























")


















 Биология
БиологияПохожие презентации:










Нуклеиновые кислоты
1. Нуклеиновые кислоты
1.2.
3.
4.
5.
6.
История открытия НК
Функции НК
Компоненты НК
Нуклеозиды и нуклеотиды
Строение полинуклеотидной цепи
Уровни организации молекулы ДНК
2.
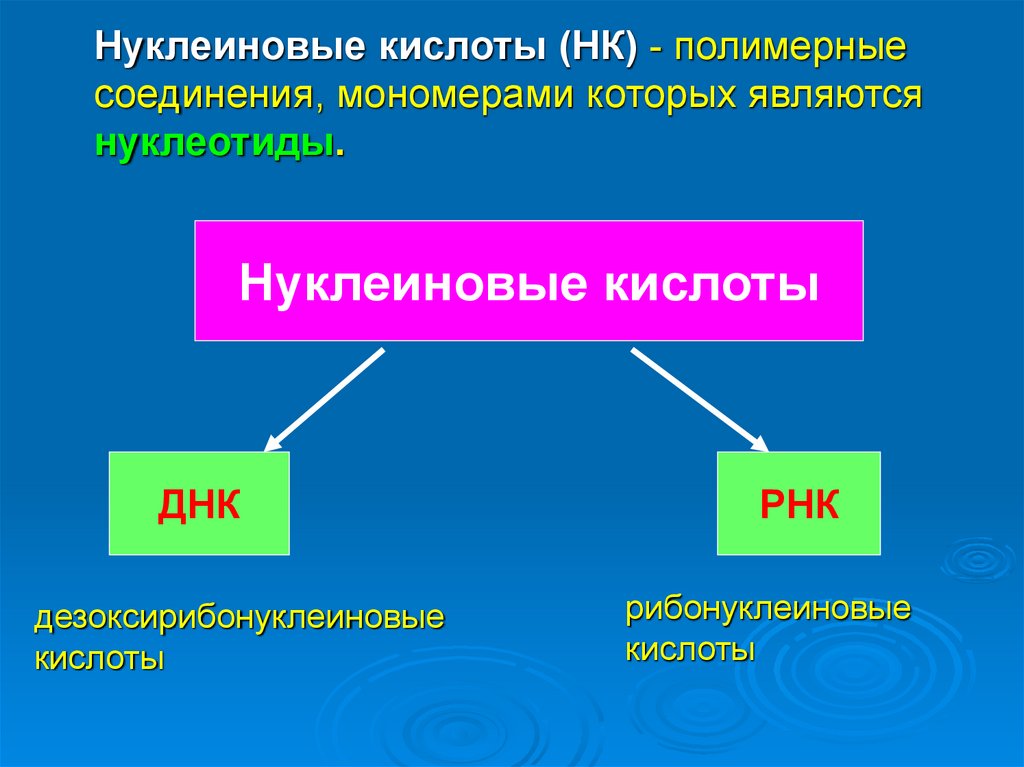
Нуклеиновые кислоты (НК) - полимерныесоединения, мономерами которых являются
нуклеотиды.
Нуклеиновые кислоты
ДНК
дезоксирибонуклеиновые
кислоты
РНК
рибонуклеиновые
кислоты
3. История открытия НК
1868г – Ф.Мишер выделил из ядер лейкоцитовчеловека соединения нового типа, которые он
назвал нуклеинами (от лат. nucleus - ядро).
1889г – Ф.Мишер определил, что нуклеин обладает
кислотными свойствами
1928 г – Ф.Гриффитс открыл явление генетической
трансформации свойство патогенности можно
передавать используя ДНК)
40-50-е гг ХХ в – получены доказательства того, что
именно ДНК, а не белки, является носителем
генетической информации
- 1950г- определен нуклеотидный состав ДНК из
разных объектов ( Чаргафф)
- 1953 г – Р.Франклин и М.Вилкинс (ДНК –это спираль
с шагом 3,4нм, на основе рентгеноструктурного
анализа)
- 1953г- Д.Уотсон и Ф.Крик – двойная спираль ДНК
4. Функции НК
1. хранение информации2. перенос информации ( передача)
3. реализация информации ( синтез
белка)
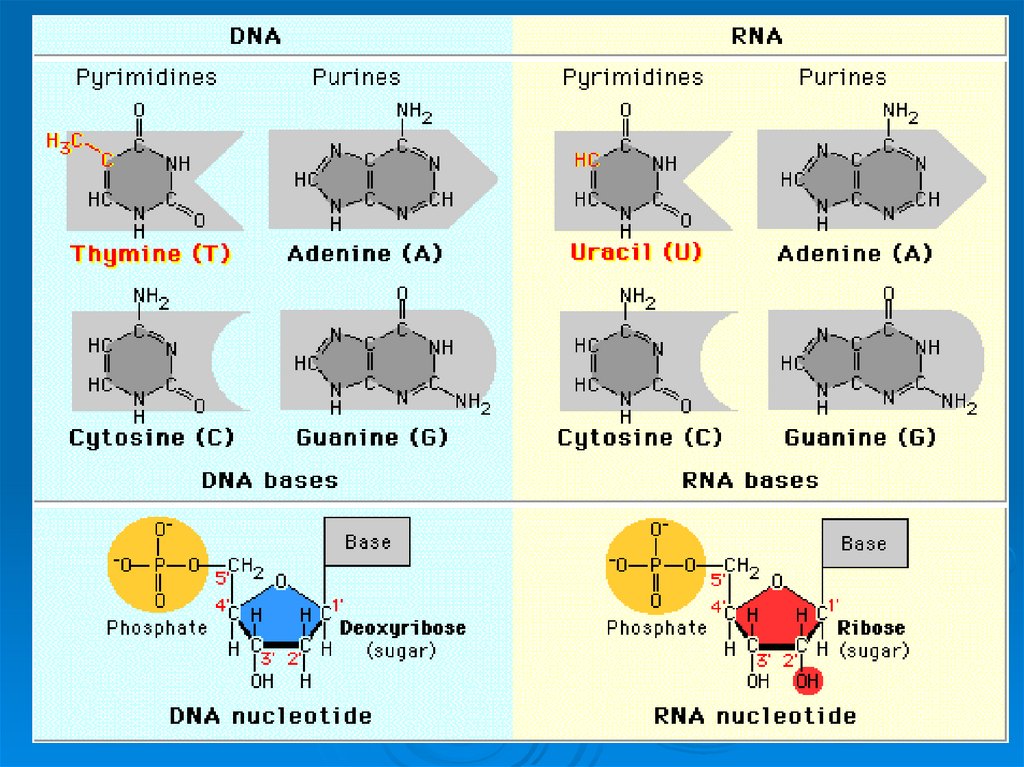
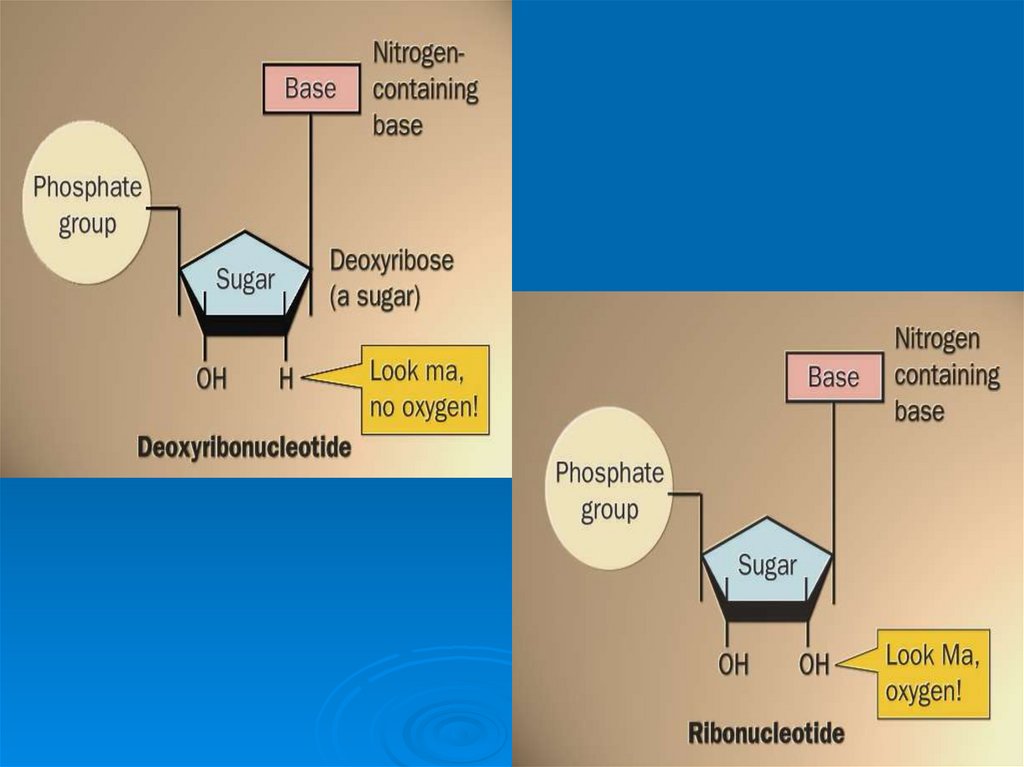
5. Компоненты нуклеотидов:
Углевод (пентоза)Азотистое основание
Остаток фосфорной кислоты
6. Сахара, входящие в состав нуклеотидов:
рибозадезоксирибоза
7. Пуриновые азотистые основания
Пуриновые азотистыеоснования являются
производными
ароматического
гетероциклического
соединения - пурина
8. Пиримидиновые азотистые основания
Пиримидиновыеазотистые основания
являются производными
ароматического
гетероциклического
соединения пиримидина
9. Минорные азотистые основания
Минорные пиримидиновые основания:Минорные пуриновые основания:
гипоксантин
ксантин
10. Остаток фосфорной кислоты
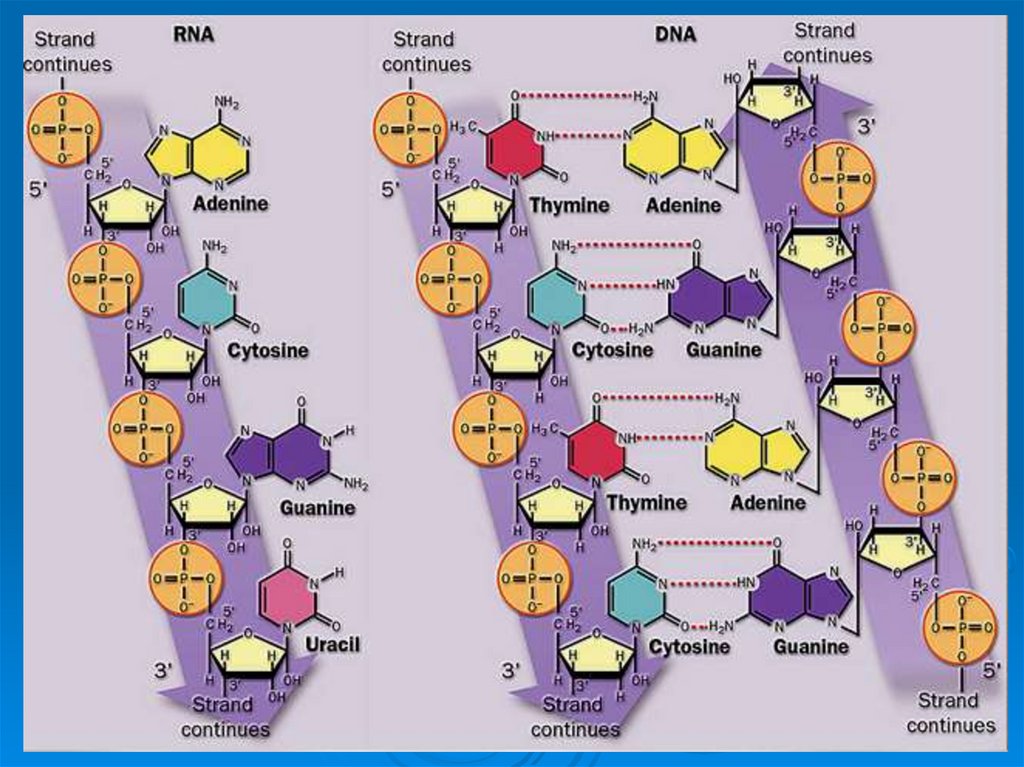
11.
