



























































 Биология
БиологияПохожие презентации:


")







Прокариотическая клетка
1.
GeneticsAli Tilek
2.
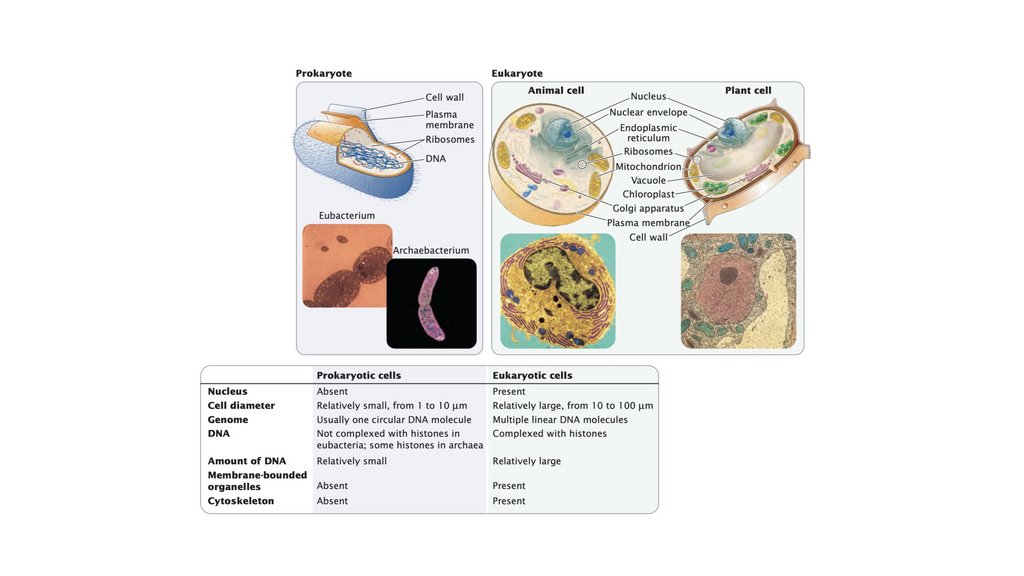
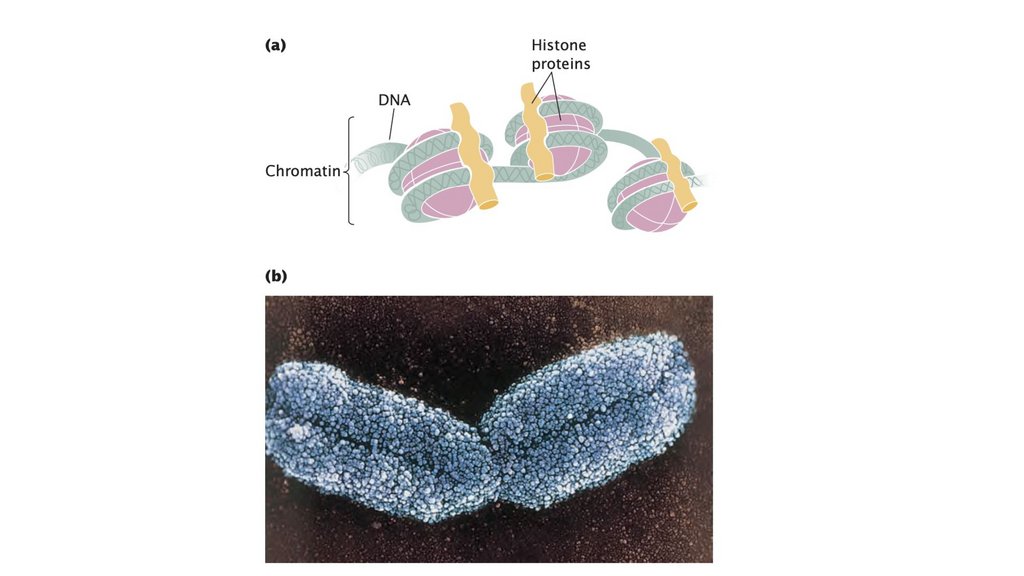
• Прокариотическая клетка обладает простой структурой, безядерной оболочки и, как правило, с единственной кольцевой
хромосомой. Эукариотическая клетка обладает более
сложной структурой, с ядром и множеством линейных
хромосом, состоящих из ДНК, связанной с белками гистонов.
3.
4.
5.
6.
7.
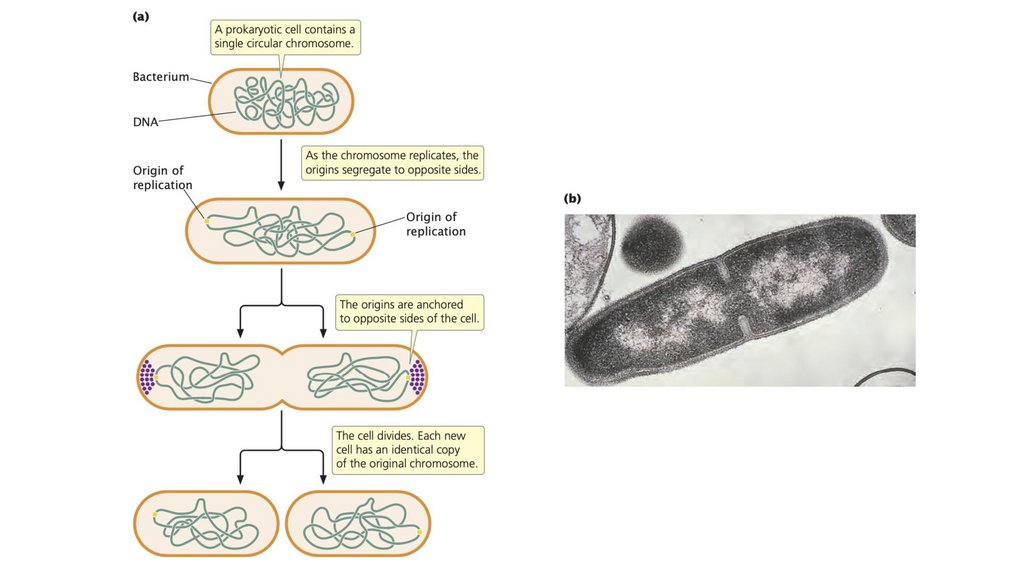
• Размножение клеток требует копирования генетическогоматериала, разделения копий и деления клеток.
8.
• В прокариотической клетке реплицируется одна хромосома,каждая копия перемещается в противоположные стороны
клетки, и клетка делится. В эукариотических клетках
размножение является более сложным процессом, чем в
прокариотических клетках, требующим митоза и мейоза для
обеспечения передачи полного набора генетической
информации каждой новой клетке.
9.
10.
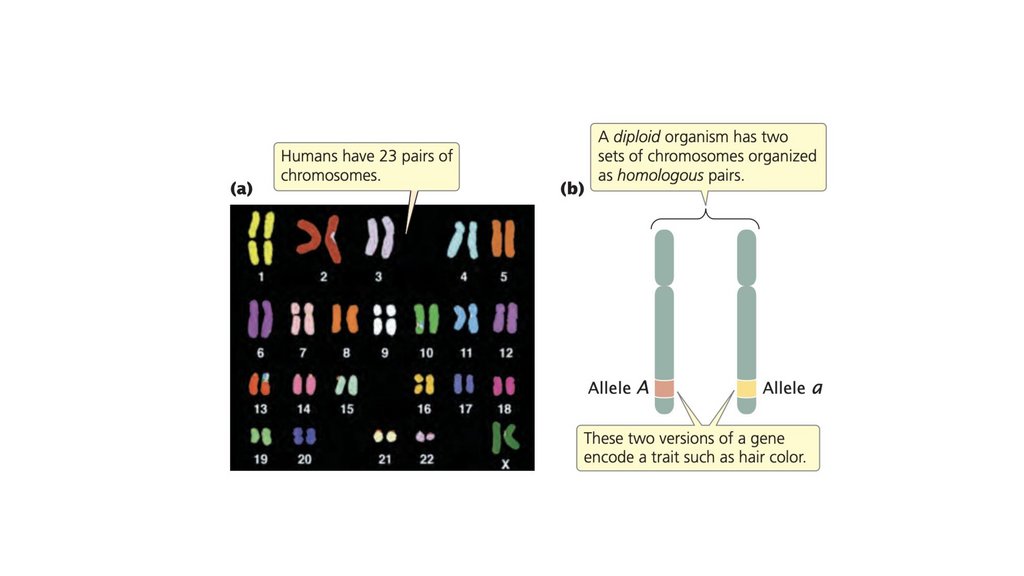
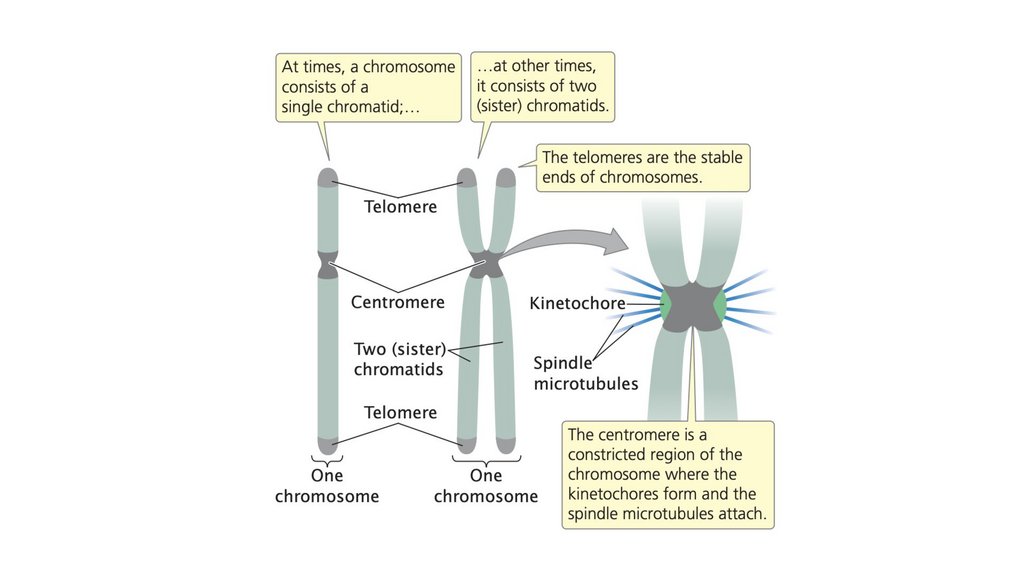
• В эукариотических клетках хромосомы обычно находятся вгомологичных парах. Каждая функциональная хромосома
состоит из центромеры, теломер и множества источников
репликации. После того, как хромосома была скопирована,
две копии остаются прикрепленными к центромере, образуя
сестринские хроматиды.
11.
12.
13.
14.
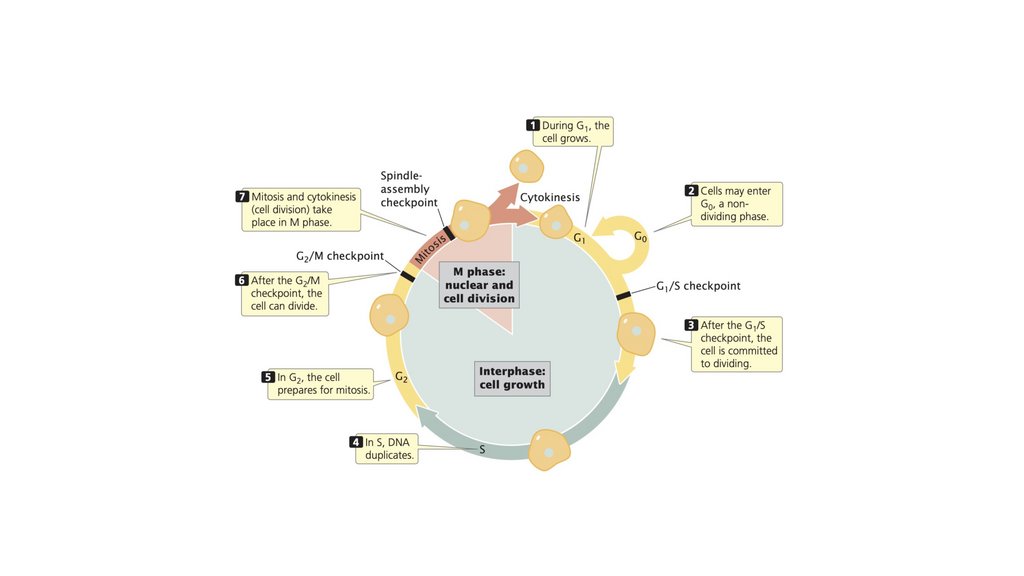
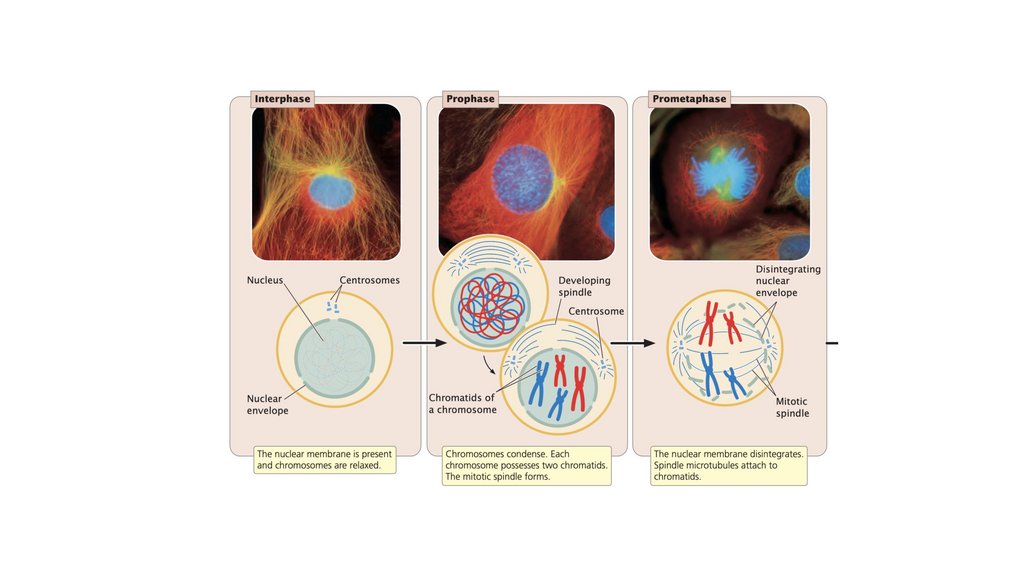
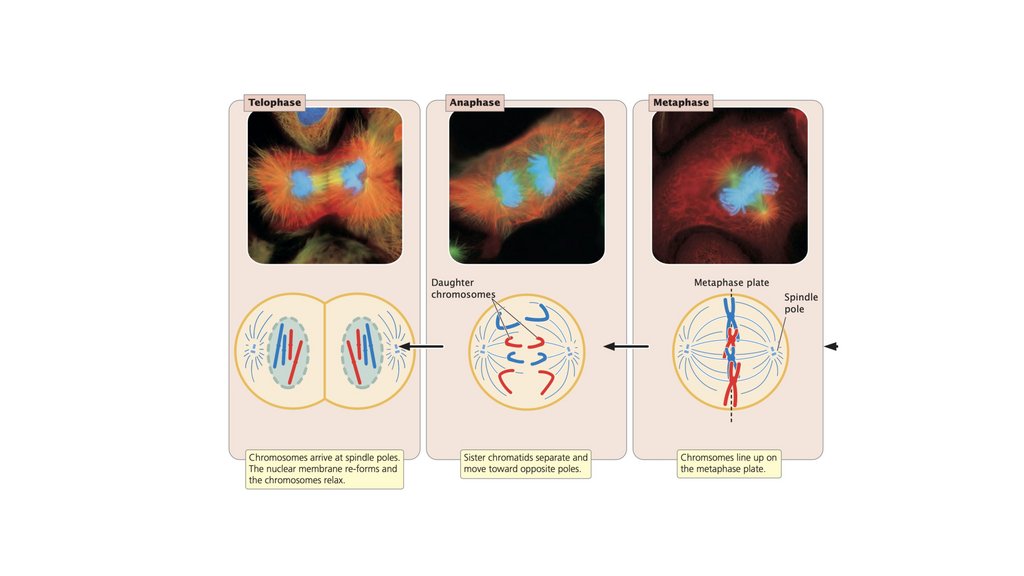
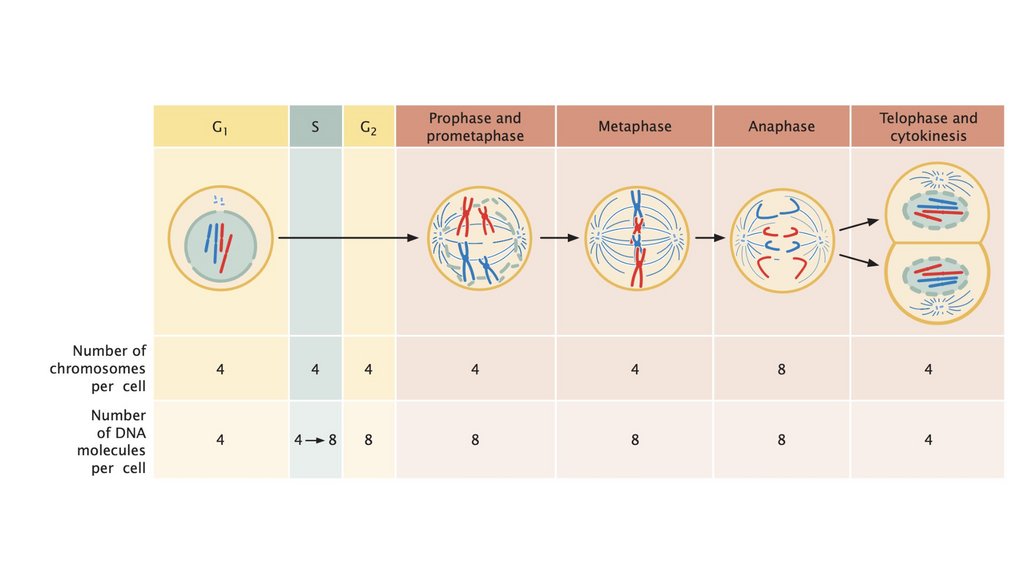
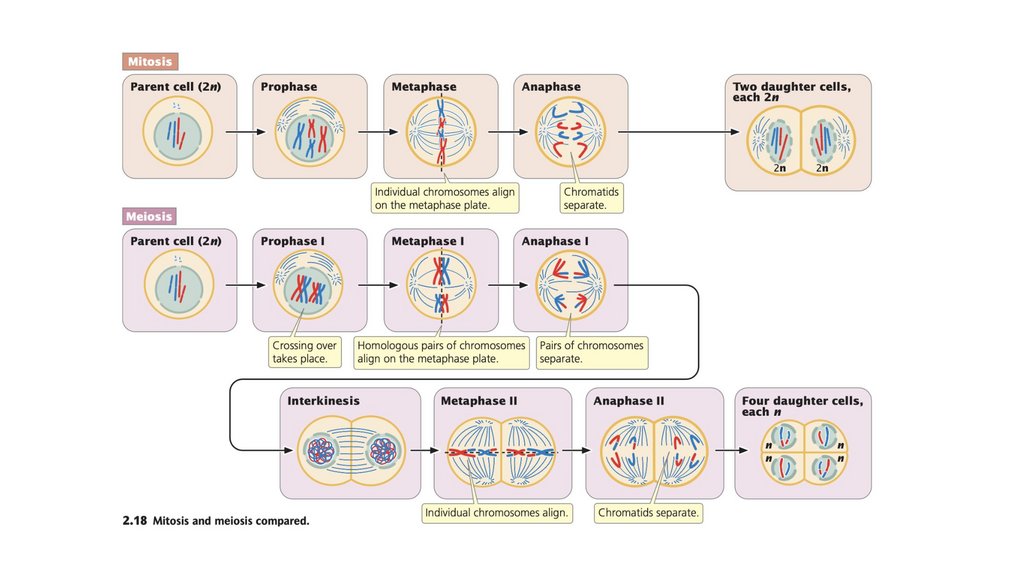
• Клеточный цикл состоит из стадий, через которые проходитэукариотическая клетка между клеточными делениями. Он
состоит из (1) интерфазы, в которой клетка растет и
готовится к делению, и (2) М-фазы, в которой происходит
ядерное и клеточное деление. Фаза М состоит из (1) митоза,
процесса деления ядра, и (2) цитокинеза, деления
цитоплазмы.
15.
• Прохождение клеточного цикла контролируетсяконтрольными точками, которые регулируют клеточный цикл,
позволяя или запрещая клетке переходить к следующей
стадии.
• Митоз обычно приводит к образованию двух генетически
идентичных клеток.
16.
17.
18.
19.
20.
21.
22.
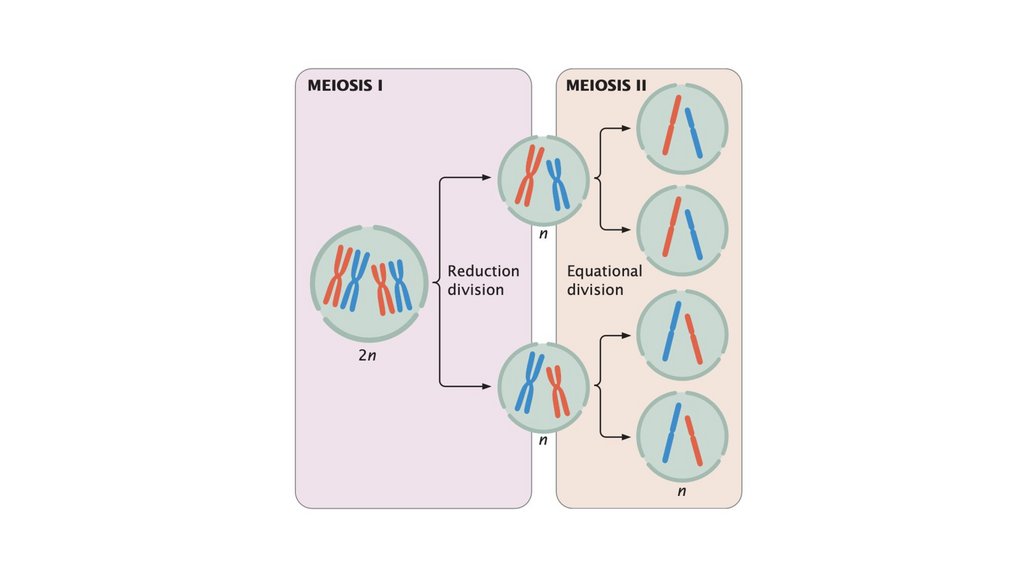
• Половое размножение производит генетически изменчивоепотомство и обеспечивает ускоренную эволюцию. Оно
включает мейоз, при котором образуются гаплоидные
половые клетки, и оплодотворение, слияние половых клеток.
