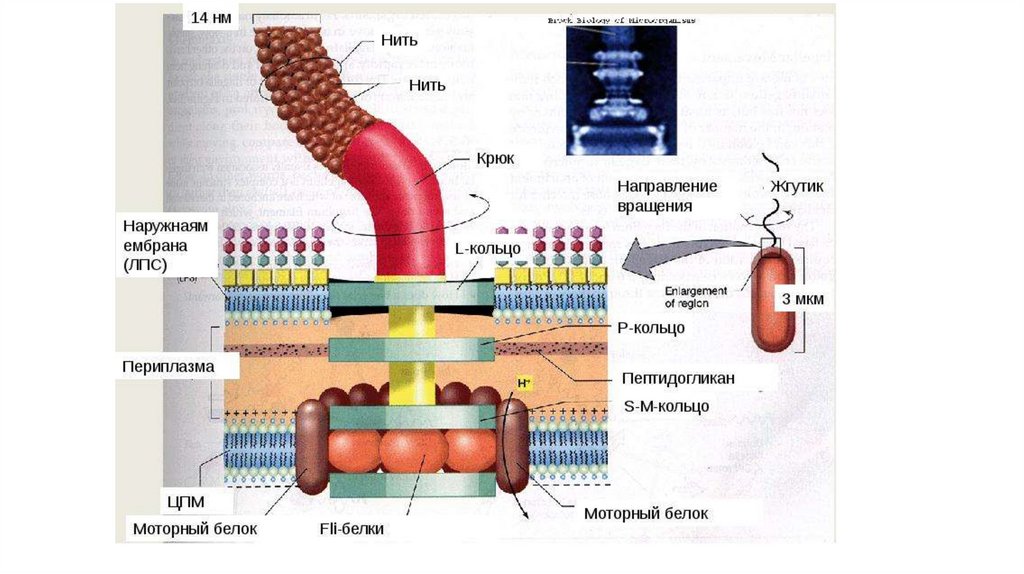
выполняют большинство
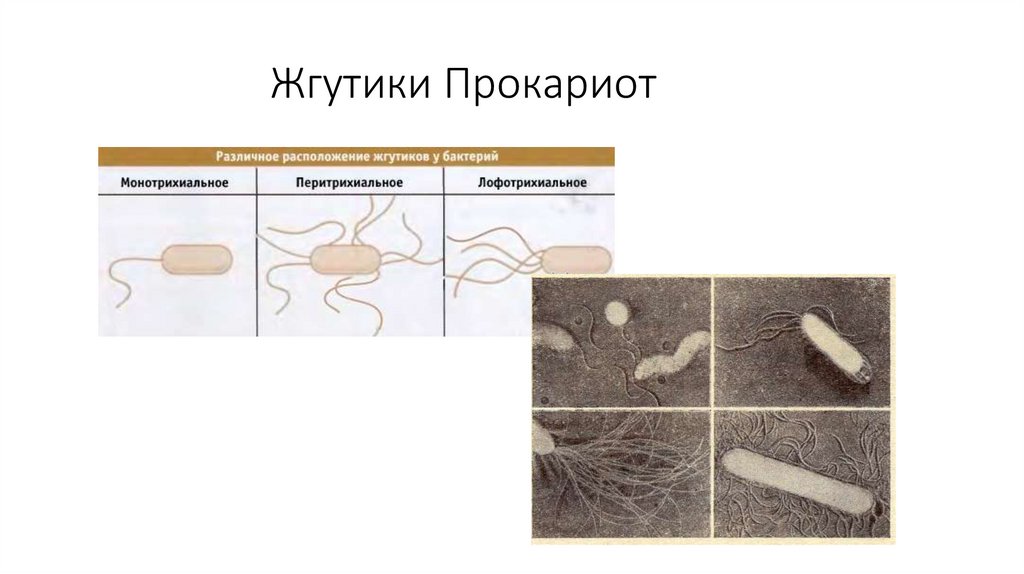
специфических функций
мембран.
• Белки составляют около
половины массы мембраны.
• Мембранные белки
различаются по структуре и
положению в липидном
бислое, что отражает их
разнообразные функции.
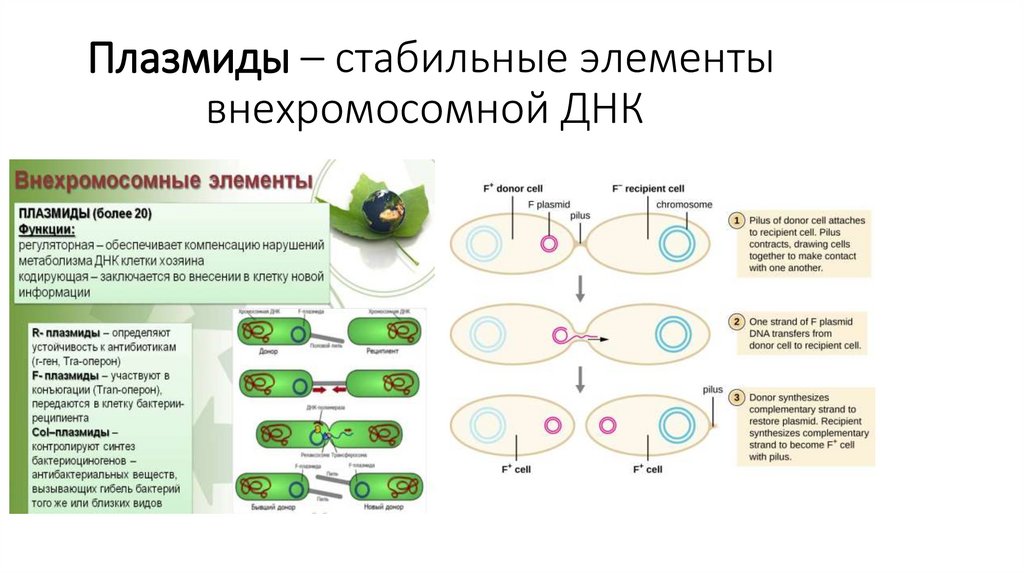
24.
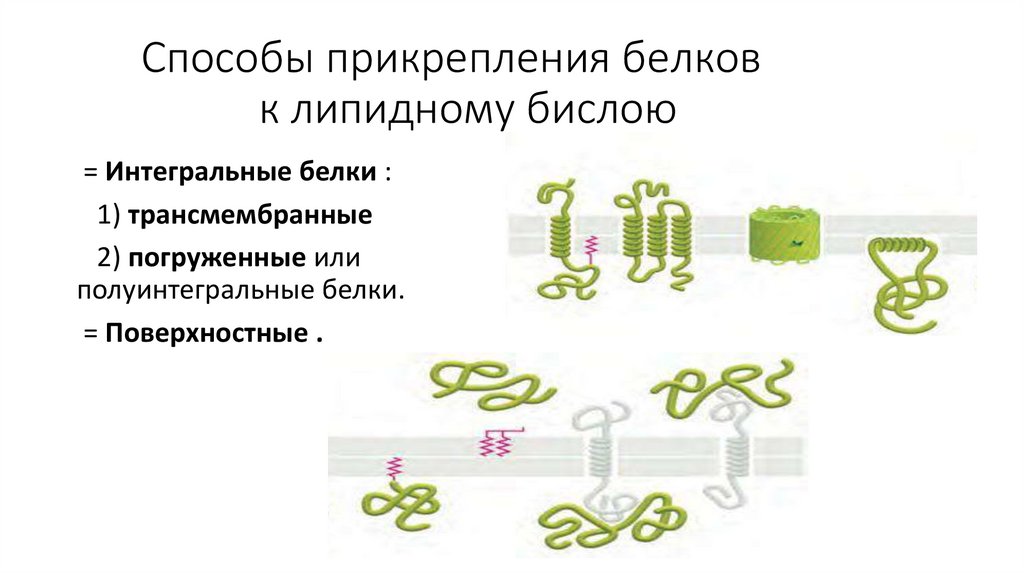
Способы прикрепления белковк липидному бислою
= Интегральные белки :
1) трансмембранные
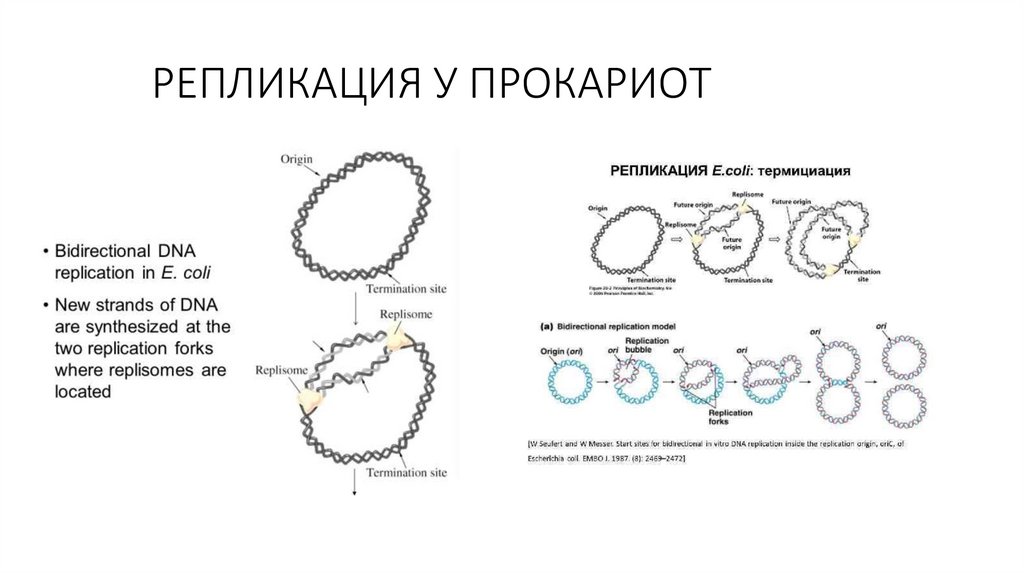
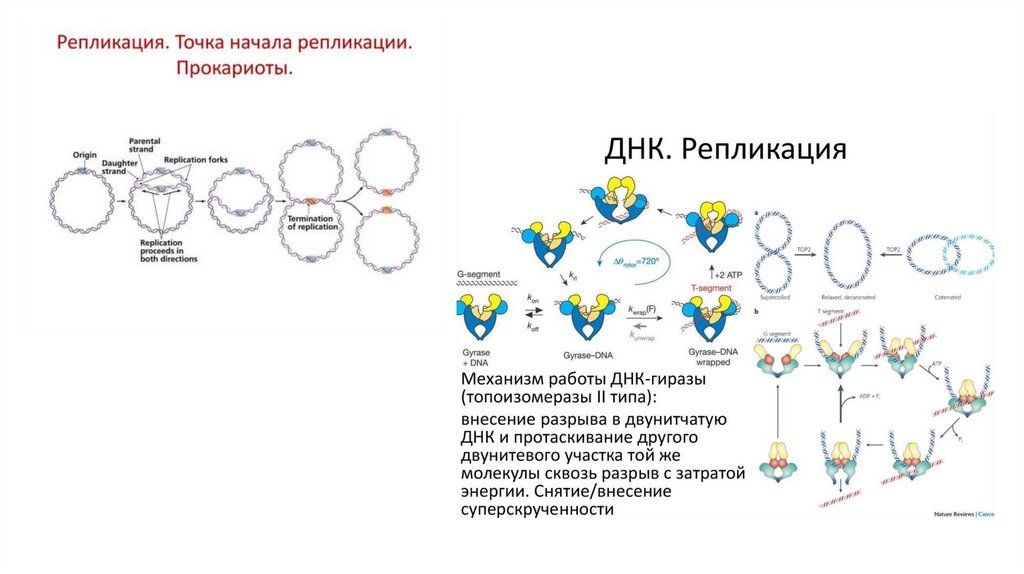
2) погруженные или
полуинтегральные белки.
= Поверхностные .
25.
• Трансмембранные белкиамфифильны ( гидрофильные и
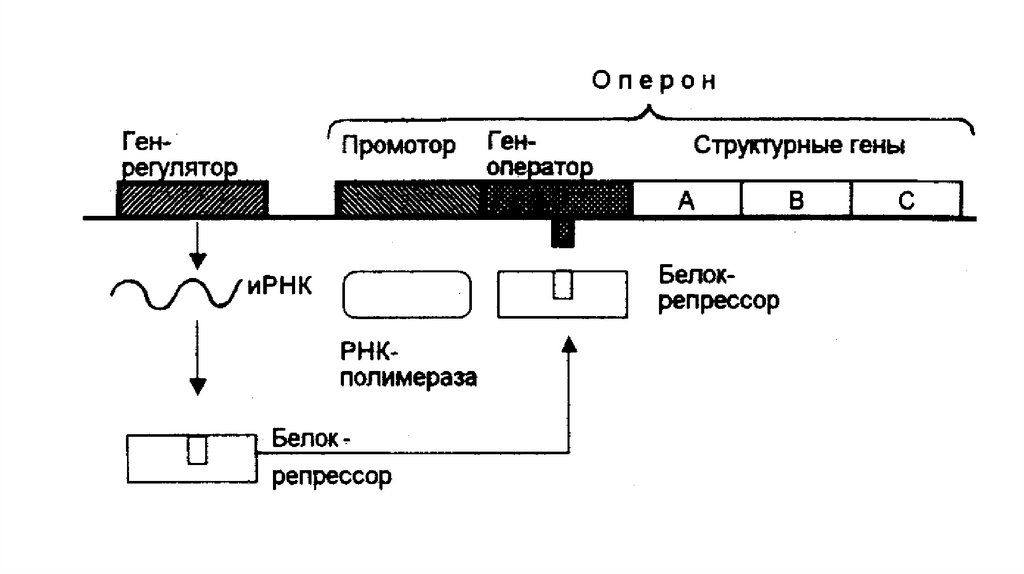
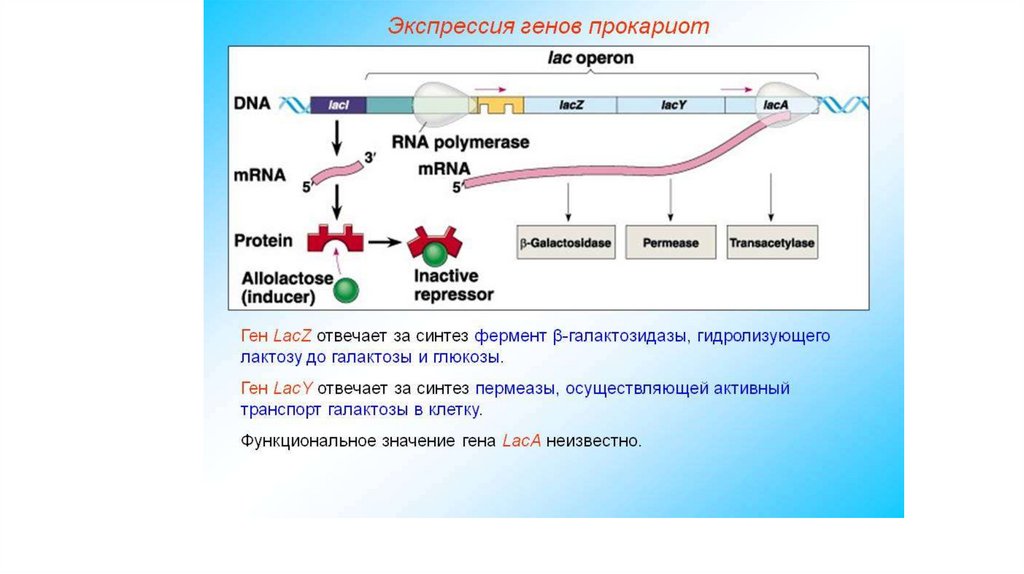
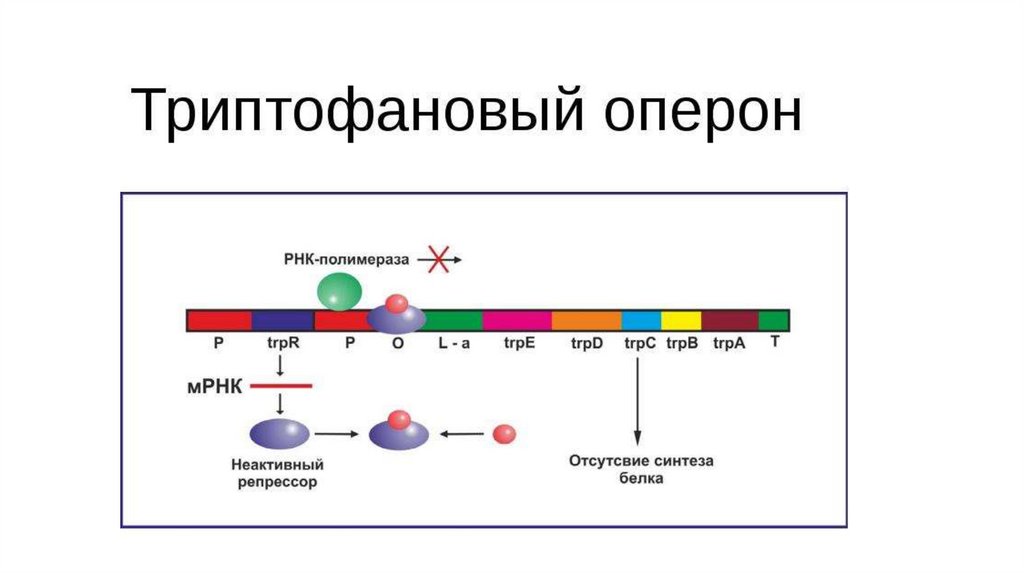
гидрофобные участки).
• Гидрофобные области находятся
в мембране и взаимодействуют с
гидрофобными «хвостами»
липидных молекул внутри
бислоя.
• Их гидрофильные участки
взаимодействуют с водой по
обеим сторонам мембраны.
26.
• Белки, как и липиды, обладают латеральной подвижностью• По расположению белков мембраны асимметричны. Особенно
это характерно для интегральных белков: их N-концы смотрят во
внешнюю для клетки среду или в вакуоли.
27.
Функции мембранных белков• По биологической роли мембранные белки делятся на 3
основных группы:
- ферменты (н-р, K-Na-зависимая АТФ-аза плазматической
мембраны, переносчики электронов и АТФ-синтетаза в
митохондриях )
- рецепторные белки (рецепторы для гормонов, вирусов,
токсинов, фоторецепторные белки и др.)
- структурные белки:
соединяют цитоскелет клетки через липидный бислой либо с
внеклеточным матриксом, либо с соседней клеткой
28.
Плазматическая мембрана и ее функции1) Барьерно-транспортная роль а) окружает клетку, определяет ее границы. Дополнительная
механическая прочность усиливается гликокаликсом и
кортикальным слоем цитоплазмы, содержащим актиновые
филаменты.
б) ограничивает свободную диффузию веществ: проницаема для
воды, газов, малых молекул жирорастворимых молекул.
Непроницаема для ионов и крупных незаряженных молекул
(сахаров). Тем самым поддерживает различия между цитозолем и
внеклеточной средой. Обладает свойством полупроницаемости.
29.
• В связи с этой функцией необходимо отметить, что прежде всегомембраны разделяют внутриклеточное пространство на
компартменты. В каждом компартменте (люмене) происходят
специфические процессы.
30.
• 2) Трансмембранный перенос ионов и низкомолекулярныхсоединений
• 3) Везикулярный транспорт – экзоцитоз и эндоцитоз .
31.
• 4) Рецепторно-сигнальная функция. В качестве рецепторов могутвыступать белки или элементы гликокаликса (гликопротеиды).
Рецепторы не только связывают специфические вещества или
реагируют на физические факторы, но и передают межклеточные
сигналы внутрь клетки.
Н-р,- гормоны связываются с рецепторами мембраны,
- происходит активация фермента аденилатциклазы, расположенной на
внутренней поверхности мембраны.
- Аденилатциклаза обеспечивает образование цАМФ из АТФ.
- цАМФ активирует другие ферменты – киназы, вызывающие
модификацию других ферментов.
32.
• 5) Межклеточное узнавание ( кадгерины, селектины, интегрины,иммуноглобулины).
На поверхности всех клеток позвоночных располагаются белки
главного комплекса гистосовместимости (major histocompatibility
complex – MHC). Это интегральные белки гликопротеины. Каждый
индивидум имеет свой набор таких белков. Т-лимфоциты узнают
МНС своего организма. При изменении структуры МНС ( в
результате мутации или связи с вирусом) Т-лимфоциты уничтожают
такие клетки.