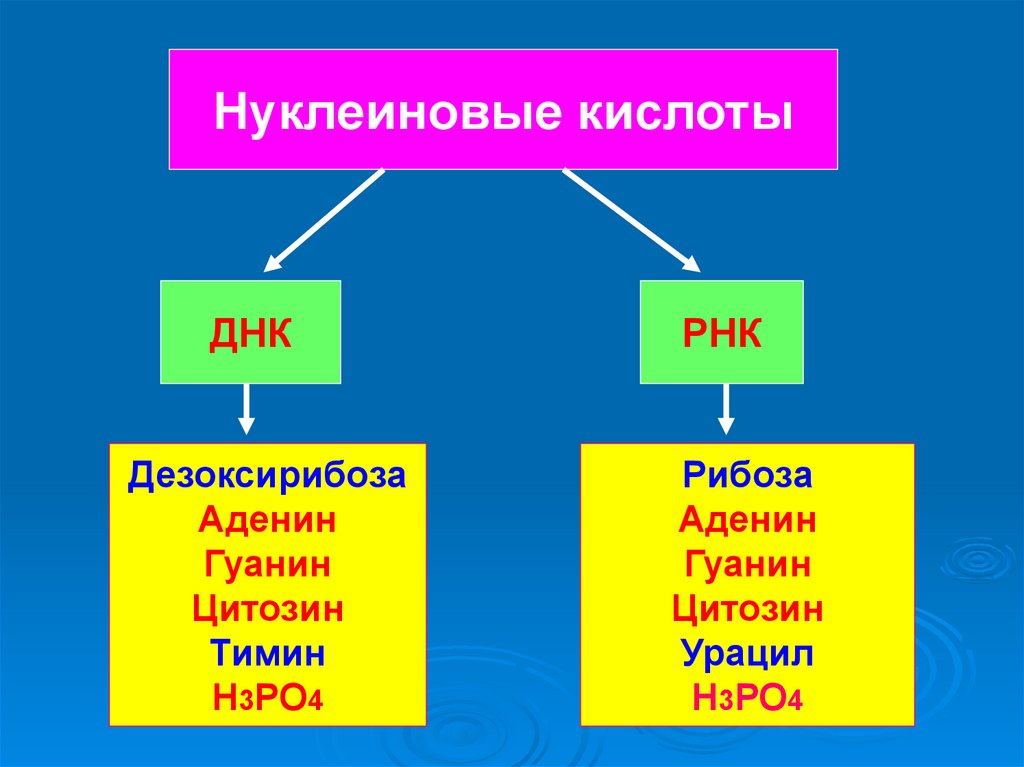
Нуклеиновые кислотыДНК
РНК
Дезоксирибоза
Аденин
Гуанин
Цитозин
Тимин
Н3РО4
Рибоза
Аденин
Гуанин
Цитозин
Урацил
Н3РО4
12.
13.
14.
15. Нуклеозиды и нуклеотиды
Комбинацияазотистого основания
с углеводным
компонентом
приводит к
образованию
нуклеозида
Фосфорные эфиры
нуклеозидов
называются
нуклеотидами
16. Нуклеозид
В НК пиримидиновые азотистыеоснования через 1-й атом, а пуриновые –
через 9-й образуют N – гликозидную связь
с пентозой рибозой или дезоксирибозой.
17. Образование гликозидной связи
18. Нуклеотид
Азотистое основание, пентоза ифосфорная кислота, соединяясь друг с
другом, образуют молекулу нуклеотида
19.
20. Образование нуклеотида
21.
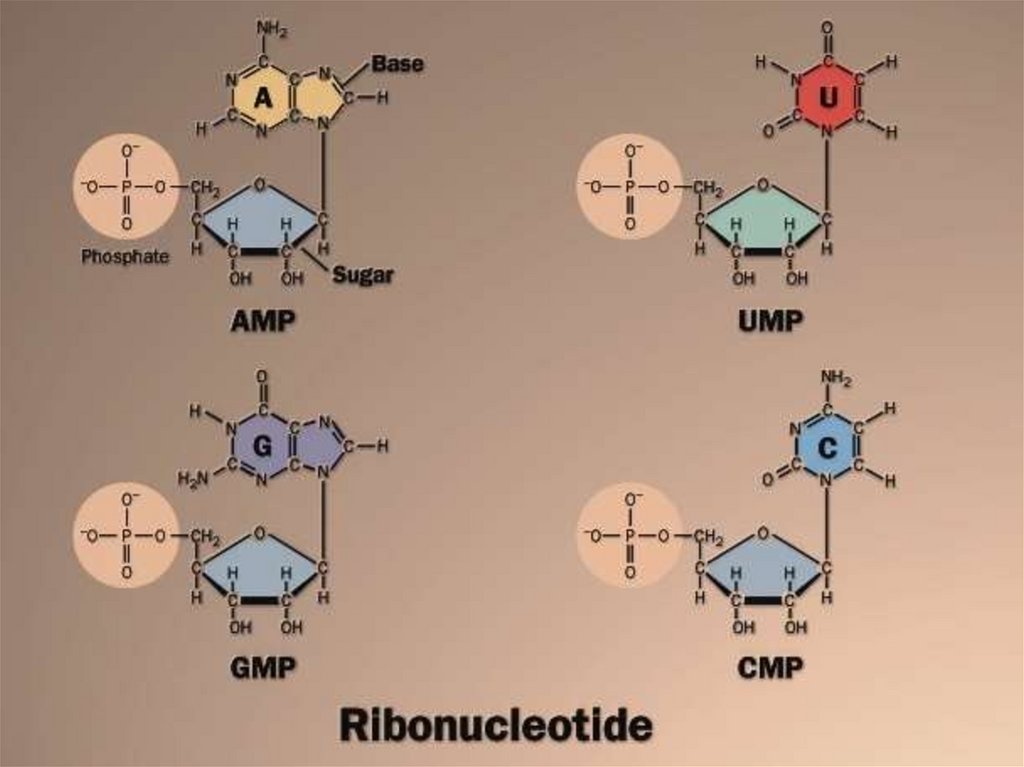
22. Номенклатура нуклеозидов и нуклеотидов
НуклеотидыАзотистое Нуклеозиды
основание
Аденин
Аденозин
Гуанин
Гуанозин
Цитозин
Цитидин
Урацил
Уридин
Тимин
Тимидин
Полное название
Сокращенное
название
Адениловая кислота,
аденозинмонофосфат
Гуаниловая кислота,
гуанозинмонофосфат
Цитидиловая кислота,
цитидинмонофосфат
Уридиловая кислота,
уридинмонофосфат
Тимидиловая кислота,
тимидинмонофосфат
АМФ
ГМФ
ЦМФ
УМФ
ТМФ
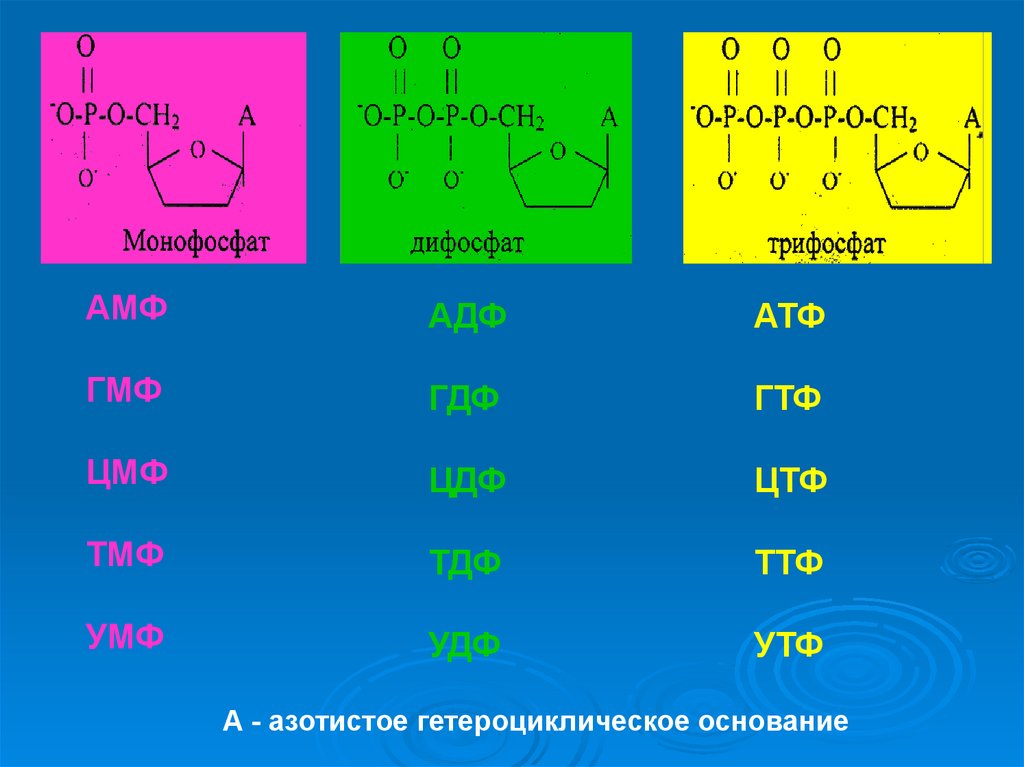
23.