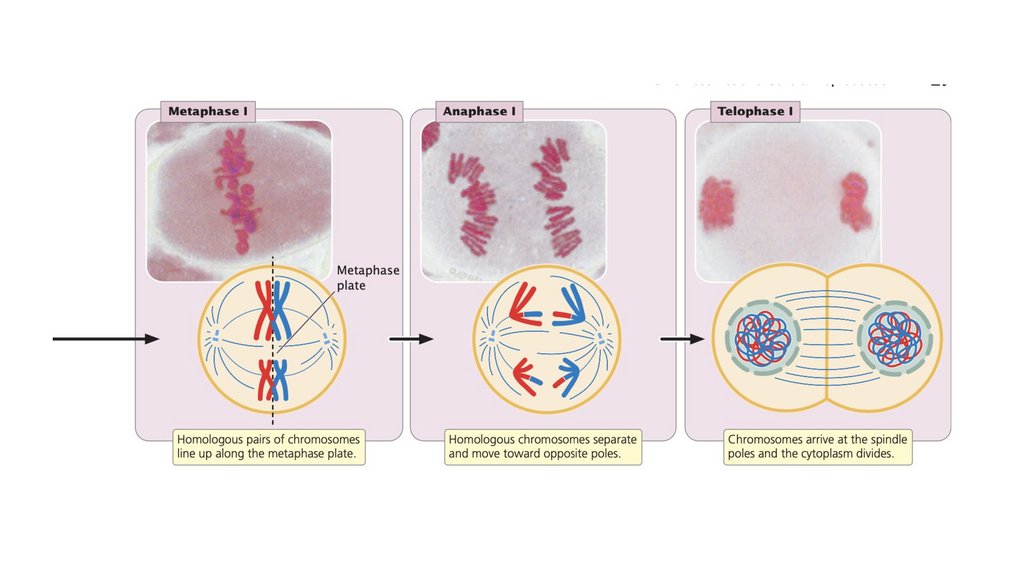
Мейоз включает в себя два клеточных деления. При мейозе I
происходит кроссинговер и гомологичные хромосомы
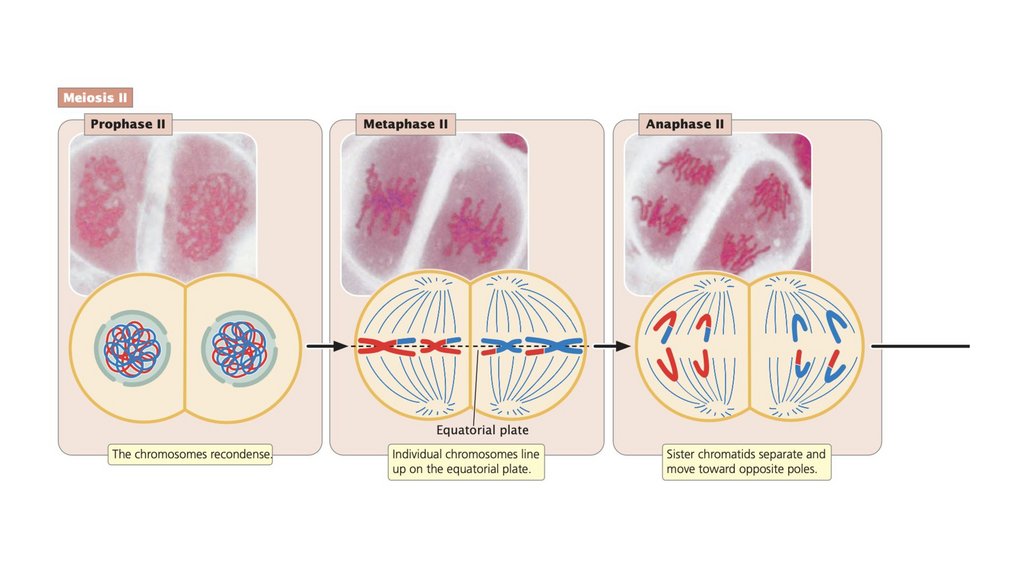
разделяются. При мейозе II хроматиды разделяются.
23.
24.
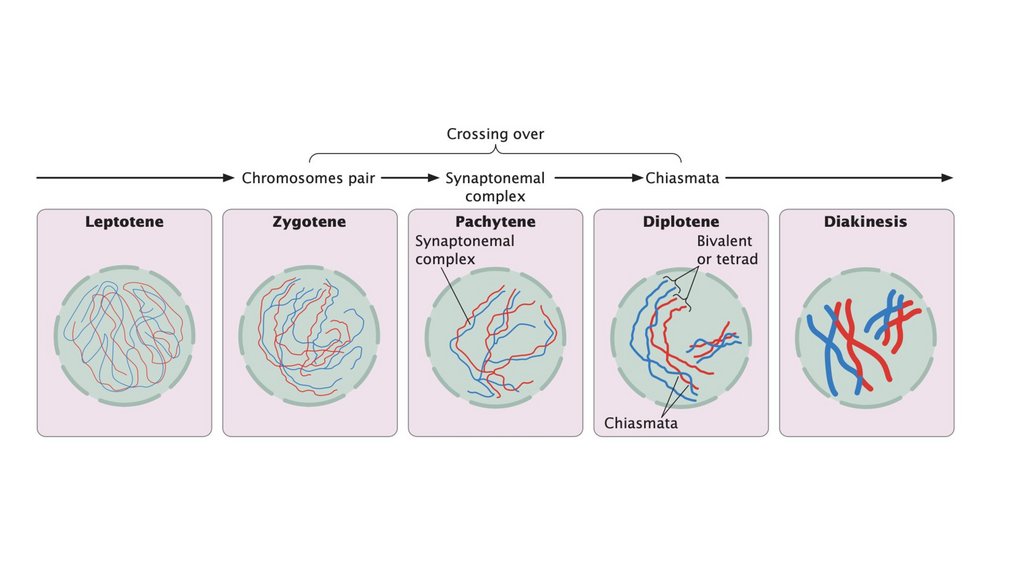
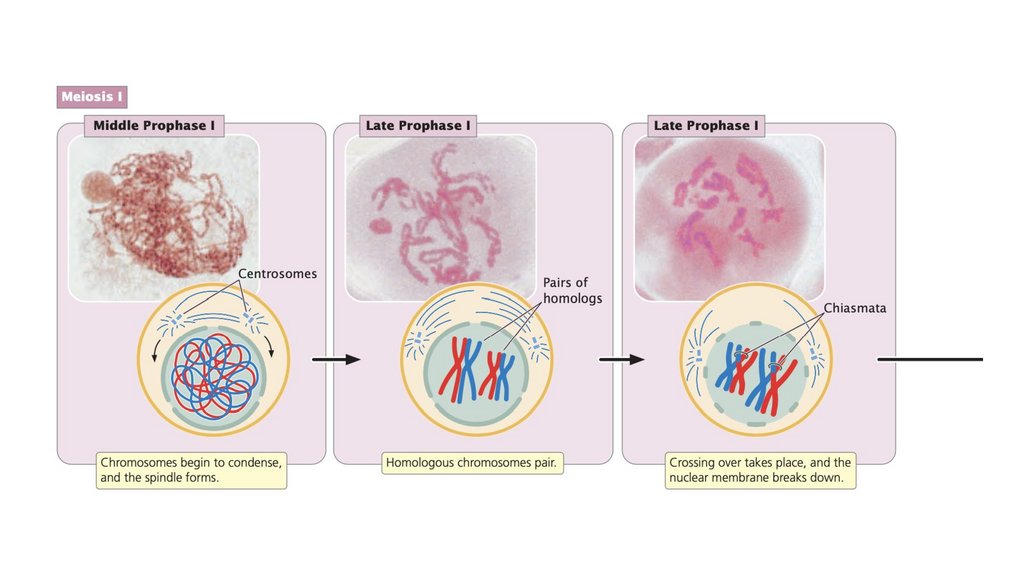
• Профаза I - это продолжительная стадия, разделенная на пятьподэтапов.
• В лептотене хромосомы сокращаются и становятся видимыми.
• В зиготине хромосомы продолжают конденсироваться;
гомологичные хромосомы объединяются в пары и начинают
синапсис, очень тесную ассоциацию.
• Каждая гомологичная пара синапсированных хромосом состоит
из четырех хроматид, называемых бивалентной или тетрадой.
• При пахитене хромосомы становятся короче и толще, и между
гомологичными хромосомами развивается синаптонемный
комплекс из трех частей. Функция синаптонемного комплекса
неясна, но хромосомы многих клеток с дефицитом этого
комплекса не разделяются должным образом.
25.
26.
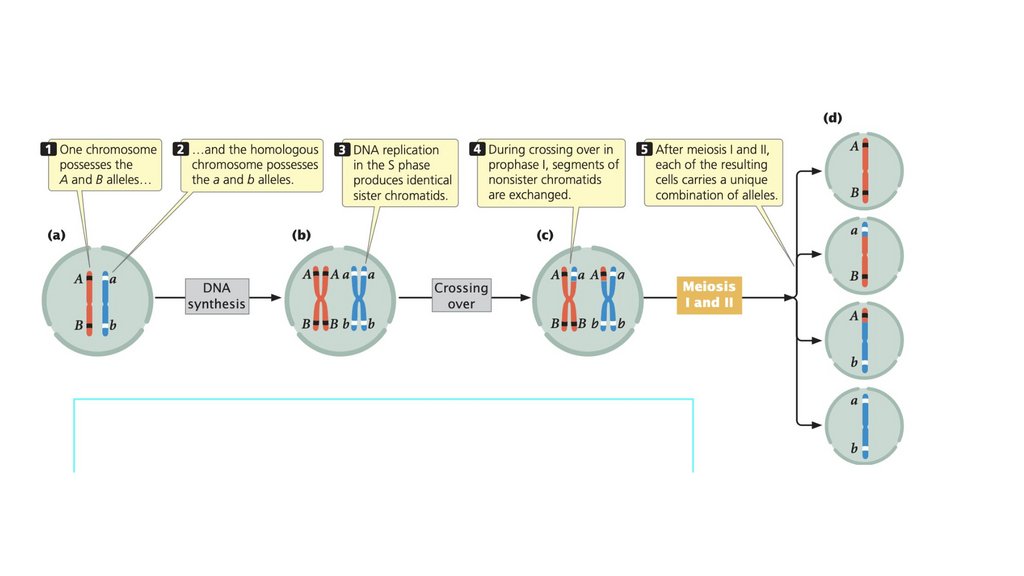
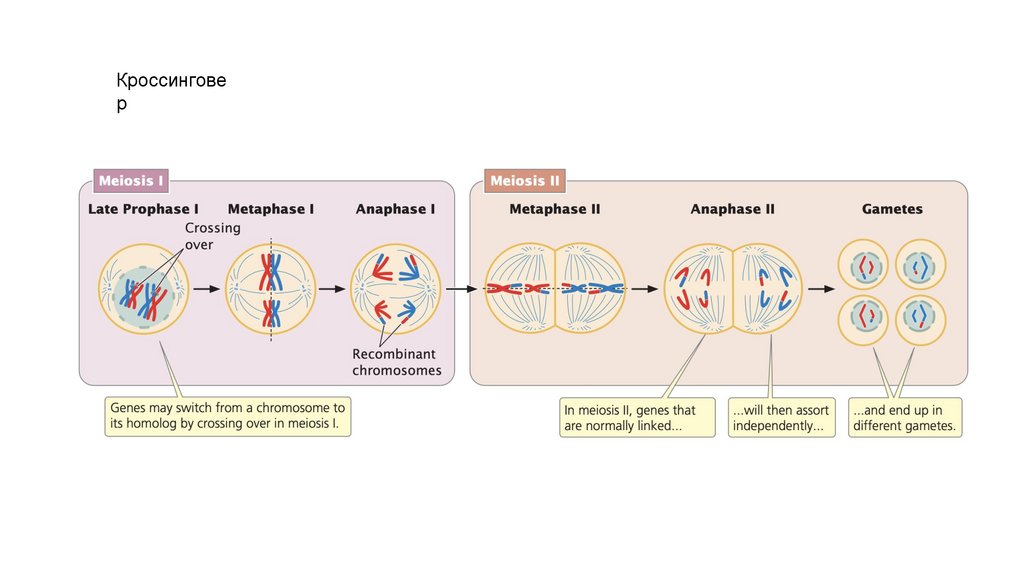
• Кроссинговер происходит в профазе I, в которой гомологичныехромосомы обмениваются генетической информацией.
• Кроссинговер порождает генетические вариации и необходим для
правильного выравнивания и разделения гомологичных
хромосом.
• Центромеры парных хромосом раздвигаются в диплотене; два
гомолога остаются прикрепленными в каждой хиазме что
является результатом кроссинговера. При диакинезе
продолжается конденсация хромосом, и хиазматы перемещаются
к концам хромосом по мере того, как нити раздвигаются; таким
образом, гомологи остаются спаренными только на кончиках.
Ближе к концу профазы I ядерная мембрана разрушается и
образуется веретено, подготавливая почву для метафазы I.
27.
28.
29.
30.
31.
32.
33.
34.
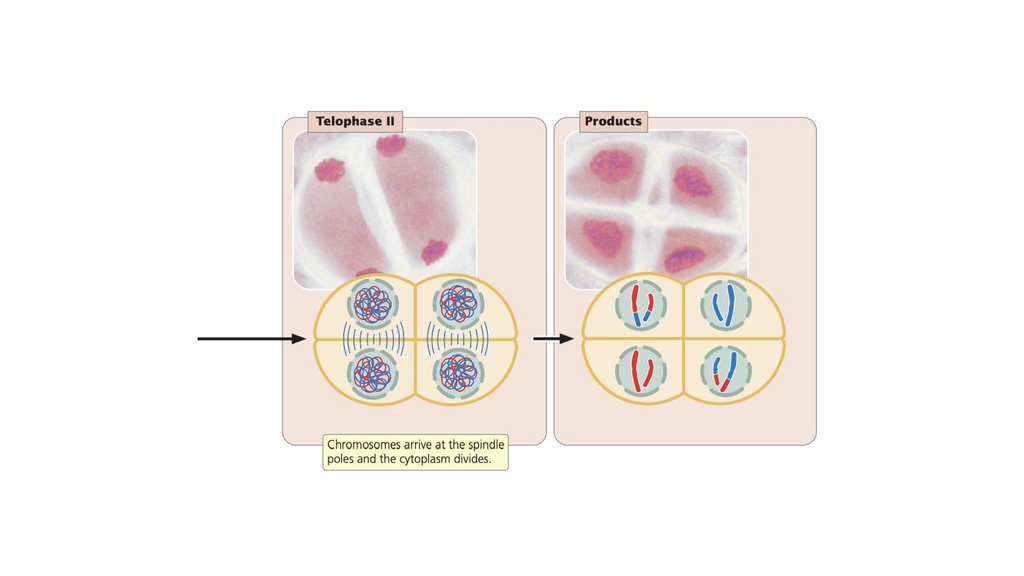
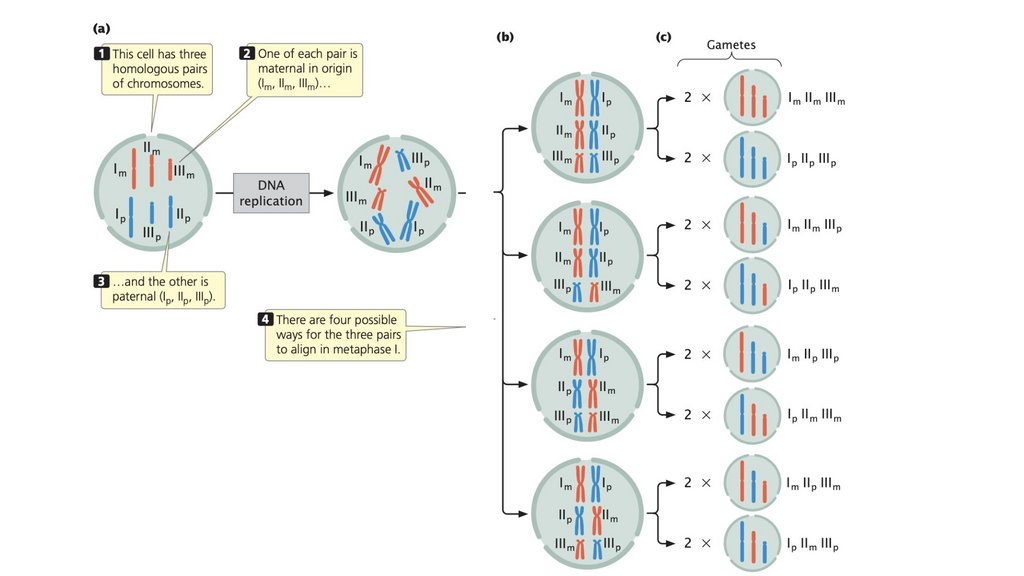
• Обычным результатом мейоза является образованиечетырех гаплоидных клеток, которые генетически
изменчивы. Генетическая изменчивость при мейозе
возникает в результате скрещивания и случайного
распределения материнских и отцовских хромосом.
35.
36.
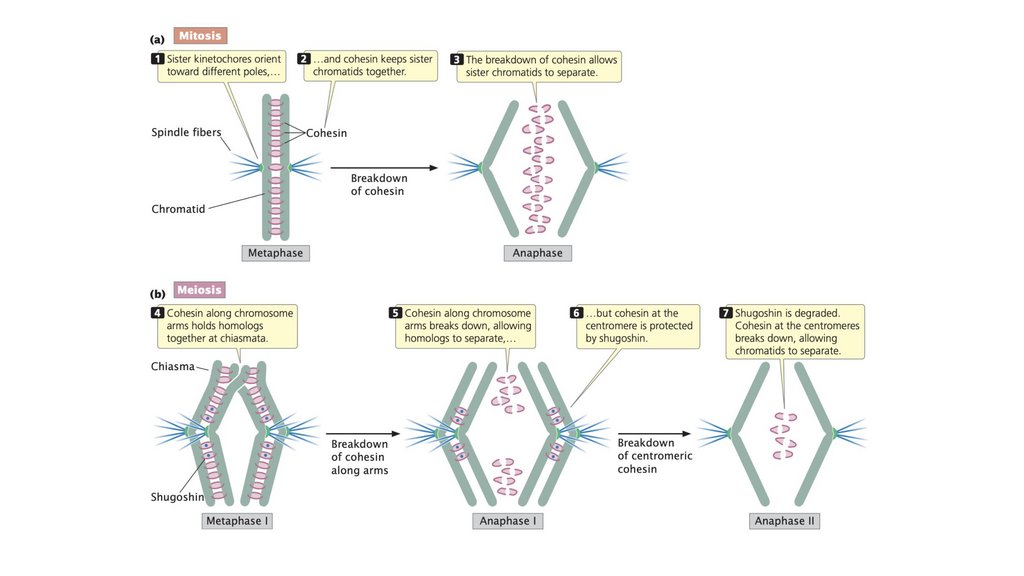
• Когезин удерживает сестринские хроматиды вместе. Вметафазе митоза и в метафазе II мейоза разрушение
когезина позволяет сестринским хроматидам разделяться.
При мейозе I центромерный когезин остается интактным и
удерживает сестринские хроматиды вместе, так что
гомологичные хромосомы, но не сестринские хроматиды,
разделяются в анафазе I.
37.
38.
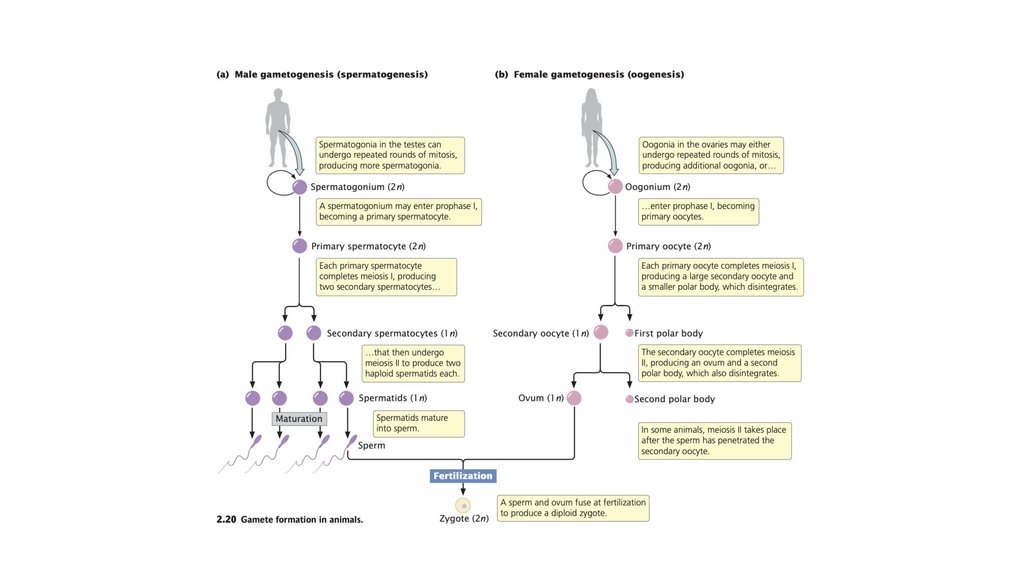
• У животных диплоидный сперматогоний подвергаетсямейозу с образованием четырех гаплоидных
сперматозоидов. Диплоидный оогоний подвергается мейозу
с образованием одной крупной гаплоидной яйцеклетки и
одного или нескольких меньших полярных телец.
39.
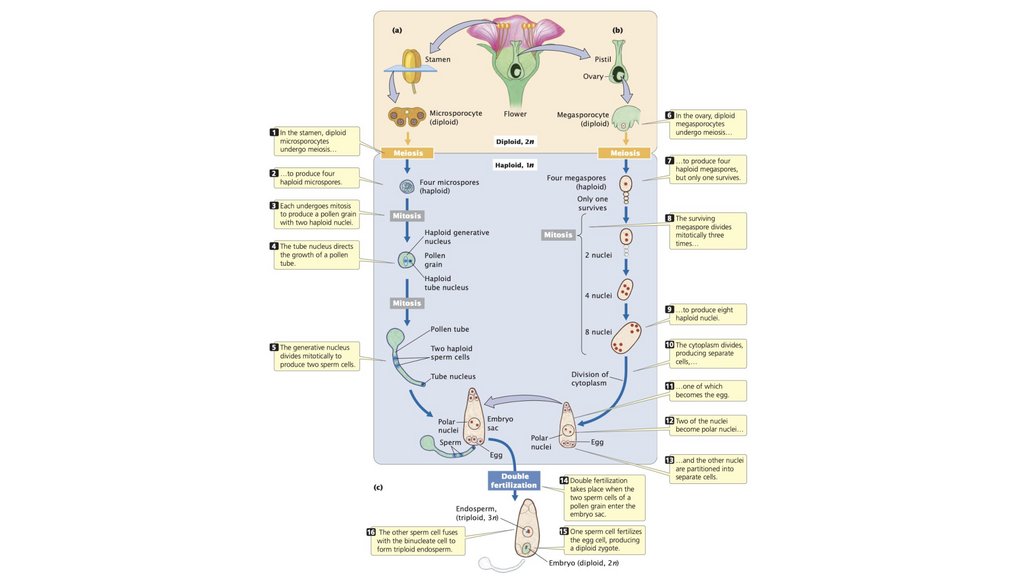
40.
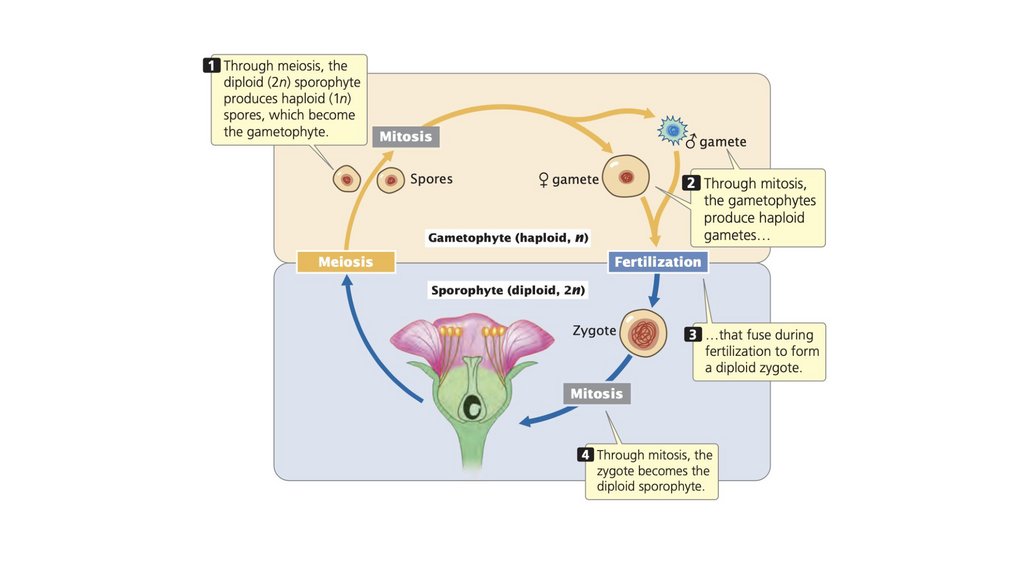
• У растений диплоидный микроспороцит в тычинкеподвергается мейозу с образованием четырех пыльцевых
зерен, каждое из которых содержит по два гаплоидных
сперматозоида. В яичнике диплоидный мегаспороцит
подвергается мейозу с образованием восьми гаплоидных
ядер, одно из которых образует яйцеклетку. Во время
опыления один сперматозоид оплодотворяет яйцеклетку, а
другой сливается с двумя гаплоидными ядрами, образуя
эндосперм 3n.