33.
• 6) Плазматическая мембрана обеспечивает формирование всехтипов межклеточных контактов.
34.
Транспортные процессы через мембрану• Транспорт м.б.
- активным
- пассивным
• Транспорт веществ внутрь клетки называется эндоцитоз, из
клетки - экзоцитоз.
35.
Мембранные белки, которые обеспечиваюттранспорт молекул через липидный бислой:
- транспортеры, обладающие подвижными частями для
транспорта специфических молекул
- каналы, которые формируют узкую гидрофильную пору,
способствующую пассивному трансмембранному движению
молекул
36.
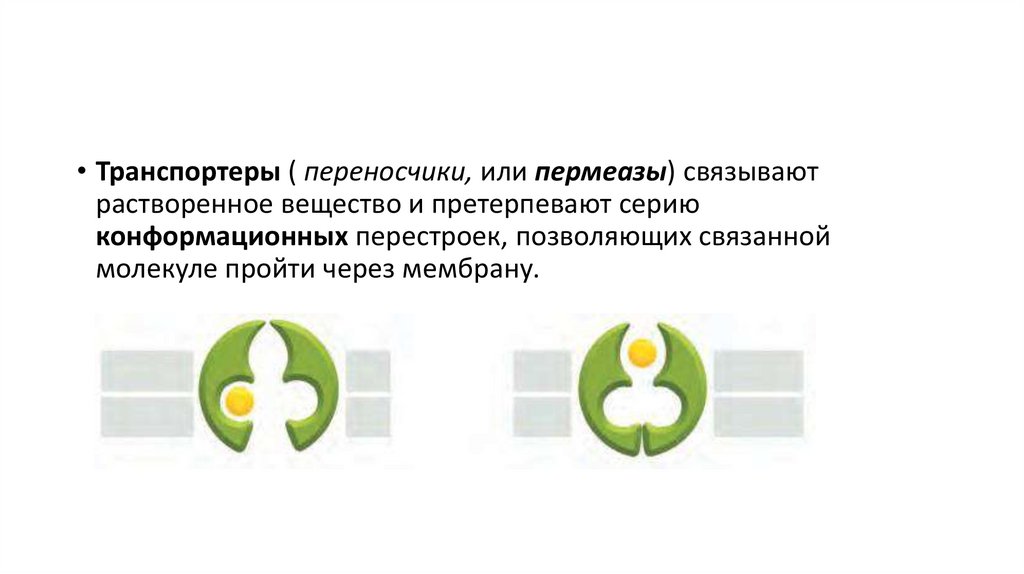
• Транспортеры ( переносчики, или пермеазы) связываютрастворенное вещество и претерпевают серию
конформационных перестроек, позволяющих связанной
молекуле пройти через мембрану.
37.
• Каналы слабее взаимодействуют с растворенным веществом.Они формируют в липидном бислое заполненные водой поры.
Когда они открыты, растворенные вещества проходят через них и
пересекают мембрану. Транспорт через каналы происходит
значительно быстрее, чем транспорт, опосредованный
транспортерами (облегченная диффузия).
38.
• Вода способна диффундировать через искусственные липидныебислои. Однако, клетки содержат специфические канальные
белки (так называемые водные каналы, или аквапорины),
которые значительно увеличивают проницаемость мембран для
воды.
39.
• Все каналы и многие транспортеры позволяют молекулампересекать мембрану пассивно без затрат энергии («по
градиенту концентрации»). Этот процесс называется пассивным
транспортом, или облегченной диффузией.
40.
• Почти все плазматические мембраны обладают разностьюэлектрических потенциалов (градиентом потенциала);
внутренняя сторона мембраны обычно заряжена отрицательно
по отношению к внешней.
• Эта разность потенциалов способствует входу положительно
заряженных ионов в клетку, но мешает входу отрицательно
заряженных ионов.
41.
• В мембранах существуют транспортные белки, которые могутактивно перекачивать определенные растворенные вещества
через мембрану против их электрохимических градиентов.
• Такой активный транспорт, опосредуется транспортерами,
которые также называют насосами.
• При активном транспорте активность переносчика сопряжена с
гидролизом ATP или ионным градиентом.
42.
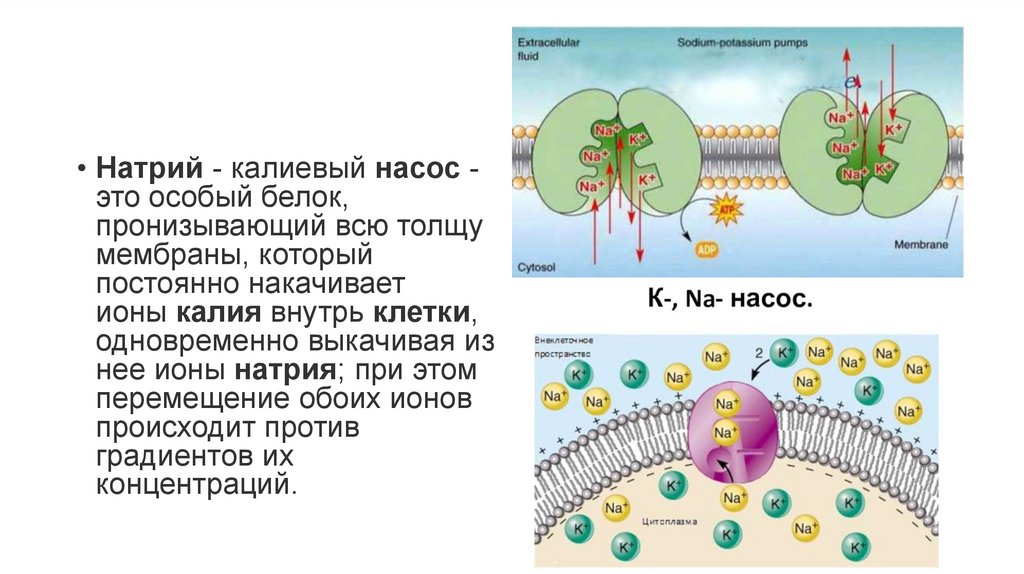
• Натрий - калиевый насос это особый белок,пронизывающий всю толщу
мембраны, который
постоянно накачивает
ионы калия внутрь клетки,
одновременно выкачивая из
нее ионы натрия; при этом
перемещение обоих ионов
происходит против
градиентов их
концентраций.
43.
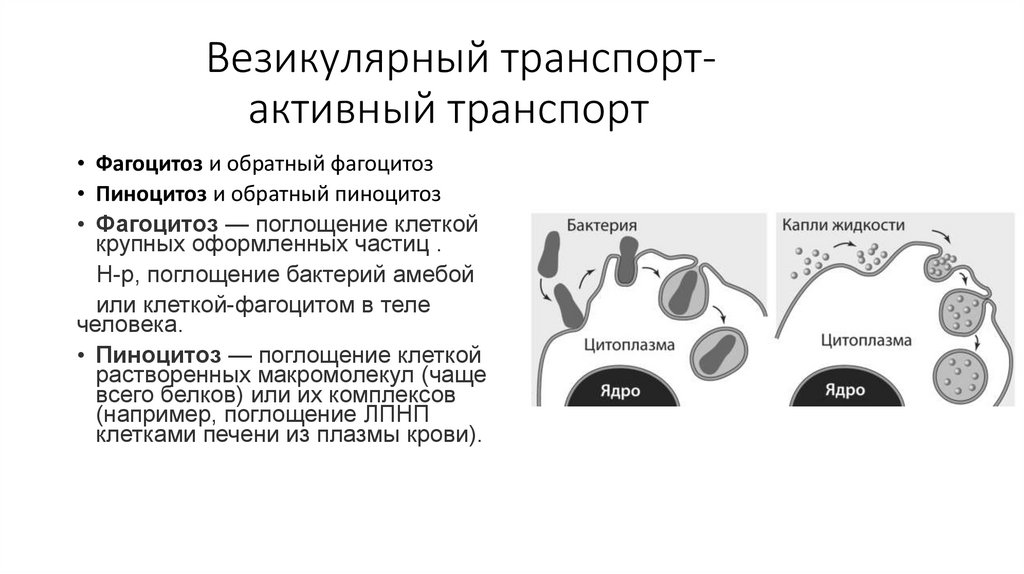
Везикулярный транспортактивный транспорт• Фагоцитоз и обратный фагоцитоз
• Пиноцитоз и обратный пиноцитоз
• Фагоцитоз — поглощение клеткой
крупных оформленных частиц .
Н-р, поглощение бактерий амебой
или клеткой-фагоцитом в теле
человека.
• Пиноцитоз — поглощение клеткой
растворенных макромолекул (чаще
всего белков) или их комплексов
(например, поглощение ЛПНП
клетками печени из плазмы крови).
44.
45.
ГиалоплазмаОсновная плазма(греч. hyalos стекло ), матрикс цитоплазмы,
сложная бесцветная коллоидная система в клетке, способная
к обратимым переходам из золя в гель.
В состав гиалоплазмы входят растворимые белки,
растворимые РНК, полисахариды, липиды.
Гиалоплазма эукариот находится в движении – циклоз.
46.
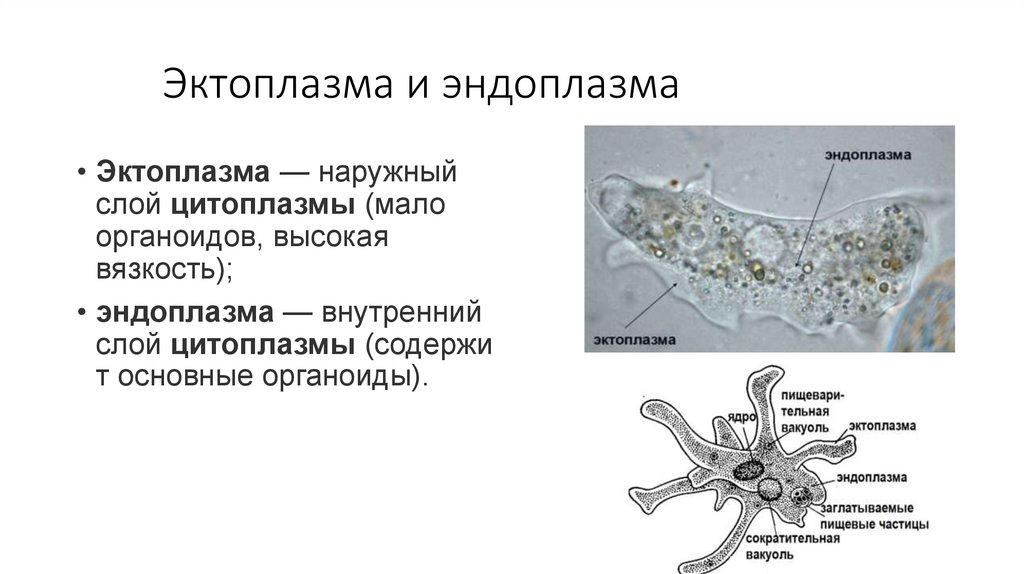
Эктоплазма и эндоплазма• Эктоплазма — наружный
слой цитоплазмы (мало
органоидов, высокая
вязкость);
• эндоплазма — внутренний
слой цитоплазмы (содержи
т основные органоиды).
47.
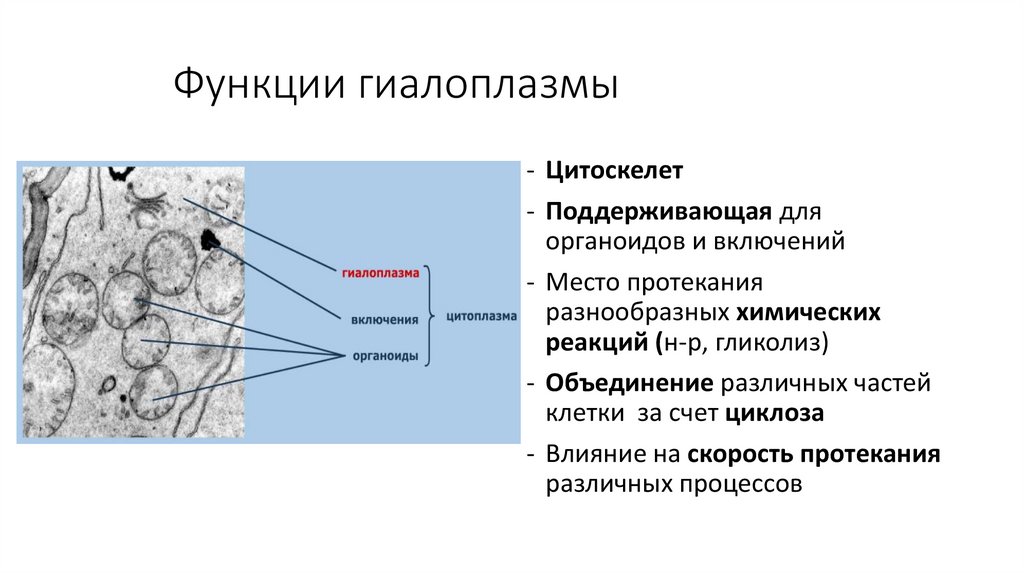
Функции гиалоплазмы- Цитоскелет
- Поддерживающая для
органоидов и включений
- Место протекания
разнообразных химических
реакций (н-р, гликолиз)
- Объединение различных частей
клетки за счет циклоза
- Влияние на скорость протекания
различных процессов
48.
Включения• Включения цитоплазмы —
это необязательные
компоненты клетки,
появляющиеся и
исчезающие в зависимости
от интенсивности и
характера обмена
веществ в клетке и от
условий существования
организма.
49.
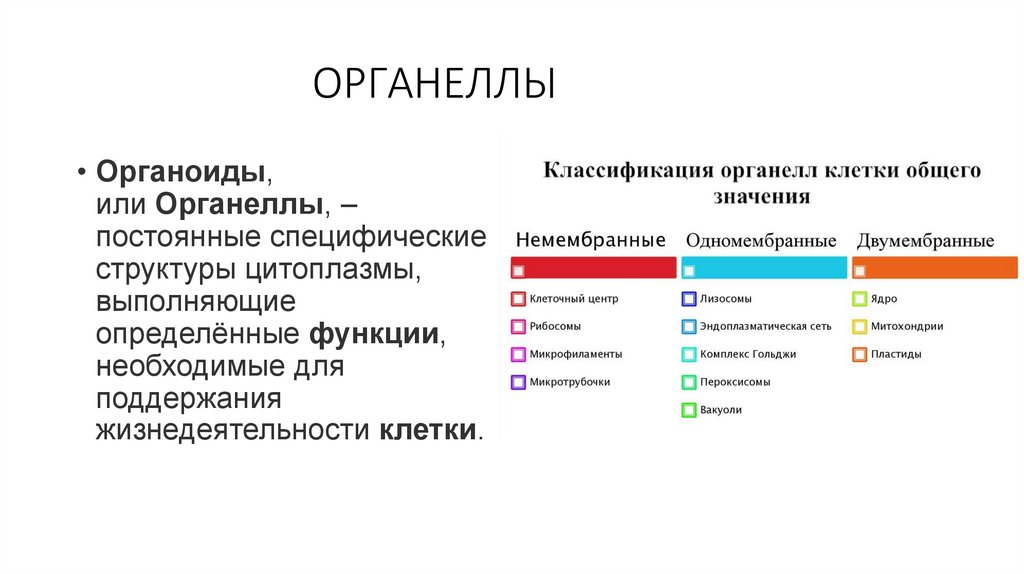
ОРГАНЕЛЛЫ• Органоиды,
или Органеллы, –
постоянные специфические
структуры цитоплазмы,
выполняющие
определённые функции,
необходимые для
поддержания
жизнедеятельности клетки.
50.
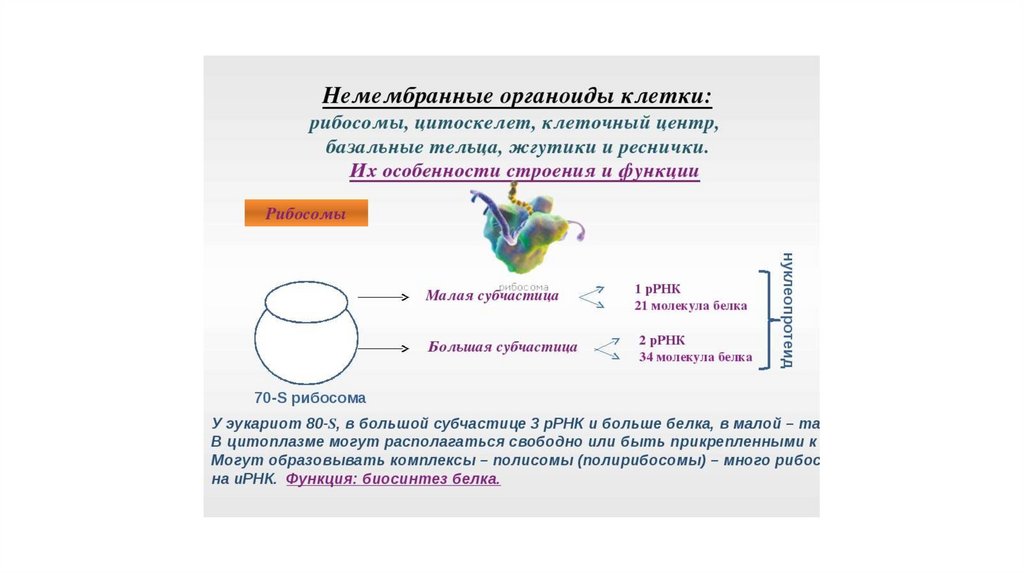
Немембранные органеллы• Рибосомы
• Цитоскелет клетки
• Клеточный центр
• Органоиды движения
51.
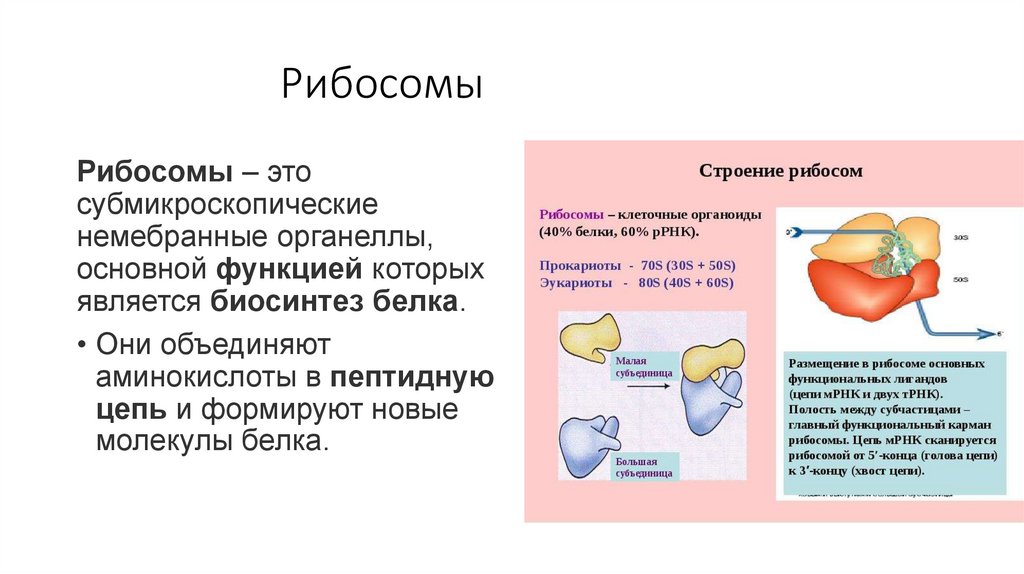
РибосомыРибосомы – это
субмикроскопические
немебранные органеллы,
основной функцией которых
является биосинтез белка.
• Они объединяют
аминокислоты в пептидную
цепь и формируют новые
молекулы белка.
52.