АМФАДФ
АТФ
ГМФ
ГДФ
ГТФ
ЦМФ
ЦДФ
ЦТФ
ТМФ
ТДФ
ТТФ
УМФ
УДФ
УТФ
А - азотистое гетероциклическое основание
24. Функции нуклеотидов
Они являются активированными предшественникамиДНК и РНК.
Нуклеотиды являются переносчиками молекул.
Например, глюкоза при синтезе гликогена
переносится с помошью УТФ; холин при синтезе
лецитинов переносится с помощью ЦТФ.
Производные нуклеотидов – активированные
промежуточные продукты многих биосинтетических
реакций. Например, УДФ-глюкоза и ЦДФдиацилглицеролы - предшественники гликогена и
фосфоацилглицеролов (фосфоглицеридов)
соответственно.
АТФ – универсальная энергетическая валюта в
биологических системах
Адениннуклеотиды – компоненты трех основных
коферментов – НАД+, ФАД и кофермент А.
Нуклеотиды – регуляторы метаболизма ( ц АМФуниверсальный посредник действия некоторых
классов гормонов)
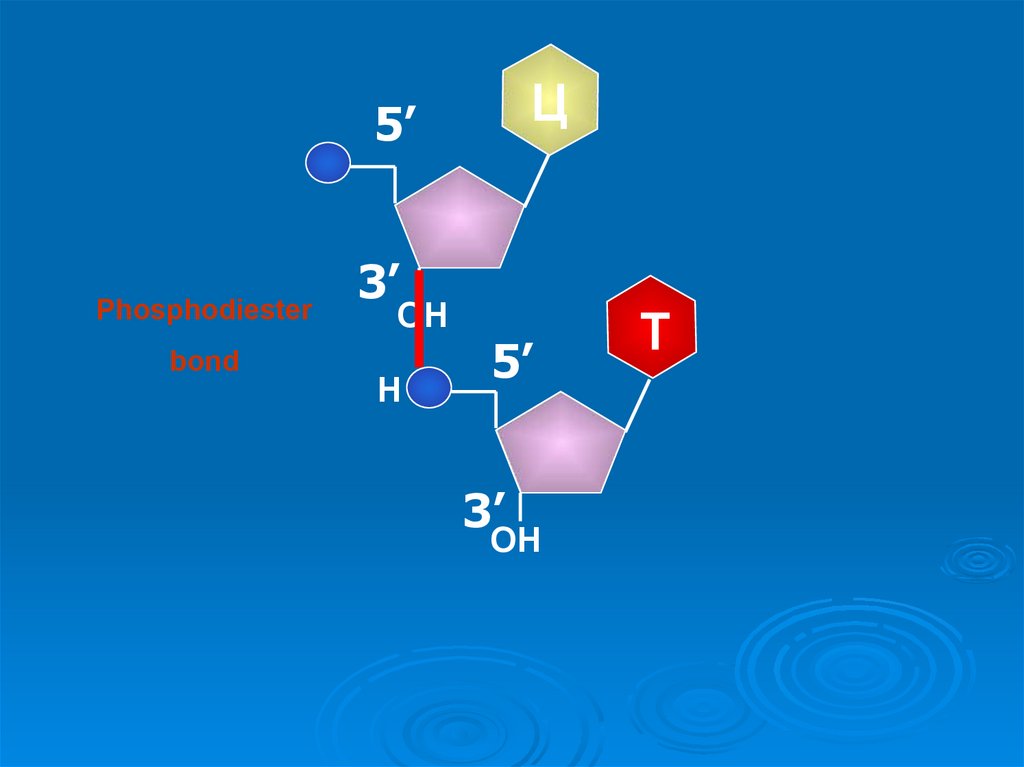
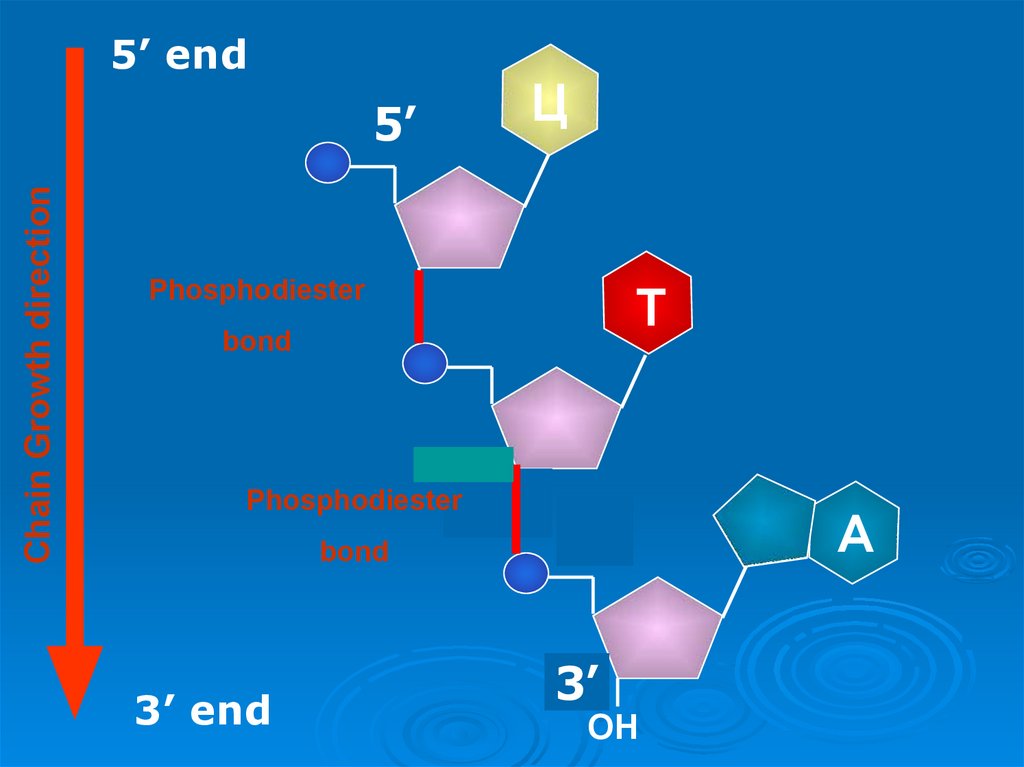
25. Строение полинуклеотидной цепи
- Отдельные нуклеотидысоединяются между
собой в полимерную
цепь за счет
образования 3’, 5’ фосфодиэфирной связи.
Межнуклеотидная связь
осуществляется за счет
фосфатной группы, а
также 3'- и 5'гидроксильных групп
пентоз нуклеотидов.
- Полинуклеотидная цепь
имеет определенное
направление: 5’ → 3’. На
одном ее конце остается
свободной 5’-Р-группа
(начало цепи), на другом
- 3’-ОН-группа (конец
цепи)
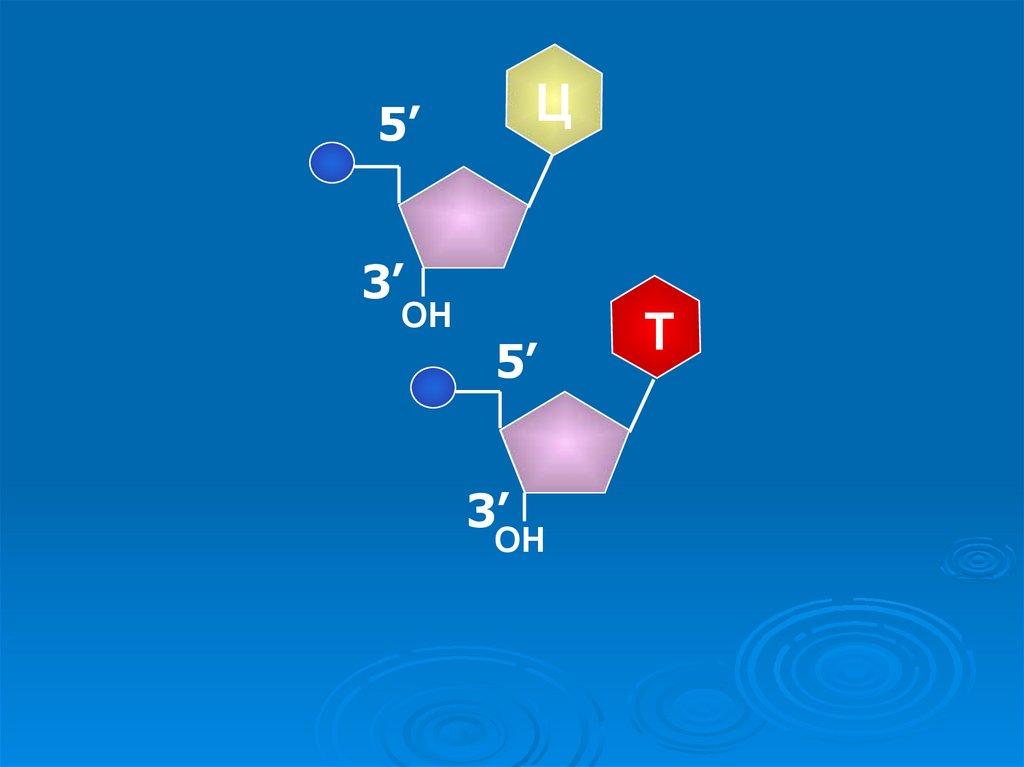
26.
Ц5’
3’
ОН
5’
3’
ОН
Т
27.
Ц5’
Phosphodiester
bond
3’
ОН
Н
5’
3’
ОН
Т
28.