41.
42.
43.
44.
45.
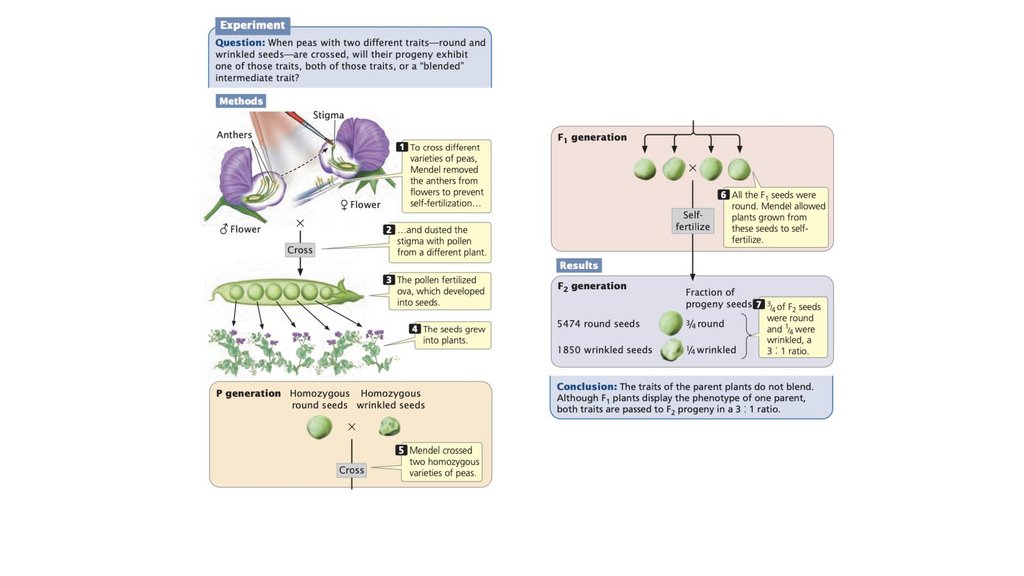
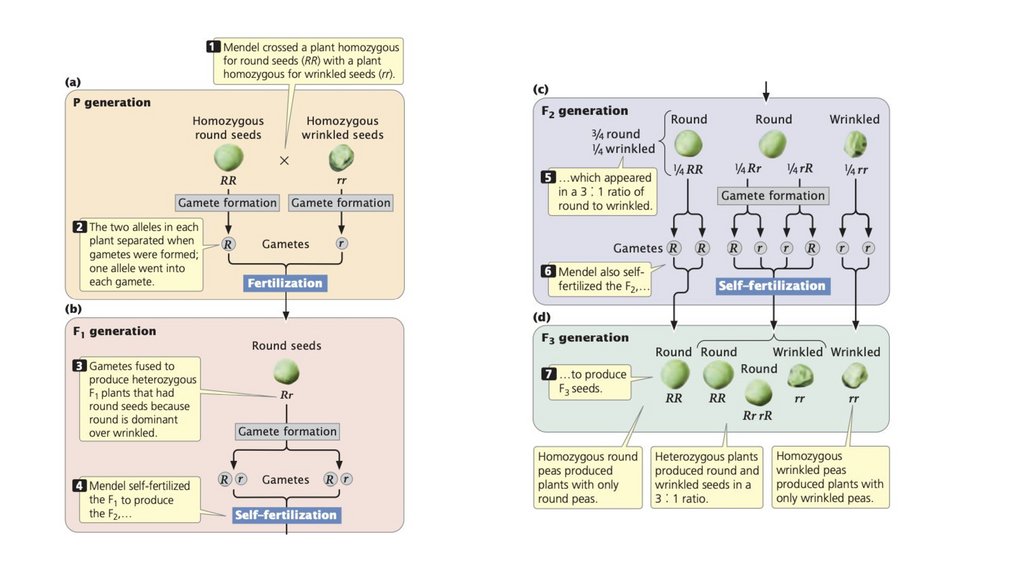
• Грегор Мендель открыл принципы наследственности. Егоуспех можно объяснить выбором растения горох в качестве
экспериментального организма, использованием признаков
с несколькими легко различимыми фенотипами, его
экспериментальным подходом, использованием математики
для интерпретации результатов и тщательным вниманием к
деталям.
46.
47.
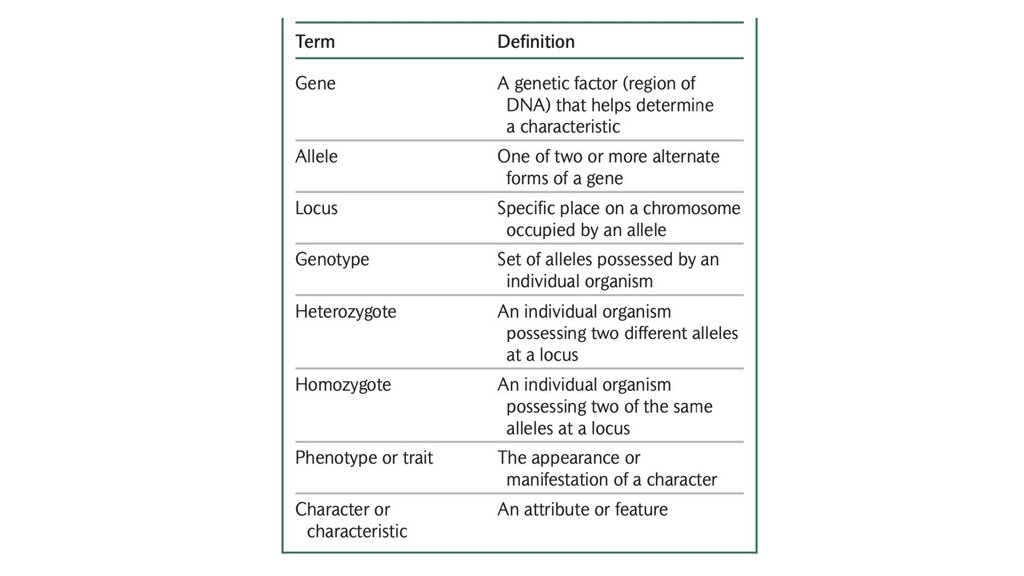
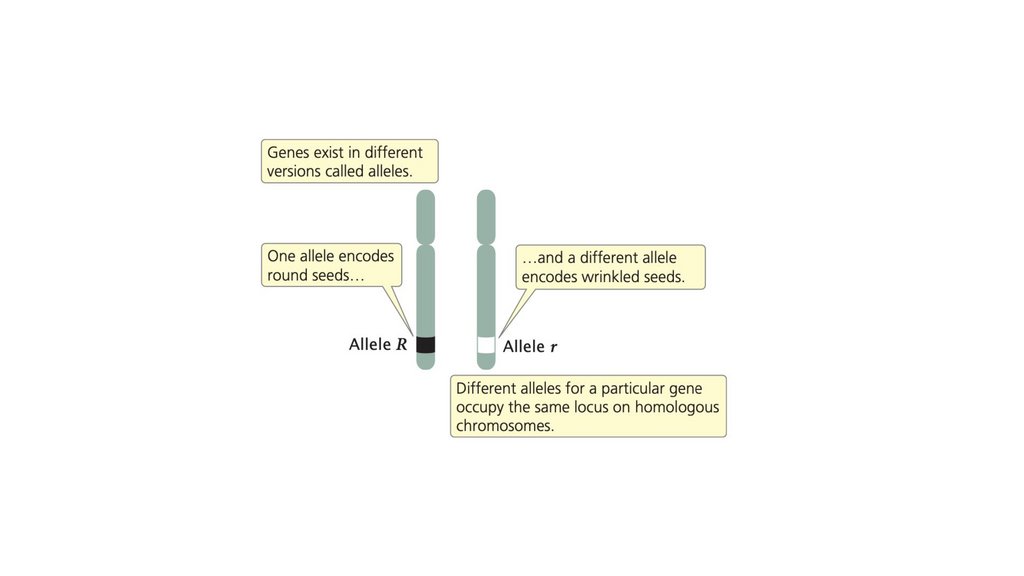
• Гены - это наследуемые факторы, которые определяютхарактеристику. Альтернативные формы гена называются
аллелями. Аллели расположены в определенном месте,
локусе, на хромосоме, а набор генов, которым обладает
отдельный организм, является его генотипом. Фенотип - это
проявление или видимость признака и может относиться к
физической, биохимической или поведенческой
характеристике. Наследуется только генотип, а не фенотип.
48.
49.
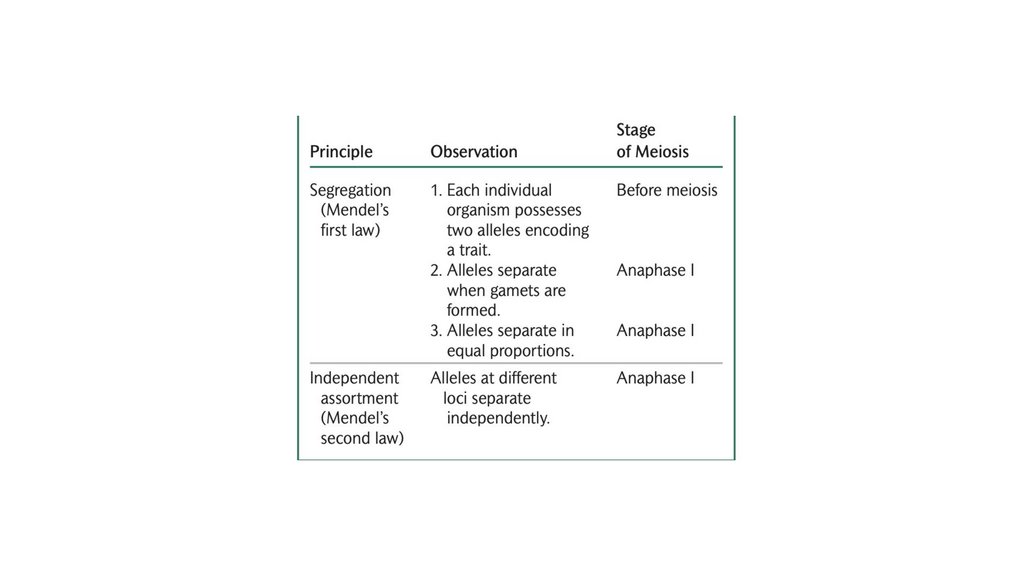
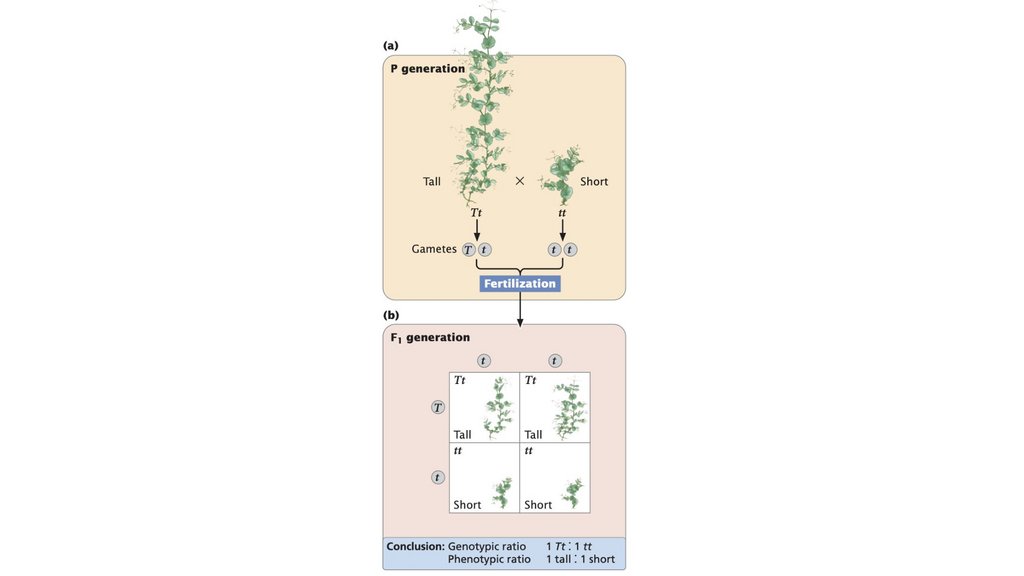
• Принцип сегрегации гласит, что отдельный организмобладает двумя аллелями, кодирующими признак, и что эти
две аллели разделяются в равных пропорциях при
формировании гамет.
50.
51.
• Концепция доминантности указывает на то, что, когда вгетерозиготе присутствуют два разных аллеля, в фенотипе
наблюдается только признак одного из них, доминантный
аллель. Другой аллель называется рецессивным.
52.
53.
54.
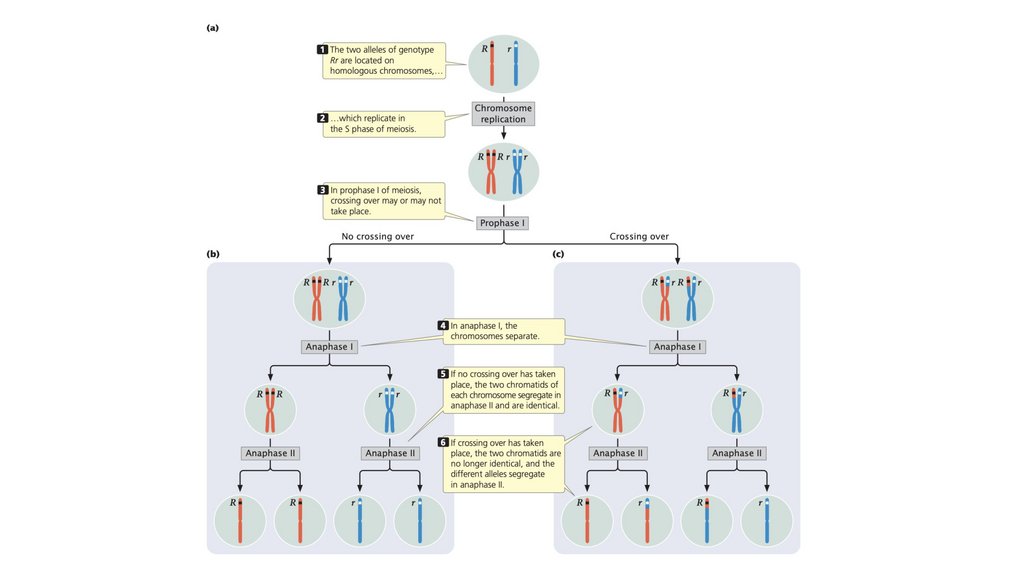
• Два аллеля генотипа расположены на гомологичныххромосомах. Разделение гомологичных хромосом в анафазе
I мейоза приводит к сегрегации аллелей.
55.
56.
57.
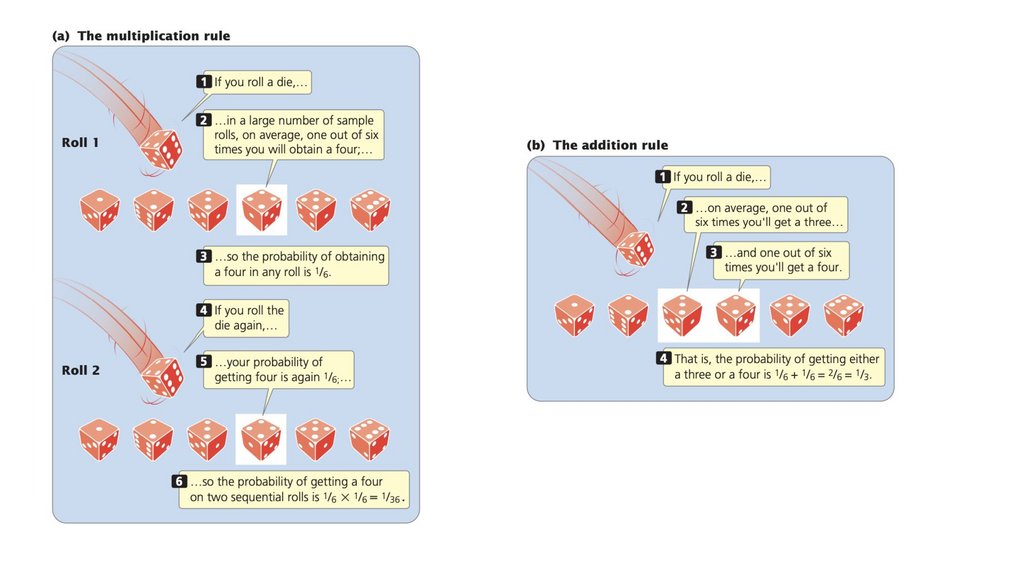
• Вероятность - это вероятность того, что произойдетконкретное событие. Правило умножения вероятности
гласит, что вероятность того, что два или более независимых
события произойдут вместе, вычисляется путем умножения
вероятностей независимых событий. Правило сложения
вероятностей гласит, что вероятность возникновения любого
из двух или более взаимоисключающих событий
вычисляется путем сложения вероятностей этих событий.
58.
59.
• Биномиальное разложение может быть использовано дляопределения вероятности определенной комбинации
событий.
60.
61.
62.
63.
64.
65.
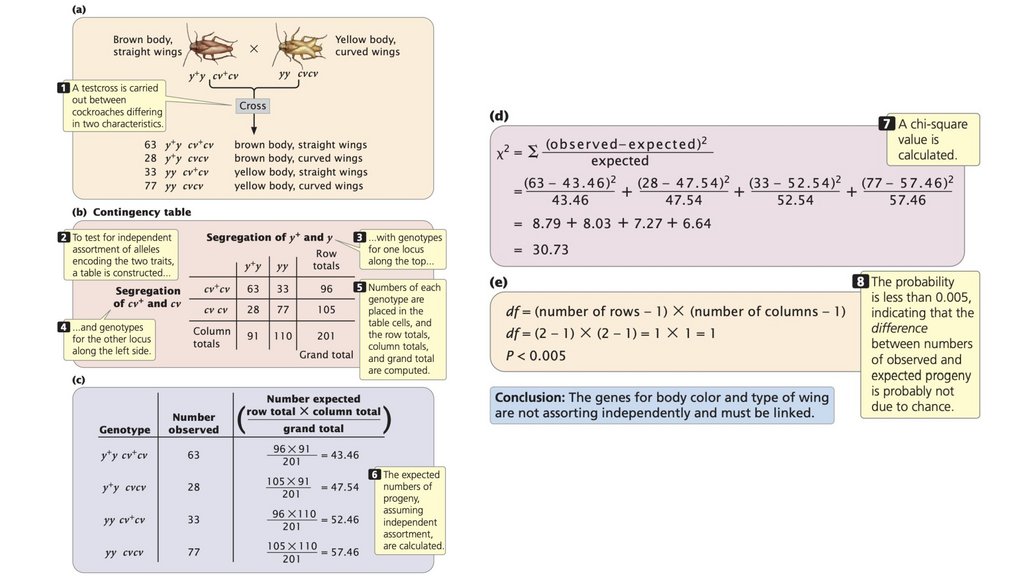
• Тест-кросс выявляет генотип (гомозиготный илигетерозиготный) отдельного организма, обладающего
доминантным признаком, и состоит из скрещивания этого
организма с особью, имеющей гомозиготный рецессивный
генотип.
66.
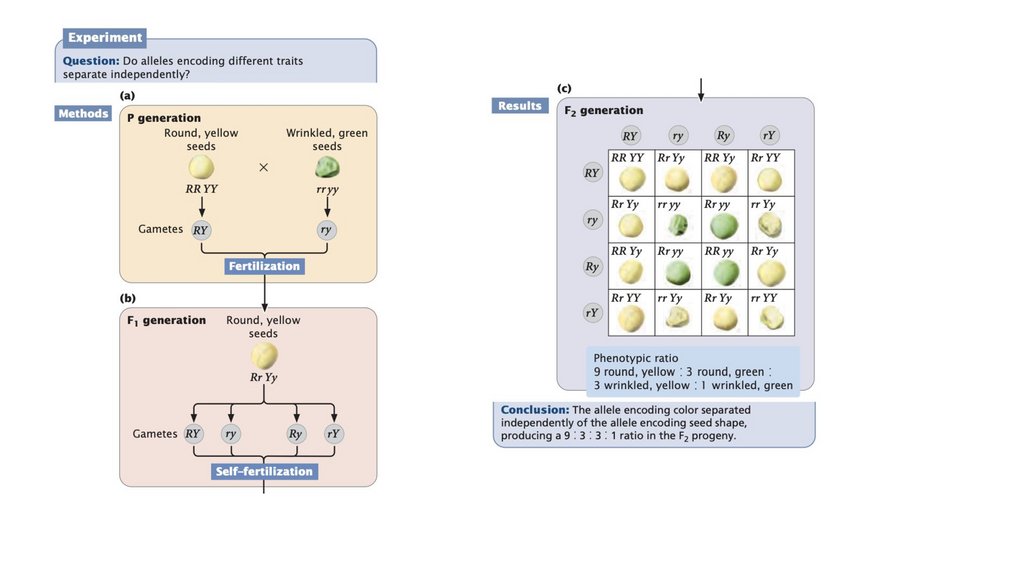
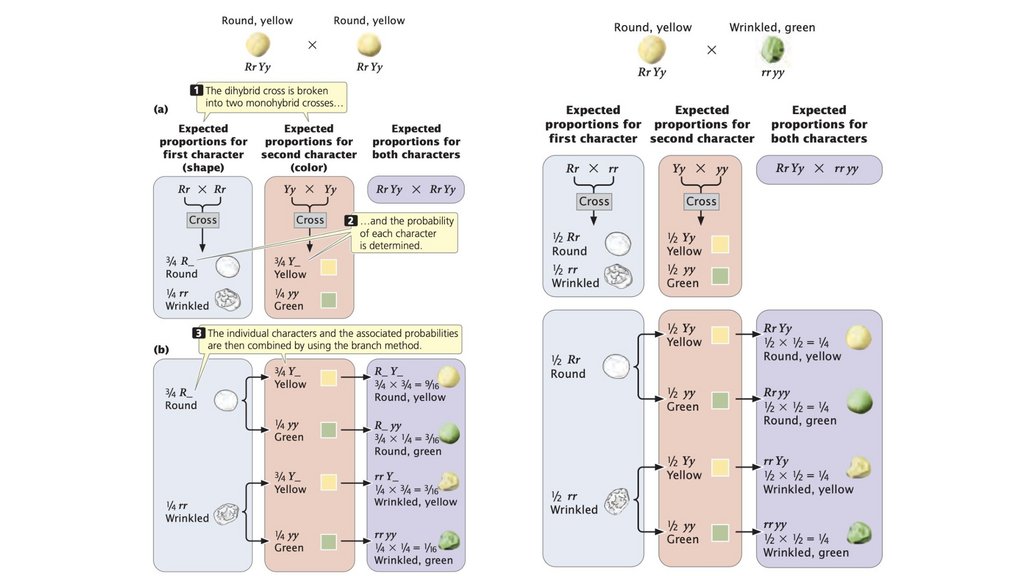
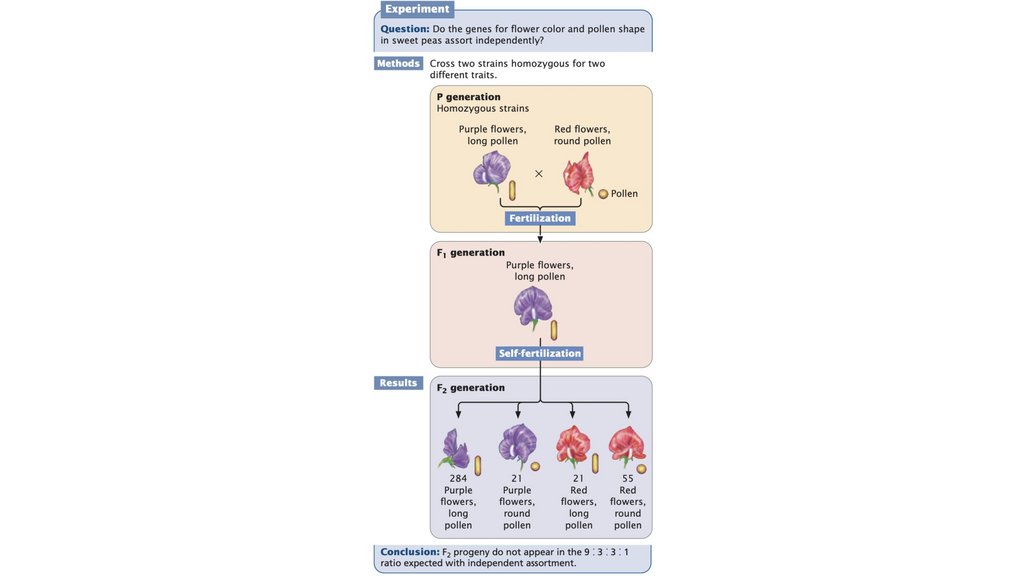
• Принцип независимой сортировки гласит, что гены,кодирующие разные признаки, сортируются независимо при
формировании гамет. Независимая сортировка основана на
случайном разделении гомологичных пар хромосом в
анафазе I мейоза; это происходит, когда гены, кодирующие
два признака, расположены на разных парах хромосом.