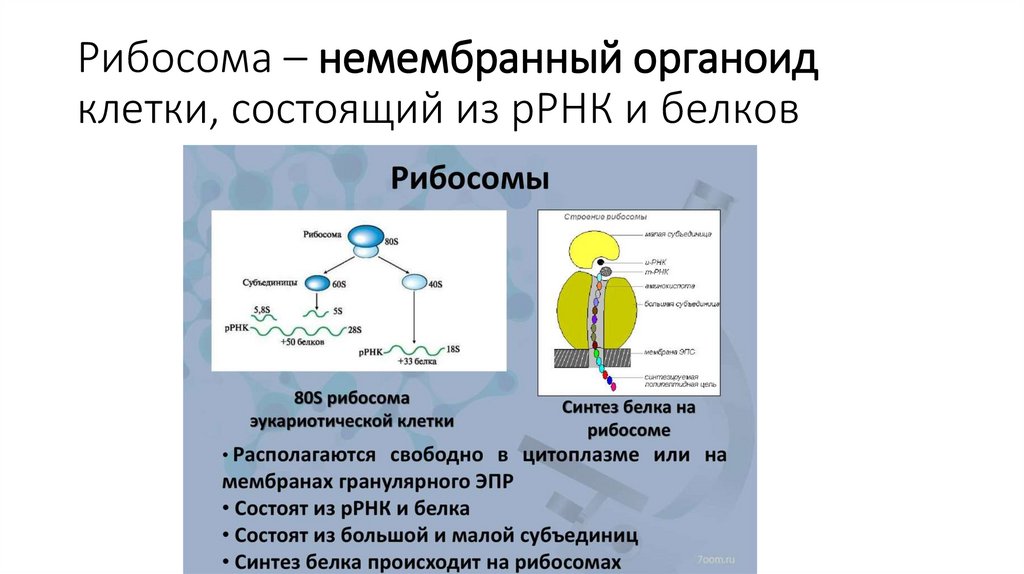
Рибосома – немембранный органоидклетки, состоящий из рРНК и белков
53.
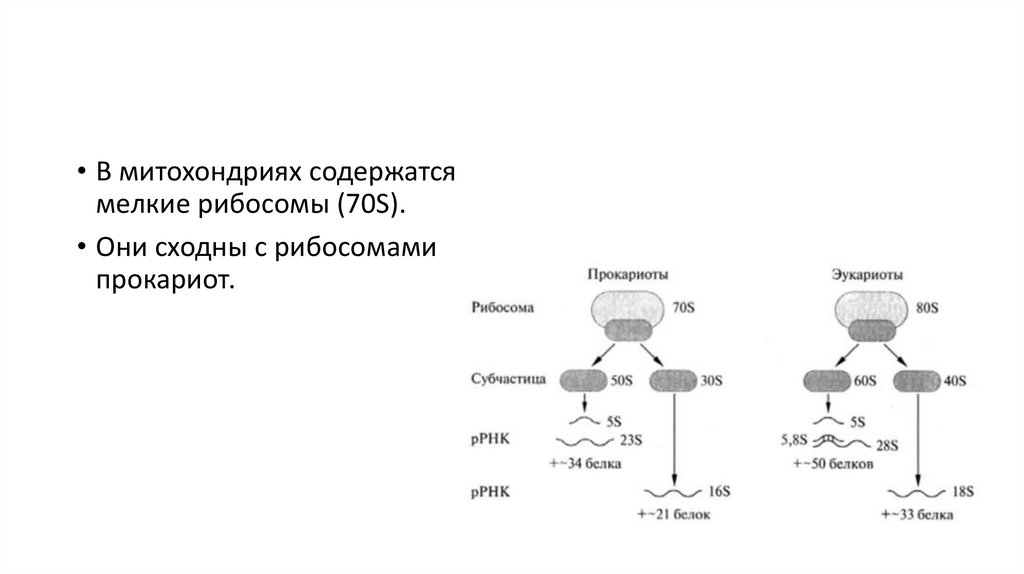
• В митохондриях содержатсямелкие рибосомы (70S).
• Они сходны с рибосомами
прокариот.
54.
Образование рибосомпроисходит в ЯДРЫШКЕ
• Ядрышко находится внутри ядра и не имеет собственной мембранной
оболочки
• Ядрышко формируется в связи с ядрышковыми организаторами,
содержащими гены, определяющие структуру рРНК. Эти гены
образуют многочисленные копии
(амплифицированы).
• В геноме человека гены,
определяющие структуру рРНК,
находятся в 13, 14, 15, 21, 22
хромосомах
(область вторичной перетяжки)
55.
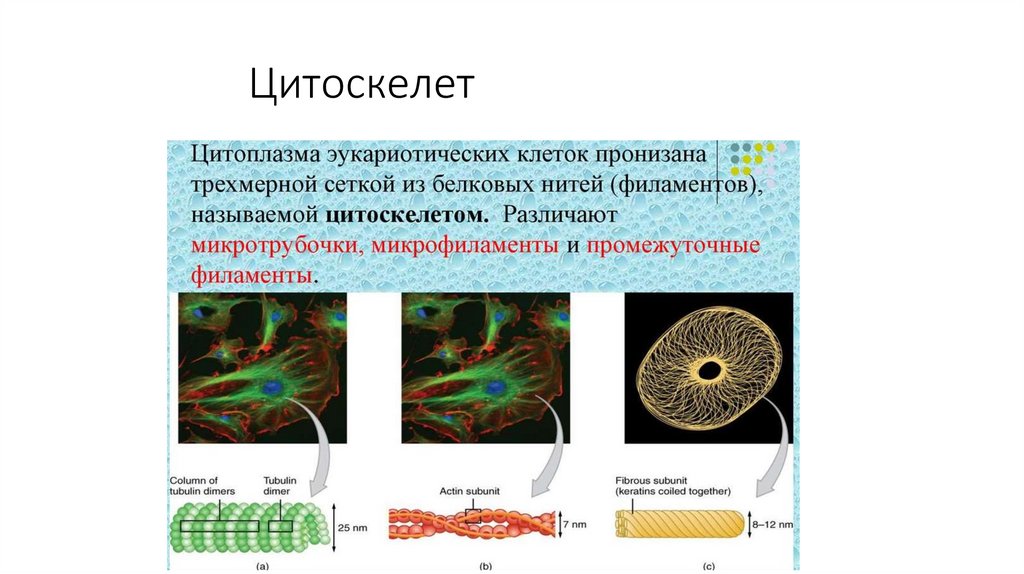
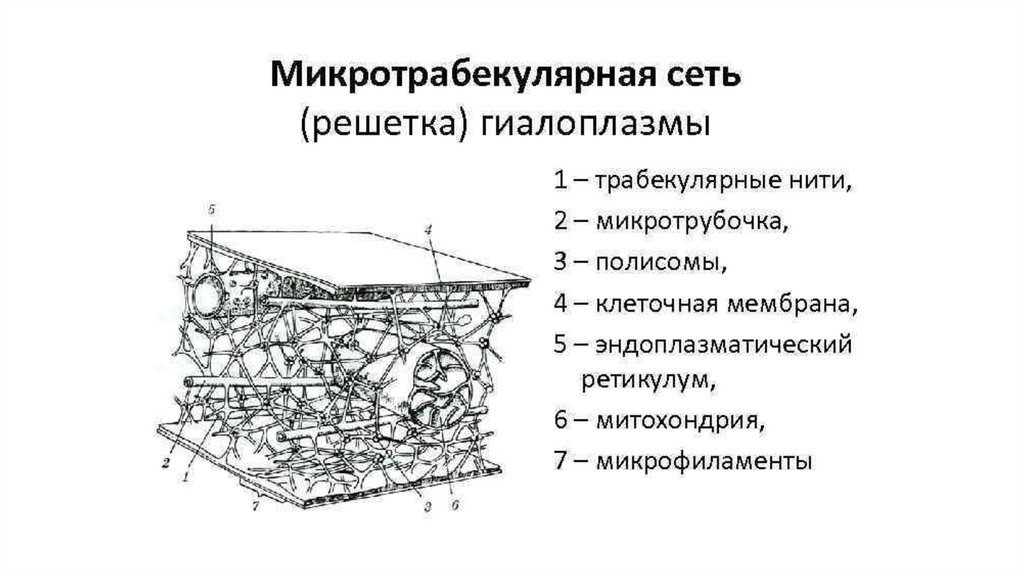
Цитоскелет56.
57.
Функции цитоскелета• Поддержание формы клетки
• Обеспечение внутриклеточного транспорта
• Создает механическое сочленение
• Поддержание мембраны клетки
• Растаскивает хромосомы при делении клетки
• Обеспечивает движение клеток (н-р, сперматозоидов,
фибробластов, лейкоцитов)
• Мышечное сокращение
• Обеспечивает удлинение аксонов и дендритов и др.
58.
Одномембранные органеллы• Эндоплазматическая сеть (ЭПС)
• Комплекс или аппарат Гольджи
• Лизосомы
• Пероксисомы
• Вакуоли
59.
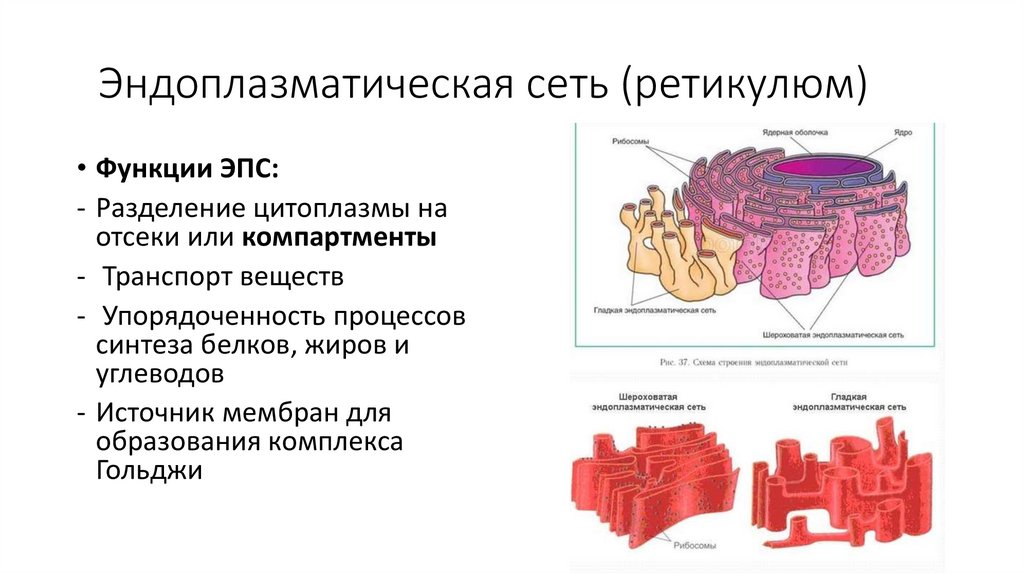
Эндоплазматическая сеть (ретикулюм)• Функции ЭПС:
- Разделение цитоплазмы на
отсеки или компартменты
- Транспорт веществ
- Упорядоченность процессов
синтеза белков, жиров и
углеводов
- Источник мембран для
образования комплекса
Гольджи
60.
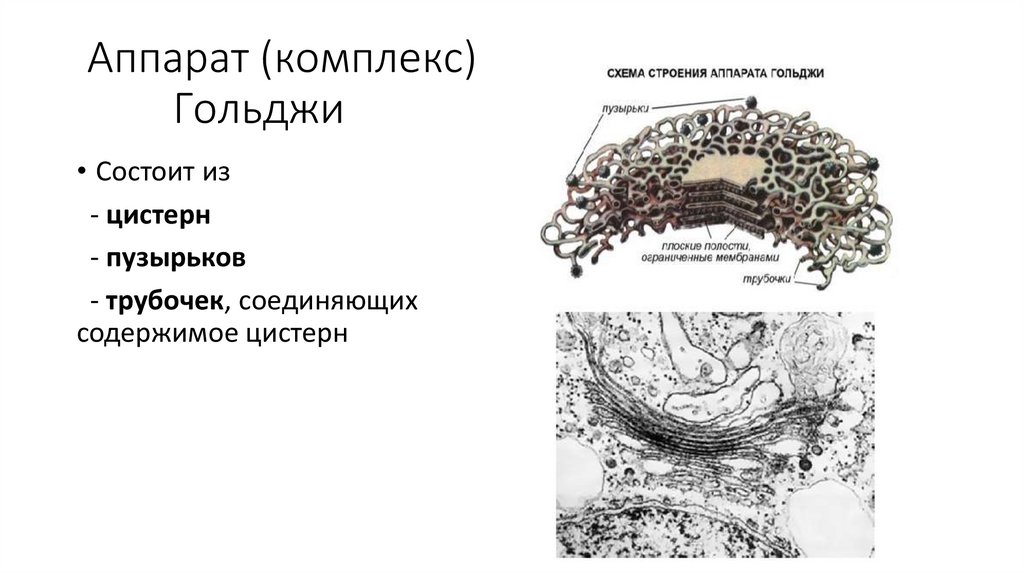
Аппарат (комплекс)Гольджи
• Состоит из
- цистерн
- пузырьков
- трубочек, соединяющих
содержимое цистерн
61.
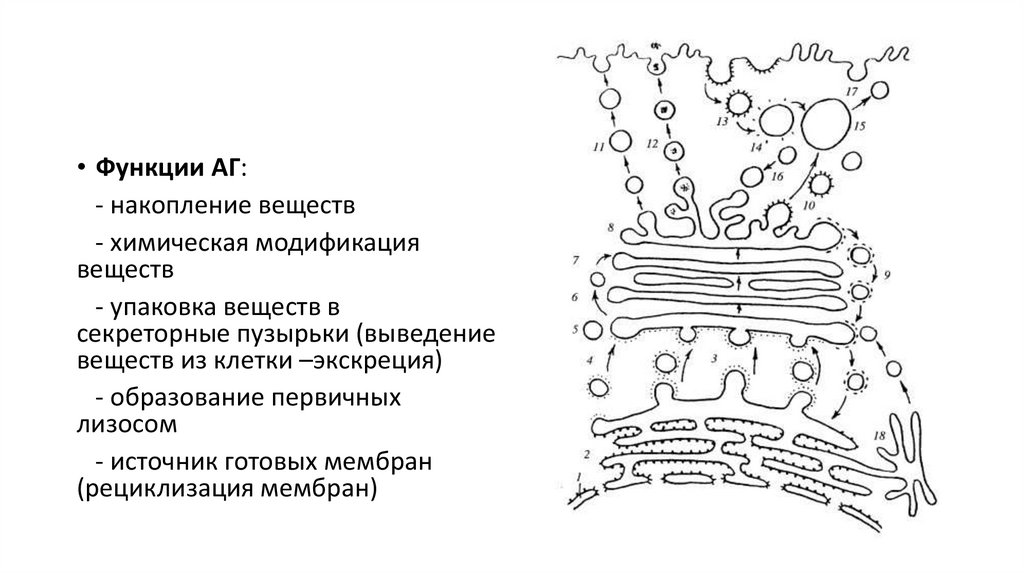
• Функции АГ:- накопление веществ
- химическая модификация
веществ
- упаковка веществ в
секреторные пузырьки (выведение
веществ из клетки –экскреция)
- образование первичных
лизосом
- источник готовых мембран
(рециклизация мембран)
62.
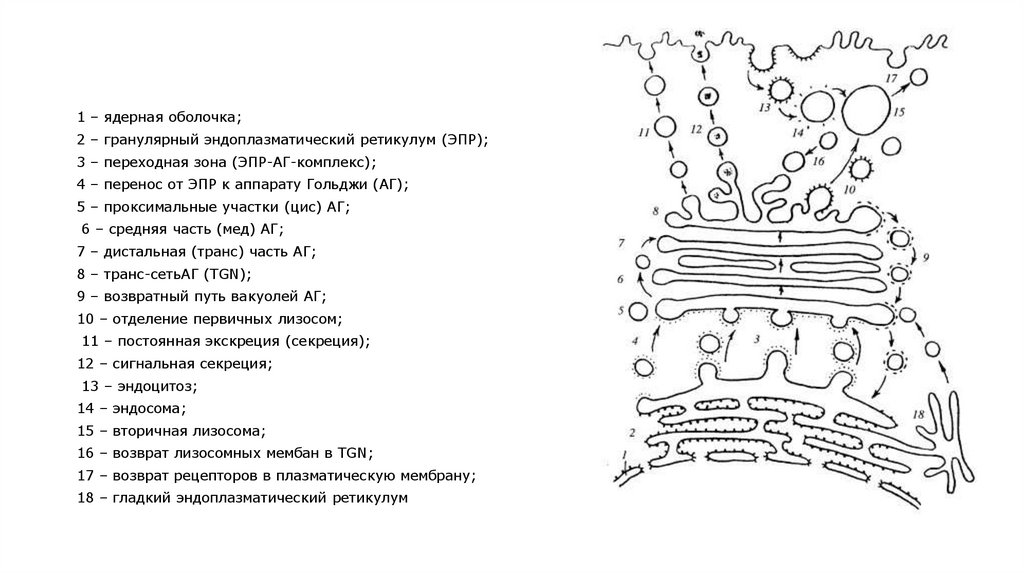
1 – ядерная оболочка;2 – гранулярный эндоплазматический ретикулум (ЭПР);
3 – переходная зона (ЭПР-АГ-комплекс);
4 – перенос от ЭПР к аппарату Гольджи (АГ);
5 – проксимальные участки (цис) АГ;
6 – средняя часть (мед) АГ;
7 – дистальная (транс) часть АГ;
8 – транс-сетьАГ (TGN);
9 – возвратный путь вакуолей АГ;
10 – отделение первичных лизосом;
11 – постоянная экскреция (секреция);
12 – сигнальная секреция;
13 – эндоцитоз;
14 – эндосома;
15 – вторичная лизосома;
16 – возврат лизосомных мембан в TGN;
17 – возврат рецепторов в плазматическую мембрану;
18 – гладкий эндоплазматический ретикулум
63.
• Лизосомы – одномембранные органоиды• Первичные лизосомы образуются из аппарата
Гольджи;
*Сходны с секреторными вакуолями: в некоторых
случаях могут подходить к ЦПМ и выбрасывать
свое содержимое путем экзоцитоза
*Содержат около 40 гидролитических ферментов
(кислая фосфатаза – маркерный фермент)
64.
• В мембране лизосом- АТФ-зависимые протонные насосы.Поэтому внутри – сильно кислая среда (рН 4,5-5.0 по сравнению с
рН 7,0-7,3 в цитоплазме.
• В мембрану встроены также белки-переносчики для транспорта
продуктов расщепления.
• Содержат около 40 гидролаз, расщепляющих все
макромолекулы.
65.
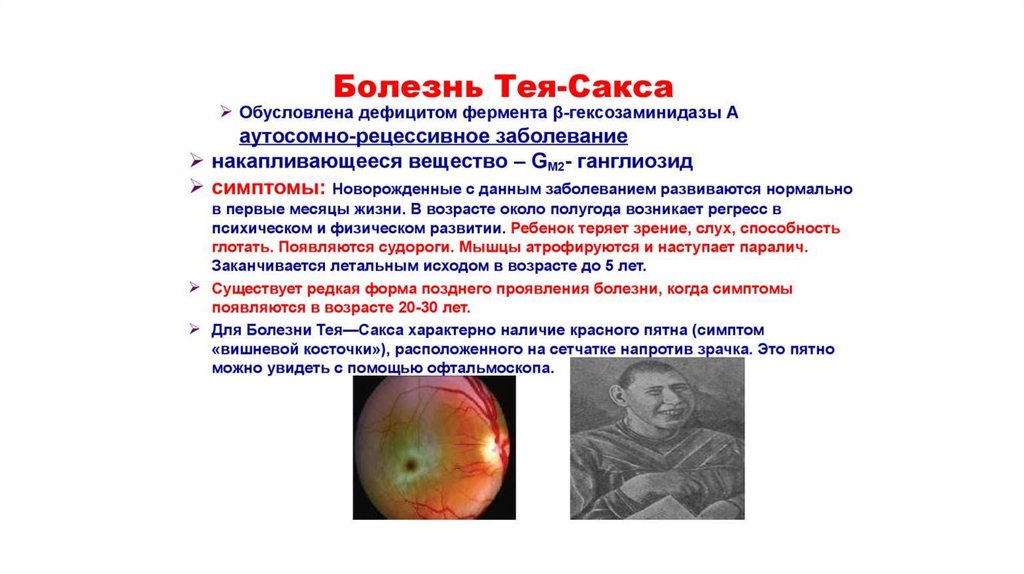
• Лизосомальные болезни накопления ( известно более 25 такихзаболеваний).
• Н-р, синдром Тея-Сакса – нарушение расщепления ганглиозидов;
в результате – они откладываются в ЦНС и вызывают
дегенерацию нервной ткани.
66.
67.