5’ endЦ
Chain Growth direction
5’
Phosphodiester
Т
bond
3’
Phosphodiester
bond
3’ end
ОН
5’
3’
ОН
A
29. Правила Чаргаффа
Пур = Пир или Пур / Пир = 1.2. А = Т или А/Т = 1.
3. Г = Ц или Г/Ц = 1.
4. Г + Т = А + Ц или Г + Т / А + Ц = 1.
5. В ДНК различных источников
соотношение нуклеотидов:
либо (А+Т)>(Г+Ц) – АТ-тип ДНК,
либо (Г+Ц)>(А+Т)- ГЦ-тип ДНК
1.
Эрвин Чаргафф
1950 г –правила
Чаргаффа
Нуклеотидный состав ДНК специфичен для каждого вида.
Установлено, что у большинства животных преобладает
АТ-тип ДНК.
У бактерий наблюдается разброс нуклеотидного состава от
сильно выраженного ГЦ-типа до АТ-типа.
В геноме высших эукариот ДНК содержится на 3-4 порядка
больше, чем у бактерий.
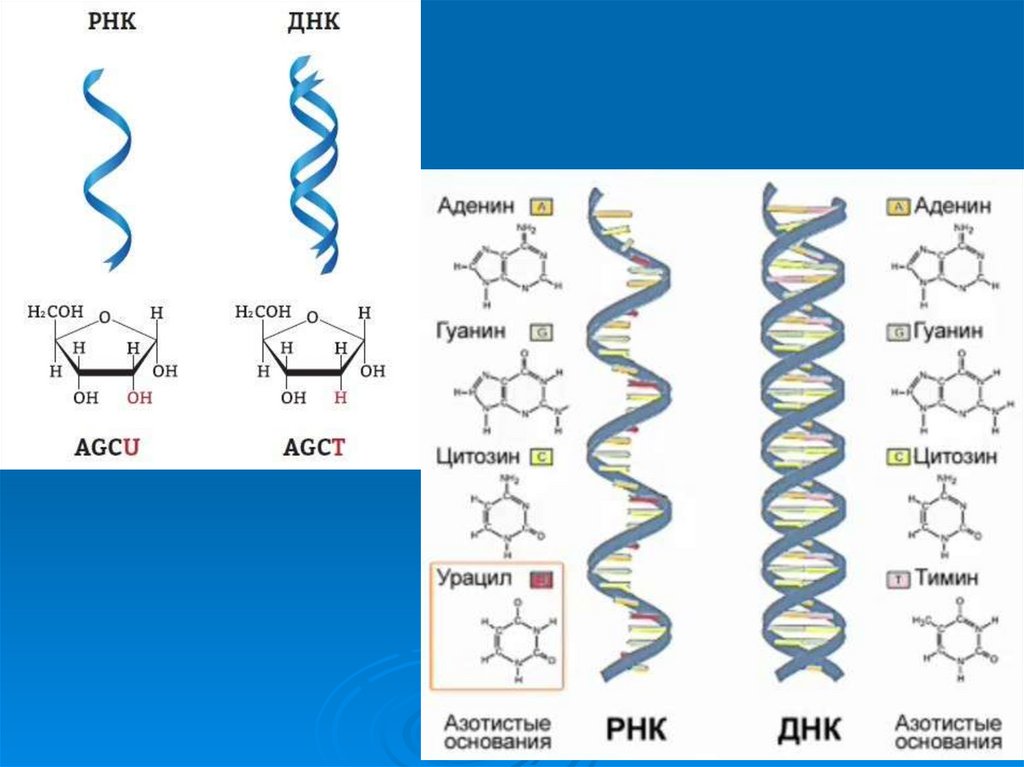
30. Вторичная структура ДНК
1953 год Джэймс Уотсон и Фрэнсис КрикРентгенограммы Розалинды Франклин и Мориса Уилкинса
31.
32. Принцип комплементарности спаривания азотистых оснований
Ц- ГА - Т
Азотистые основания, принадлежащие двум цепочкам, избирательно
соединяются водородными связями, образуя специфические пары:
А-Т
Г-Ц
А и Т соединяются двумя водородными связями в положении 1:3 и 6:4,
Г и Ц, тремя водородными связями в 1:3, 2:2 и 6-4.
Эти азотистые основания называются комплементарными.
Комплементарность – дополнительность цепей ДНК друг другу.
33. Двойная спираль ДНК
34. Характеристики двойной спирали
В модели двойнойспирали Уотсона и
Крика две
полинуклеотидные цепи
обвивают друг друга,
образуя правую спираль
(хеликс)
цепи антипараллельны
углеводно-фосфатные
группы располагаются
снаружи,
пуриновые и
пиримидиновые
основания – внутри.
35. Характеристики двойной спирали:
1. Двойная спираль - это 2 полинуклеотидные цепи, обвивающиедруг друга.
2. Эти полинуклеотидные цепи ДНК - антипараллельны.
3. Азотистые основания располагаются внутри спирали,
а остатки сахара и фосфорной кислоты снаружи.
4. Азотистые основания соединяются друг с другом
по принципу
комплементарности
5. Последовательность нуклеотидов одной цепи автоматически
определяет последовательность нуклеотидов комплементарной
цепи.
6. Диаметр спирали 2 нм.
7. На один виток спирали приходится 10 пар оснований.
8. Шаг спирали ( длина одного витка) = 3,4 нм
9. Расстояние между плоскостями оснований =0,34нм.
10.Между стопками азотистых оснований возникают стэкингвзаимодействия.
36. Конформационные характеристики ДНК (В-форма)
Стабильность двойнойспирали определяется
взаимодействием
расположенных друг
над другом азотистых
оснований.
Диаметр – 2,0 нм
Длина витка спирали –
3,4 нм
На один виток спирали
– 10 нуклеотидов
Межнуклеотидное
расстояние – 0,34 нм
Влажность ДНК в
форме В выше 75%.
37. Форма молекулы ДНК
При изменении внешних условийдвойная спираль может принимать
конформации, отличные от уотсонкриковской В-формы. Так существует:
А-форма,
С-форма,
Z-форма.
38. А-форма ДНК
В-форма превращается вА- форму, когда
влажность препарата
ДНК становится менее
75%.
В А-форме пары
оснований наклонены к
оси спирали под углом
около 20 градусов, в
результате чего шаг
спирали уменьшается с
3,4 до 2,8 нм.
В А-форме
насчитывается 11 пар
оснований на виток, что
приводит к укорачиванию
цепи приблизительно на
25%.
Предполагают, что в Аформе ДНК
функционирует в
процессе транскрипции.
39. Z-форма
В участках, содержащихчередующуюся
последовательность Г и
Ц, ДНК приобретает Zформу.
Это левая спираль, на
один виток которой
приходится 12 пар
оснований.
Сахарофосфатный остов
имеет форму не спирали,
а зигзагообразный вид
(Z).
40. С-форма
С-форма очень сходна с В-формой,имеет 9,3 пар оснований на виток,
основания наклонены под углом 5
градусов.
Полагают, что форму, близкую к С
имеет ДНК в составе надмолекулярных
структур хроматина и у ряда вирусов.
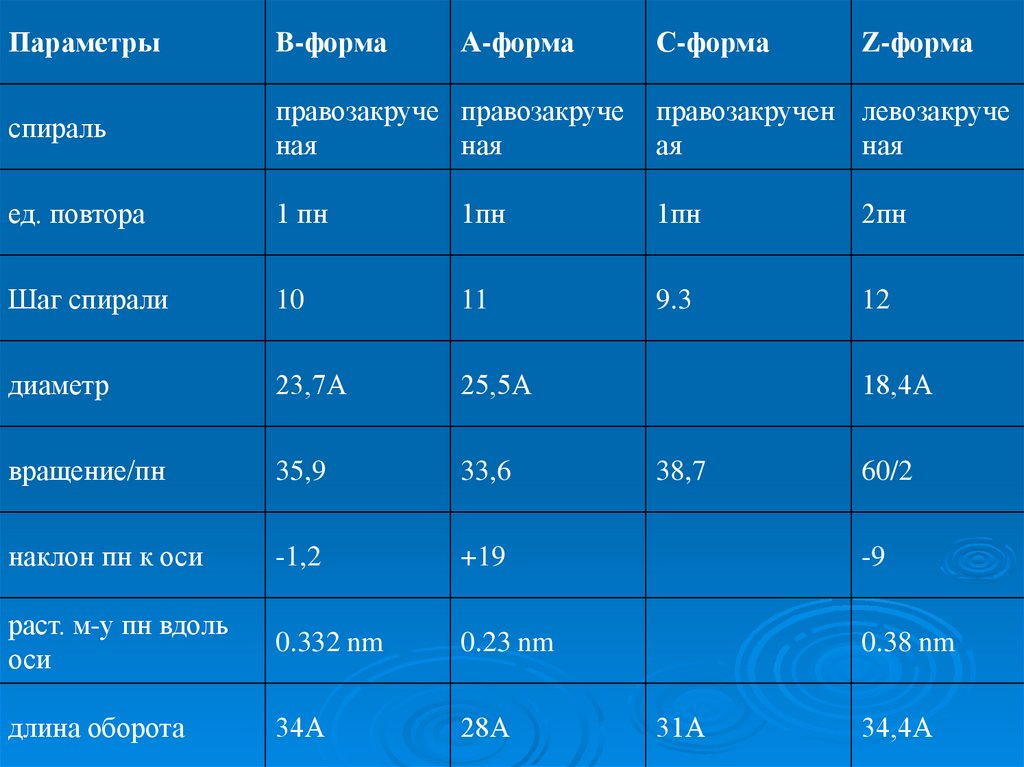
41.
ПараметрыB-форма
А-форма
С-форма
спираль
правозакруче правозакруче
ная
ная
правозакручен левозакруче
ая
ная
ед. повтора
1 пн
1пн
1пн
2пн
Шаг спирали
10
11
9.3
12
диаметр
23,7А
25,5А
вращение/пн
35,9
33,6
наклон пн к оси
-1,2
+19
-9
раст. м-у пн вдоль
оси
0.332 nm
0.23 nm
0.38 nm
длина оборота
34А
28А
Z-форма
18,4А
38,7
31А
60/2
34,4А
42. Размеры ДНК
Всредней эукариотической клетке
общая протяженность геномной ДНК
составляет около 2 м, диаметр ее ядра
всего ~10-20 мкм.
У E.coli в клетке диаметром несколько
микрометров находится молекула ДНК
длиной около 1 мм.