67.
68.
69.
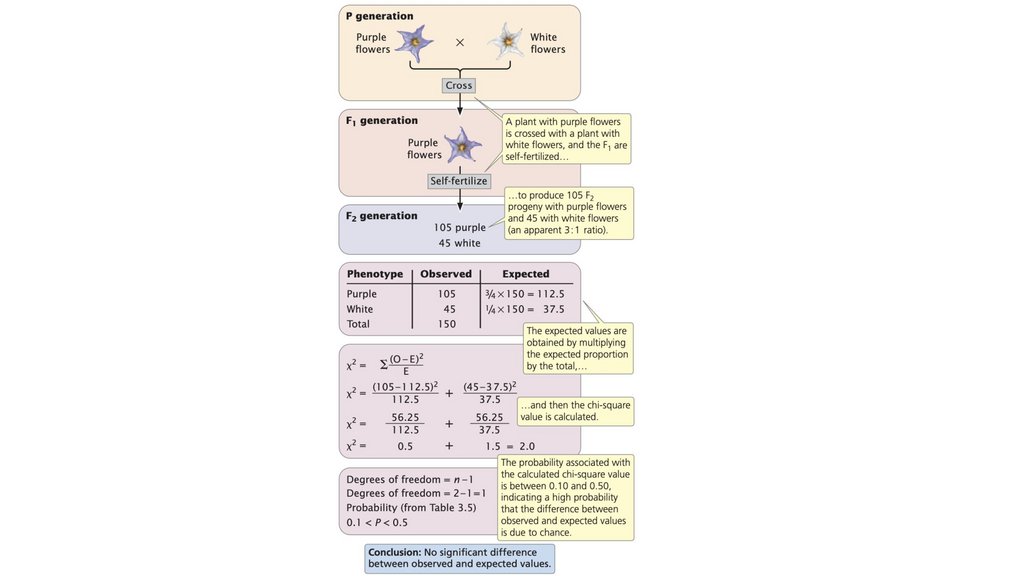
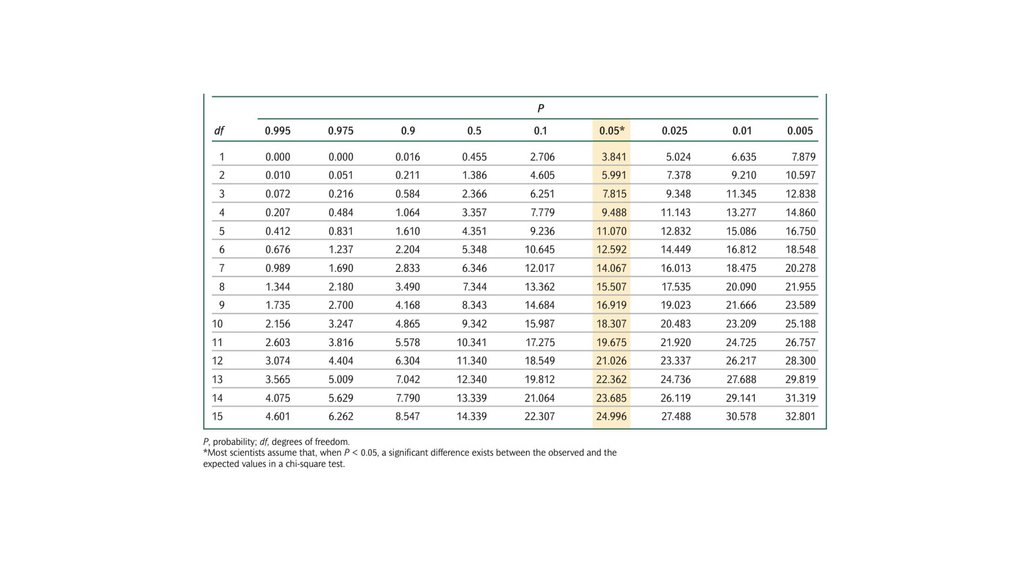
• Наблюдаемые соотношения потомства от генетическогоскрещивания могут отличаться от ожидаемых соотношений
из-за случайности. Критерий соответствия хи-квадрат может
быть использован для определения вероятности того, что
разница между наблюдаемыми и ожидаемыми числами
обусловлена случайностью.
70.
• Большинство ученых используют уровень вероятности 0,05 вкачестве своего предельного значения: если вероятность
того, что случайность ответственна за отклонение, больше
или равна 0,05, они допускают, что случайность может быть
ответственна за отклонение между наблюдаемыми и
ожидаемыми значениями. Когда вероятность меньше 0,05,
ученые предполагают, что случайность здесь ни при чем и
существует значительная разница. Выражение значительная
разница означает, что какой-то фактор, отличный от
случайности, ответственен за то, что наблюдаемые значения
отличаются от ожидаемых значений.
71.
• Выбрав 0,05 в качестве предельного значения, ученыесогласились предположить, что за отклонения между
наблюдаемыми и ожидаемыми значениями отвечает
случайность, если только нет убедительных доказательств
обратного. Имейте в виду, что, даже если мы получим
вероятность, скажем, 0,01, все равно существует 1%-ная
вероятность того, что отклонение между наблюдаемыми и
ожидаемыми числами вызвано не более чем случайностью.
72.
73.
74.
75.
• Половое размножение - это производство потомства, генетическиотличного от своих родителей. Большинство организмов имеют два
половых фенотипа — самцов и самок. Самцы производят маленькие
гаметы; самки производят большие гаметы.
• Механизм, с помощью которого определяется пол, называется
определением пола. Пол может определяться различиями в конкретных
хромосомах, генотипах или окружающей среде.
• Половые хромосомы мужчин и женщин различаются по количеству и
внешнему виду. Гомогаметный пол производит гаметы, которые все
идентичны в отношении половых хромосом; гетерогаметный пол
производит гаметы, которые отличаются по своему составу половых
хромосом.
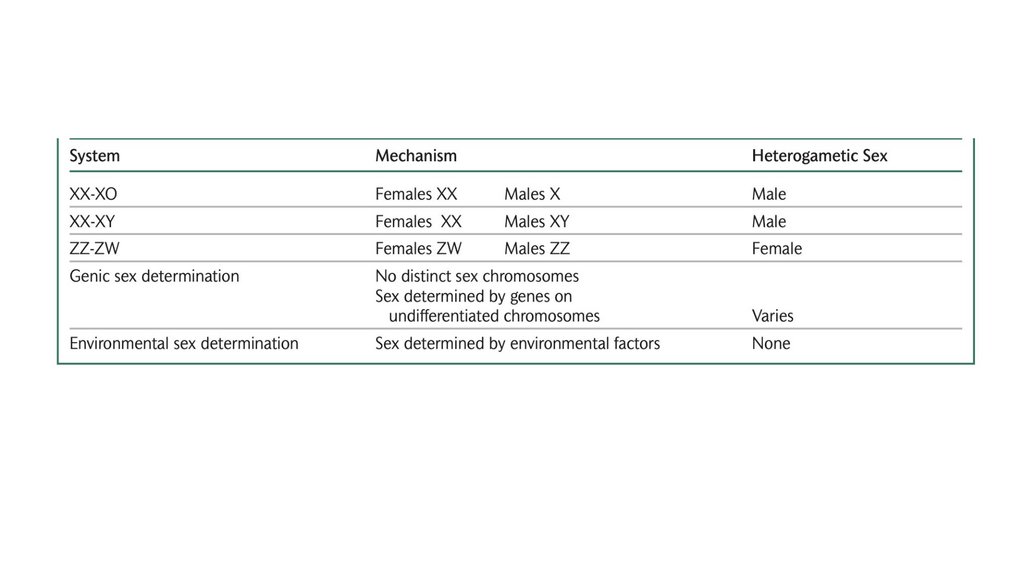
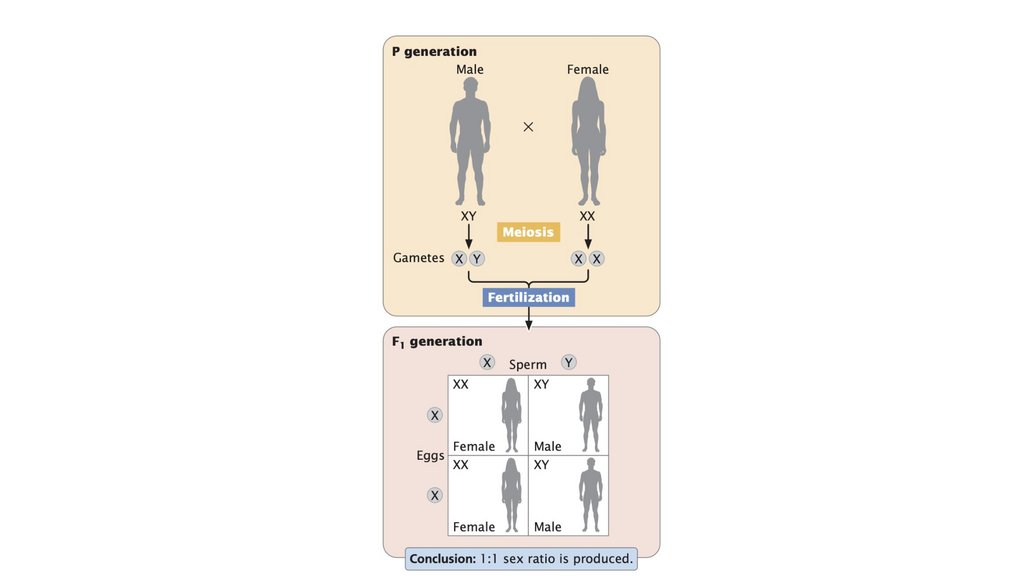
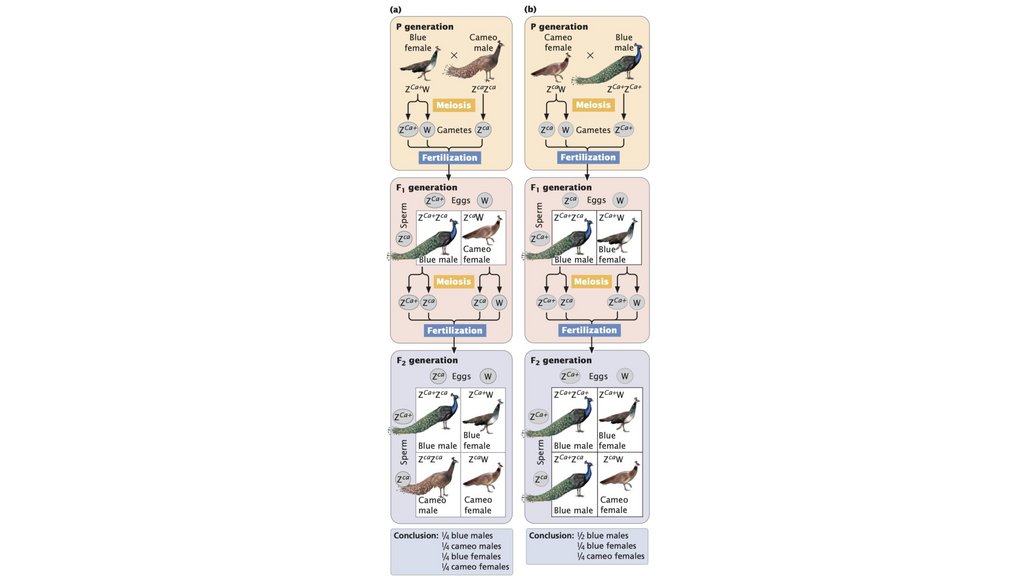
• В системе определения пола XX-XO женщины обладают двумя Xхромосомами, тогда как мужчины обладают одной X-хромосомой. В
системе XX-XY женщины обладают двумя X-хромосомами, тогда как
мужчины обладают одной X-хромосомой и одной Y-хромосомой. В
системе ZZ-ZW самцы обладают двумя Z-хромосомами, тогда как самки
обладают Z-хромосомой и W-хромосомой.
76.
• У некоторых организмов есть генетическая детерминацияпола, при которой генотипы в одном или нескольких локусах
определяют пол отдельного организма. У других есть
экологическая детерминация пола.
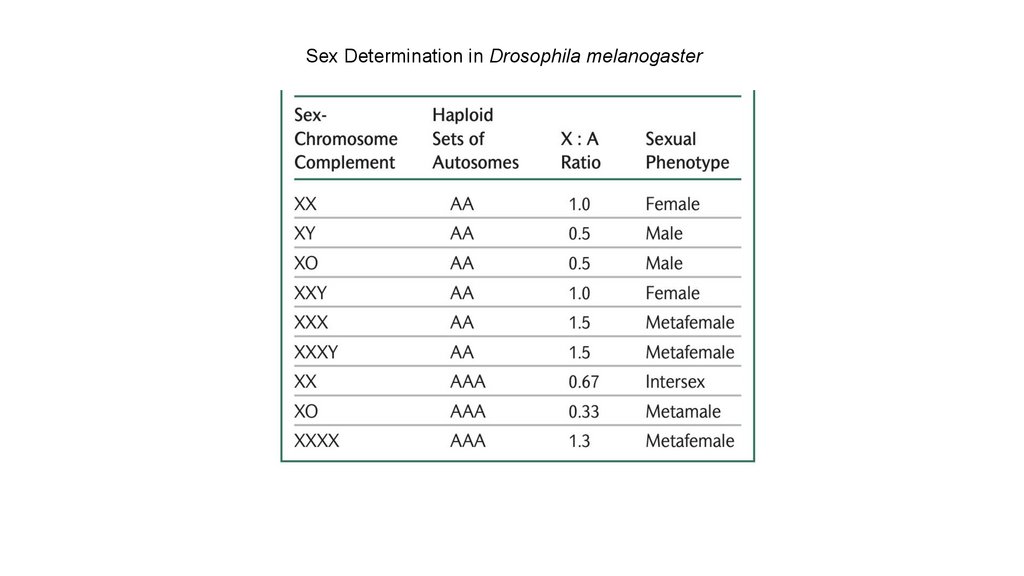
• У Drosophila melanogaster пол определяется балансом между
генами на X-хромосомах и генами на аутосомах,
соотношением X:A.
77.
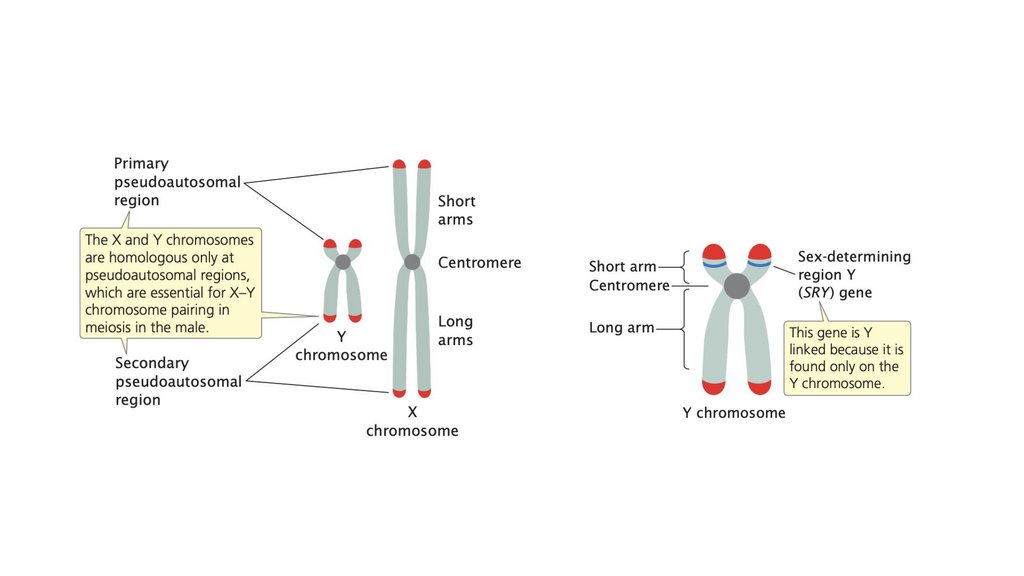
• У людей пол в конечном счете определяется наличием илиотсутствием гена SRY, расположенного на Y-хромосоме.
• Характеристики, связанные с полом, определяются генами на
половых хромосомах; характеристики, связанные с X, кодируются
генами на X-хромосоме, а характеристики, связанные с Y,
кодируются генами на Y-хромосоме.
• Самка наследует Х-сцепленные аллели от обоих родителей;
самец наследует Х-сцепленные аллели только от своей
родительницы.
• Половые хромосомы эволюционировали из аутосом.
Кроссинговер между X и Y-хромосомами был подавлен, но
палиндромные последовательности внутри Y-хромосомы
допускают внутреннюю рекомбинацию на Y-хромосоме. Эта
внутренняя рекомбинация иногда приводит к хромосомным
перестройкам, которые могут отрицательно сказаться на половом
развитии.
78.
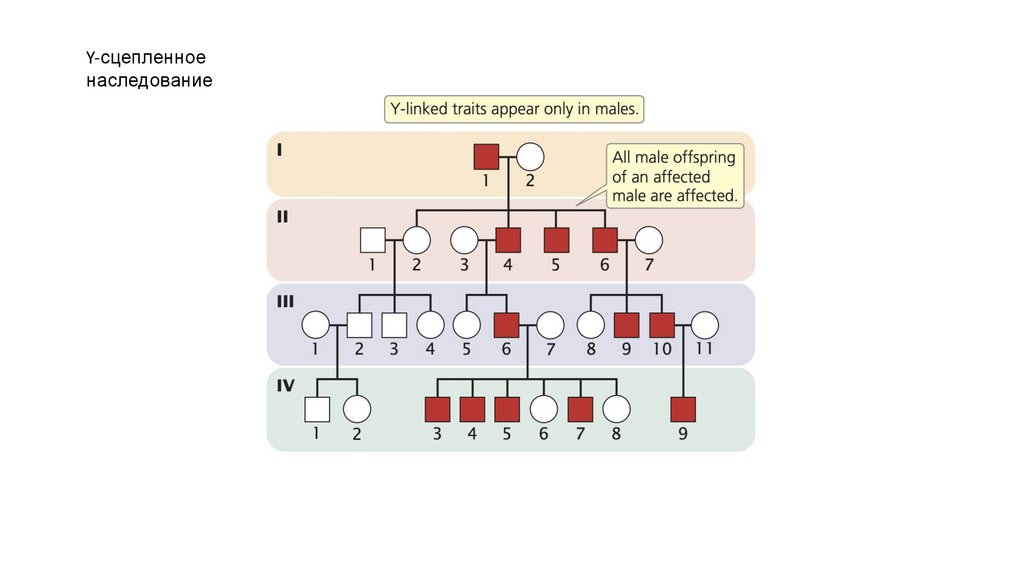
• Y-сцепленные признаки встречаются только у самцов ипередаются от отца ко всем сыновьям.
• Компенсация дозы выравнивает количество белка,
вырабатываемого X-сцепленными генами у самцов и самок.
У плацентарных млекопитающих одна из двух X-хромосом у
самок обычно становится инактивированной. Какая именно
Х-хромосома инактивирована, является случайным и
варьируется от клетки к клетке. Некоторые Х-сцепленные
гены избегают Х-инактивации, а другие Х-сцепленные гены
могут быть инактивированы у одних женщин, но не у других.
Х-инактивация контролируется геном Xist.
79.
80.
81.
82.
83.
84.
Sex Determination in Drosophila melanogaster85.
86.
87.
88.
89.
В результате инактивации X самкифункционально гемизиготны на клеточном
уровне по X-сцепленным генам. У самок,
гетерозиготных по X-сцепленному локусу,
приблизительно 50% клеток будут
экспрессировать один аллель, а 50% - другой
аллель; таким образом, у гетерозиготных
самок продуцируются белки, кодируемые
обоими аллелями, хотя и не в пределах одной
и той же клетки.
90.
91.
• Доминирование всегда относится к генам в одном и том желокусе (аллельные гены) и может быть понято в отношении
того, как фенотип гетерозиготы соотносится с фенотипами
гомозигот.
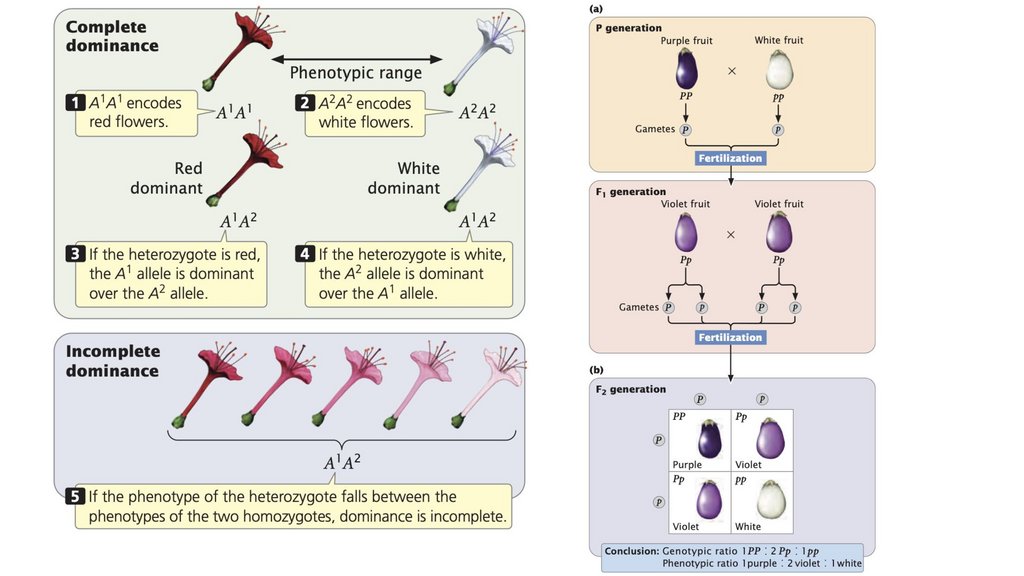
• Доминирование является полным, когда гетерозигота имеет
тот же фенотип, что и гомозигота, неполным, когда
гетерозигота имеет фенотип, промежуточный между
фенотипами двух родительских гомозигот, и кодоминантным,
когда гетерозигота проявляет признаки обеих родительских
гомозигот.