Типы лизосом• Первичные лизосомы – отделяются от АГ и содержат гидролазы
• Вторичные лизосомы (пищеварительная вакуоль)– результат
слияния первичных лизосом с фагоцитарными и пиноцитозными
вакуолями (эндосомами)
• Телолизосомы или остаточные тельца – содержат
непереваренные продукты. У человека при старении эти тельца
содержат пигмент старения – липофусцин ( в клетках печени,
мозга, мышц)
• Аутолизосомы - уничтожают собственные «сломанные»
органоиды (н-р, в клетках печени уничтожают митохондрии,
которые живут 10 дней)
68.
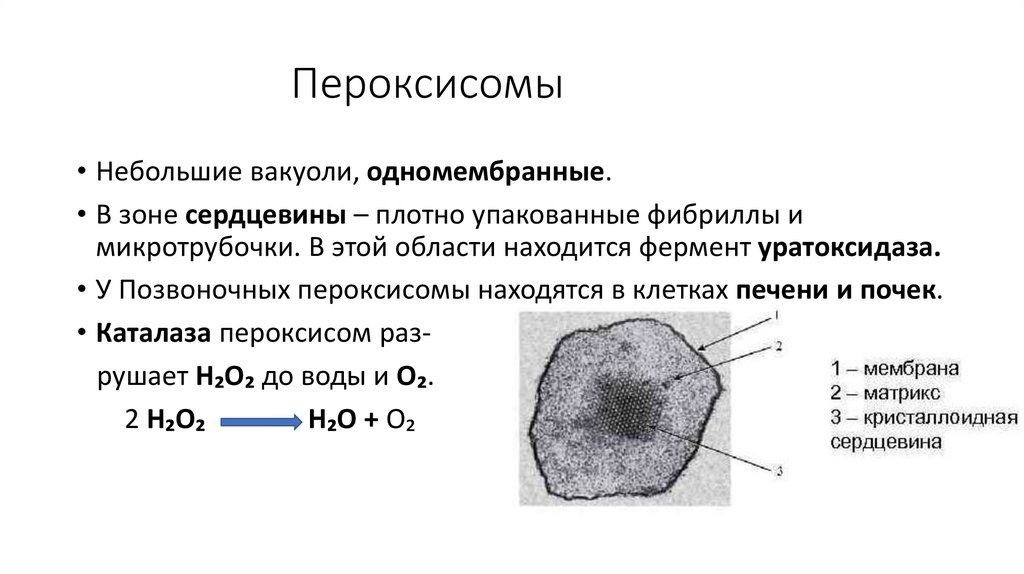
Пероксисомы• Небольшие вакуоли, одномембранные.
• В зоне сердцевины – плотно упакованные фибриллы и
микротрубочки. В этой области находится фермент уратоксидаза.
• У Позвоночных пероксисомы находятся в клетках печени и почек.
• Каталаза пероксисом разрушает Н₂О₂ до воды и О₂.
2 Н₂О₂
Н₂О + О₂
69.
• Ферменты пероксисомы могут использовать пероксид водородадля окисления субстратов – метанола, формальдегида,
муравьиной кислоты, нитритов, обезвреживая их
Н₂О₂ + АН₂
Н₂О + А
• Пероксисомные нарушения приводят к поражению печени,
головного мозга, атрофии мозгового вещества надпочечников,
нарушению зрения и слуха, ухудшению работы сердца.
70.
Образование пероксисом71.
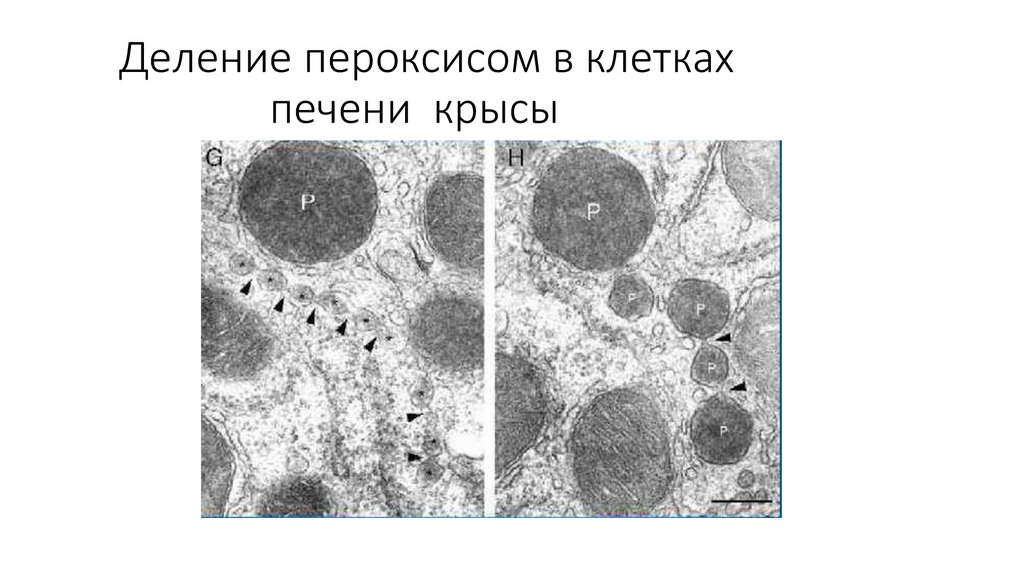
Деление пероксисом в клеткахпечени крысы
72.
Вакуолярная система внутриклеточноготранспорта
• Вакуолярная система – система одномембранных органелл:
- ЭПР
- аппарат Гольджи
- лизосомы
- эндосомы
- секреторные вакуоли
Функция : синтез
перестройка(модификация)
сортировка
выведение различных биополимеров
73.
74.
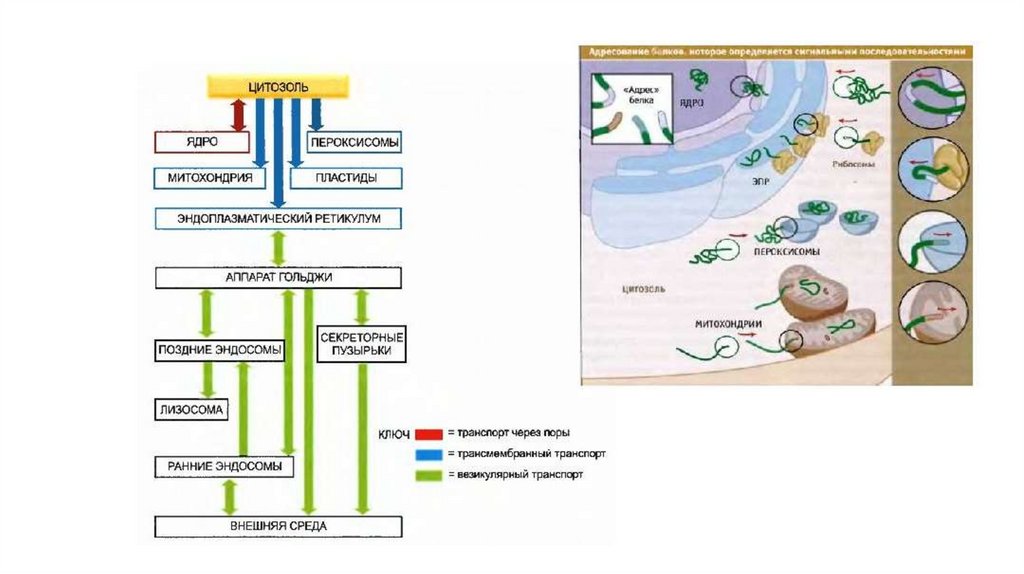
• Белки имеют сигнальные последовательности –последовательности аминокислот в первичной структуре,
которые распознаются рецепторами органеллы-мишени
(мембранное адресование белков). Если белок не имеет
адресного сигнала, он остается в цитозоле.
• Для большинства секреторных и трансмембранных белков
адресование и транслокация происходят во время синтеза белка.
75.
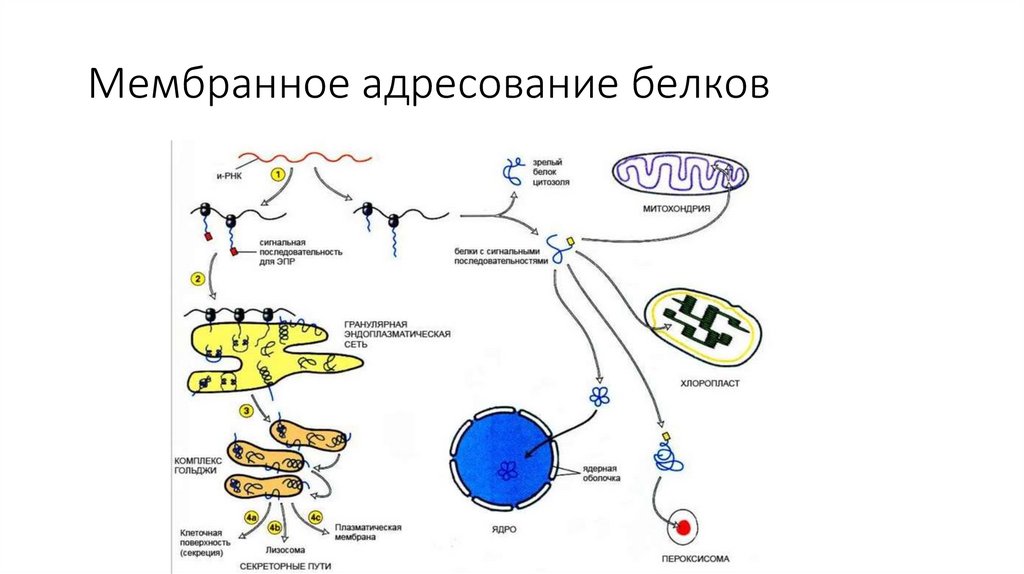
Мембранное адресование белков76.
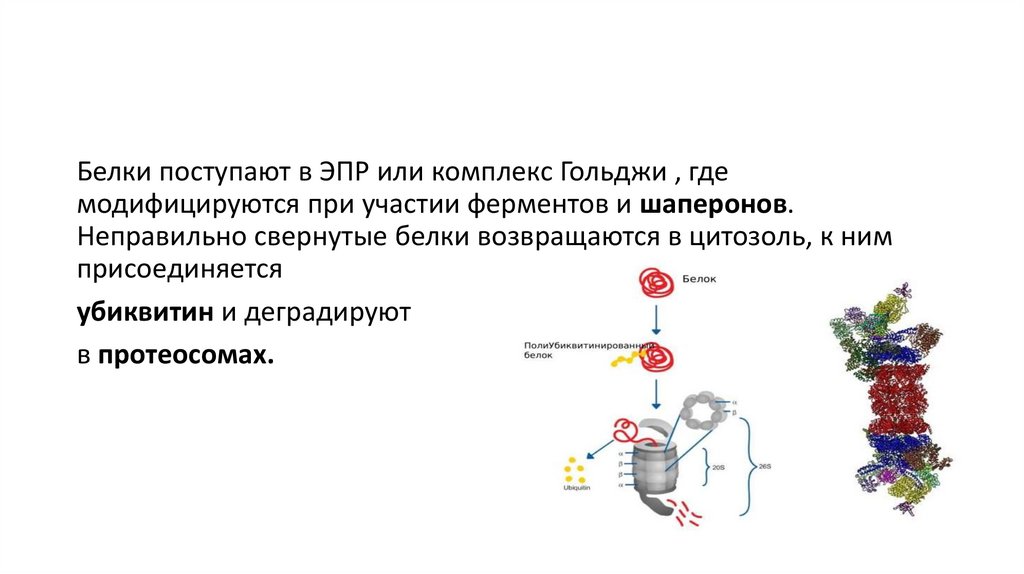
Белки поступают в ЭПР или комплекс Гольджи , гдемодифицируются при участии ферментов и шаперонов.
Неправильно свернутые белки возвращаются в цитозоль, к ним
присоединяется
убиквитин и деградируют
в протеосомах.
77.
78.
79.
• Важнейшей функцией вакуолярной системы является такжефункция рециклизации мембран,
в том числе – плазматической
мембраны.
80.
Двумембранные органоиды• Митохондрии
• Пластиды
• Ядро (?)
81.
МИТОХОНДРИИ82.
83.
• Гипотеза симбиотического происхождения митохондрий:древние прокариоты, имеющие разный набор ферментов,
перешли к симбиозу после того, как одна из них фагоцитировала
другую, но не переварила.
• Доказательства:
- наличие в митохондриях мелких рибосом
- наличие кольцевой молекулы ДНК
- наличие двух мембран
84.
85.
Митохондрии способны ксамовоспроизведению
86.
МИТОХОНДРИАЛЬНАЯ ДНК• Кольцевая молекула ДНК митохондрий у человека состоит из
16 569 пар нуклеотидов и содержит 37 генов, кодирующих рРНК и
тРНК и 13 генов кодируют субъединицы белков дыхательной цепи.
Митохондриальные заболевания, связанные с мутациями
мтДНК, затрагивают органы с интенсивным энергетическим
обменом ( чаще- мышечная система и нервная система).
Н-р, парез мышц глазного яблока и век, миопатия, нарушение
координации движений, полинейропатия и т.д.
87.
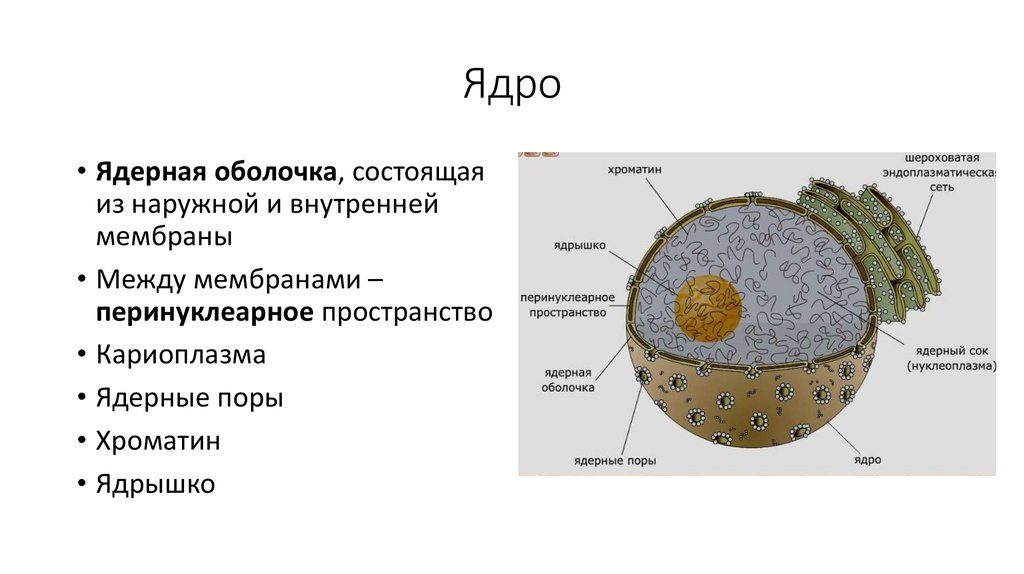
Ядро• Ядерная оболочка, состоящая
из наружной и внутренней
мембраны
• Между мембранами –
перинуклеарное пространство
• Кариоплазма
• Ядерные поры
• Хроматин
• Ядрышко
88.
Функции ядра• Хранение наследственной информации
• Передача наследственной информации из поколения в
поколение
• Реализация наследственной информации
89.
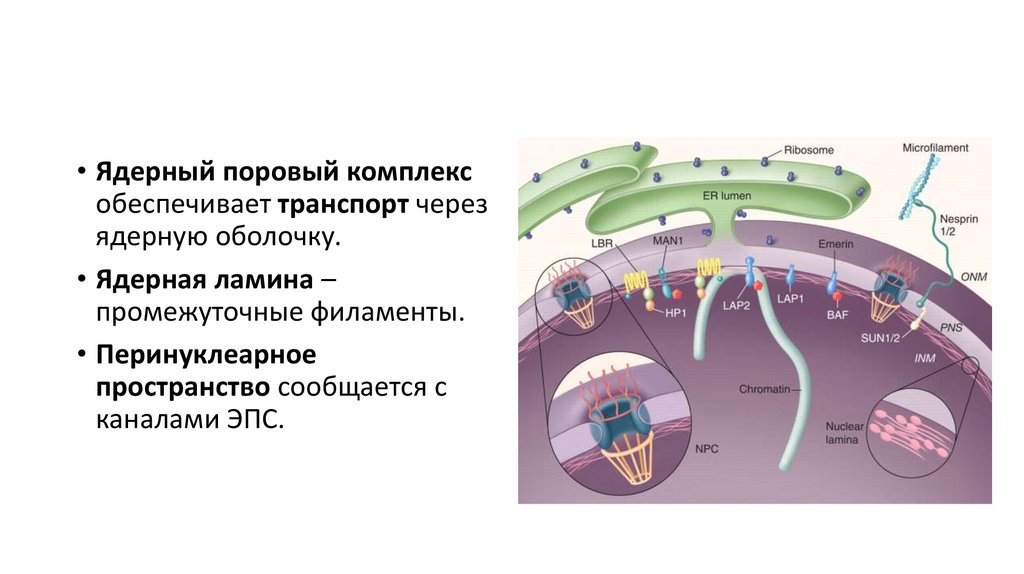
• Ядерный поровый комплексобеспечивает транспорт через
ядерную оболочку.
• Ядерная ламина –
промежуточные филаменты.
• Перинуклеарное
пространство сообщается с
каналами ЭПС.
90.
В ЯДРЫШКЕ происходит образование рибосом• Ядрышко находится внутри ядра и не имеет собственной мембранной
оболочки
• Ядрышко формируется в связи с ядрышковыми организаторами,
содержащими гены, определяющие структуру рРНК. Эти гены
образуют многочисленные копии
(амплифицированы).
• В геноме человека гены,
определяющие структуру рРНК,
находятся в 13, 14, 15, 21, 22
хромосомах
(область вторичной перетяжки)
91.
92.
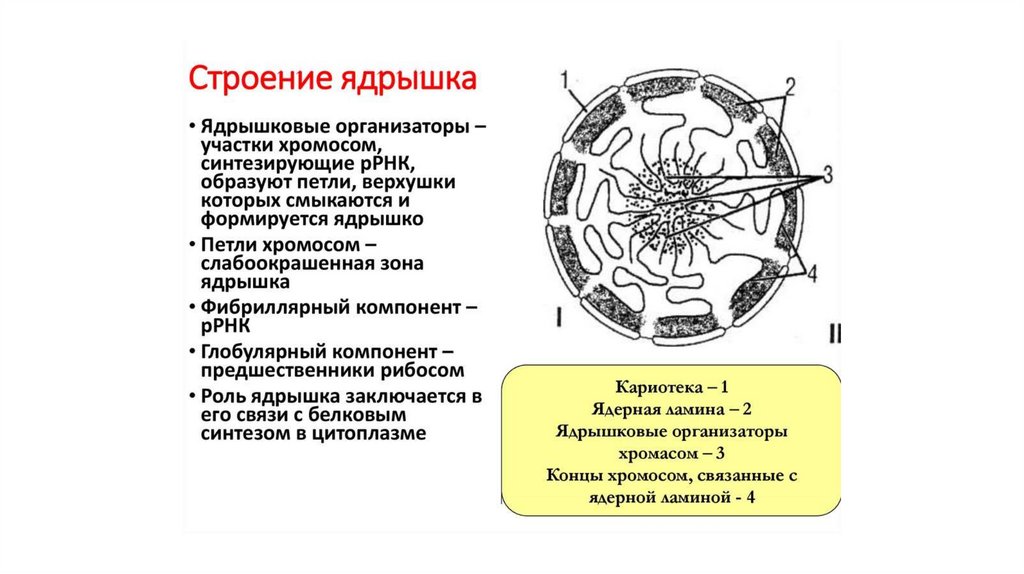
• Функция ядрышка- синтез рРНК
- сборка субъединиц рибосом
В ядрышко из цитоплазмы проникают белки, принимающие
участие в этих процессах. Большинство из этих белков имеют
специальную «метку»- сигнал ядрышковой локализации
(Nucleolus Localization Signal – NoLS).
В ядрышке наблюдается самая высокая концентрация белков в
клетке ( около 600 видов белков)
93.
94.