43.
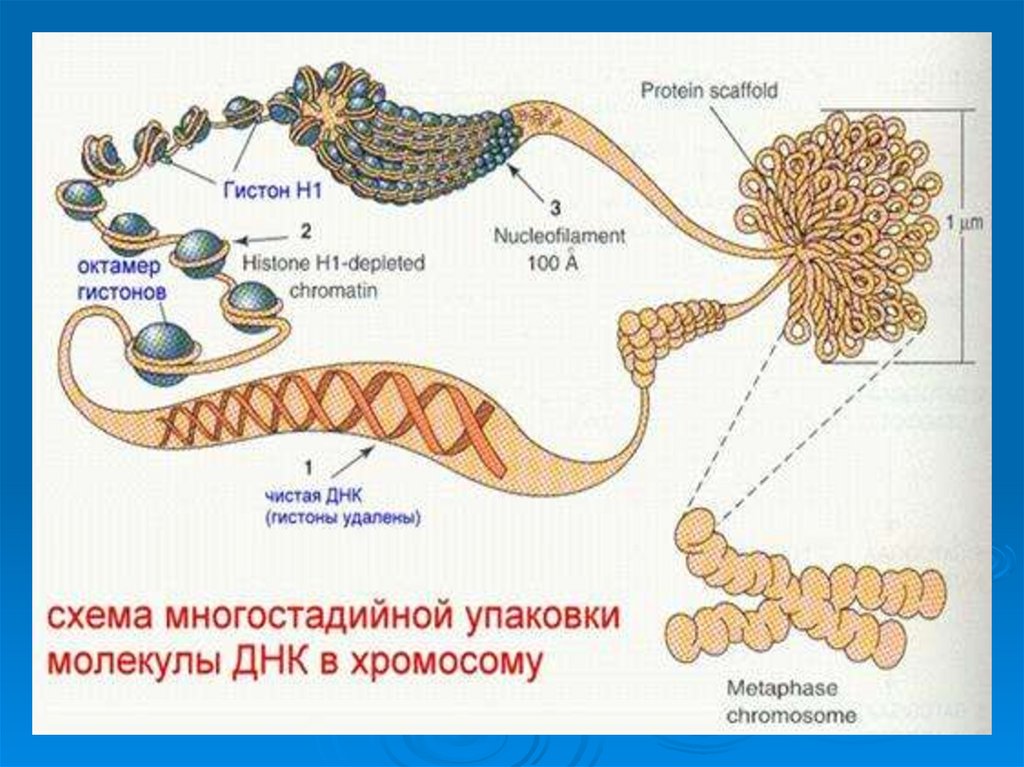
44. Как ДНК таких размеров может уместиться в микроскопически маленьких ядрах клеток?
Третичная структура ДНК эукариотических клеток- комплекс ДНК с белками.
ДНК эукариот почти вся находится в хромосомах ядер, лишь
небольшое количество ее содержится в митохондриях, а у растений
и в пластидах.
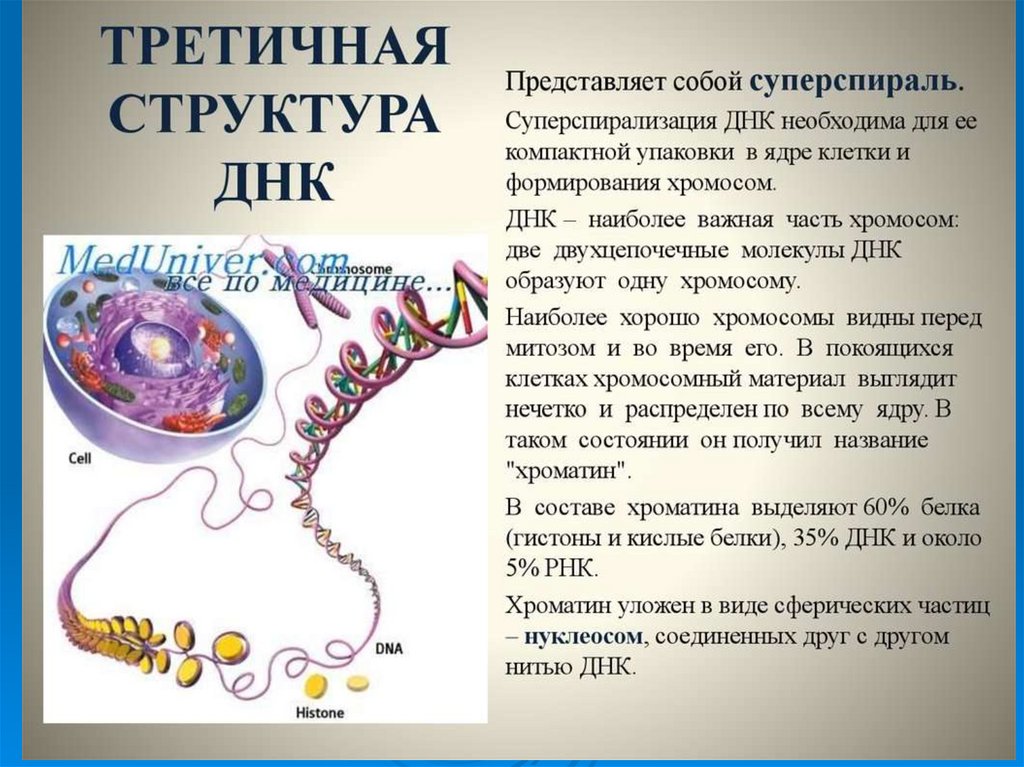
Суммарный материал хромосом – хроматин – содержит:
ДНК;
гистоны;
негистоновые белки;