• Тип доминантности не влияет на наследование аллеля; он
влияет на фенотипическую экспрессию аллеля.
Классификация доминантности зависит от уровня
изучаемого фенотипа.
92.
• Пенетрантность - это процент особей с определеннымгенотипом, которые демонстрируют ожидаемый фенотип.
Экспрессивность - это степень выраженности признака.
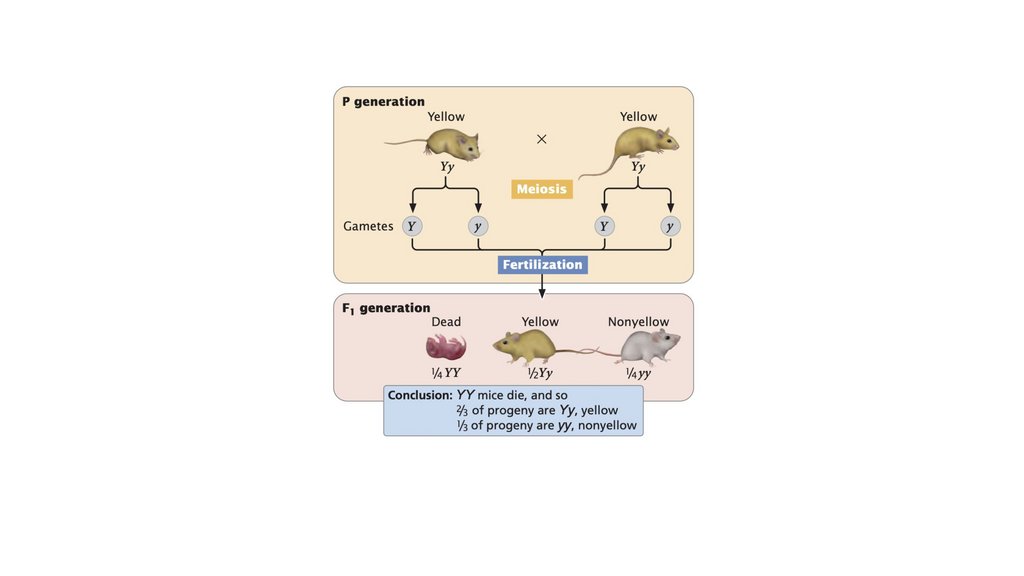
• Летальные аллели вызывают гибель отдельного организма,
обладающего ими, обычно на ранней стадии развития, и
могут изменять фенотипические соотношения.
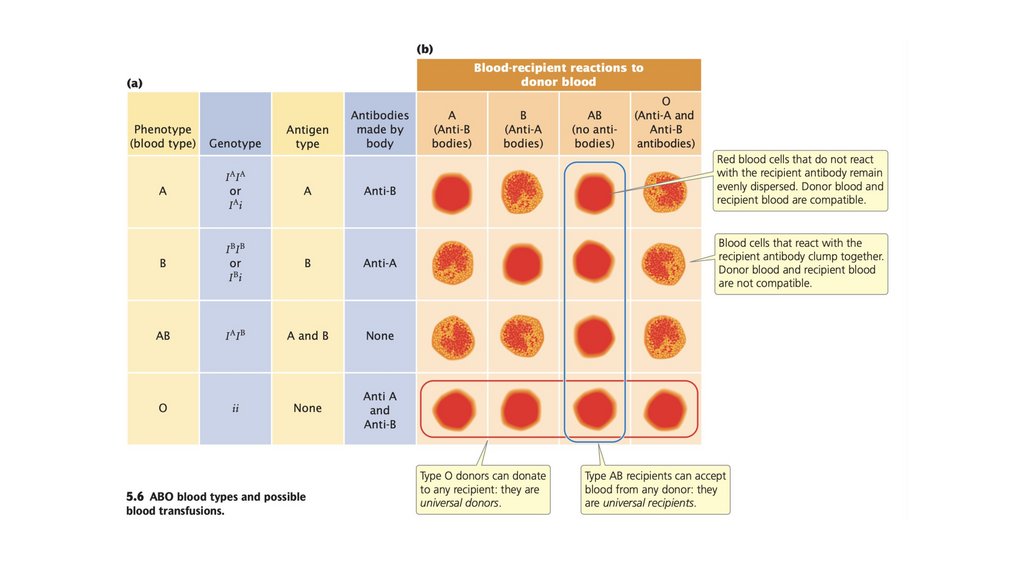
• Множественные аллели относятся к присутствию более двух
аллелей в локусе внутри группы. Их присутствие увеличивает
количество возможных генотипов и фенотипических
разновидностей.
93.
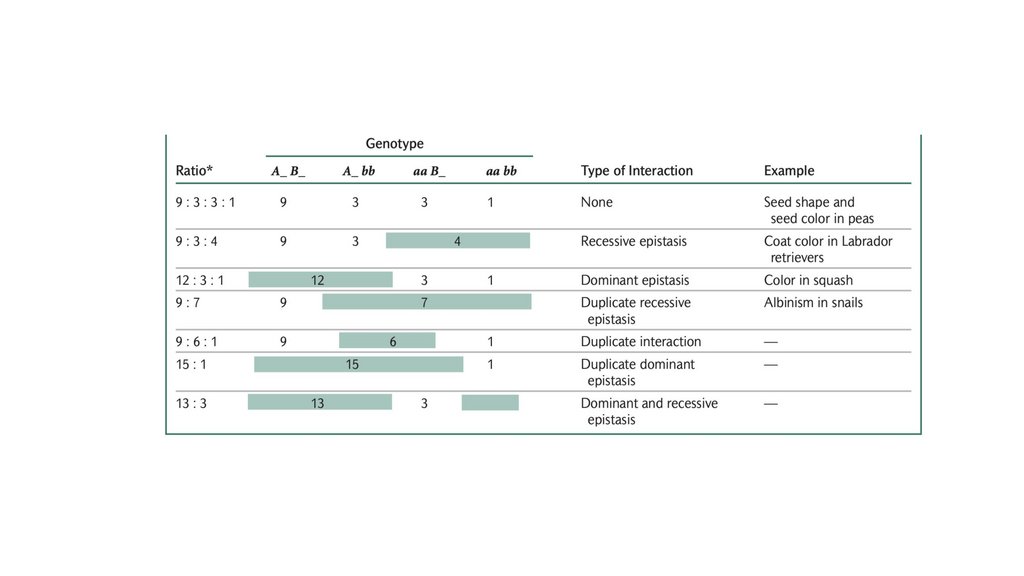
• Взаимодействие генов относится к взаимодействию между генами вразных локусах для получения единого фенотипа. Эпистатический ген в
одном локусе подавляет или маскирует экспрессию гипостатические
гены в других локусах. Взаимодействие генов часто приводит к
фенотипическим соотношениям, которые являются модификациями
дигибридных соотношений.
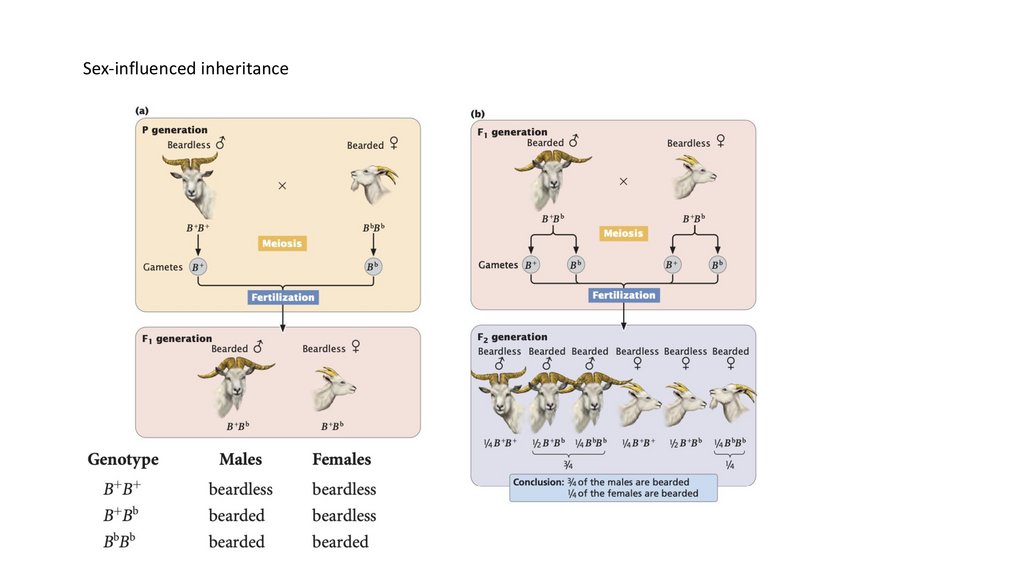
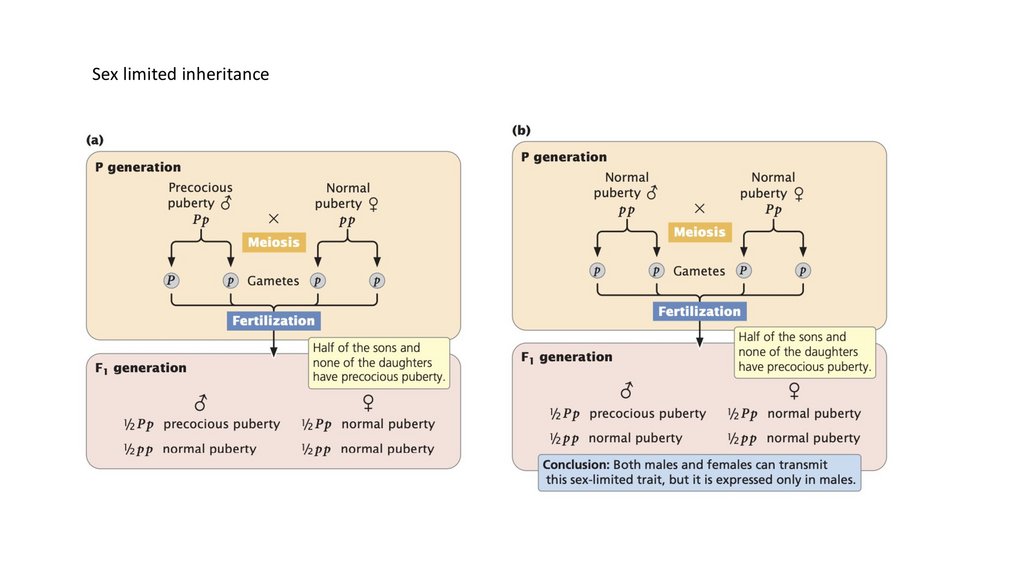
• Характеристики, зависящие от пола, кодируются аутосомными генами,
которые легче экспрессируются у представителей одного пола.
Характеристики, ограниченные полом, кодируются аутосомными
генами, экспрессируемыми только у одного пола.
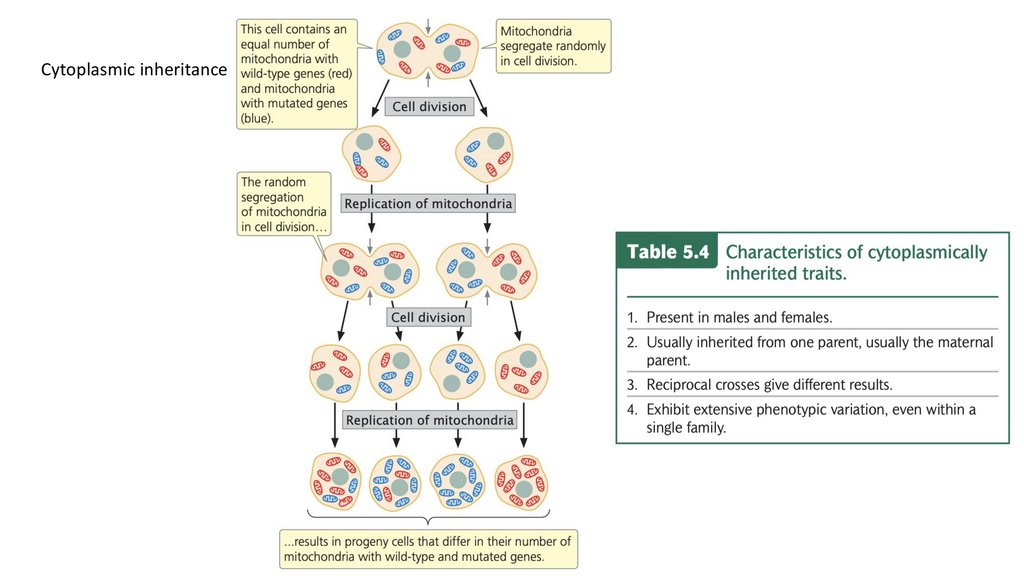
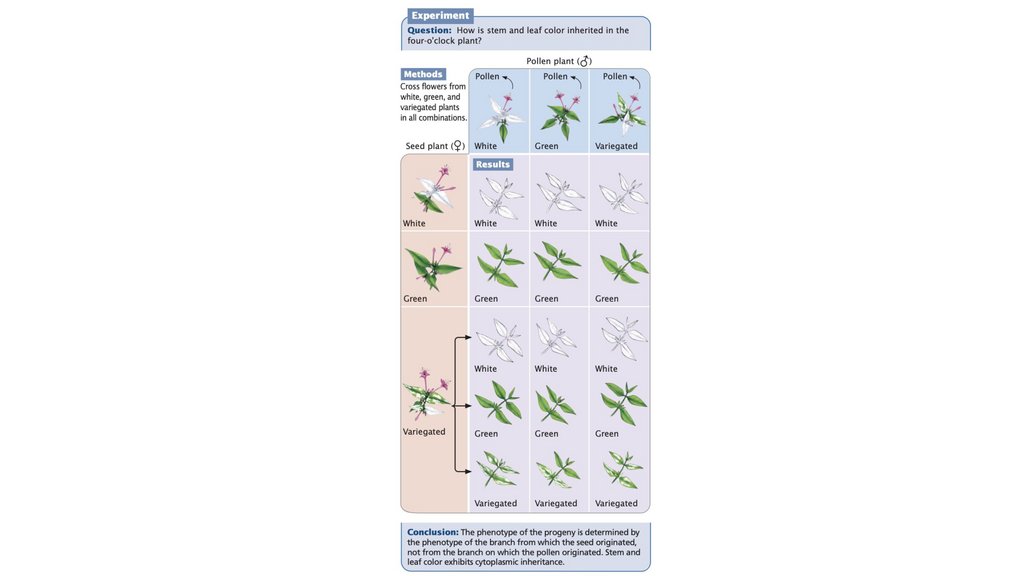
• При цитоплазматическом наследовании гены, отвечающие за данный
признак, находятся в органеллах и обычно наследуются от одного
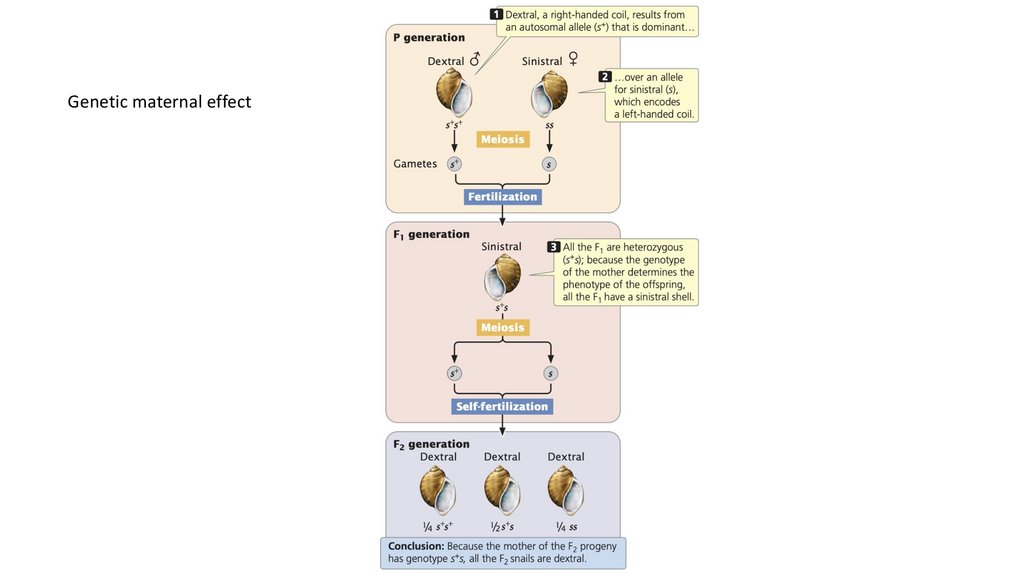
(обычно материнского) родителя. Генетический материнский эффект
присутствует, когда потомство наследует гены от обоих родителей, но
ядерные гены матери определяют фенотип потомства.
94.
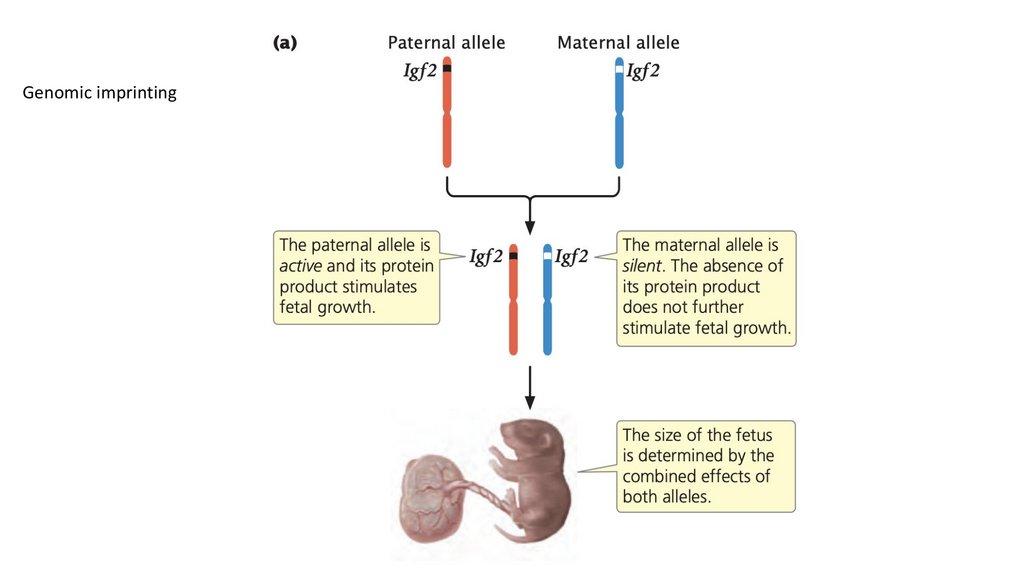
• Геномный импринтинг относится к характеристикам, кодируемымаутосомными генами, на экспрессию которых влияет пол
родителя, передающего гены.
• Предвосхищение относится к генетическому признаку, который
более сильно выражен или проявляется в более раннем возрасте
в последующих поколениях.
• Фенотипы часто изменяются под воздействием окружающей
среды. Фенокопия - это фенотип, созданный воздействием
окружающей среды, который имитирует фенотип, созданный
генотипом.
• Непрерывные характеристики - это те, которые проявляют
широкий спектр фенотипов; они часто возникают в результате
комбинированного воздействия многих генов и воздействия
окружающей среды.
95.
96.
97.
98.
99.
• Пенетрантность определяется как процент отдельныхорганизмов, имеющих определенный генотип, которые выражают
ожидаемый фенотип.
• Иногда люди обладают аллелем полидактилии (о чем
свидетельствует тот факт, что их дети наследуют полидактилию),
но, тем не менее, имеют нормальное количество пальцев на
руках и ногах
• Родственным понятием является экспрессивность, степень, в
которой выражен характер. В дополнение к неполной
проникновенности, полидактилия демонстрирует переменную
экспрессивность. Некоторые люди с полидактилией обладают
дополнительными пальцами рук и ног, которые полностью
функционируют, в то время как другие обладают лишь
небольшим участком дополнительной кожи.
• Неполная пенетрантность и переменная экспрессия обусловлены
воздействием других генов и факторов окружающей среды,
которые могут изменять или полностью подавлять действие
определенного гена.
100.
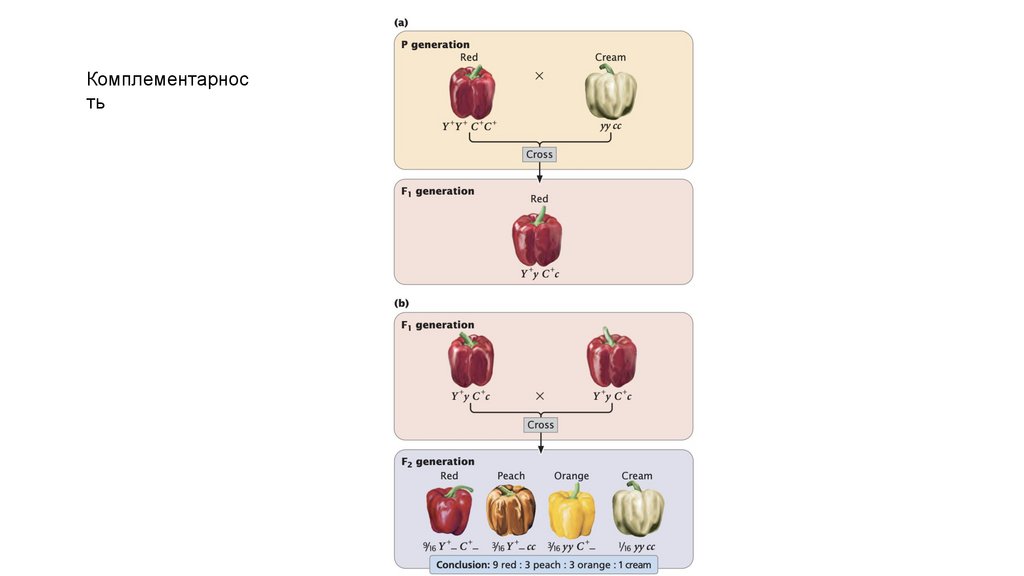
Комплементарность
101.
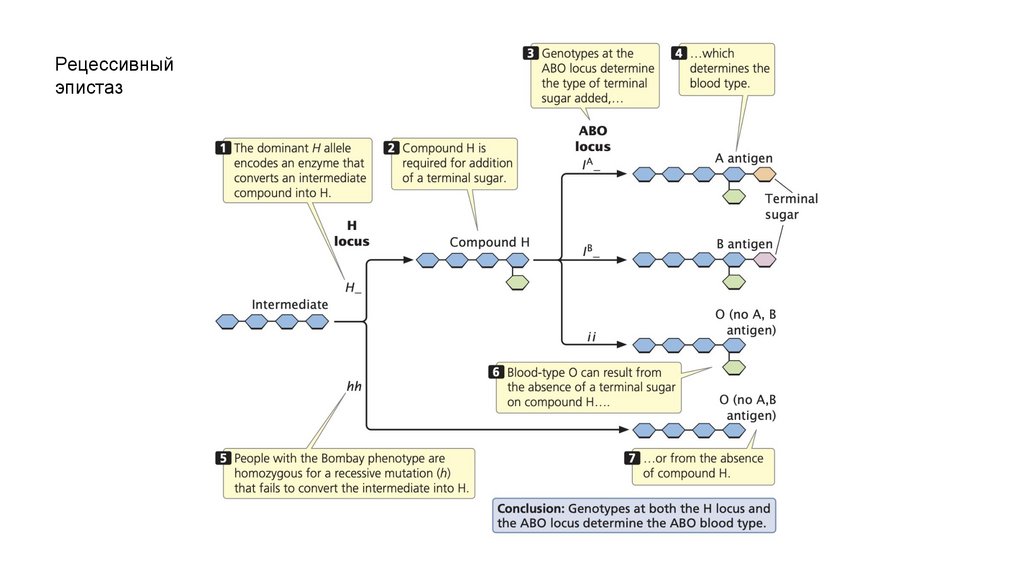
Рецессивныйэпистаз
102.