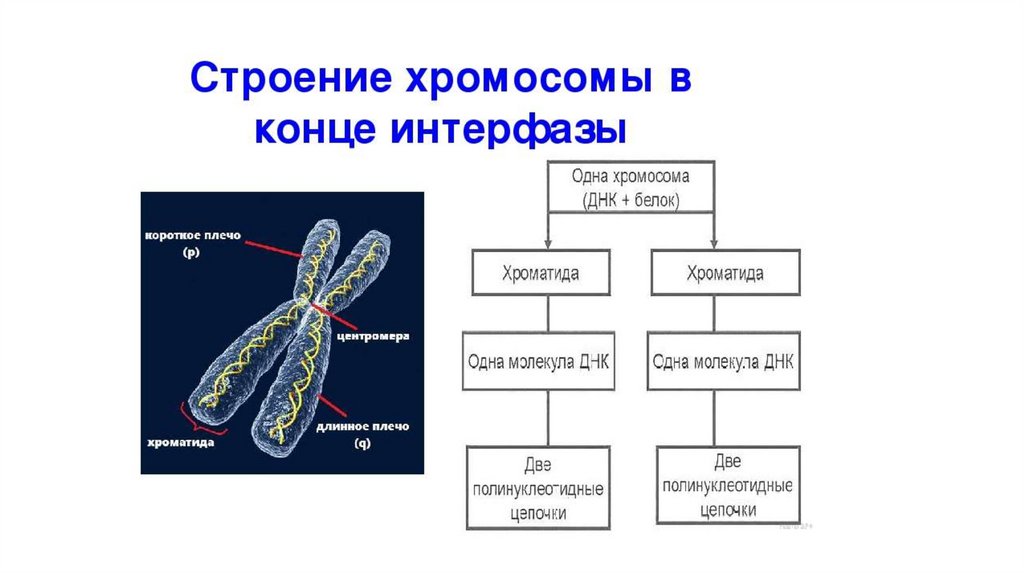
Хроматин и хромосомы• В промежутках между делениями клетки (интерфаза)
наследственный материал имеет вид
тонких нитей хроматина,
расположенных в ядре клетки.
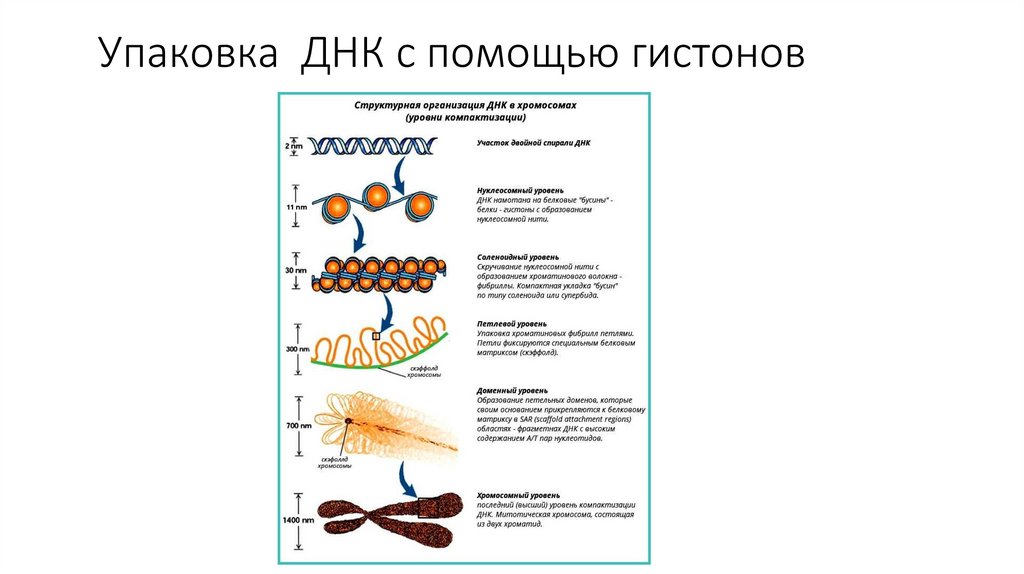
• Хроматин образуется в результате
упаковки молекул ДНК с помощью
белков – гистонов.
Количество нитей хроматина является
видоспецифическим признаком
95.
Упаковка ДНК с помощью гистонов96.
• Во время деления клетки изнитей хроматина образуются
хромосомы (путем
спирализации хроматина).
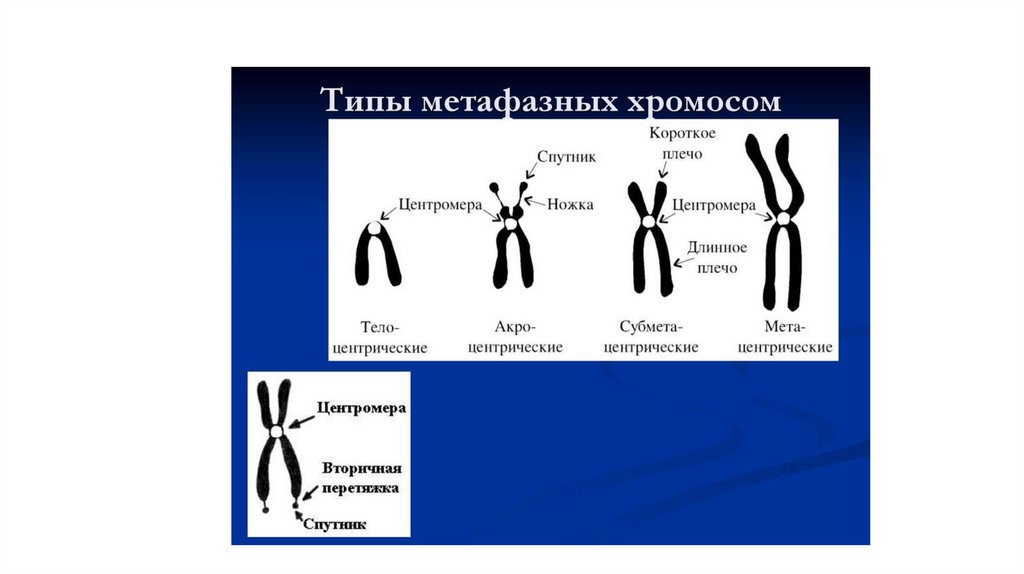
• Хромосомы отличаются по
- длине,
- положению первичной
перетяжки (центромеры)
-наличию вторичной перетяжки,
отделяющей «спутник»
97.
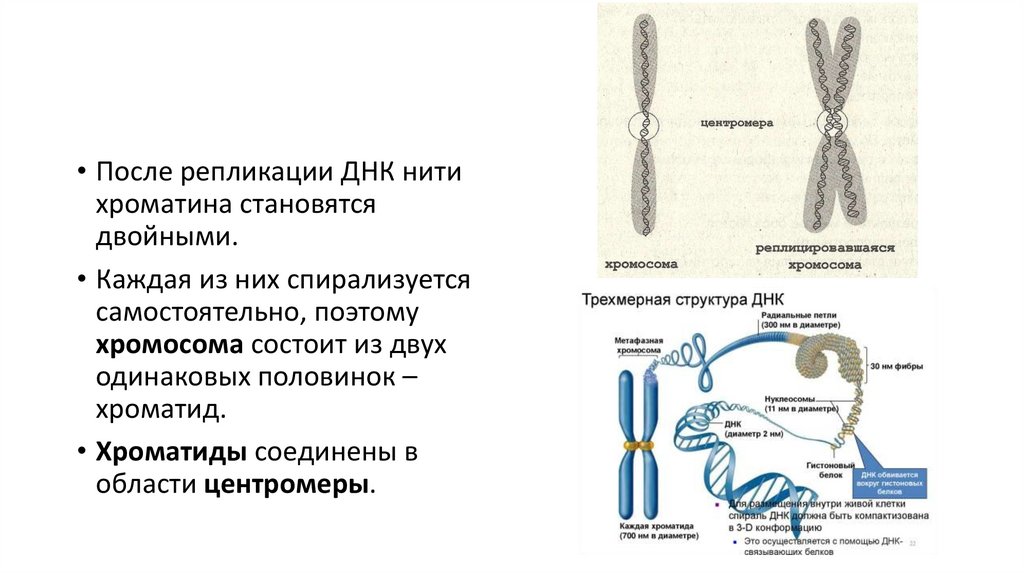
• После репликации ДНК нитихроматина становятся
двойными.
• Каждая из них спирализуется
самостоятельно, поэтому
хромосома состоит из двух
одинаковых половинок –
хроматид.
• Хроматиды соединены в
области центромеры.
98.
99.
100.
101.
Хромосомный набор и кариотип• Количество и строение
хромосом видоспецифично.
• Хромосомный набор с
особенностями строения
каждой хромосомы
называется кариотипом.
102.
• В половых клетках находитсяодинарный или гаплоидный
набор хромосом (n) , т.е.
каждая хромосома в этом
наборе уникальна.
• Гаплоидный набор хромосом
содержит уникальный набор
генов, отвечающих за все
признаки организма (ГЕНОМ)
103.
• После оплодотворения взиготе образуется
диплоидный или двойной
набор (2n), который
характерен и для
соматических клеток.
• Диплоидный набор состоит из
парных или гомологичных
хромосом.
104.
• В одинаковых локусахгомологичных хромосом
находятся гены, отвечающие
за один и тот же признак –
аллельные гены.
105.
Биология растительнойклетки
106.
Особенности растительной клетки• Аппарат Гольджи не
централизован, чаще находится в
кортексе, участвуя в диффузном
механизме роста
• Цитоскелет: микротрубочки
присоединены к растущей
поверхности клетки; актиновые
филаменты находятся внутри,
обеспечивая циклоз
• Отсутствуют центросомы
(центриоли)
107.
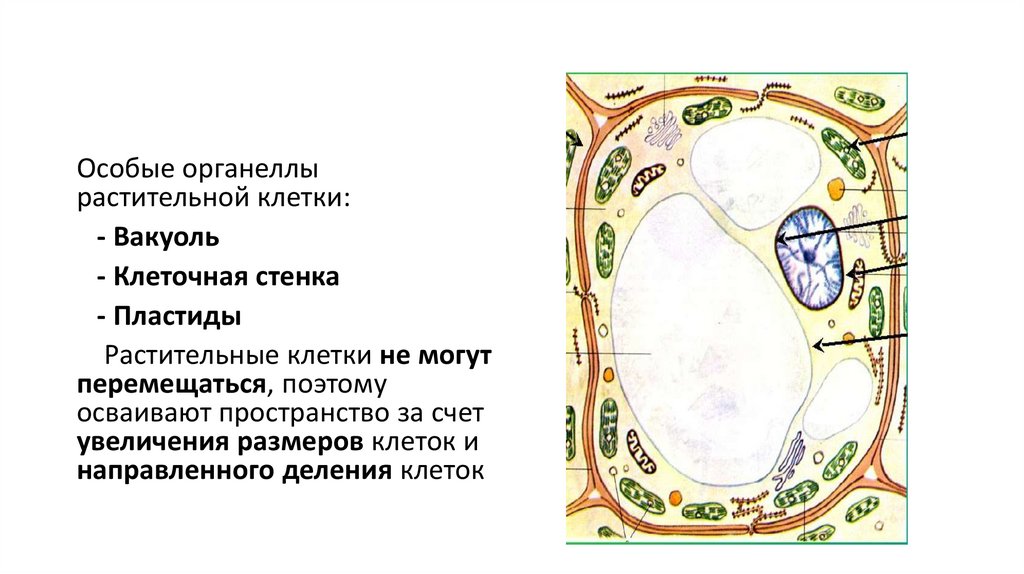
Особые органеллырастительной клетки:
- Вакуоль
- Клеточная стенка
- Пластиды
Растительные клетки не могут
перемещаться, поэтому
осваивают пространство за счет
увеличения размеров клеток и
направленного деления клеток
108.
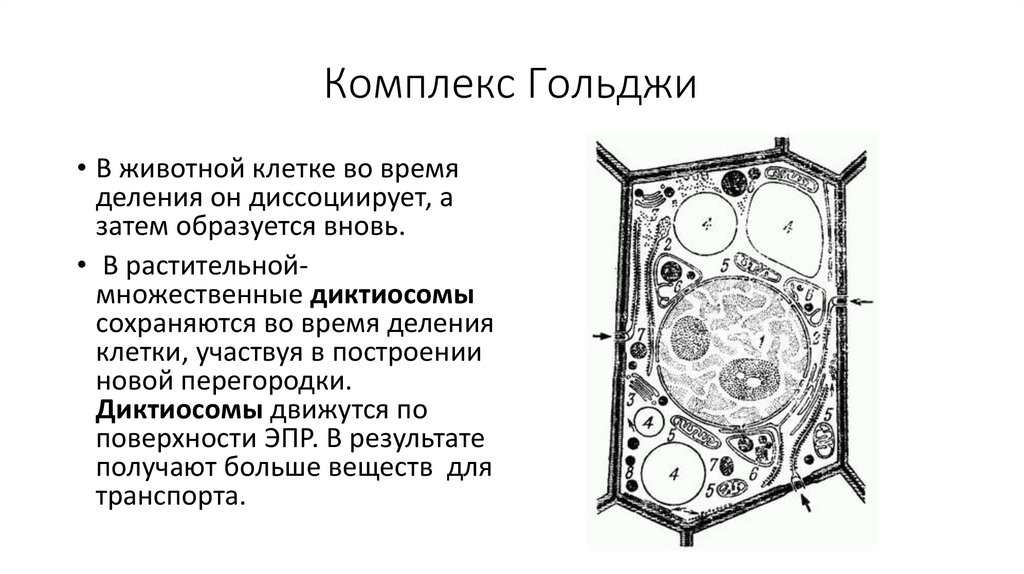
Комплекс Гольджи• В животной клетке во время
деления он диссоциирует, а
затем образуется вновь.
• В растительноймножественные диктиосомы
сохраняются во время деления
клетки, участвуя в построении
новой перегородки.
Диктиосомы движутся по
поверхности ЭПР. В результате
получают больше веществ для
транспорта.
109.
Функции аГ: - гликозилирование белков (как и в жив.кл.)- сортинг белков (как и в жив.кл.)
- синтез полисахаридов клеточной стенки (кроме
целлюлозы и каллозы) и экспорт в виде везикул
- сборка гекзогональных розеток ферментов для
синтеза целлюлозы
- в клетках семян доставляет белки и
гидролитические ферменты в вакуоли ( белки гидролизуются до
аминокислот и используются для роста)
110.
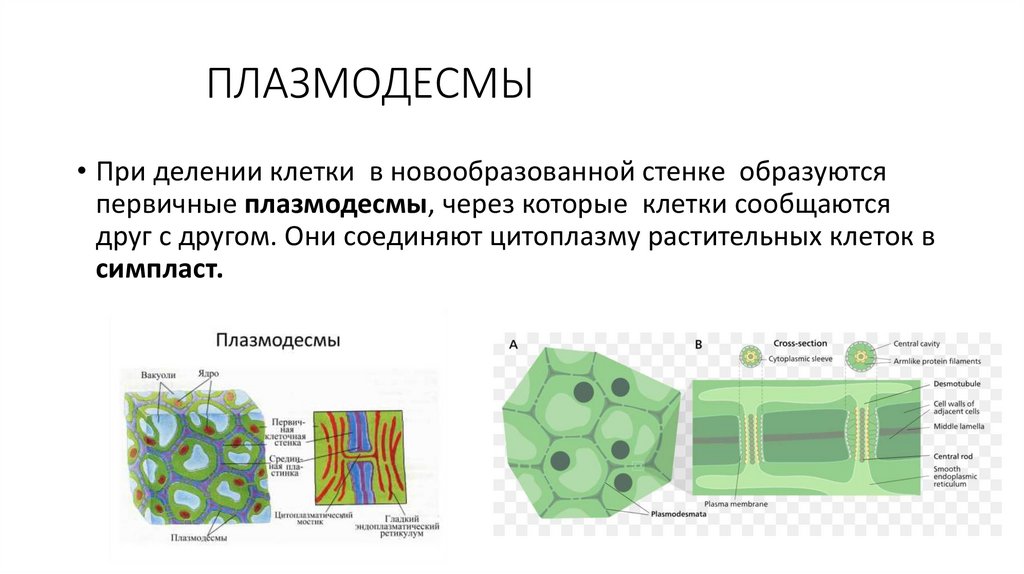
ПЛАЗМОДЕСМЫ• При делении клетки в новообразованной стенке образуются
первичные плазмодесмы, через которые клетки сообщаются
друг с другом. Они соединяют цитоплазму растительных клеток в
симпласт.
111.
• Вещества могут перемещаться через плазмодесмы с помощьюактиновых филаментов, образующих сеть. Эта сеть прикреплена
к клеточной стенке миозином (в раст кл.- особая форма миозина).
• Через плазмодесмы м. перемещаться и вирусы (н-р, ВТМ).
112.
ВАКУОЛИ• После деления клетки начинается рост клетки в зоне растяжения.
Происходит с участием вакуолей (в делящихся клетках часто
отсутствуют). Растяжение происходит в определенном
направлении
могут формироваться вытянутые клетки – прозенхимные
или изодиаметричные - паренхимные
113.
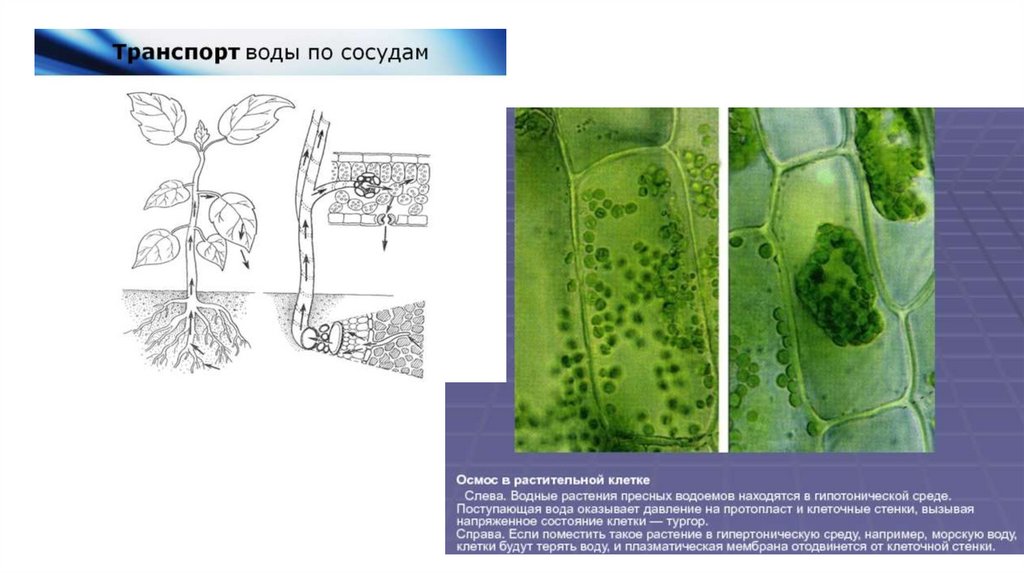
• Вода поступает в растительную клетку по законам диффузии иосмоса, стремясь разбавить более концентрированную
внутреннюю среду.
• Затем она попадает в вакуоль, где находится более
концентрированный вакуолярный сок. Вакуоль растягивается, ее
мембрана (тонопласт) расправляет клетку, создавая тургорное
давление ,н-р, в клетках тюльпана равно 0,6 мПа. Для сравненияв автошинах – 0,2мПА.
• Вакуоль может занимать до 95% объема клетки.
114.
115.
116.
Другие функции вакуоли• Детоксицирующая – разрушение ядов, поступающих извне
• Хранение ядов, которые продуцируют сами
• Хранение пигментов
• Временное депо для аминокислот и ионов
• Могут образовывать литические и пищеварительные вакуоли,
аналогичные лизосомам животной клетки
• Запасающая функция (н-р, в семенах)
• Функция осмотического насоса при транспирации
117.
КЛЕТОЧНАЯ СТЕНКА• Противостоит растягивающему усилию вакуоли. Причем клетка
управляет этим растяжением.
• Если внеклеточный матрикс у животных состоит
преимущественно из белков, то у растений – из углеводов
• Белки- лишь 10% : гликопротеины, ферменты для синтеза
компонентов клеточной стенки, структурный белок экстензин.
118.