небольшое количество РНК.
45.
46.
47.
48.
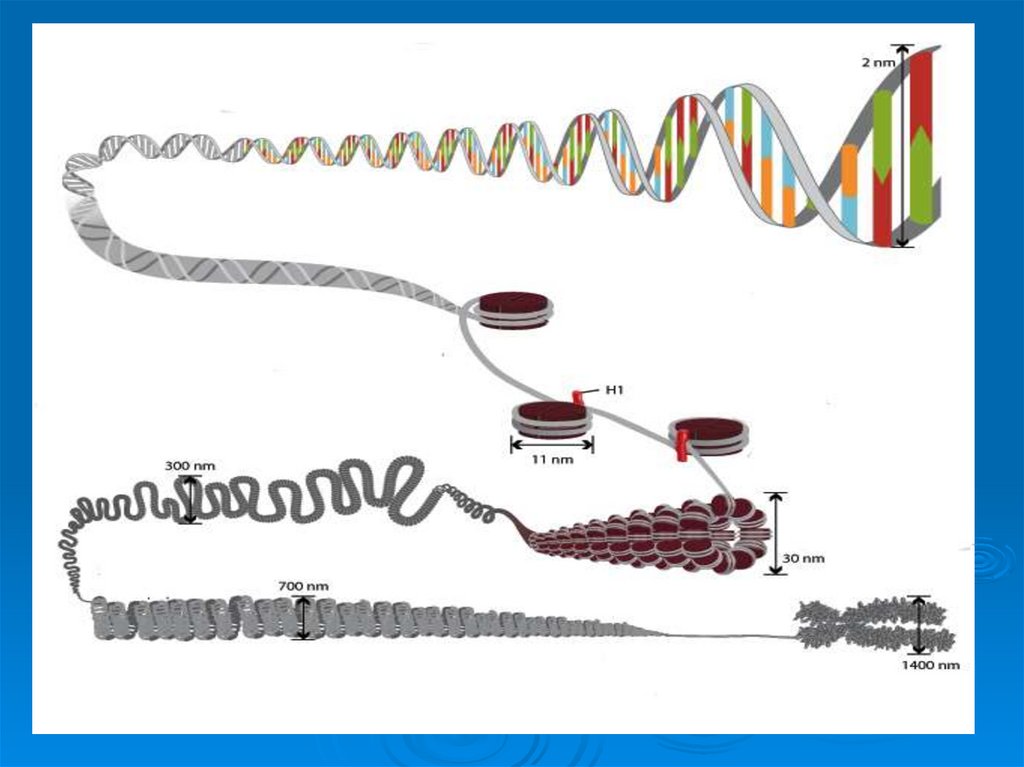
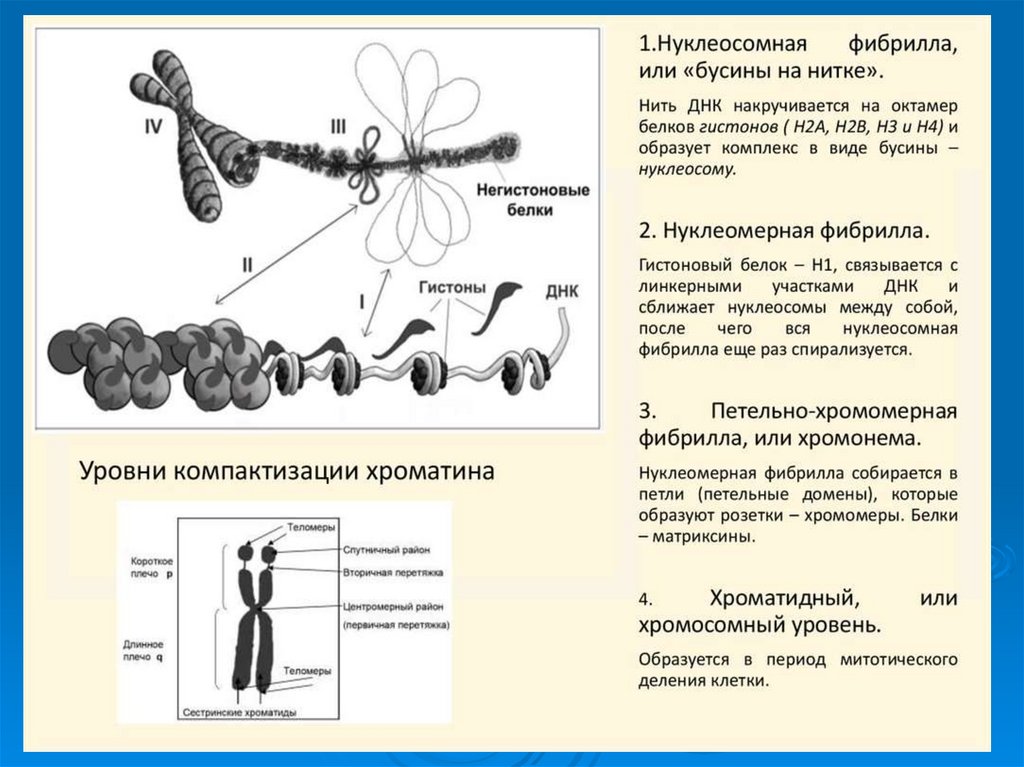
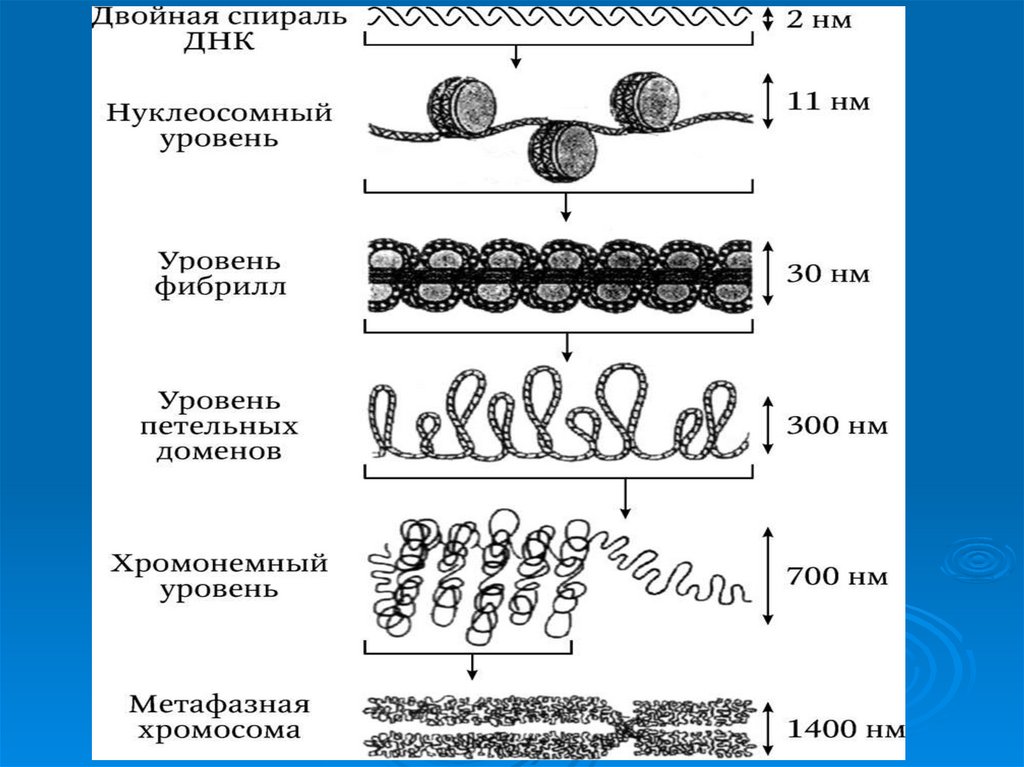
49. Третичная структура ДНК
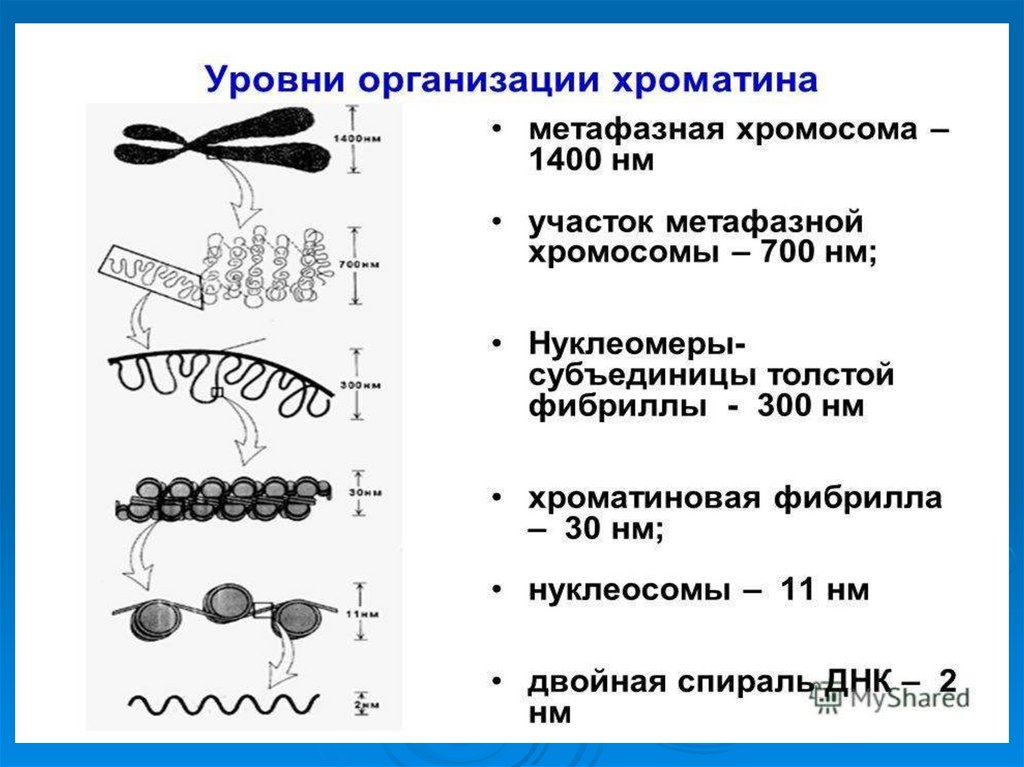
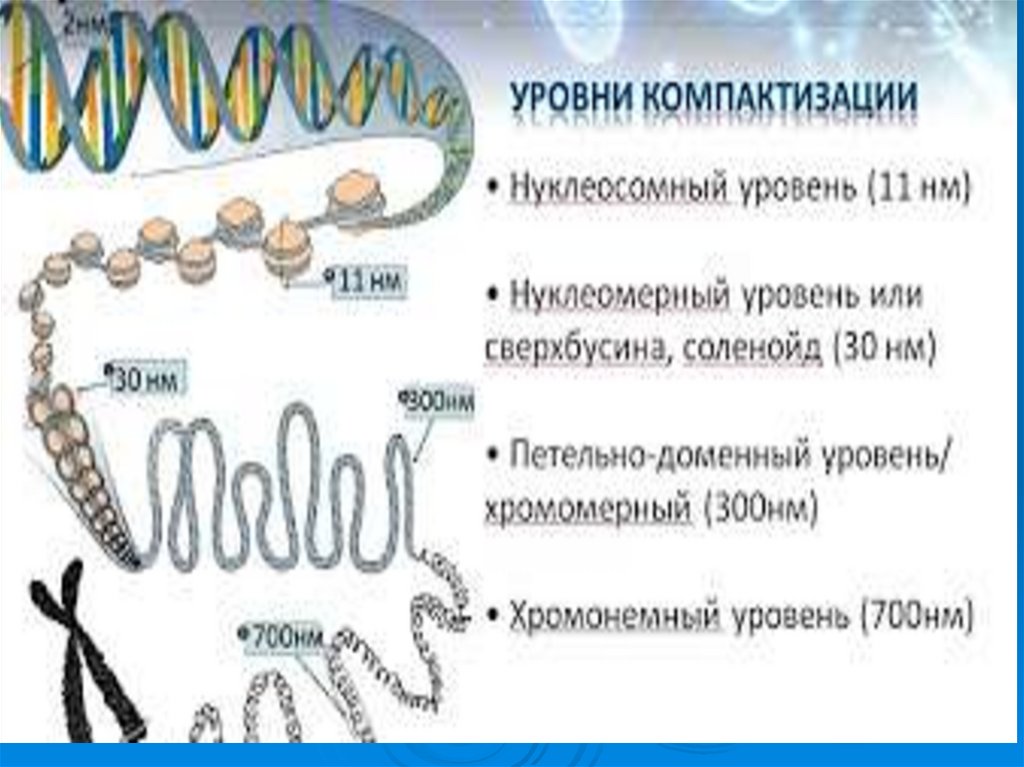
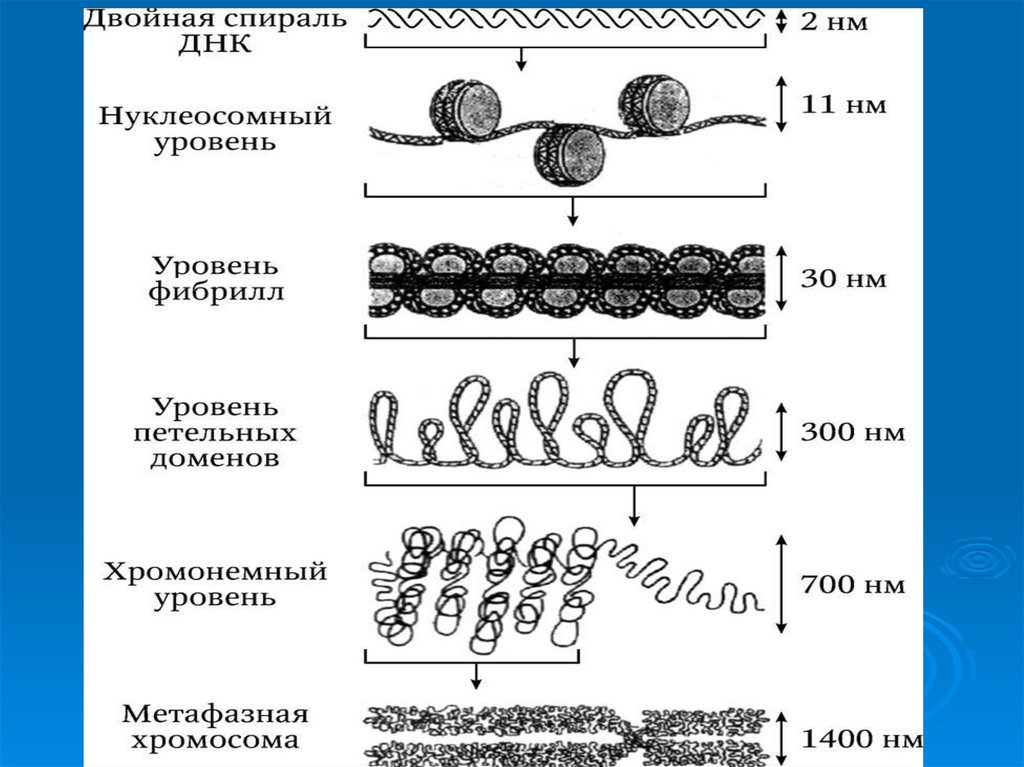
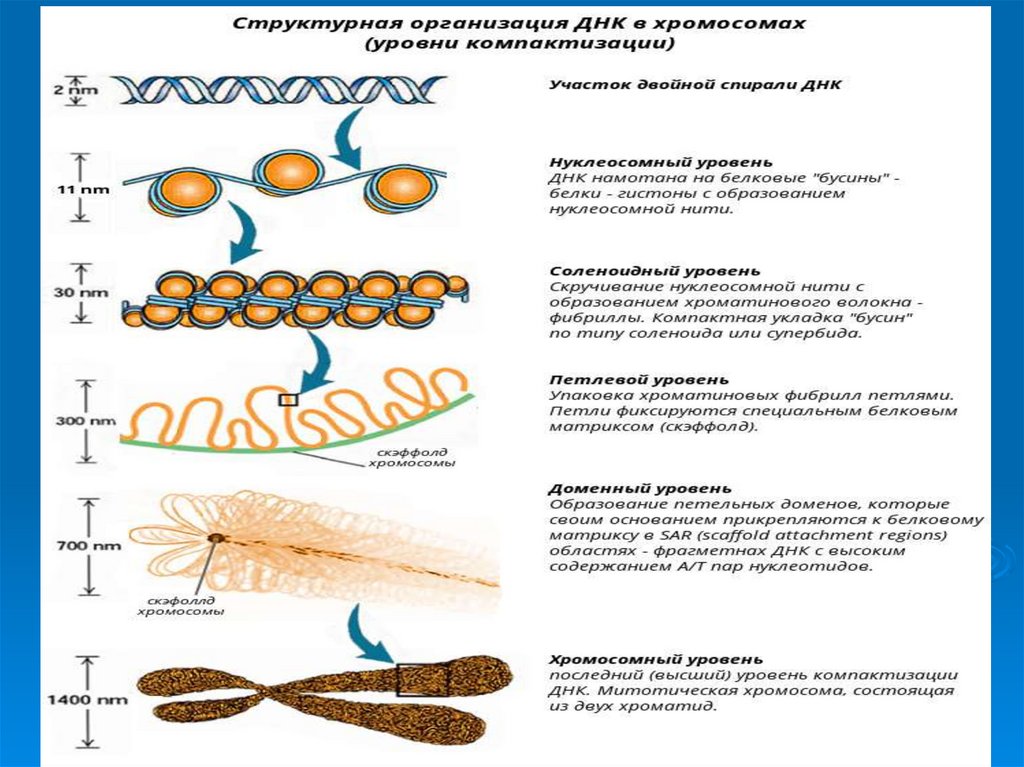
УпаковкаДНК в ядре эукариотической клетки
осуществляется в несколько этапов (уровни
третичной структуры):
1.