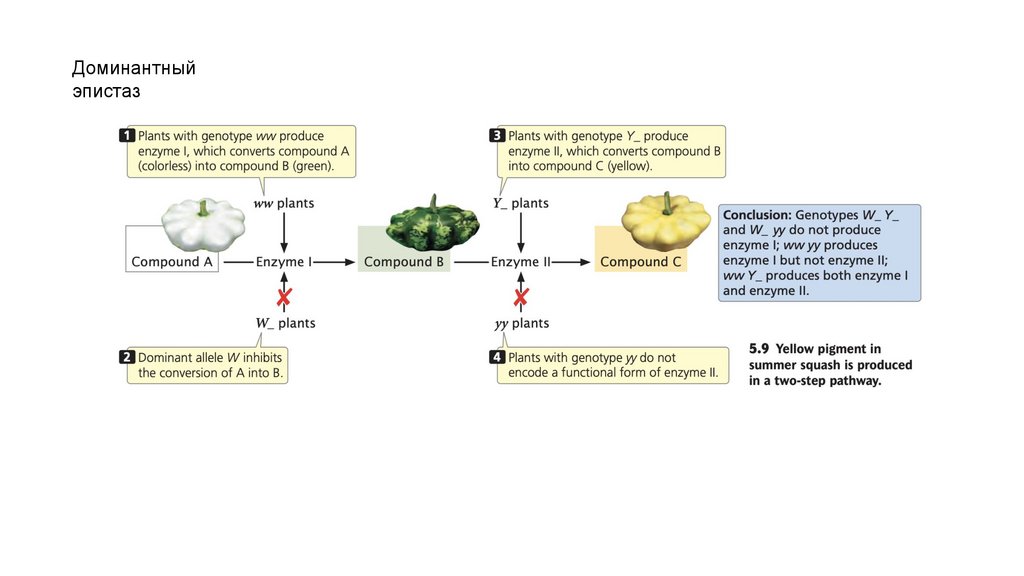
Доминантныйэпистаз
103.
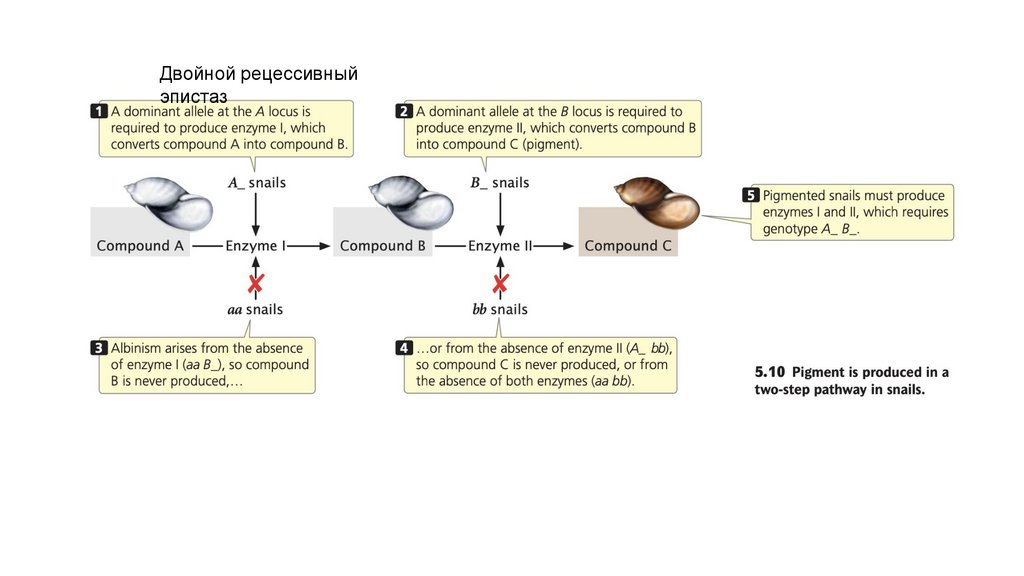
Двойной рецессивныйэпистаз
104.
105.
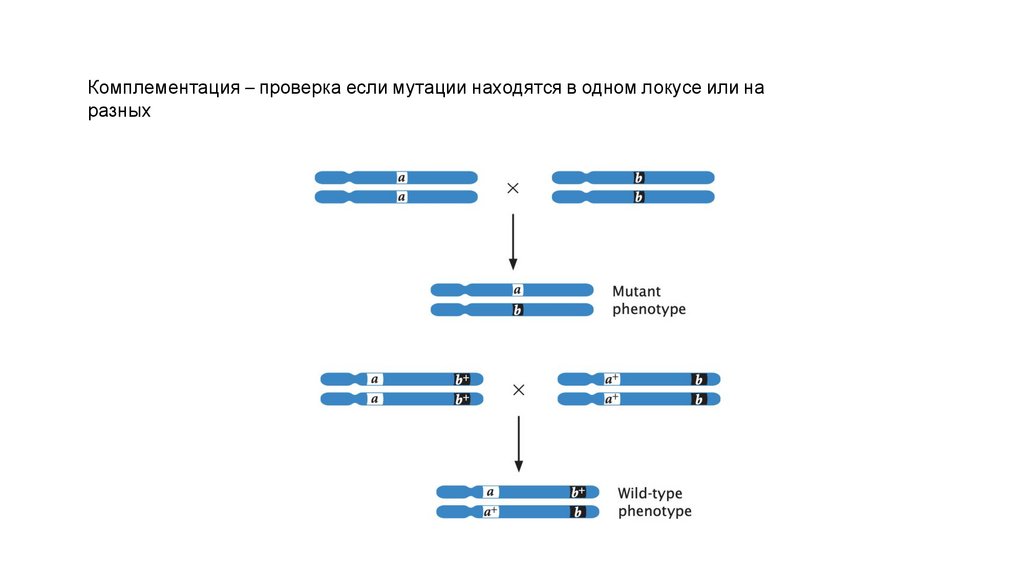
Комплементация – проверка если мутации находятся в одном локусе или наразных
106.
Sex-influenced inheritance107.
Sex limited inheritance108.
Cytoplasmic inheritance109.
110.
Genetic maternal effect111.
Genomic imprinting112.
113.
114.
• Ограничения на генетическое изучение человеческих черт включают в себяневозможность проведения контролируемых скрещиваний, длительное время
генерации, небольшой размер семьи и сложность разделения генетических
влияний и влияния окружающей среды. Родословные часто используются для
изучения наследования признаков у людей.
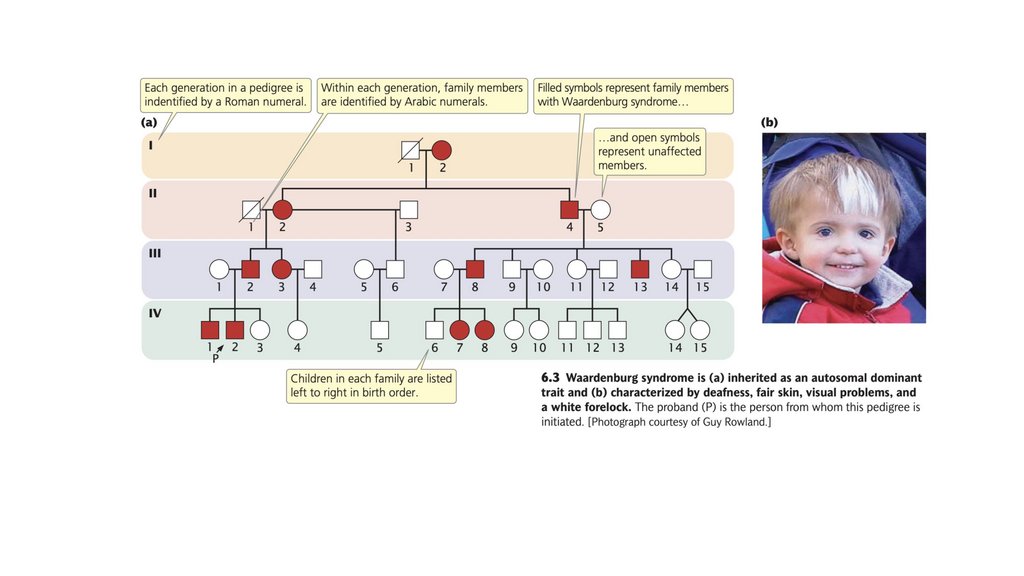
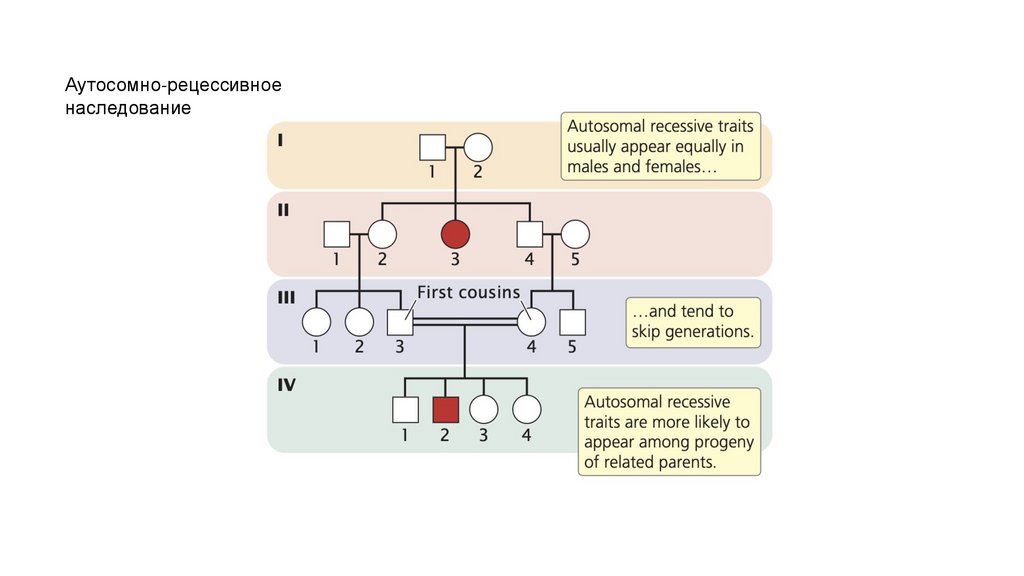
• Аутосомно-рецессивные признаки обычно проявляются с одинаковой частотой
у обоих полов и, как правило, передаются через поколения. Когда оба
родителя гетерозиготны по определенному аутосомно-рецессивному признаку,
примерно четверть их потомства будет обладать этим признаком.
Рецессивные признаки чаще проявляются в семьях с кровным родством
(спаривание между близкородственными лицами).
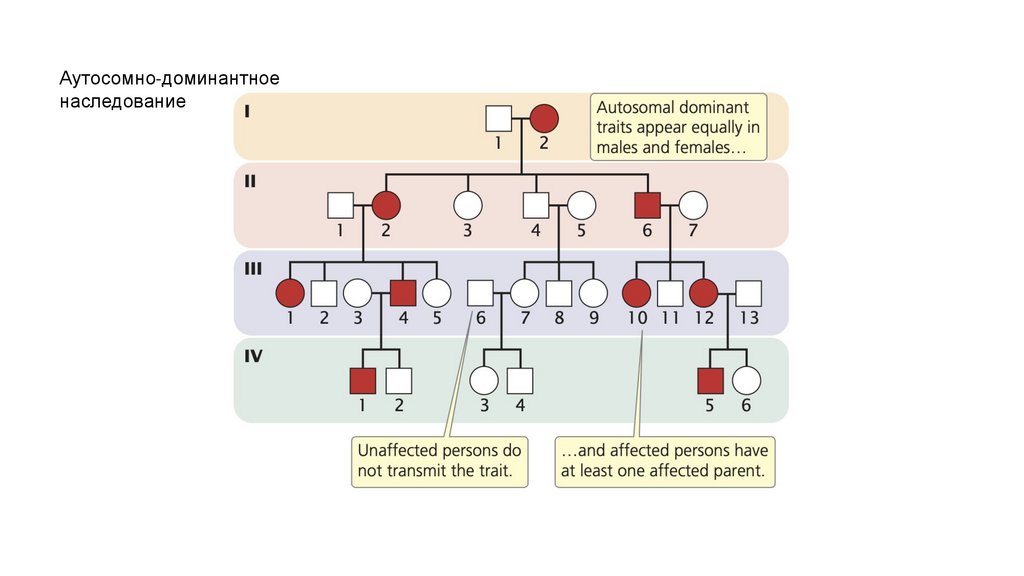
• Аутосомно-доминантные признаки обычно проявляются в равной степени у
обоих полов и не переходят из поколения в поколение. Когда один из
родителей поражен и гетерозиготен по аутосомно-доминантному признаку,
примерно половина потомства будет обладать этим признаком. Незатронутые
люди обычно не передают аутосомно-доминантный признак своему потомству.
115.
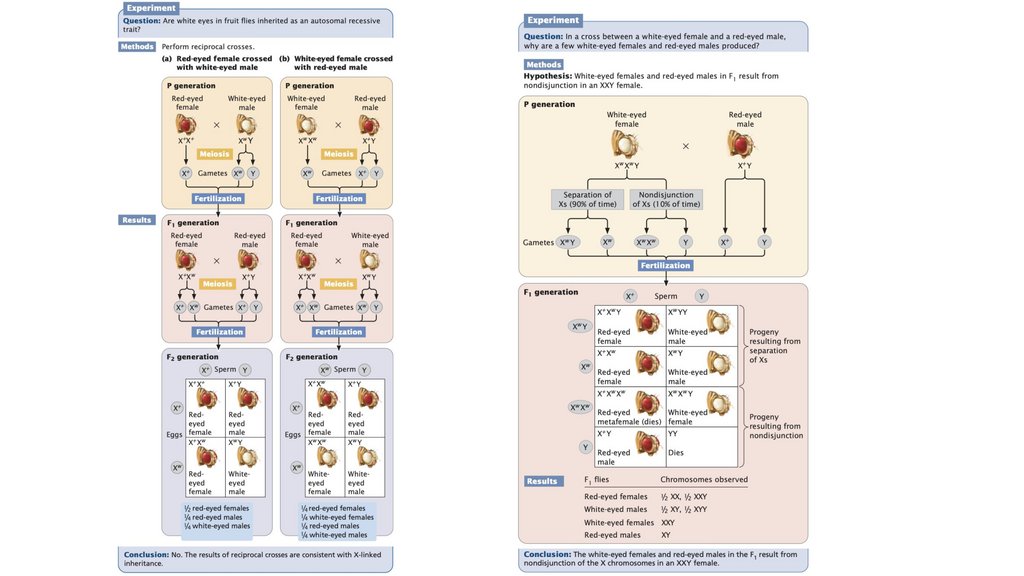
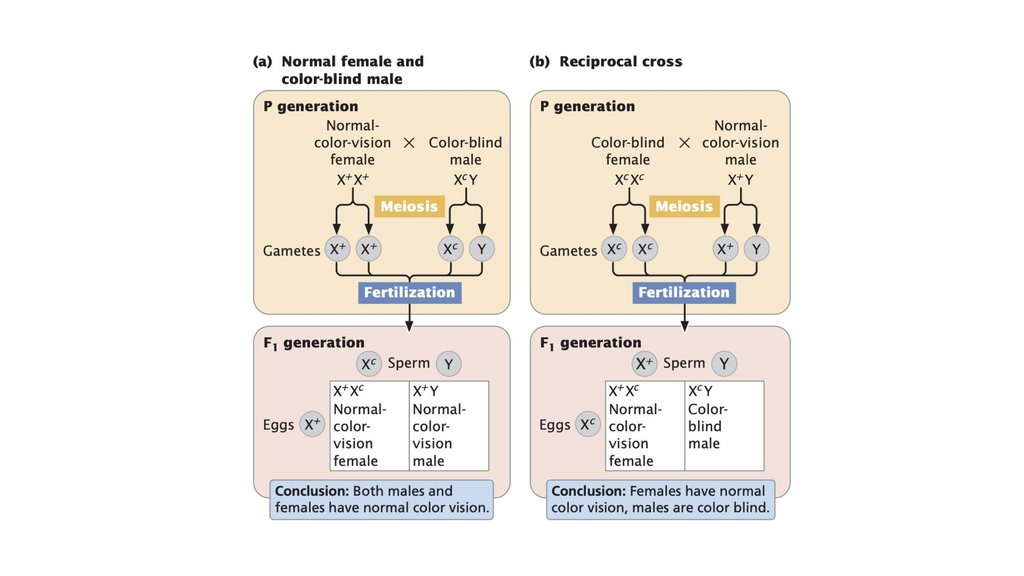
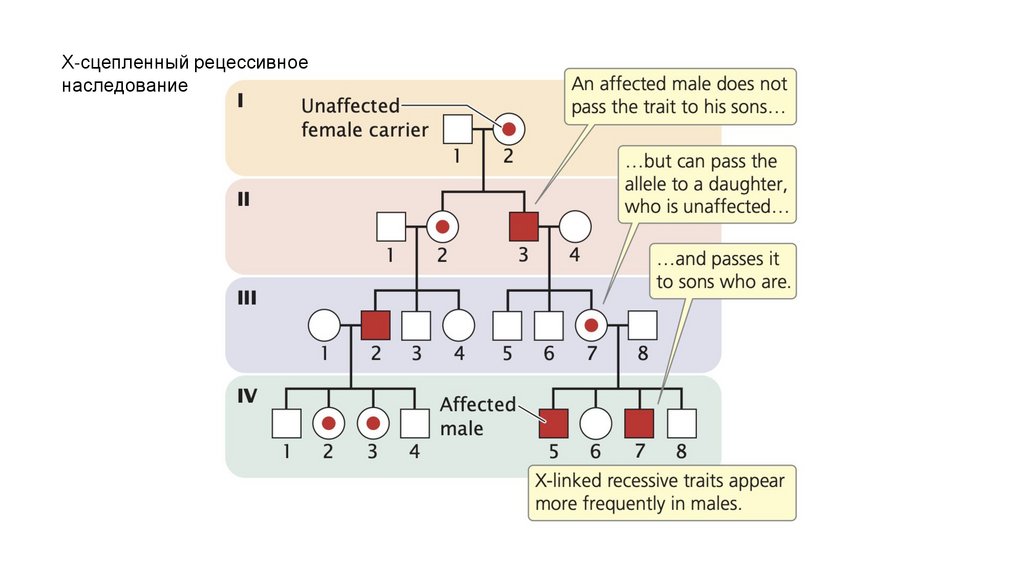
• Х-сцепленные рецессивные признаки чаще проявляются умужчин, чем у женщин. Когда женщина является гетерозиготным
носителем Х-сцепленного рецессивного признака, а мужчина не
подвержен этому заболеванию, примерно половина их сыновей
будет обладать этим признаком, а половина их дочерей будут
незатронутыми носителями. Х-сцепленные признаки не
передаются от отца к сыну.
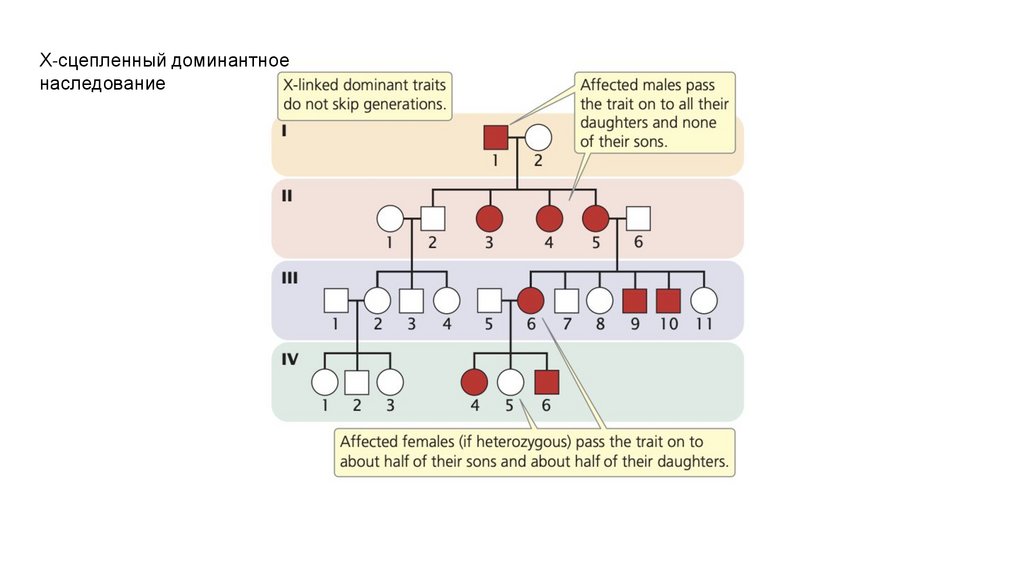
• Х-сцепленные доминантные признаки проявляются у мужчин и
женщин, но чаще у самок. Они не переходят из поколения в
поколение. Пораженные мужчины передают Х-сцепленную
доминантную черту всем своим дочерям, но ни одному из своих
сыновей. Гетерозиготные женщины передают этот признак
половине своих сыновей и половине дочерей.
• Y-сцепленные признаки проявляются только у мужчин и
передаются от отца всем сыновьям.
116.
• Более высокое соответствие признака у монозиготных близнецов, чем у дизиготных, указывает нагенетическое влияние на признак; менее 100% соответствие у монозиготных близнецов указывает на влияние
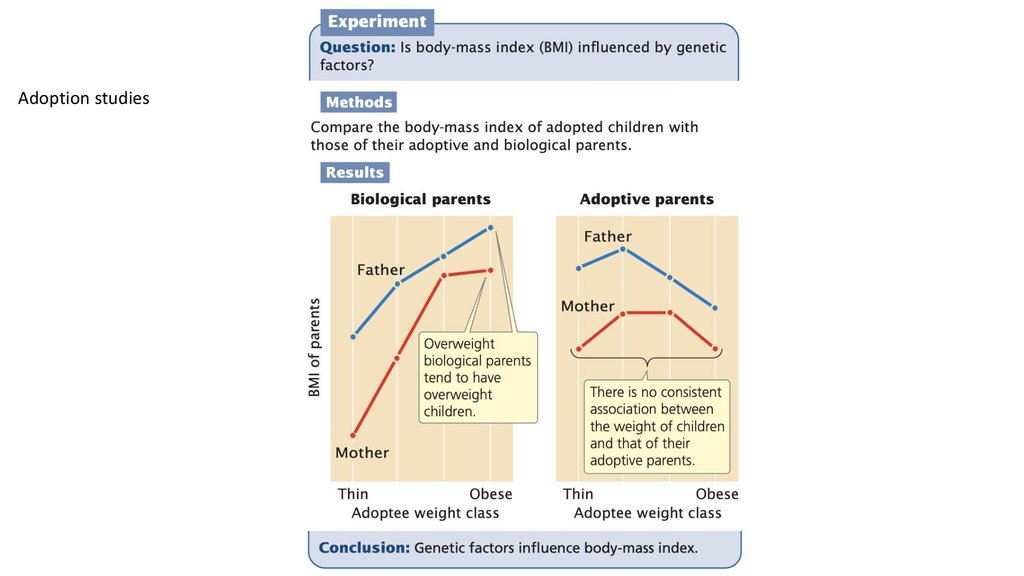
окружающей среды на признак.Сходство между усыновленными детьми и их биологическими родителями
указывает на важность генетических факторов в проявлении того или иного признака; сходство между
усыновленными детьми и их генетически неродственными приемными родителями указывает на влияние
факторов окружающей среды.