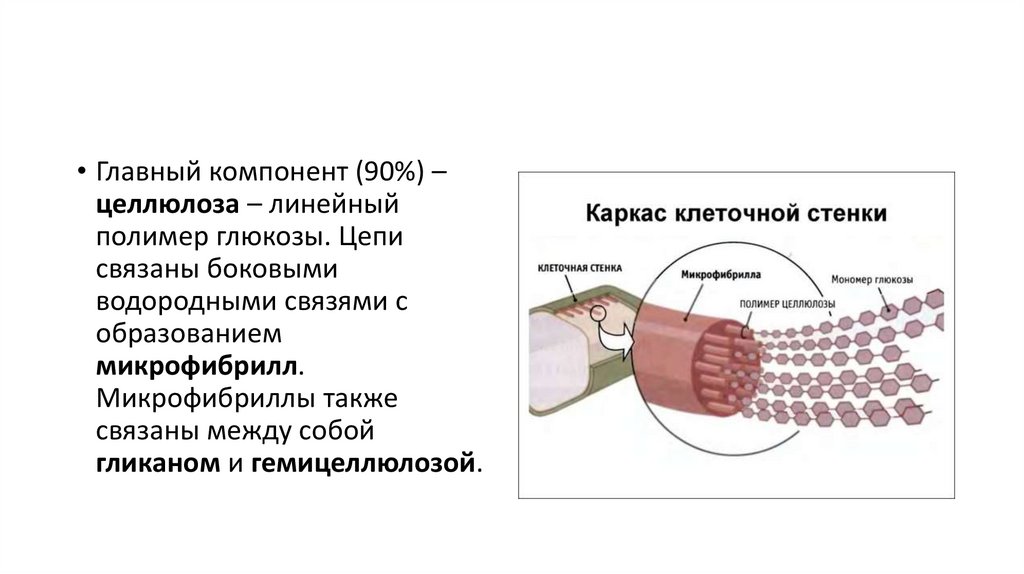
• Главный компонент (90%) –целлюлоза – линейный
полимер глюкозы. Цепи
связаны боковыми
водородными связями с
образованием
микрофибрилл.
Микрофибриллы также
связаны между собой
гликаном и гемицеллюлозой.
119.
• Сеть микрофибриллпогружена в гелеобразный
гидратированный пектин (не
пропускает крупные молекулы,
но пропускает воду и
небольшие молекулы,
связывает кальций).
120.
• Клеточная стенка состоит из концентрических слоев – ламеллей.Микрофибриллы разных слоев не пересекаются. Наиболее
старые слои находятся снаружи, отодвигаемые вновь
образующимися изнутри слоями.
121.
122.
• Ориентация микрофибрилл в клеточной стенки определяетнаправление ее растяжения.
Если они обернуты вокруг клетки поперек,
вытягивание клетки происходит в перпендикулярном направлении.
123.
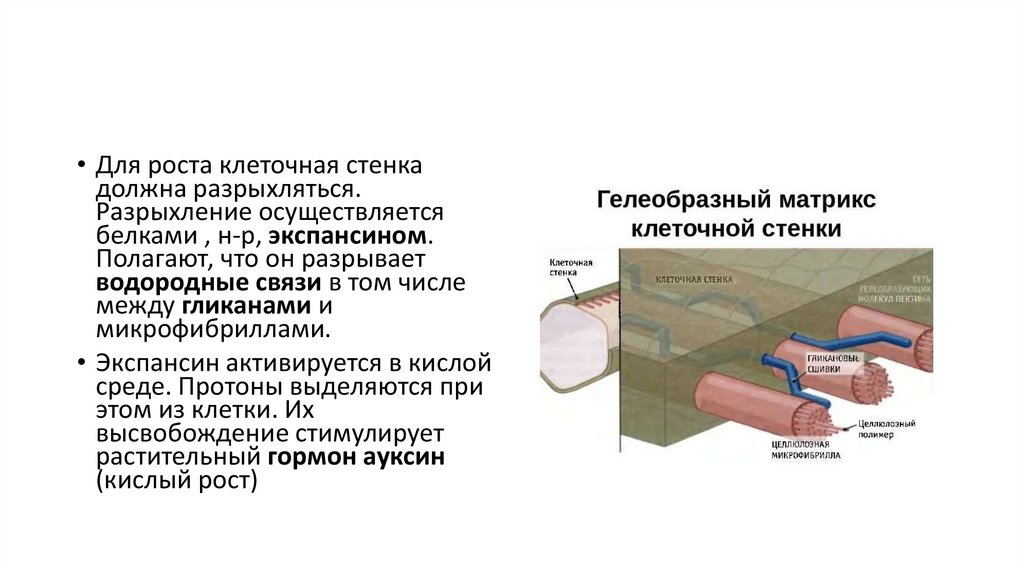
• Для роста клеточная стенкадолжна разрыхляться.
Разрыхление осуществляется
белками , н-р, экспансином.
Полагают, что он разрывает
водородные связи в том числе
между гликанами и
микрофибриллами.
• Экспансин активируется в кислой
среде. Протоны выделяются при
этом из клетки. Их
высвобождение стимулирует
растительный гормон ауксин
(кислый рост)
124.
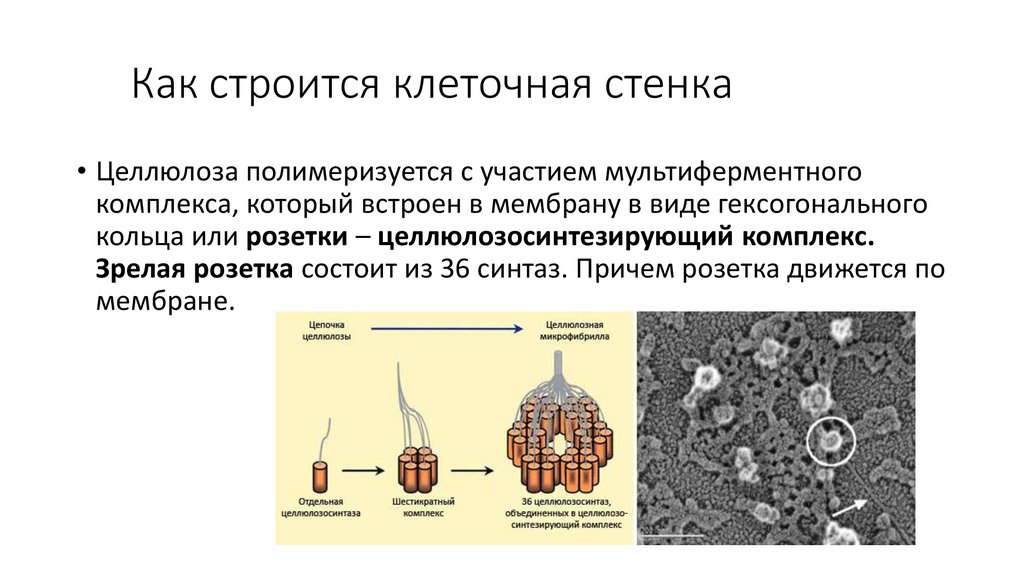
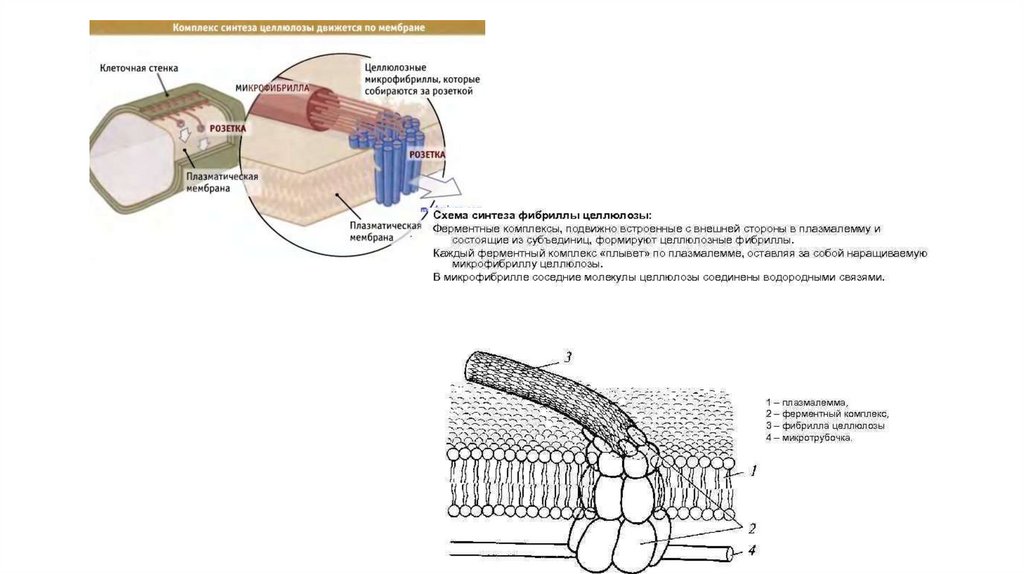
Как строится клеточная стенка• Целлюлоза полимеризуется с участием мультиферментного
комплекса, который встроен в мембрану в виде гексогонального
кольца или розетки – целлюлозосинтезирующий комплекс.
Зрелая розетка состоит из 36 синтаз. Причем розетка движется по
мембране.
125.
126.
• Другие компоненты клеточной стенки доставляются из клетки впузырьках комплекса Гольджи; поэтому комплекс Гольджи
находится на периферии цитоплазмы.
127.
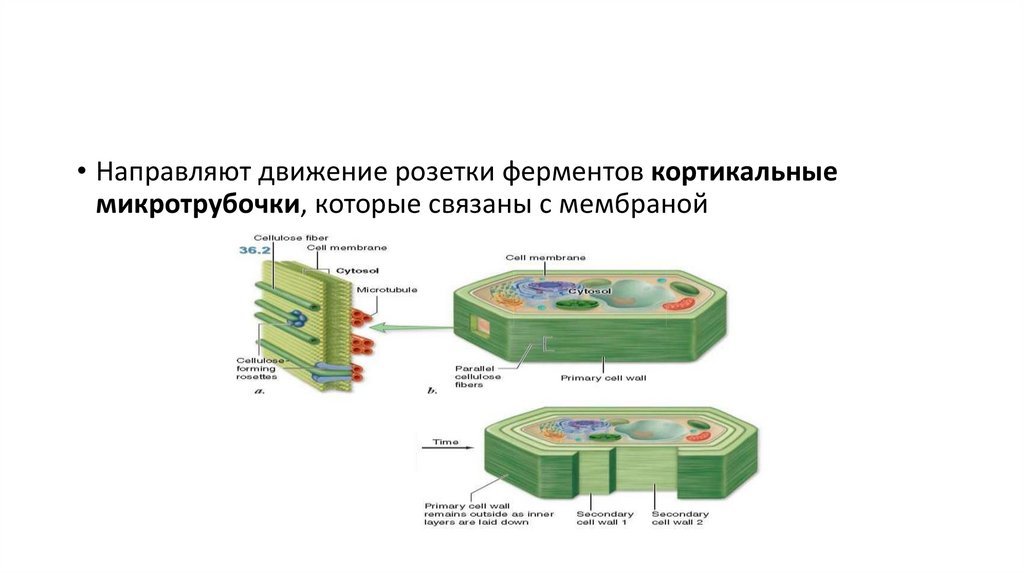
• Направляют движение розетки ферментов кортикальныемикротрубочки, которые связаны с мембраной
128.
• Микротрубочки обеспечивают также направленное растяжениеклеток. Поэтому, если клетки корня обработать колхицином или
каким-либо гербицидом, происходит деполимеризация
микротрубочек и направленный рост корня прекращается.
Корень растет во все стороны и превращается в бесформенное
скопление клеток.
129.
Влияние различных факторов намикротрубочки
• Газ этилен действует как гормон растений. Вырабатывается в клетках и
высвобождается при повреждении тканей. Если растения поместить в
атмосферу этилена, то их рост задерживается вследствие
переориентации Мктр. Образуются короткие растения с толстыми
стеблями.
• Гибберриллиновая кислота удлиняет корень, ориентируя Мктр
соответствующим образом.
• Свет и гравитация также влияют на положение Мктр (?).Показано,что
разная освещенность стебля, листьев приводит к быстрой элонгации
мктр с одной стороны и их разборке с другой стороны
поворот побега в сторону солнца.
130.
Циклоз• Осуществляется с участием
актиновых филаментов и
миозинов. При этом вдоль
актиновых филаментов
мигрируют и органеллы
(в животной клетке- вдоль
Мктр).
131.
Пластиды132.
133.
Типы пластид• Хлоропласты в зеленых частях растений с хлорофиллом
• Хромопласты – накапливают красный, оранжевый и желтый
пигменты ( каротиноиды)
• Лейкопласты подразделяются на
- амилопласты (накапливают углеводы)
- протеинопласты (накапливают белки)
- олейопласты (накапливают липиды)
134.
135.
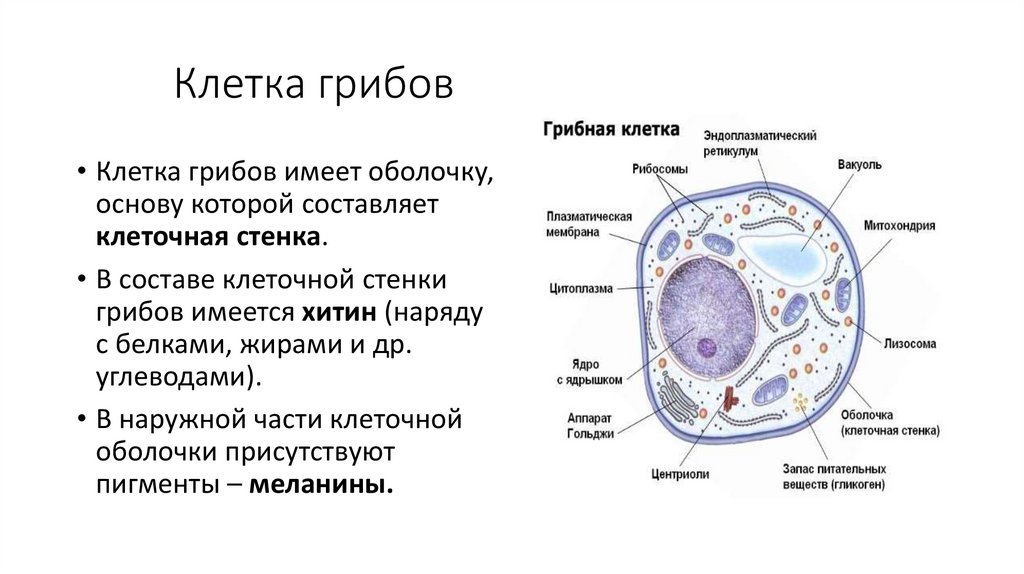
Клетка грибов• Клетка грибов имеет оболочку,
основу которой составляет
клеточная стенка.
• В составе клеточной стенки
грибов имеется хитин (наряду
с белками, жирами и др.
углеводами).
• В наружной части клеточной
оболочки присутствуют
пигменты – меланины.
136.
• Клетки грибов объединяются,образуя тонкие нити – гифы.
• Гифы имеют общую клеточную
стенку из хитина.
• Гифы чаще не имеют истинного
клеточного строения. Они
разделены перегородками –
септами. В отличие от истинной
клеточной стенки образование септ
не связано с делением.
• В септах имеются отверстия, через
которые цитоплазма м. перетекать
из одного компартмента в другой.
137.
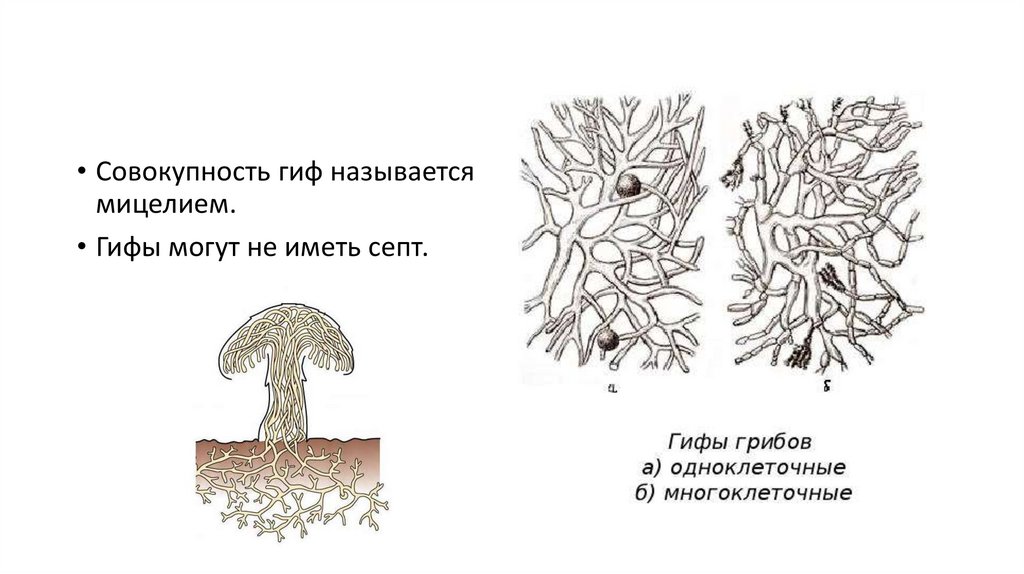
• Совокупность гиф называетсямицелием.
• Гифы могут не иметь септ.
138.
139.
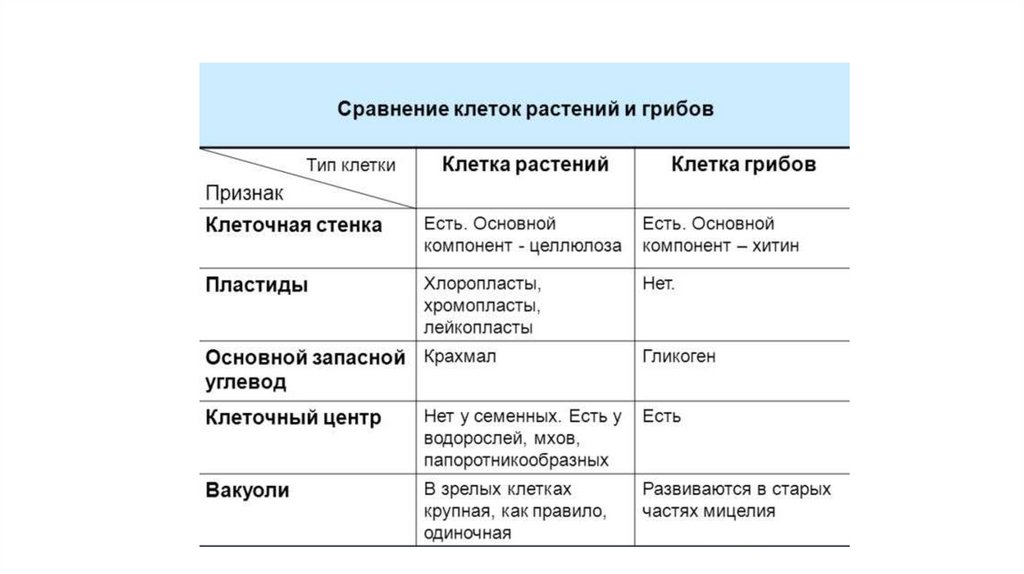
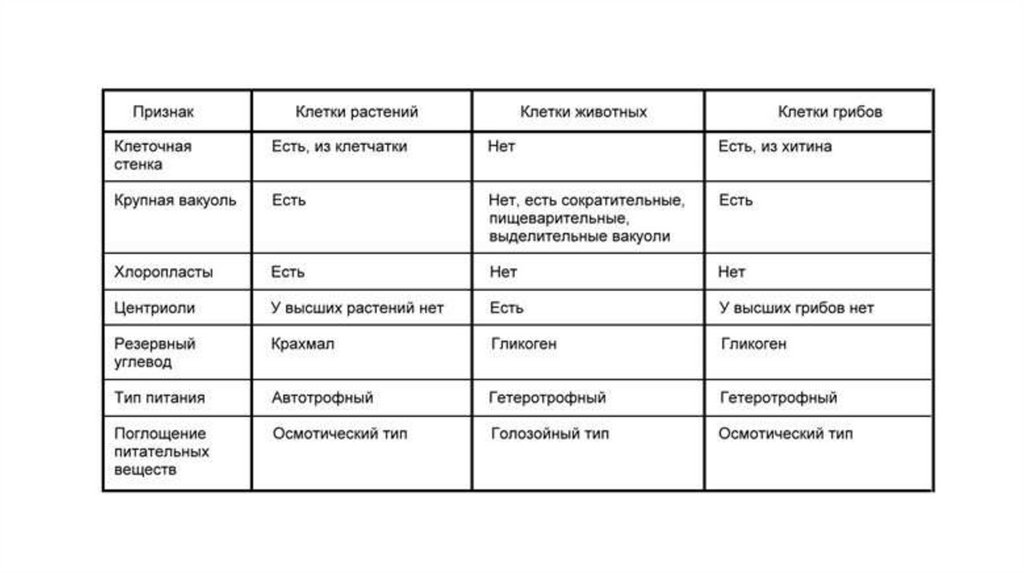
Сходство грибов и растенийСходство грибов и животных
140.