Нуклеосомный: нить ДНК укладывается в
нуклеосомы, при этом ее длина уменьшается в
шесть-семь раз.
2.
Образование
фибрилл:
нуклеосомная
нить
складывается в так называемую 30 нм фибриллу
(соленоид), что обеспечивает дополнительную
компактизацию в 40 раз.
3. Образование петель: фибрилла организуется в
большие (50 и более тысяч пар нуклеотидов) петли,
концы которых закрепляются на белковом скелете
ядра (его часто называют ядерным матриксом). На
этом этапе линейные размеры ДНК сокращаются в
700 раз
50. Уровни упаковки ДНК в ядре эукариотической клетки.
51.
52. Гистоны
До 50% хроматина составляют простые белкигистоны.
Молекула
гистона
состоит
из
одной
полипептидной цепи, которая в своей средней
части спирализована и скручена в глобулу
диаметром около 2,5 нм; от глобулы отходят 2
неспирализованых конца молекулы.
Первичная структура гистонов имеет ряд
особенностей. Так, в гистонах Н2А, Н2В, Н3, Н4 и
N-концевой области большинство аминокислотных
остатков положительно заряжено, а С-концевая
лишена заряда
53. Гистоны
Классификациягистонов основана на
содержании в них лиз
и арг.
54. Взаимодействие гистонов с ДНК
Гистонывзаимодействуют с ДНК
в основном через
ионные связи,
образующиеся между
отрицательно
заряженными
лизиновыми и
аргининовыми
остатками гистонов.
55. Гистоны в составе нуклеосом
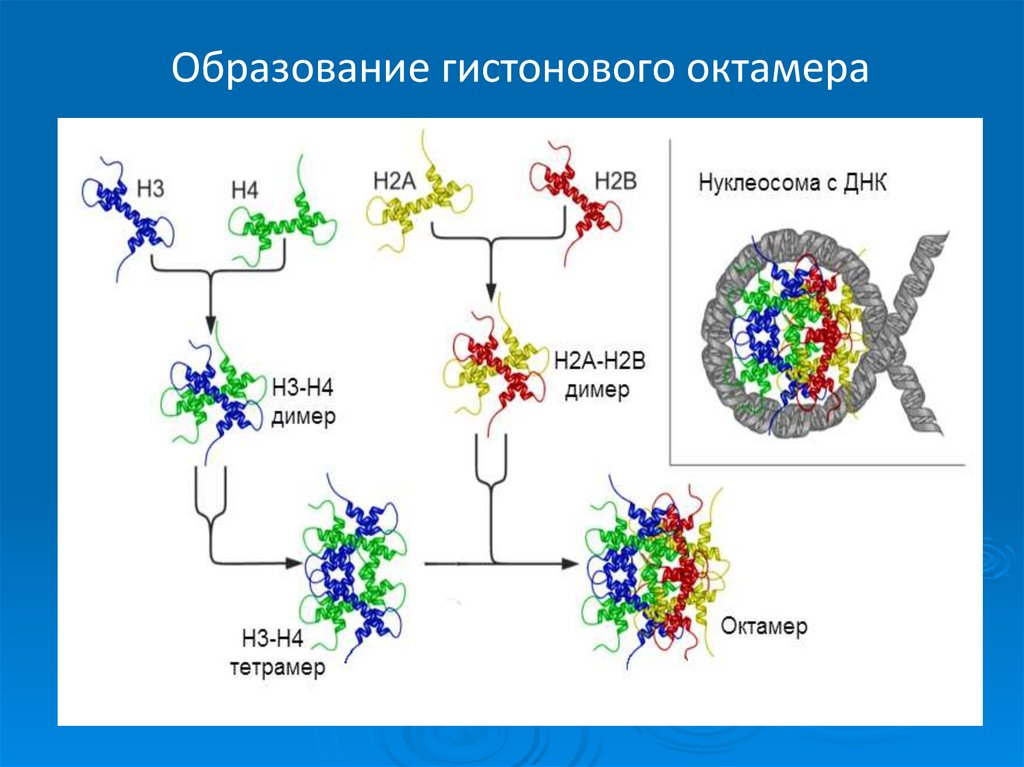
56. Строение нуклеосомы
Нуклеосома содержит ДНКдлиной
160-240
пар
нуклеотидов, одну молекулу
гистона Н1 и гистоновый
октамер. Последний состоит
из 8 молекул – по 2
молекулы из гистонов Н2А,
Н2В, Н3 и Н4.
Из
нуклеосом
можно
выделить
нуклеосомное
ядро или нуклеосомный
кор. Эта дискретная частица
содержит
гистоновый
октамер и участок ДНК
длиной
145-150
нуклеотидных пар.
57.
Образование гистонового октамера58. Строение нуклеосомы
Между коровыми частицамирасположены участки ДНК –
линкеры,
их
длина
варьирует в зависимости от
типа клеток. В результате
этого варьирует и длина
фрагмента ДНК, входящего в
состав нуклеосом.
Межкоровые
(линкерные)
участки ДНК или свободны,
или связаны с гистоном Н1 и
негистоновыми белками.
Гистон
Н1
способствует
компактной
упаковке
хроматина.
59.
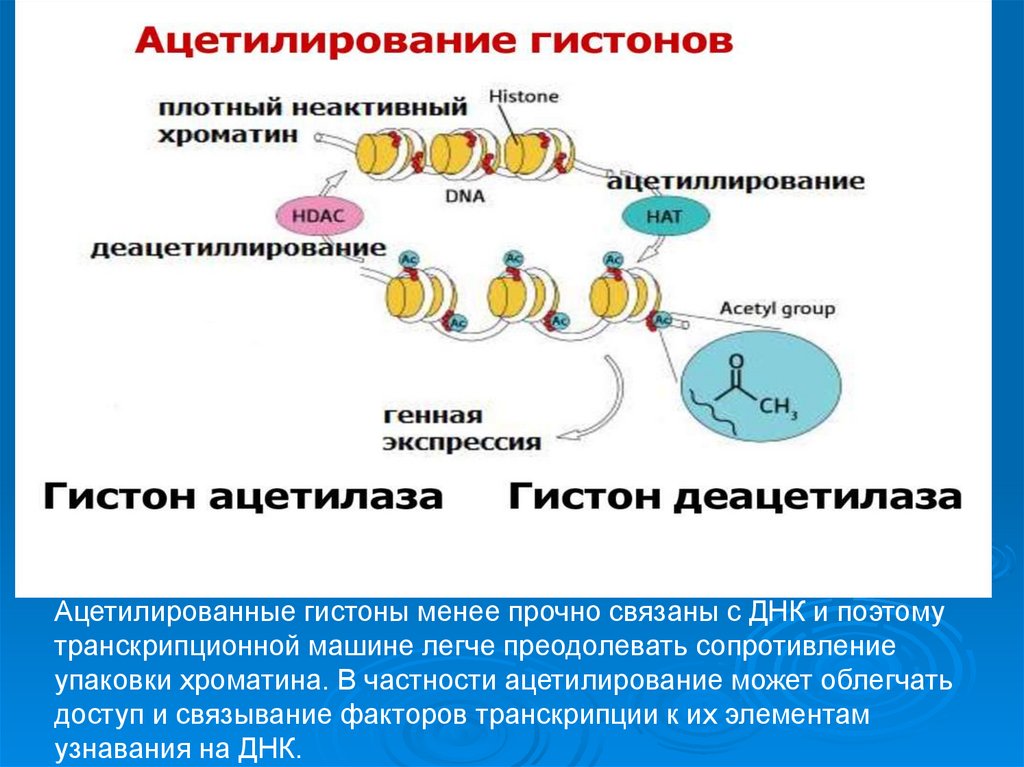
60.