• Генетическое консультирование предоставляет информацию и поддержку людям, обеспокоенным
наследственными заболеваниями в их семьях.Генетическое тестирование включает пренатальную
диагностику, скрининг на наличие болезнетворных аллелей у новорожденных, выявление людей,
гетерозиготных по рецессивным аллелям, и досимптомное тестирование на наличие болезнетворного аллеля у
людей из группы риска.Интерпретация генетических тестов может быть осложнена наличием многочисленных
причинных мутаций, неполной пенетрантностью и влиянием факторов окружающей среды.
• Недавняя доступность генетических тестов непосредственно для потребителя вызвала обеспокоенность по
поводу адекватности предоставляемой информации, отсутствия генетических консультаций, точности,
конфиденциальности и практического использования некоторых тестов.
• Генетическое тестирование вызвало обеспокоенность по поводу генетической дискриминации и
конфиденциальности результатов тестов. Закон о недискриминации генетической информации запрещает
использование генетической информации при принятии решения о возможности медицинского страхования и
трудоустройства.
• Генетические исследования выявили ряд генов, которые могут способствовать уникальности человека,
включая гены, влияющие на размер мозга и речь.
117.
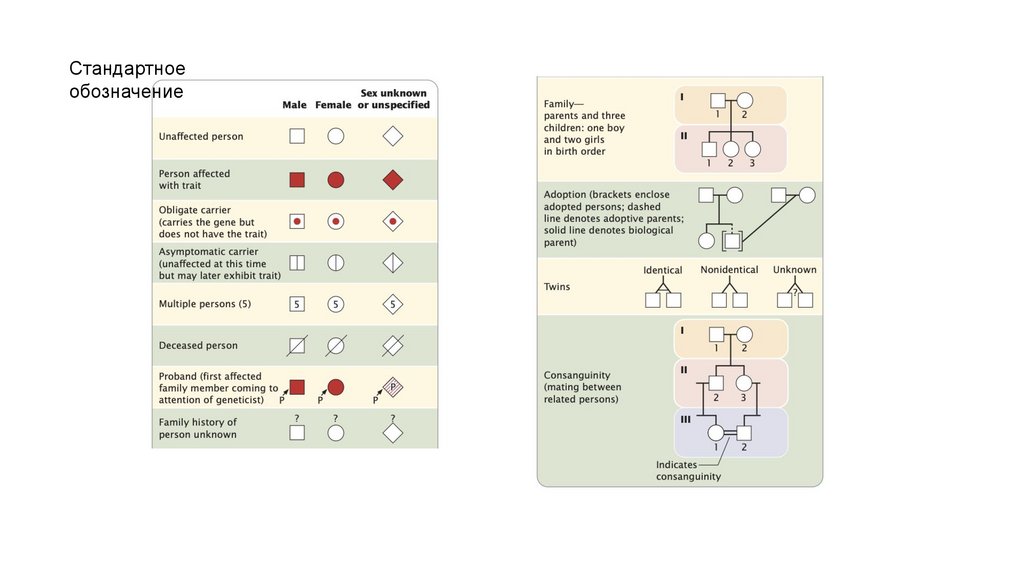
Стандартноеобозначение
118.
119.
Аутосомно-рецессивноенаследование
120.
Аутосомно-доминантноенаследование
121.
Х-сцепленный рецессивноенаследование
122.
Х-сцепленный доминантноенаследование
123.
Y-сцепленноенаследование
124.
125.
Monozygotic versus Dizygotic twins126.
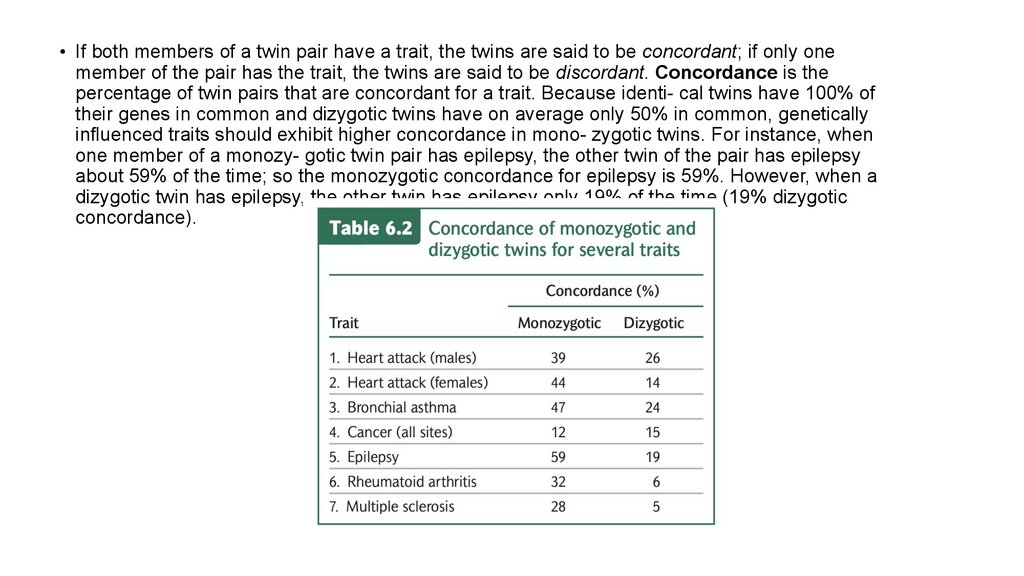
• If both members of a twin pair have a trait, the twins are said to be concordant; if only onemember of the pair has the trait, the twins are said to be discordant. Concordance is the
percentage of twin pairs that are concordant for a trait. Because identi- cal twins have 100% of
their genes in common and dizygotic twins have on average only 50% in common, genetically
influenced traits should exhibit higher concordance in mono- zygotic twins. For instance, when
one member of a monozy- gotic twin pair has epilepsy, the other twin of the pair has epilepsy
about 59% of the time; so the monozygotic concordance for epilepsy is 59%. However, when a
dizygotic twin has epilepsy, the other twin has epilepsy only 19% of the time (19% dizygotic
concordance).
127.
Adoption studies128.
Ultrasounography129.
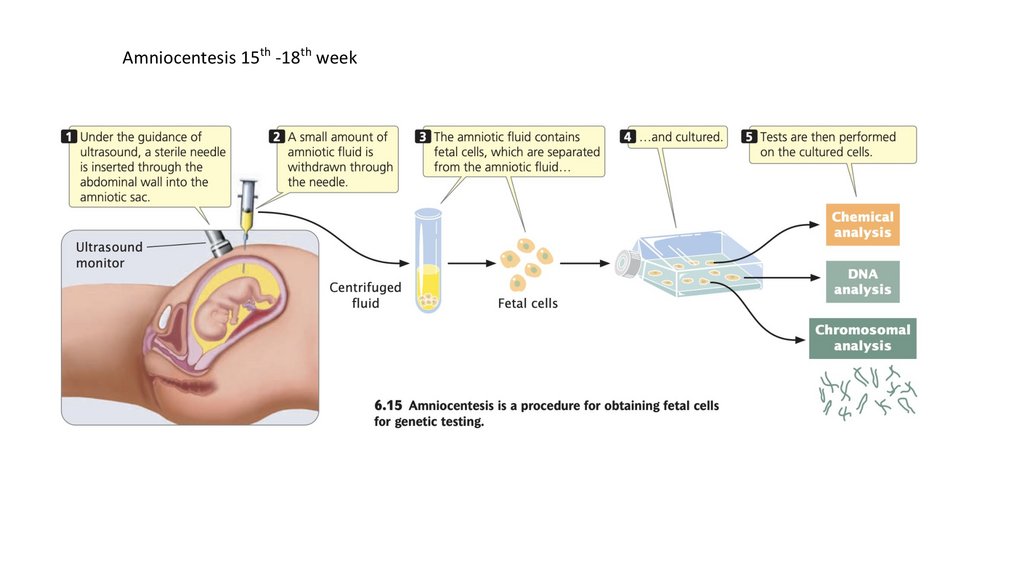
Amniocentesis 15th -18th week130.
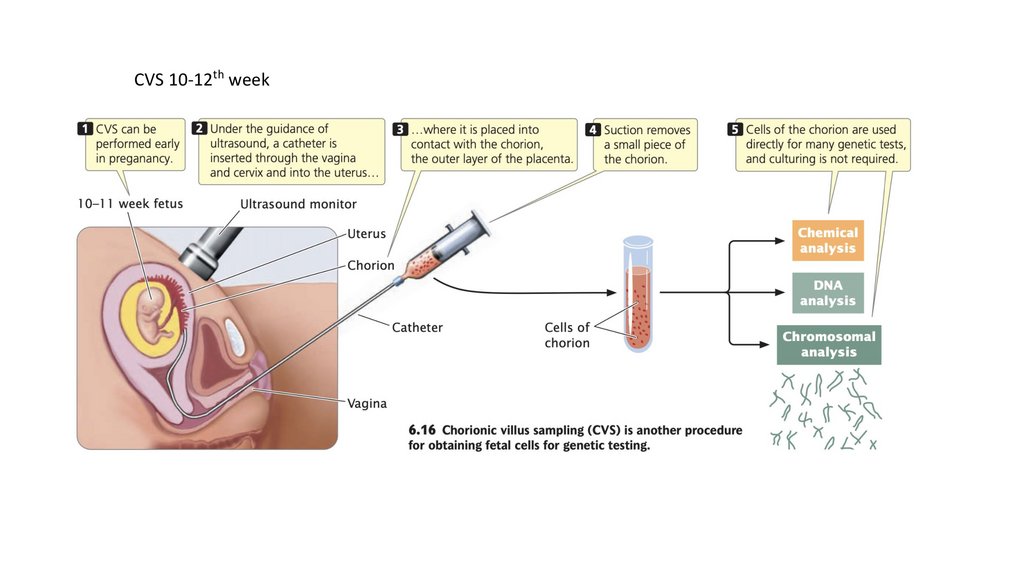
CVS 10-12th week131.
132.
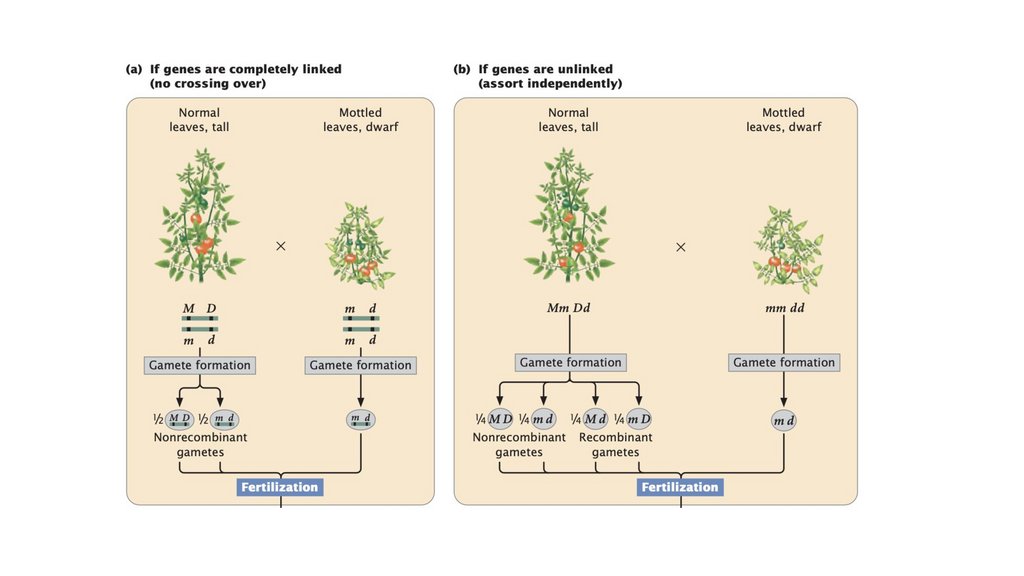
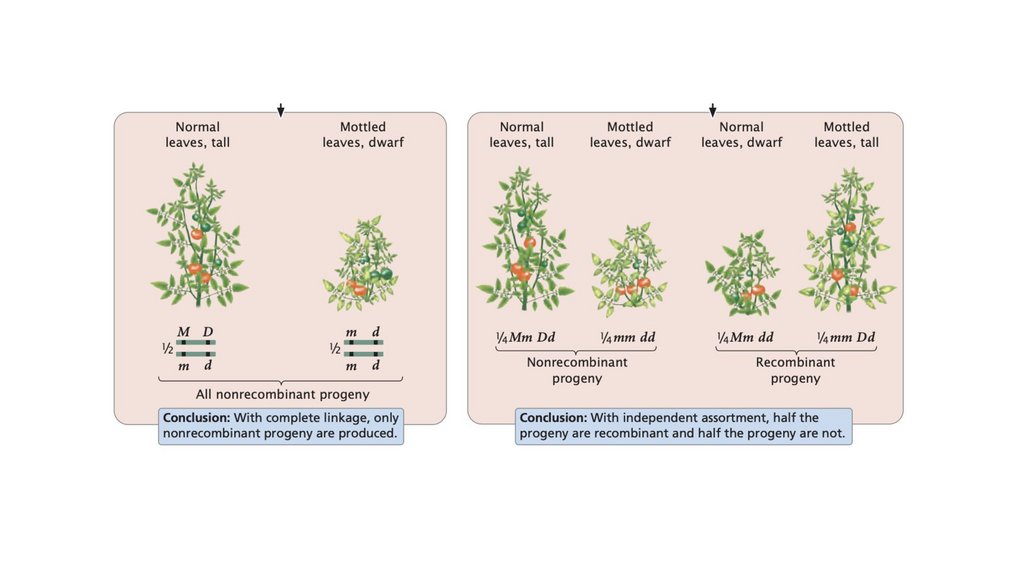
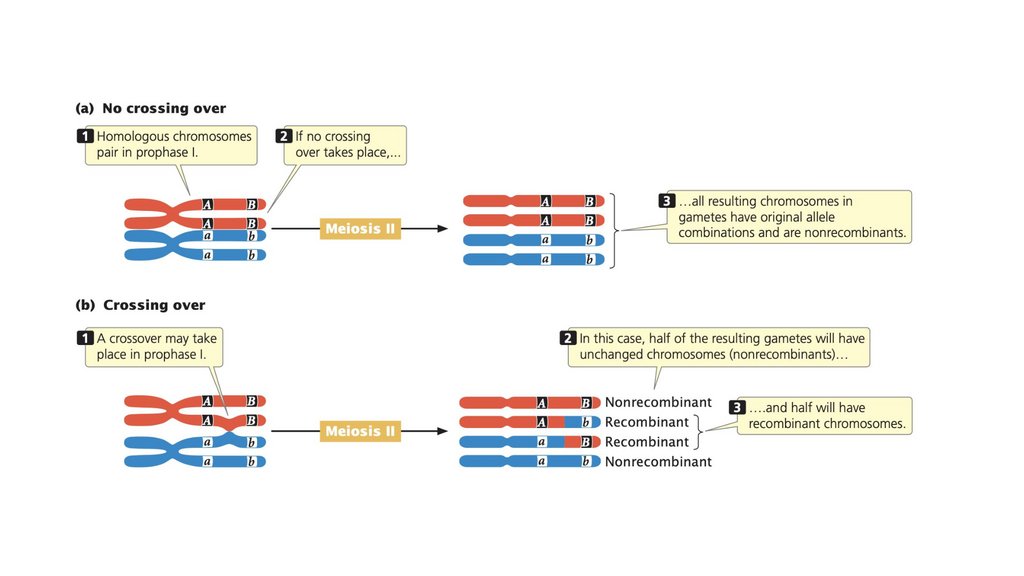
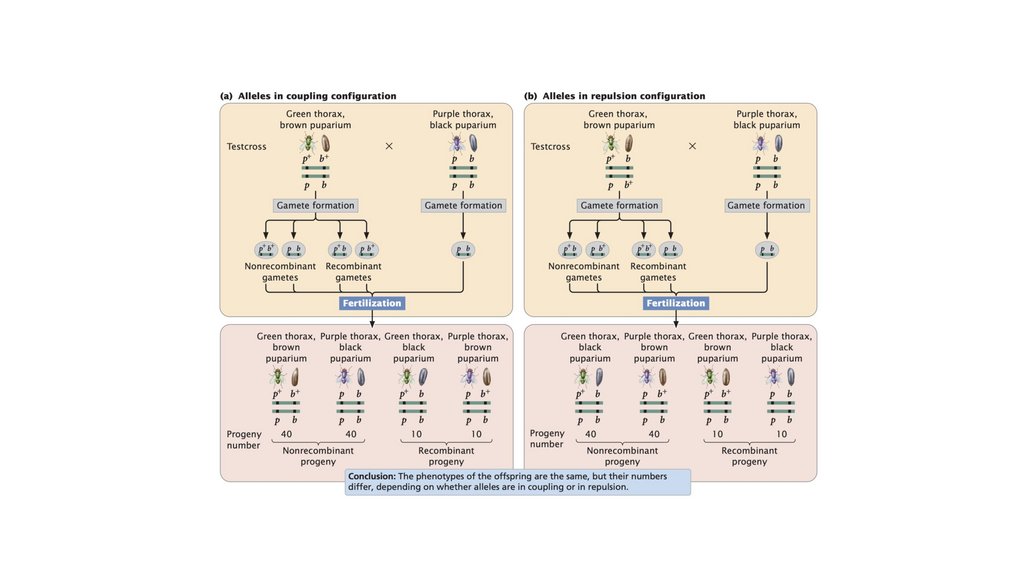
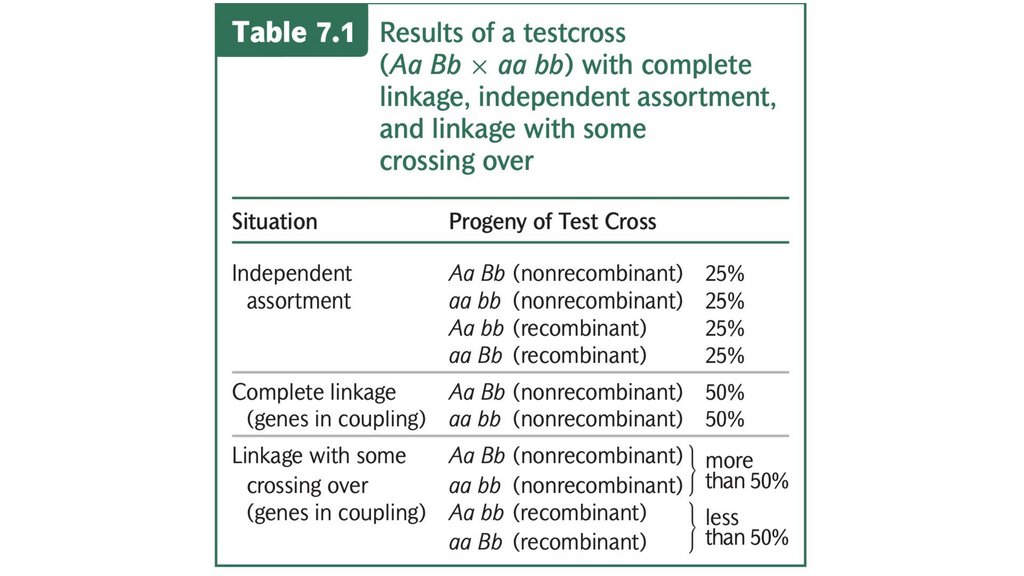
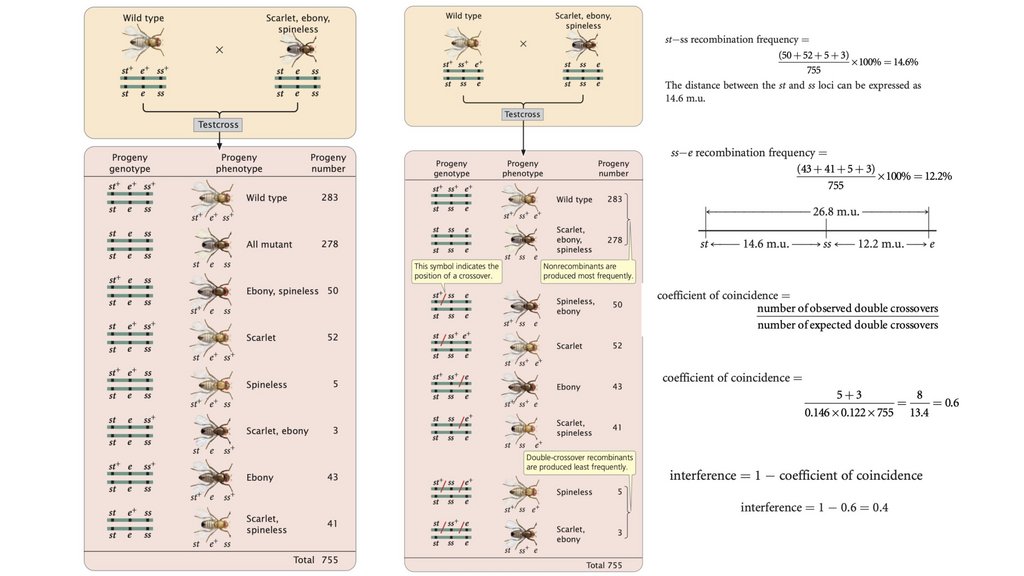
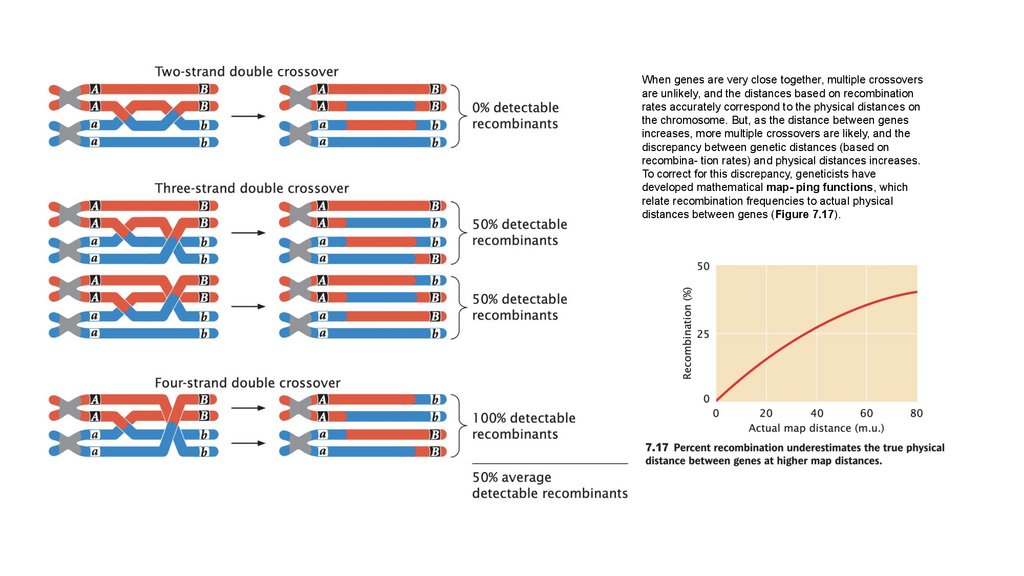
• Сцепленные гены не сортируются независимо. При тестовомскрещивании двух полностью сцепленных генов (без кроссинговера)
образуется только нерекомбинантное потомство. Когда два гена
сортируются независимо, рекомбинантное потомство и
нерекомбинантное потомство образуются в равных пропорциях. Когда
два гена связаны некоторым кроссинговером между ними, образуется
больше нерекомбинантного потомства, чем рекомбинантного
потомства.