141.
142.
Биологияпрокариотической клетки
143.
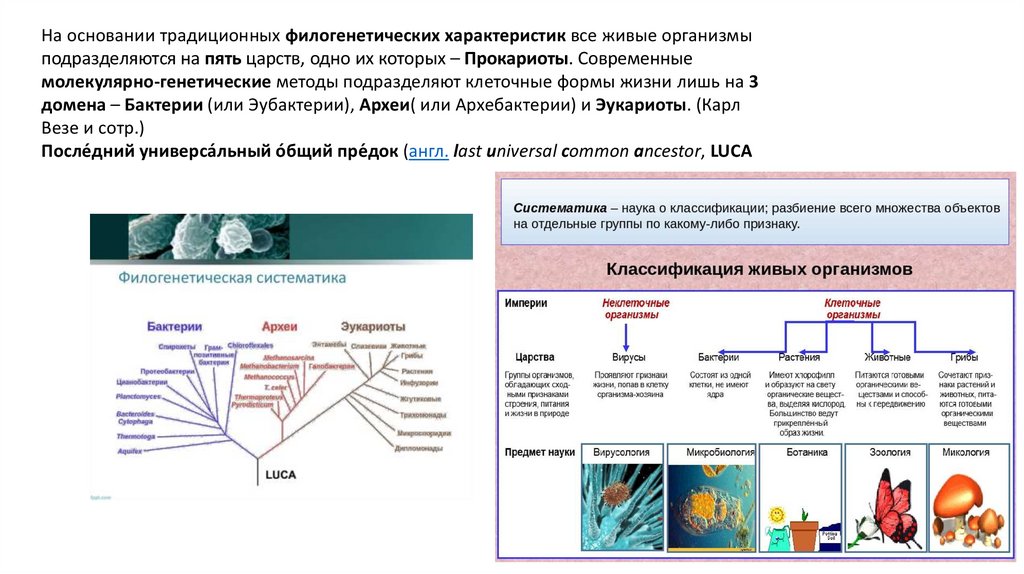
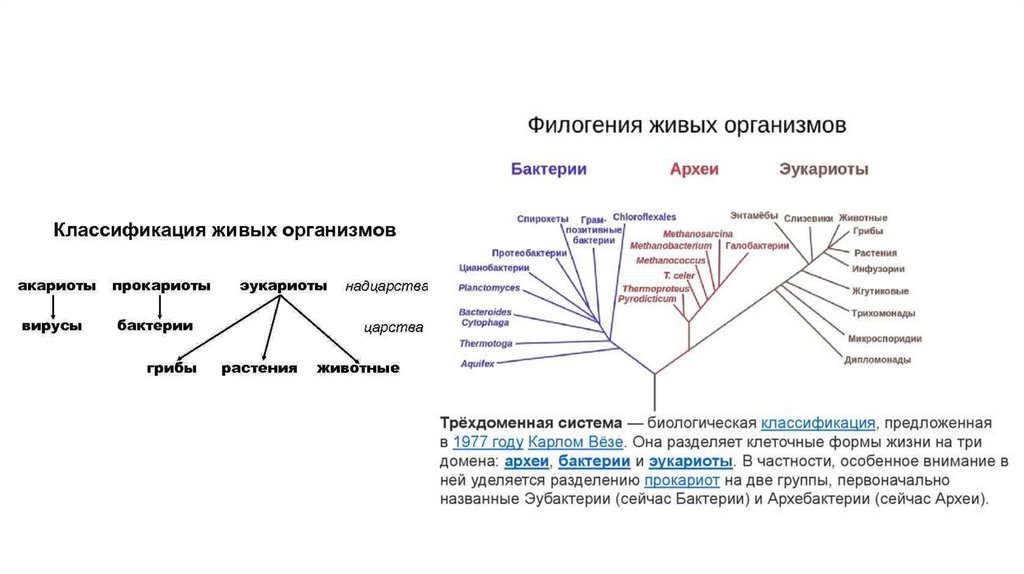
На основании традиционных филогенетических характеристик все живые организмыподразделяются на пять царств, одно их которых – Прокариоты. Современные
молекулярно-генетические методы подразделяют клеточные формы жизни лишь на 3
домена – Бактерии (или Эубактерии), Археи( или Архебактерии) и Эукариоты. (Карл
Везе и сотр.)
После́дний универса́льный о́бщий пре́док (англ. last universal common ancestor, LUCA
144.
145.
• Данные о строении прокариотической клетки были получены восновном на двух объектах - Echerichia coli (грамотрицательная)
Bacillus subtilis (грамположительная). При этом об огромном
количестве бактерий ,населяющих Землю, мы ничего не знаем.
• Трудности изучения связаны с тем, что 99% прокариот не
поддаются культивированию.
146.
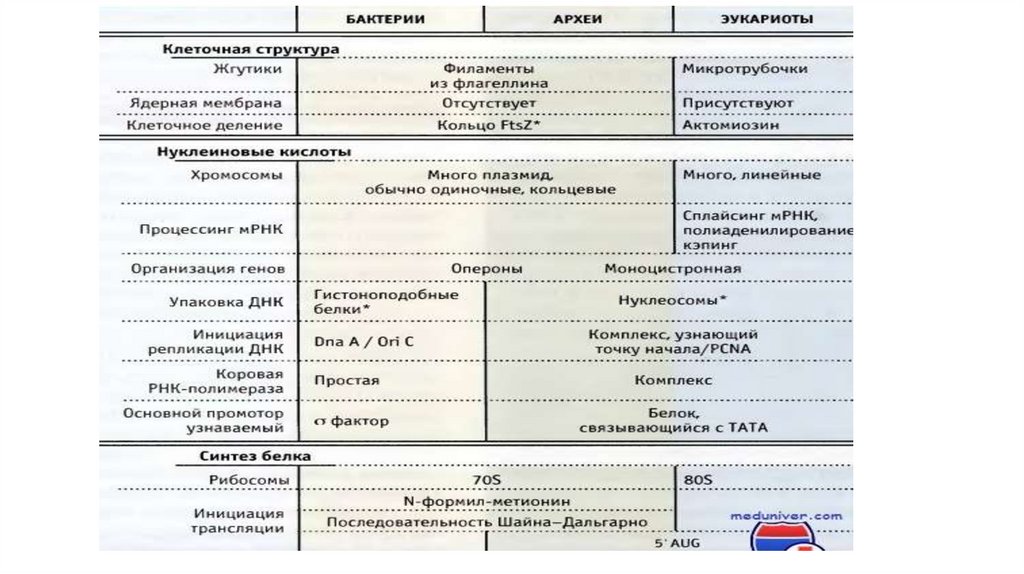
Особенности прокариотической клетки• Отсутствие оформленного ядра.
• Обычно содержат одну кольцевую молекулу ДНК (вместе со
связанными белками образует нуклеоид).
• Не содержат внутренних мембран (кроме фотосинтезирующих
бактерий - цианобактерий)
• Небольшие размеры
• Наличие клеточной стенки и капсулы
• Более мелкие рибосомы
147.
• Жгутик из филаментов белка флагеллина.• ДНК вне нуклеоида – плазмиды.
• Оперонная структура генов.
• Отсутствует циклоз
148.
Особенности Архей по сравнению сбактериями и эукариотами
• У Архей проявляются черты сходства с Эукариотами:
- отсутствие пептидогликана в клеточной стенке
- сходство в строении генома
- упаковка ДНК с образованием нуклеосом
- комплексная РНК-полимераза
- стартовый кодон 5 ′ AUG
149.
150.
• Археи характеризуются универсальностью метаболизма испособностью существовать в экстремальных условиях (высокая
температура, высокие или низкие значения рН, высокая
концентрация соли - галофилы)
• Многие имеют особые метаболические процессы: могут
использовать при дыхании соединения серы, соединения с
метильной группой или ацетат - хемосинтезирующие.
151.
152.
Оболочка Прокариот• Включает
- цитоплазматическую мембрану,
- клеточную стенку различного состава
- капсулу
- у грамотрицательных бактерий имеется наружная мембрана.
При этом их клеточная стенка тонкая и не окрашивается по Грамму
фиолетовым красителем.
153.
Большинство прокариот на поверхностиклеточной стенки образуют капсулу или
слизистый слой
• Можно сравнить с гликокаликсом животной клетки, т.к. состоит
из углеводов ( разнообразных полисахаридов) и белков.
• Обладает антигенными свойствами. Н-р, у E.coli описано более
80 капсулярных полисахаридов или К-антигенов (в том числе
связанных с патогенными штаммами)
154.
Функции капсулы• Защита от высыхания
• Прикрепление бактерий к клеткам хозяина, внешним
поверхностям и связывание друг с другом в биопленке
(Pseudomonas aeroginosa образует такую биопленку в легких
человека – защита от иммунной системы и антибиотиков)
• Защита от фагоцитов, детергентов, бактериофагов
155.
Капсула может становиться основным вирулентным факторомпатогенных бактерий:
- Bacillus anthracis (возбудитель сибирской язвы). Капсула состоит из
пептидов. Синтезируется in vivo. Штаммы неспособные синтезировать
капсулу обладают слабой патогенностью. При этом капсула защищает
микроорганизм от атаки иммунной системы человека.
156.
• - Mycobacterium tuberculosus (возбудитель туберкулеза) –капсула обеспечивает связывание с макрофагом и
проникновение в него. В составе капсулы при этом есть особые
защитные факторы ( бескапсульные бактерии обладают меньшей
вирулентностью и используются для изготовления вакцины)
157.
158.
Клеточная стенка бактерий• У большинства бактерий основным компонентом клеточной
оболочки является пептидогликан (муреин). Он выполняет
формообразующую функцию и противостоит высокой
внутренней осмолярности клетки.
• Предшественниками пептидогликана являются пентапептиды
дисахаридной природы . Они образуются в цитоплазме,
экспортируются из клетки и собираются над цитоплазматической
мембраной.
159.
• Гликановые нити состоят из повторяющихся дисахаридов (вколичестве около 30) - N -ацетилглюкозамин-N ацетилмурамовой кислоты (NAG-NAM) - муреин.
• К ним присоединяется олигопептид, который синтезируется вне
рибосом и содержит необычные аминокислоты (н-р, Dглутаминовую). Пептид сшивает гликановые нити.
• Антибиотики пенициллинового ряда связывают эти пептиды и
т.о. разрушают клеточную стенку бактерий.
160.
У грамположительныхбактерий имеется несколько
слоев петидогликана,
у грамотрицательных –
один.
161.
У грамположительных бактерий пептидогликаны клеточнойстенки могут связываться с фосфолипидами и белками в составе
мембраны.
Действие многих антибиотиков основано на подавлении роста
клеточной стенки бактерий.
162.
Наружная мембрана грамотрицательныхбактерий
• У грамотрицательных бактерий имеется наружная мембрана
наряду с цитоплазматической мембраной. Мембраны отличаются
по составу. Наружная мембрана асимметрична. Ее внутренний
слой состоит из фосфолипидов, а наружный – из
липополисахаридов (LPS).
• LPS токсичны для животных. Поэтому они являются ведущим
фактором в патогенном действии грамотрицательных бактерий.
У человека и животных лимфоциты и клетки эпителия имеют
рецепторы, которые узнают LPS и активируют воспалительную
реакцию.
163.
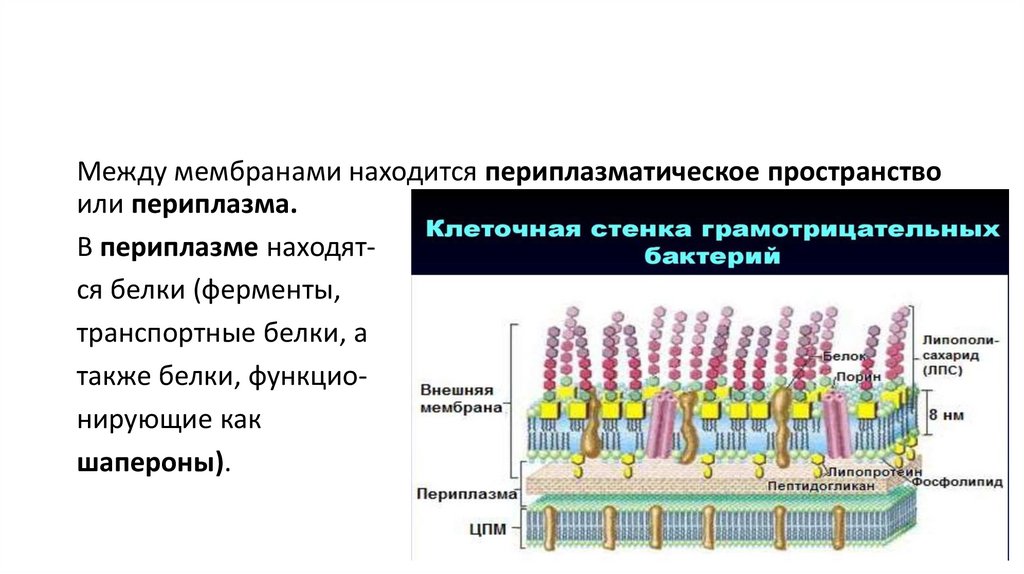
Между мембранами находится периплазматическое пространствоили периплазма.
В периплазме находятся белки (ферменты,
транспортные белки, а
также белки, функционирующие как
шапероны).
164.
Цитоплазматическая мембрана и ее рольв транспорте
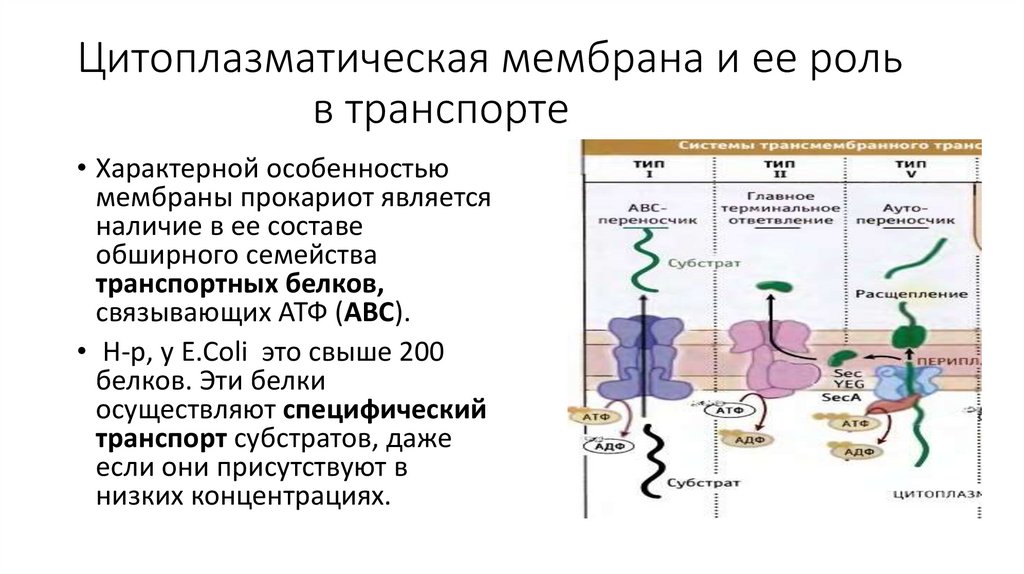
• Характерной особенностью
мембраны прокариот является
наличие в ее составе
обширного семейства
транспортных белков,
связывающих АТФ (АВС).
• Н-р, у E.Coli это свыше 200
белков. Эти белки
осуществляют специфический
транспорт субстратов, даже
если они присутствуют в
низких концентрациях.
165.
• Через мембрану транспортируются протоны, так что снаружимембрана имеет слабый положительный заряд , а внутриотрицательный. Возникает протонный градиент. Эта энергия
используется для синтеза АТФ, синтеза молекул, транспорта
белков, движения. Изучение этих механизмов продолжается
166.
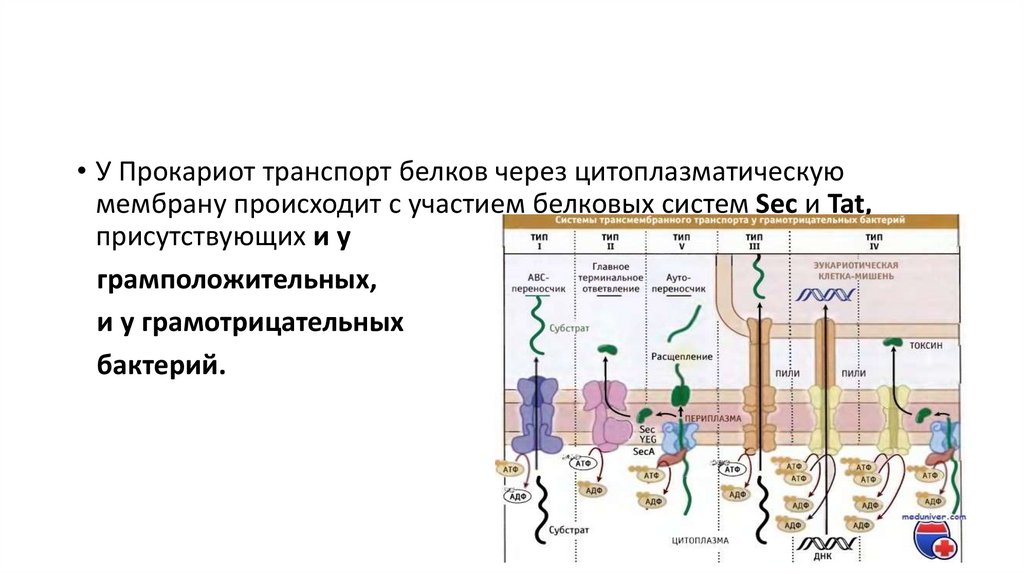
• У Прокариот транспорт белков через цитоплазматическуюмембрану происходит с участием белковых систем Sec и Tat,
присутствующих и у
грамположительных,
и у грамотрицательных
бактерий.
167.
• У грамотрицательныхбактерий существуют особые
транспортные системы (тип lll),
которые секретируют белки не
только через
цитоплазматическую
мембрану, но и через
наружную мембрану.
168.
• Одна из этих системтранспортирует субстраты из
цитоплазмы клетки бактерий
непосредственно в
цитоплазму клетки хозяина.
Таким путем переносятся
многие вирулентные
факторы-молекулы, которые
позволяют бактериям
инфицировать организм и
размножаться в нем.
169.
Система секреции белков типа lVтакже секретируют субстраты
непосредственно в клетку
хозяина, н-р, коклюшный токсин.
Эта система может переносить
не только белки, но и ДНК.
170.
ПИЛИ ПРОКАРИОТ• Пили - внеклеточные белковые структуры, которые опосредуют
разнообразные процессы:
- обмен ДНК
- адгезию
- образование биопленки клетками Прокариот
Впервые были обнаружены F-пили, которые участвуют в переносе
ДНК и в клеточной агрегации. Их присутствие коррелировало со
способностью E.сoli агглютинировать эритроциты. Сначала
называли фимбрии (от fimbria – нить, волокно) , а потом назвали
пили ( pilus – волос).
171.
• Основная функция пилей состоит в позиционированииспецифических молекул, участвующих в клеточной адгезии.
• Н-р, при инфекциях мочевых путей патогенными E.сoli они
прикрепляются к клеткам мочевого пузыря пилями.
• Структурные компоненты пилей перемещаются через
цитоплазматическую мембрану в периплазму. Сборка
происходит здесь с участием шаперонов. Затем комплекс
перемещается через наружную мембрану.
172.
Жгутики Прокариот173.
174.
Жгутик состоит из трех основных частей:- Филамент – из повторяющихся структур белка флагеллина,
расположенных по спирали.
- крючок для соединения филамента и базального тела
- базальное тело из множества белков
175.
176.
177.
Хемотаксис• Направление перемещения жгутика определяется наличием или
отсутствием питательных веществ. При отсутствии питательных
веществ жгутик вращается по часовой стрелке, что вызывает
поворот клетки.
• Направленное движение к молекулам химического соединения
называется хемотаксисом. Когда жгутик вращается против
часовой стрелки, клетка плывет по прямой линии по
направлению к пищевому аттрактанту.
• Практически у всех Прокариот обнаружены консервативные
белки хемотаксиса (кроме Mycoplasma, не имеющей клеточной
стенки)
178.
Геном прокариот• У большинства Прокариот присутствует одна кольцевая
хромосома и они гаплоидны.