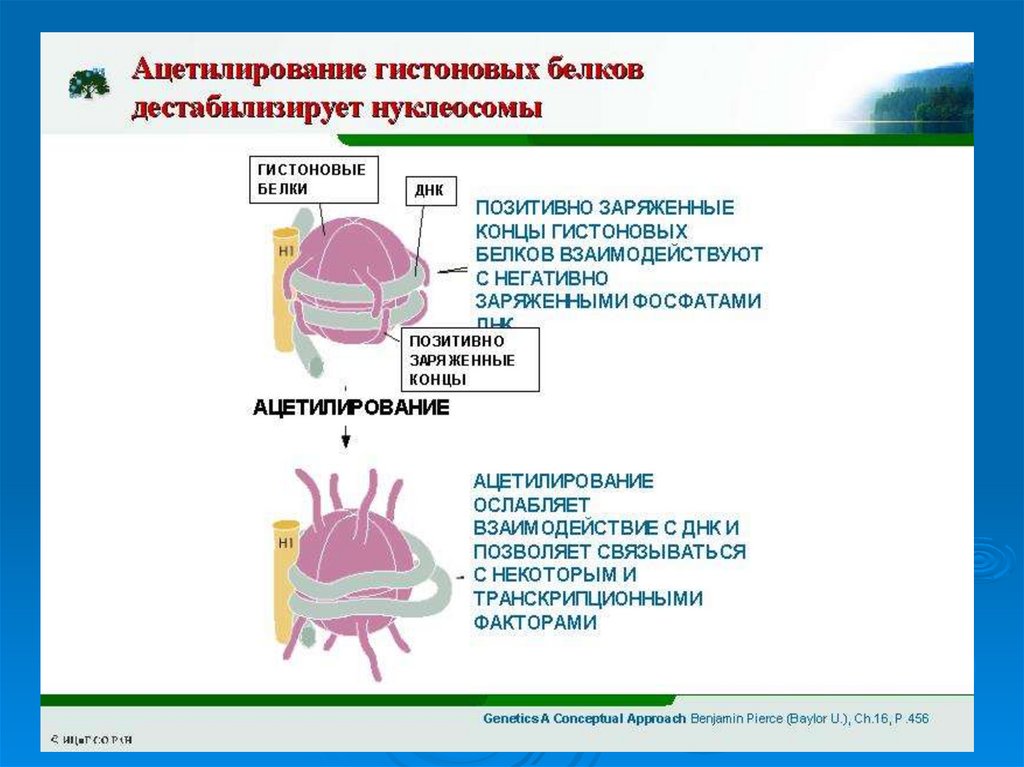
Ацетилированные гистоны менее прочно связаны с ДНК и поэтомутранскрипционной машине легче преодолевать сопротивление
упаковки хроматина. В частности ацетилирование может облегчать
доступ и связывание факторов транскрипции к их элементам
узнавания на ДНК.
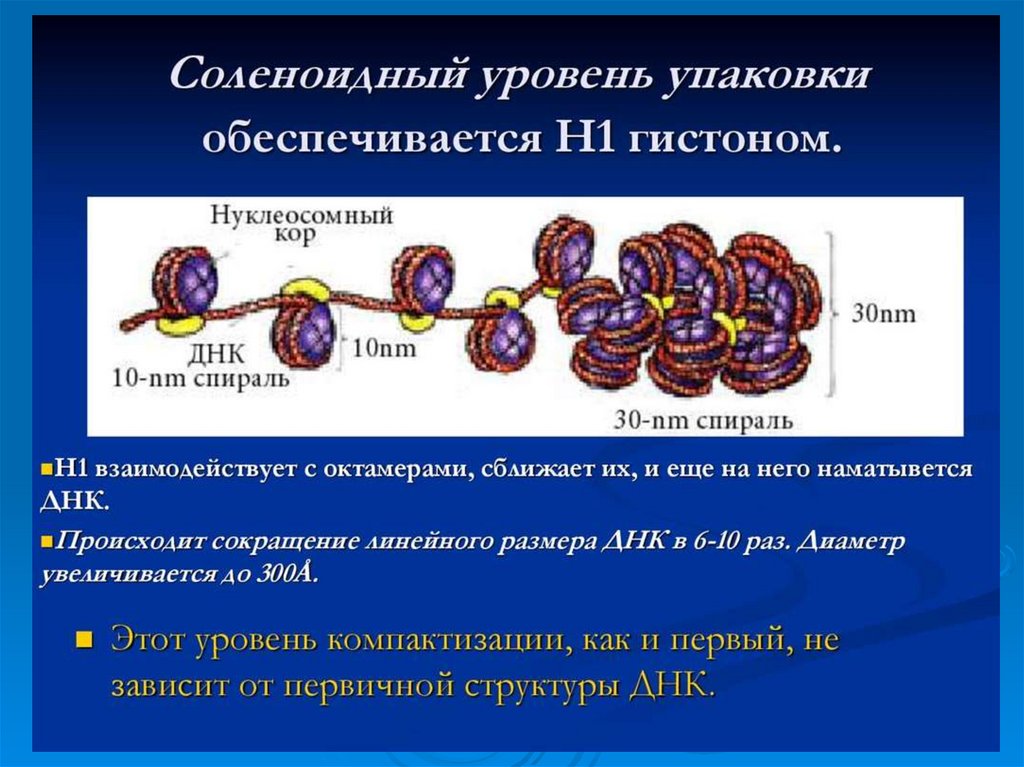
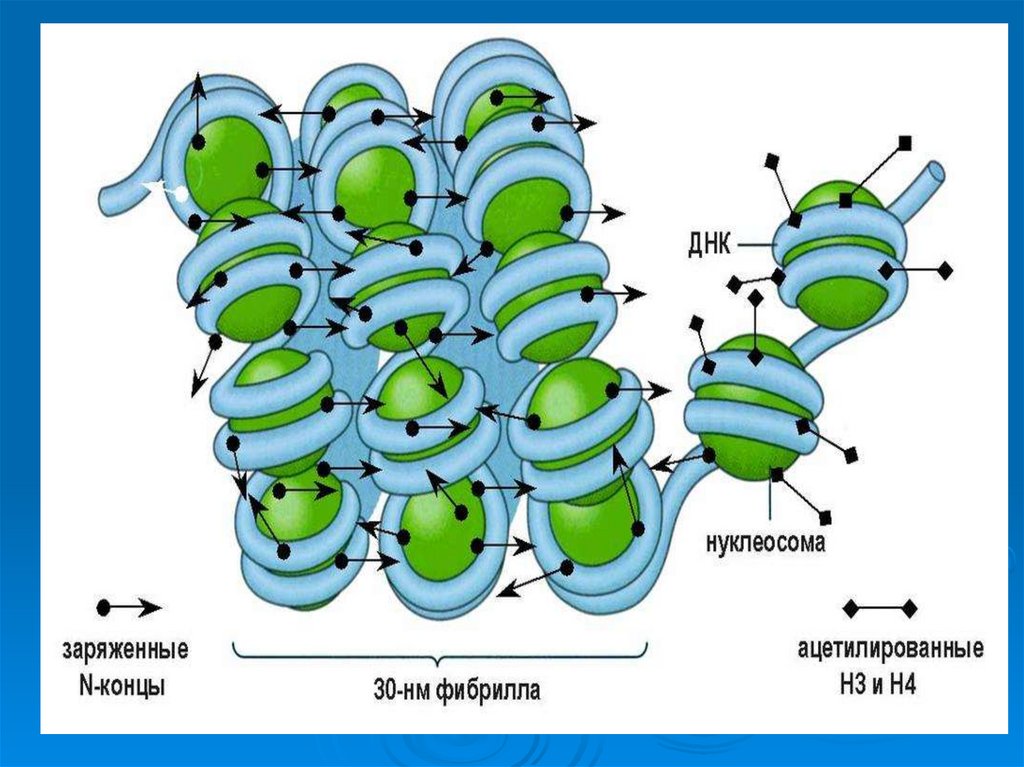
61. Второй уровень укладки ДНК -30 нм фибрилла
Второй уровень укладки ДНК -30 нмфибрилла
Второй
уровень организации хромосом – это
образование из нуклеосомной нити более толстых
фибрилл (20-35 нм.). Предполагают, что фибриллы
представляют по форме соленоиды, образующиеся
в результате скручивания нуклеосомной нити.
Шаг соленоида равен 11 нм, на один виток
приходится около 6-10 нуклеосом.
Соленоидная
упаковка
считается
наиболее
вероятной, однако существуют и другие модели
организации хроматина, например супербидная.
Согласно
последней
фибрилла
хроматина
диаметром 20-30 нм представляет собой цепь
гранул, или супербидов, каждая из которых состоит
из 8 нуклеосом.
В сумме 1-й и 2-й уровни обеспечивают уменьшение
линейных размером ДНК в 40-50 раз.
62.
63.
64.
65. Третий уровень компактизации – петлистые структуры
Фибриллаорганизуется
в
большие
(50
и
более
тысяч
пар
нуклеотидов)
петли,
концы
которых
закрепляются
на
белковом скелете ядра
(его часто называют
ядерным
матриксом).
На
этом
этапе
линейные размеры ДНК
сокращаются
в
700
раз.
66.
67.
68.
69.
Термин стэкинг в молекулярной химии относится к такомурасположению ароматических молекул, которое напоминает
расположение монет в стопке и поддерживается ароматическими
взаимодействиями. Наиболее популярный пример такого
расположения наблюдается в последовательных парах оснований ДНК.
Стэкинг также часто наблюдается в белках, когда два относительно
неполярных кольца имеют перекрывающиеся π-орбитали. Точная
природа таких взаимодействий (электростатическая или
неэлектростатическая) остаётся предметом обсуждений.
В ДНК параллельный стэкинг имеет место между соседними
парами нуклеотидов и повышает стабильность молекулярной
структуры. Азотистые основания нуклеотидов
имеют пуриновые или пиримидиновые группы в своем составе,
состоящие, в свою очередь, из ароматических колец. В молекуле ДНК
ароматические кольца расположены примерно перпендикулярно оси
спирали, поэтому их поверхности расположены параллельно, что
способствует перекрыванию p-орбиталей этих оснований.
Похожий эффект, именуемый T-стэкинг, часто наблюдается в белках,
когда атомы водорода с частичным положительным зарядом одной
ароматической системы направлены к центру другой ароматической
системы перпендикулярно плоскости последней.