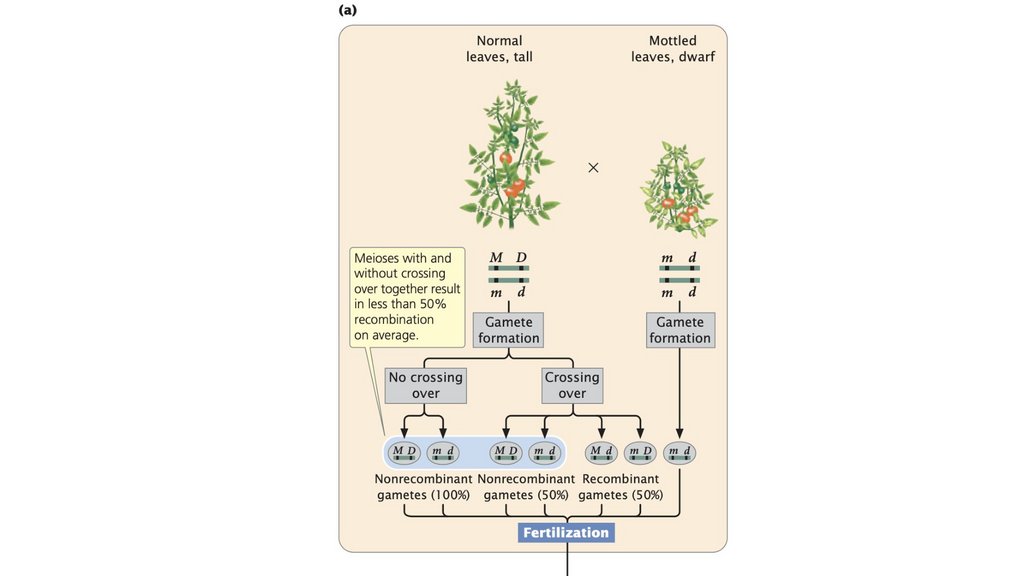
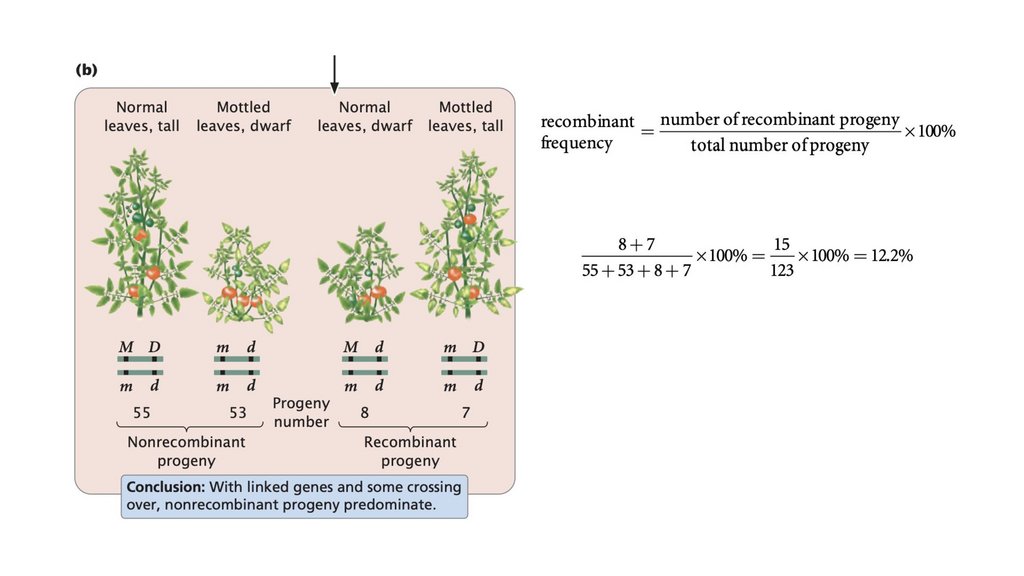
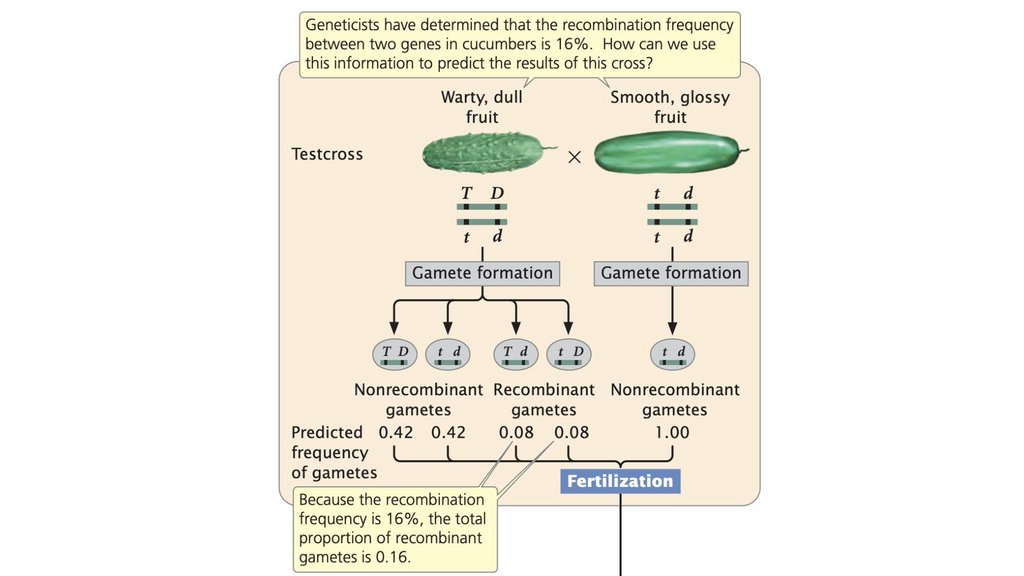
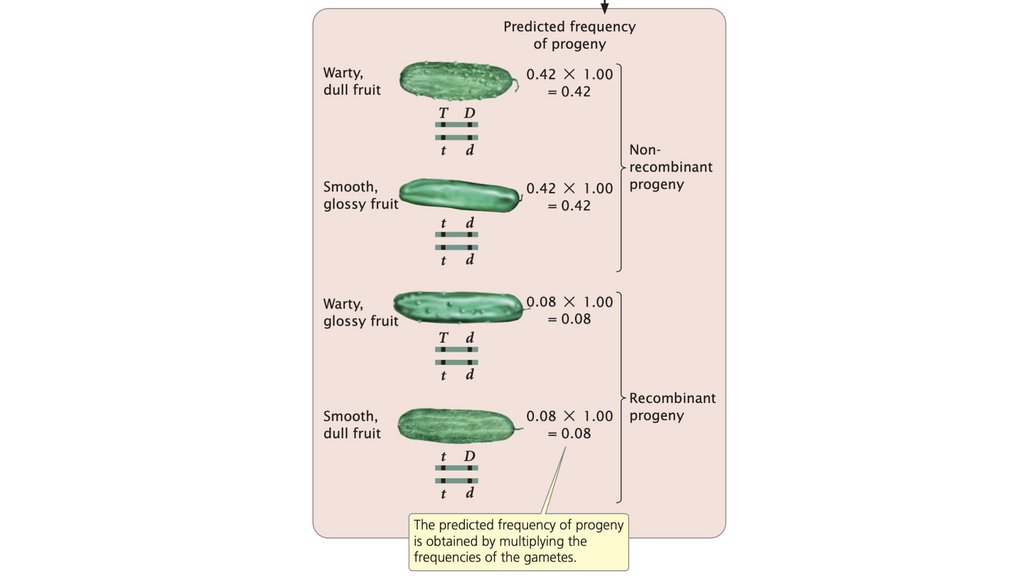
• Частота рекомбинации рассчитывается путем суммирования
количества рекомбинантного потомства, деления на общее количество
потомства, полученного в результате скрещивания, и умножения на
100%. Частота рекомбинации составляет половину частоты
кроссинговера, а максимальная частота рекомбинантных гамет
составляет 50%.
• Сцепление и отталкивание относятся к расположению аллелей на
хромосоме. То, находятся ли гены в конфигурации сцепления или
отталкивания, определяет, какая комбинация фенотипов будет
наиболее частой в потомстве тест-скрещивания.
133.
• Межхромосомная рекомбинация происходит между генами,расположенными на разных хромосомах, путем случайной
сегрегации хромосом в мейозе. Внутрихромосомная
рекомбинация происходит между генами, расположенными
на одной хромосоме, путем кроссинговера.
• Критерий независимости хи-квадрат может быть
использован для определения того, связаны ли гены.
• Скорость рекомбинации может использоваться для
определения относительного порядка генов и расстояний
между ними в хромосоме. Один процент рекомбинации
равен одной единице карты. Карты, основанные на частоте
рекомбинации, называются генетическими картами; карты,
основанные на физических расстояниях, называются
физическими картами.
134.
• Генетические карты могут быть составлены путем изученияскорости рекомбинации в серии двухточечных скрещиваний
или путем изучения потомства в результате трехточечного
тест-скрещивания.
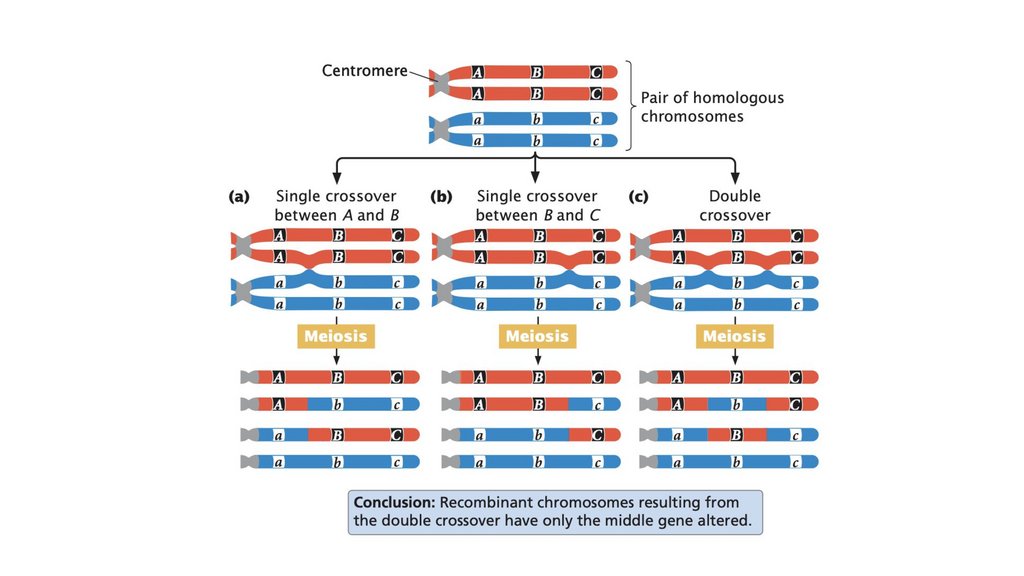
• Некоторые множественные кроссинговеры остаются
незамеченными; таким образом, генетические карты,
основанные на частоте рекомбинации, недооценивают
истинные физические расстояния между генами.
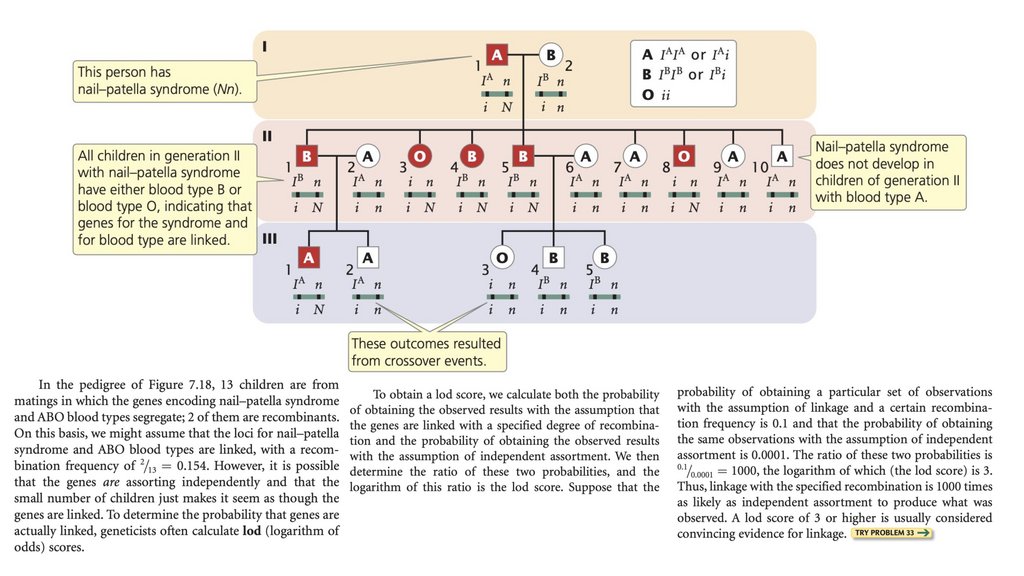
• Гены человека можно картировать, изучая косегрегацию
признаков в родословных.
135.
• Оценка lod - это логарифм отношения вероятностиполучения наблюдаемого потомства при предположении о
связи к вероятности получения наблюдаемого потомства при
предположении о независимом ассортименте. Оценка lod,
равная 3 или выше, обычно считается доказательством
связи.
• Молекулярные методы, позволяющие обнаруживать
вариабельные различия в последовательности ДНК,
значительно облегчили картирование генов.
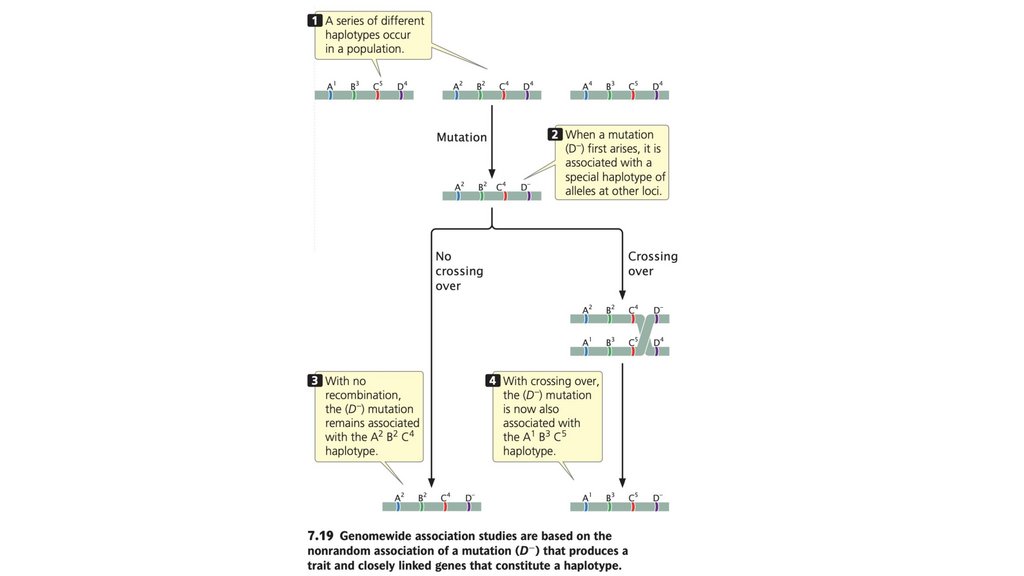
• Общегеномные ассоциативные исследования выявляют
гены, влияющие на определенные признаки, путем изучения
неслучайной ассоциации признака с генетическими
маркерами по всему геному.
136.
• Секвенирование нуклеотидов - еще один метод физическогокартирования генов.
• Скорости рекомбинации широко варьируются, различаясь у
разных видов, между хромосомами и вдоль них в пределах
одного вида и даже между самцами и самками одного и того
же вида.
137.
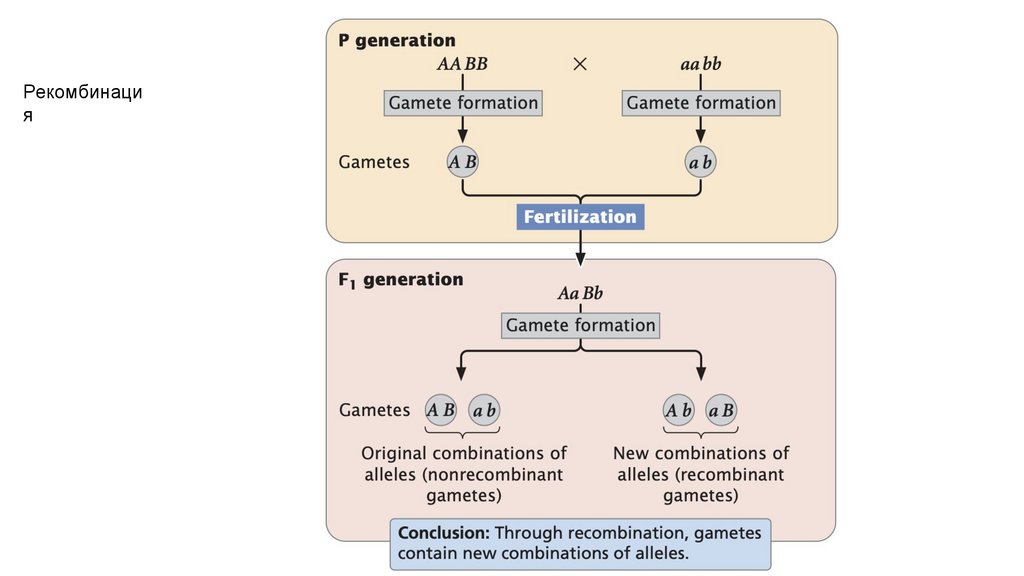
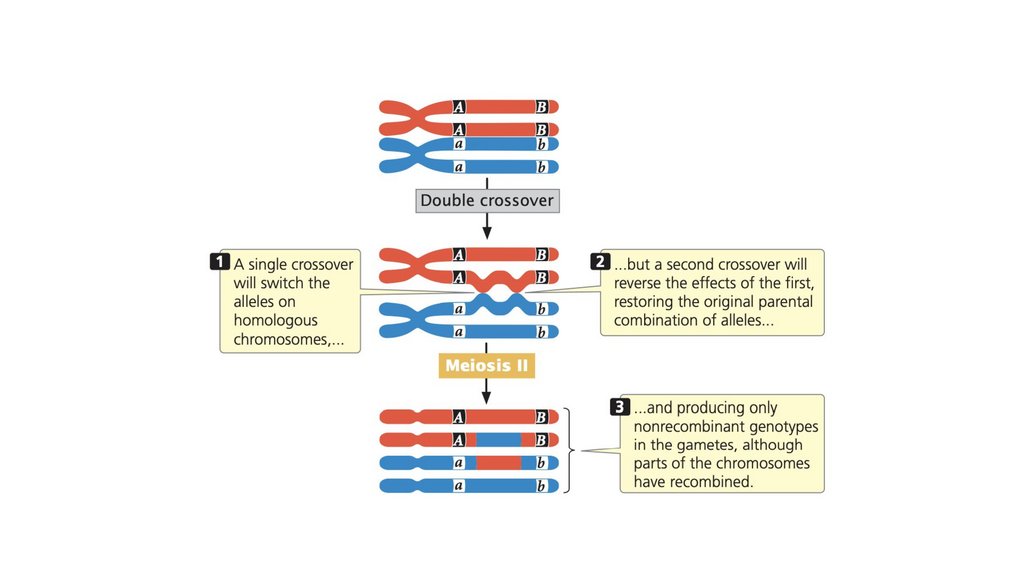
Рекомбинация
138.
139.
Кроссинговер
140.
141.
142.
143.
144.
145.
146.
147.
148.
149.
150.
151.
152.
153.
When genes are very close together, multiple crossoversare unlikely, and the distances based on recombination
rates accurately correspond to the physical distances on
the chromosome. But, as the distance between genes
increases, more multiple crossovers are likely, and the
discrepancy between genetic distances (based on
recombina- tion rates) and physical distances increases.
To correct for this discrepancy, geneticists have
developed mathematical map- ping functions, which
relate recombination frequencies to actual physical
distances between genes (Figure 7.17).
154.
155.
156.
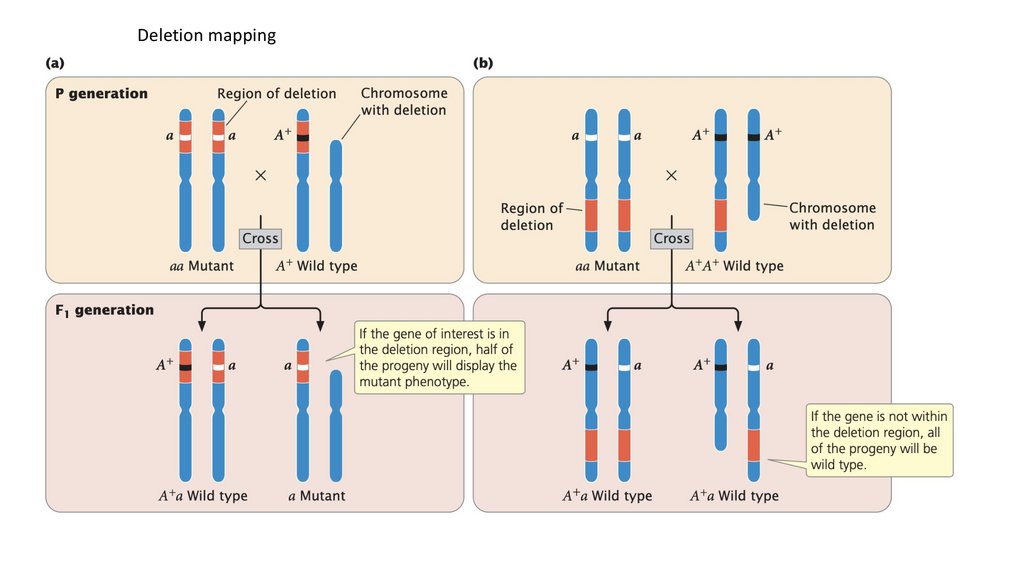
Deletion mapping157.
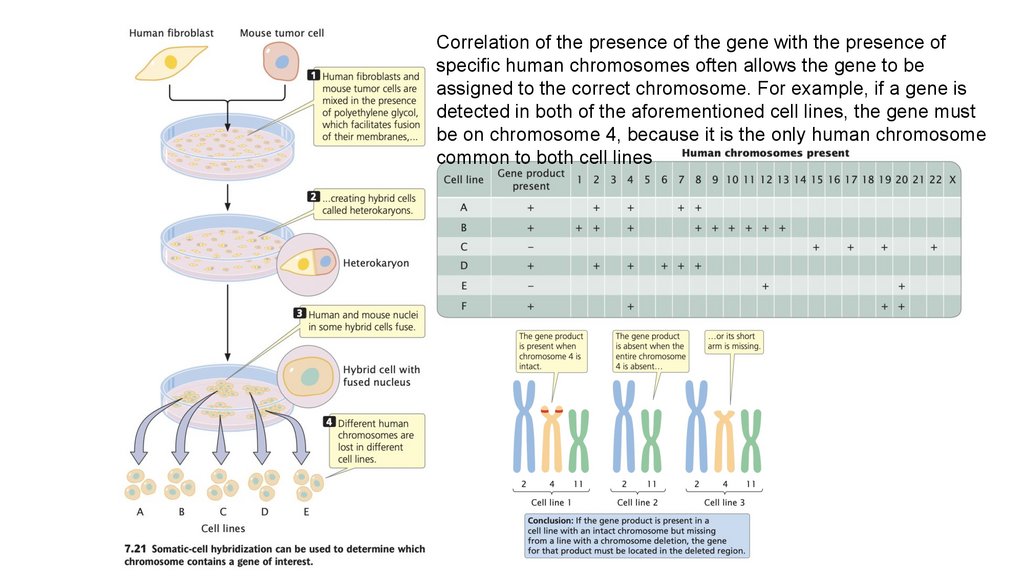
Correlation of the presence of the gene with the presence ofspecific human chromosomes often allows the gene to be
assigned to the correct chromosome. For example, if a gene is
detected in both of the aforementioned cell lines, the gene must
be on chromosome 4, because it is the only human chromosome
common to both cell lines