• Количество пар нуклеотидов – от 580 тысяч у микоплазмы до
9 млн. пар оснований у стрептомицетов.
• Некодирующие последовательности практически отсутствуют.
• Генетическая гибкость обеспечивается мобильными элементами
(н-р, плазмидами).
• Некоторые бактерии имеют линейные хромосомы – н-р, Borrelia.
• «Гены домашнего хозяйства» находятся в кольцевой ДНК.
179.
• Несмотря на отсутствие ядерной оболочки, геном прокариотхарактеризуется высокой упорядоченностью и неслучайным
расположением генов. Он организован в виде нуклеоида,
занимающего центральную часть цитоплазмы. Он состоит из
кольцевой ДНК и связанных с ней белков. Нуклеосом нет, но
белки часто гистоноподобные.
• Белки нуклеоида E.coli : гиразы, топоизомеразы, РНКполимеразы, гистоноподобные белки, фактор интеграции в
клетки хозяина, белки конденсации ДНК.
180.
181.
- Плазмиды, определяющие способность синтезироватьантибиотики (Streptomyces)
- Плазмиды, определяющие устойчивость к тяжелым металлам
- Плазмиды, определяющие способность к симбиозу (нодуляция)
и фиксации азота
- Плазмиды, определяющие устойчивость к ультрафиолету
(сальмонелла)
- Плазмиды, определяющие способность использовать
различные источники углерода (н-р, толуола)
182.
Плазмиды – стабильные элементывнехромосомной ДНК
183.
• Плазмиды могут распространяться различными путями:- Конъюгация – напоминает спаривание. В донорской плазмиде
закодированы функции, необходимые для обеспечения контакта и
переноса плазмиды, н-р, пили.
- Трансформация – прямой захват ДНК
- Трансдукция – с помощью бактериофага. При этом геном
бактериофага может сохраняться в бактерии, превращаясь в
плазмиду (профаг)
184.
• Бактериальный геном содержит множество мобильныхэлементов, распространяющихся путем транспозиции –
транспозоны.
• Наиболее известные транспозоны у бактерий – гены ,
определяющие их устойчивость к антибиотикам.
185.
186.
РЕПЛИКАЦИЯ У ПРОКАРИОТ187.
188.
• ДНК-хеликаза раскручивает двойную спираль со скоростью 1000пар нуклеотидов в секунду . Возникают супервитки впереди от
хеликазы. Их необходимо убрать. Осуществляет это
топоизомераза.
• Single strand DNA-binding proteins – SSB у прокариот (RPA replication protein A у эукариот) препятствуют образованию
«шпилек на одноцепочечной ДНК»
189.
190.
• Пространственное разделение образующихся молекул ДНК уE.coli находится под контролем особого белка FtsK . Он
необходим
- для образования септы деления
- способен транслоцировать ДНК
- приводит к разделению молекул ДНК и обеспечивает их
распределение в новые клетки
Несмотря на отсутствие митотического веретена деления,
сегрегация хромосом у прокариот происходит безошибочно.
Механизмы сегрегации изучены недостаточно.
191.
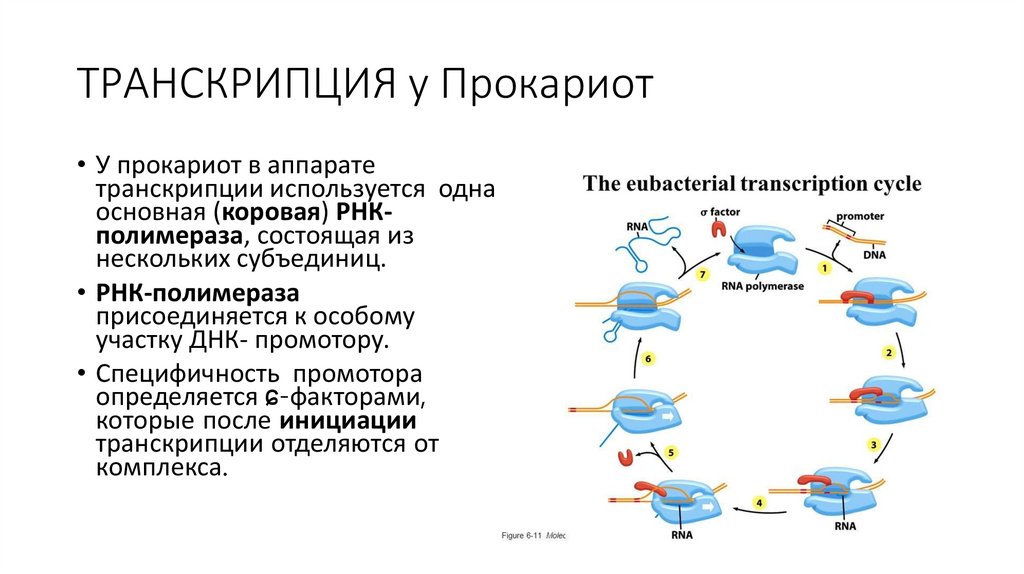
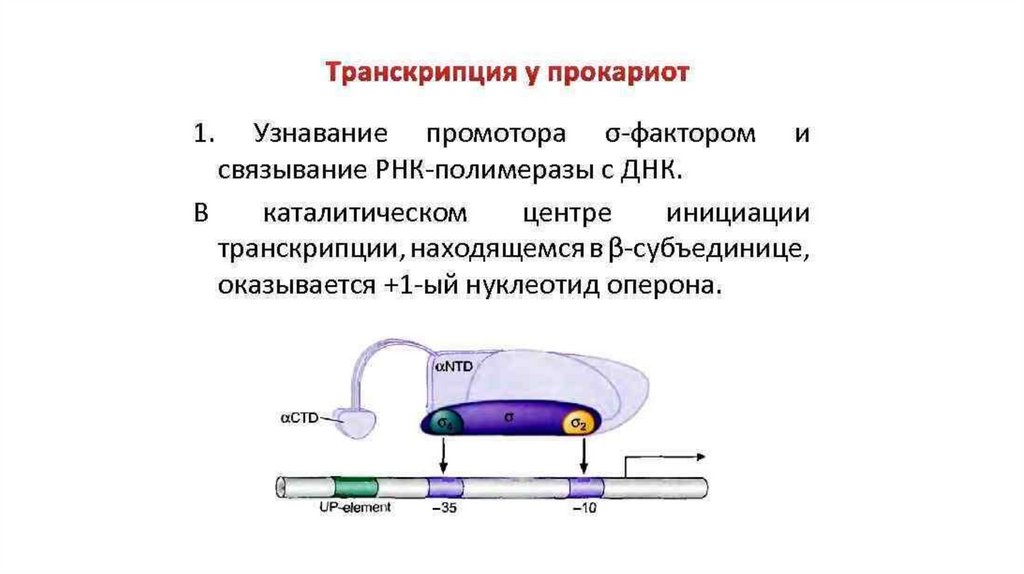
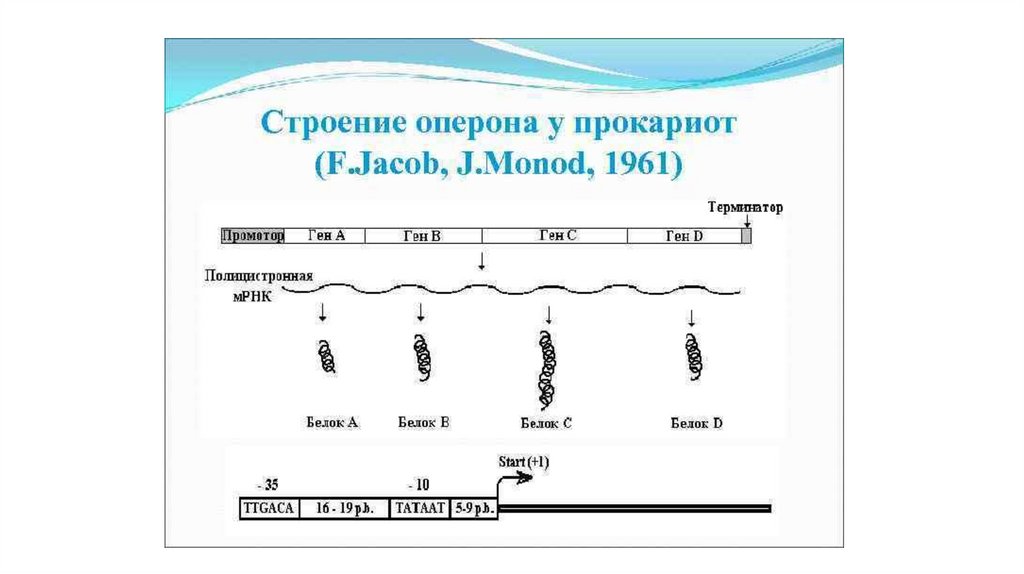
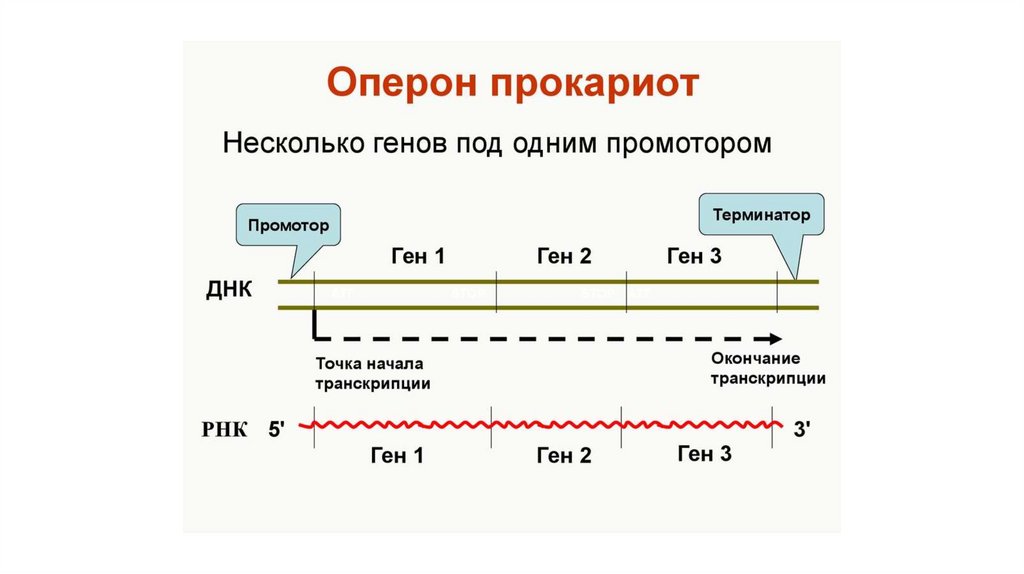
ТРАНСКРИПЦИЯ у Прокариот• У прокариот в аппарате
транскрипции используется одна
основная (коровая) РНКполимераза, состоящая из
нескольких субъединиц.
• РНК-полимераза
присоединяется к особому
участку ДНК- промотору.
• Специфичность промотора
определяется ɕ-факторами,
которые после инициации
транскрипции отделяются от
комплекса.
192.
• Большая часть молекул коровой РНК- полимеразы находится вцентре нуклеоида. Очевидно, именно здесь происходит
транскрипция.
193.
194.
195.
196.
197.
198.
199.
ТРАНСЛЯЦИЯ У ПРОКАРИОТРибосомы и белки, участвующие в трансляции, находятся на
периферии нуклеоида. Очевидно, именно здесь начинается
процесс трансляции. Однако, вопрос до конца не изучен.
200.
201.
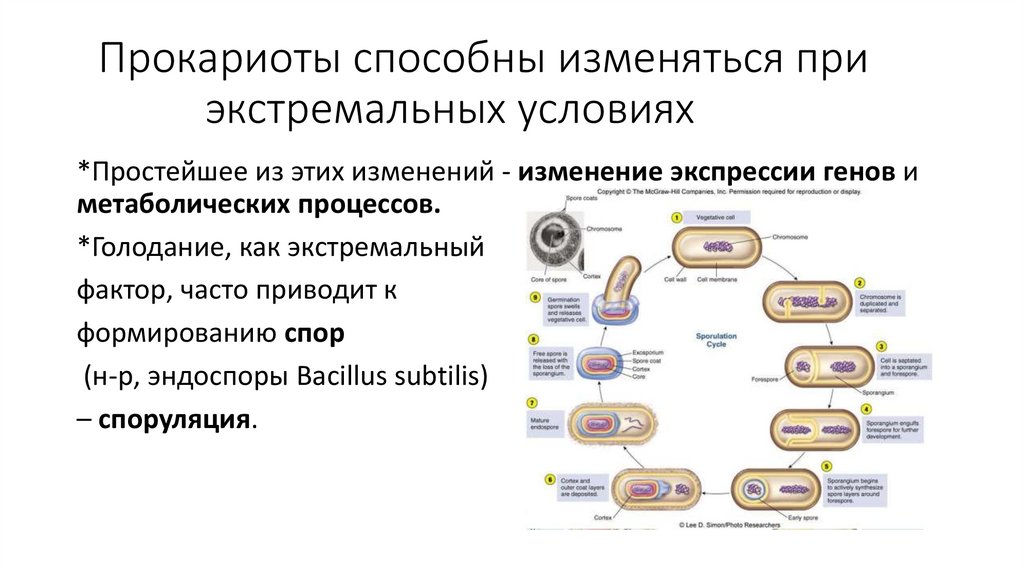
Прокариоты способны изменяться приэкстремальных условиях
*Простейшее из этих изменений - изменение экспрессии генов и
метаболических процессов.
*Голодание, как экстремальный
фактор, часто приводит к
формированию спор
(н-р, эндоспоры Bacillus subtilis)
– споруляция.
202.
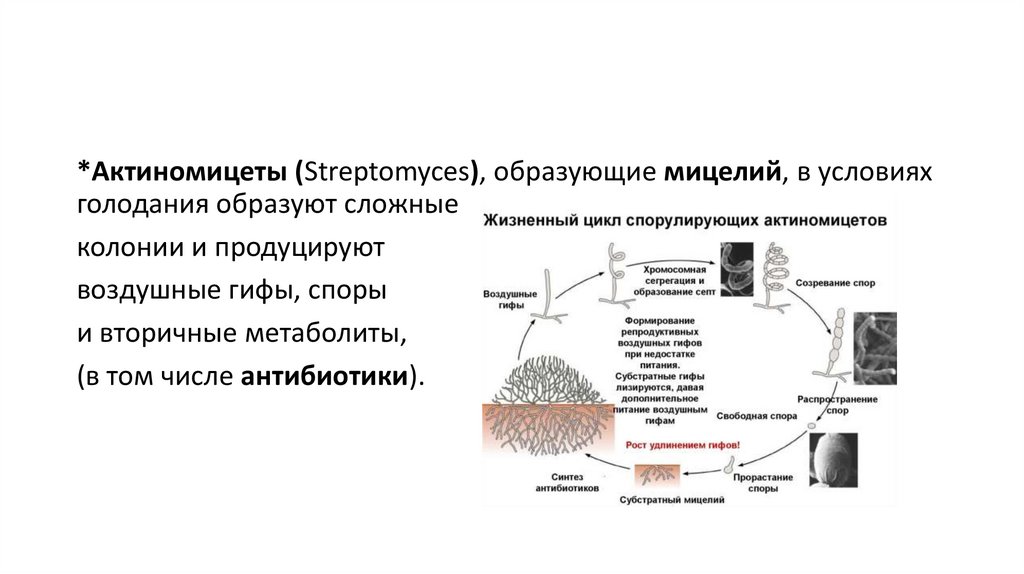
*Актиномицеты (Streptomyces), образующие мицелий, в условияхголодания образуют сложные
колонии и продуцируют
воздушные гифы, споры
и вторичные метаболиты,
(в том числе антибиотики).
203.
*У миксобактерий при недостатке питательных веществотдельныеклетки объединяются в
плодовые тела, устойчивые к
обезвоживанию. Комплексы
плодовых тел образуют ветвящиеся
ножки со спорами. При этом
существует межклеточная система
передачи сигналов, контролируемая
определенными генами.
204.
ЭНДОСИМБИОЗ*Митохондрии и пластиды образовались в результате интеграции
свободноживущих прокариот в цитоплазму простейших
эукариотических клеток, которые стали постоянными
эндосимбионтами.
*Эндосимбиоз азотфиксирующих
бактерий рода Rhizobium и растений (клубеньки бобовых).
205.
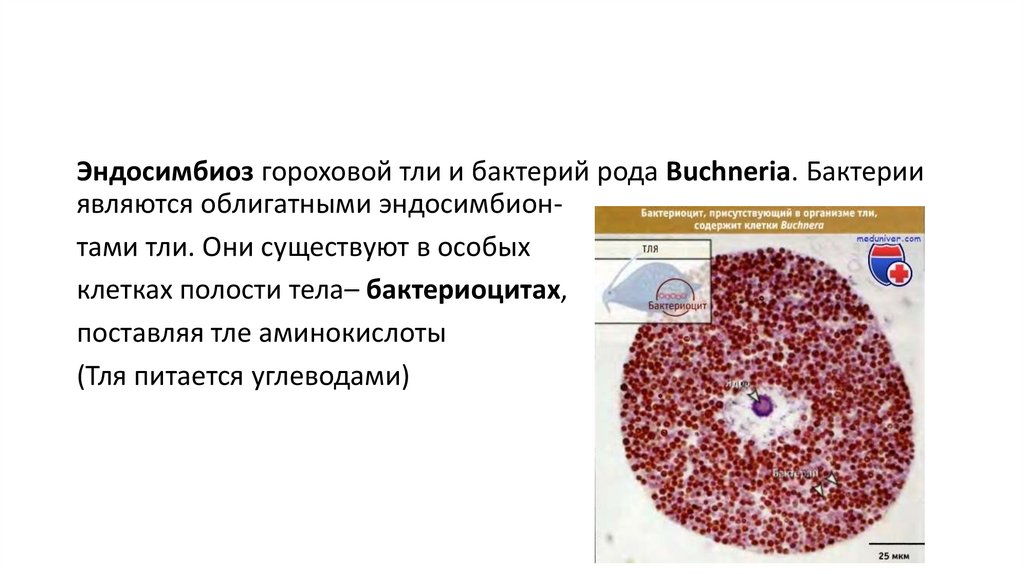
Эндосимбиоз гороховой тли и бактерий рода Buchneria. Бактерииявляются облигатными эндосимбионтами тли. Они существуют в особых
клетках полости тела– бактериоцитах,
поставляя тле аминокислоты
(Тля питается углеводами)
206.
Симбиоз бактерий и эукариот• В зависимости от степени выгоды и вреда симбиоз между
хозяином и микроорганизмом может быть
- взаимным (н-р, эндосимбионты, бактерии в кишечнике
жвачных)
- комменсальным (слабо влияют на организм хозяина или не
влияют). Но комменсалы могут быть очень полезны, как, н-р,
микрофлора кишечника.
- паразитическим – незначительный вред
- патогенным – выраженный вред (н-р, сибирская язва, чума)
207.
208.
209.
БИОПЛЕНКИ• Большинство прокариот на Земле образуют сообщества –
биопленки – грибообразные колонии, пронизанные системой
каналов.
• Образование биопленки включает несколько этапов:
прикрепление к субстрату, рост и деление микроорганизмов,
синтез полисахаридов, распространение биопленки.
210.
• Сидячие биопленки формируются при многих заболеванияхчеловека – при отите, кистозном фиброзе (муковисцидоз), на
поверхности искусственных сердечных клапанов и других
имплантов.
• Микроорганизмы в составе биопленки взаимодействуют между
собой, выделяя аутоиндукторы. Регуляция экспрессии генов с
участием аутоиндукторов, называется quorum sensing.
• Полагает, что такая система существует и в микрофлоре
кишечника, что играет важную роль в предупреждении
колонизации кишечника патогенными бактериями.
211.
Значение бактерий в природеи жизни человека
• Болезнетворные бактерии
• Бактерии в составе микрофлоры кишечника
• Бактерии в пищевой промышленности, осуществляющие
молочнокислое брожение
• Бактерии могут вызывать порчу продуктов (маслянокислое
брожение)
• Бактерии используются для очистки сточных вод.
• Почвенные бактерии (н-р,азотфиксирующие)
• Участие бактерий в круговоре веществ (хемосинтезирующие
бактерии)
212.
• Использование бактерий в биотехнологии (генная инженерия,микробиологическая промышленность)
• CRISPR-Cas –технологии (см.далее)