










 Биология
БиологияПохожие презентации:








")

Физиология ЦНС: ствол мозга, мозжечок, промежуточный мозг
1.
ПЛАН ЛЕКЦИИ:1. Продолговатый мозг, его функции и центры.
2. Проводниковая функция продолговатого мозга.
3. Роль продолговатого мозга в регуляции мышечного тонуса.
4. Ретикулярная формация ствола мозга. Восходящие активирующие
влияния на кору больших полушарий.
5. Средний мозг. Функции четверохолмия, красных ядер, черного вещества,
ядер III и IY пар ЧМН.
6. Участие среднего мозга в осуществлении фазно-тонической деятельности
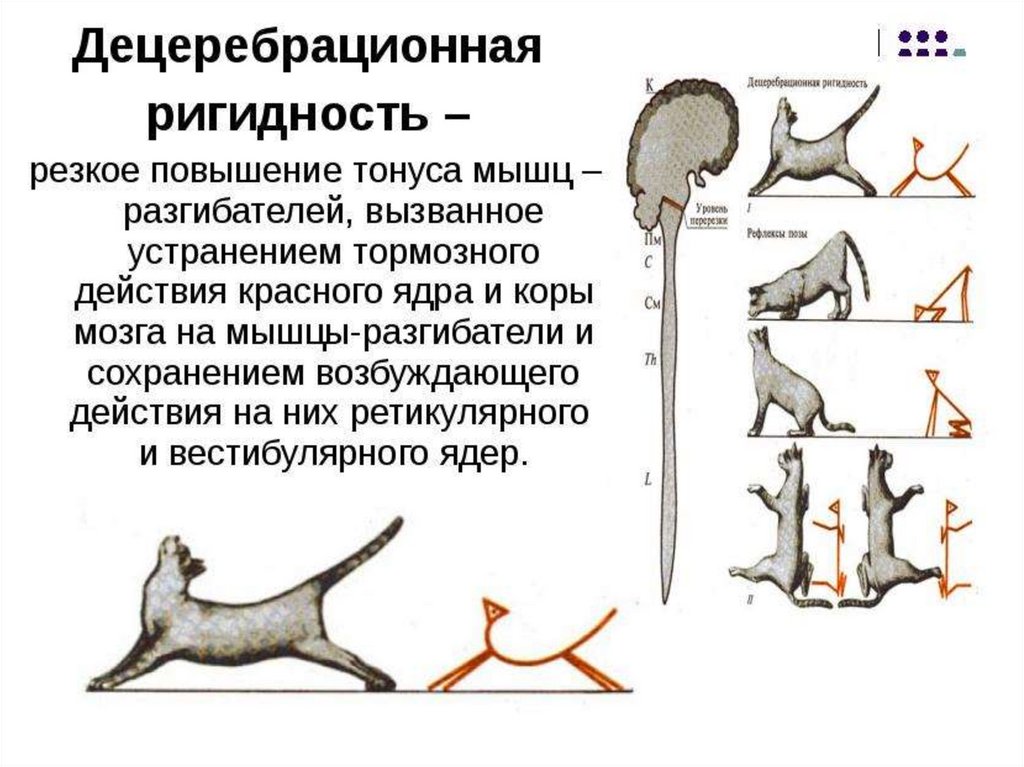
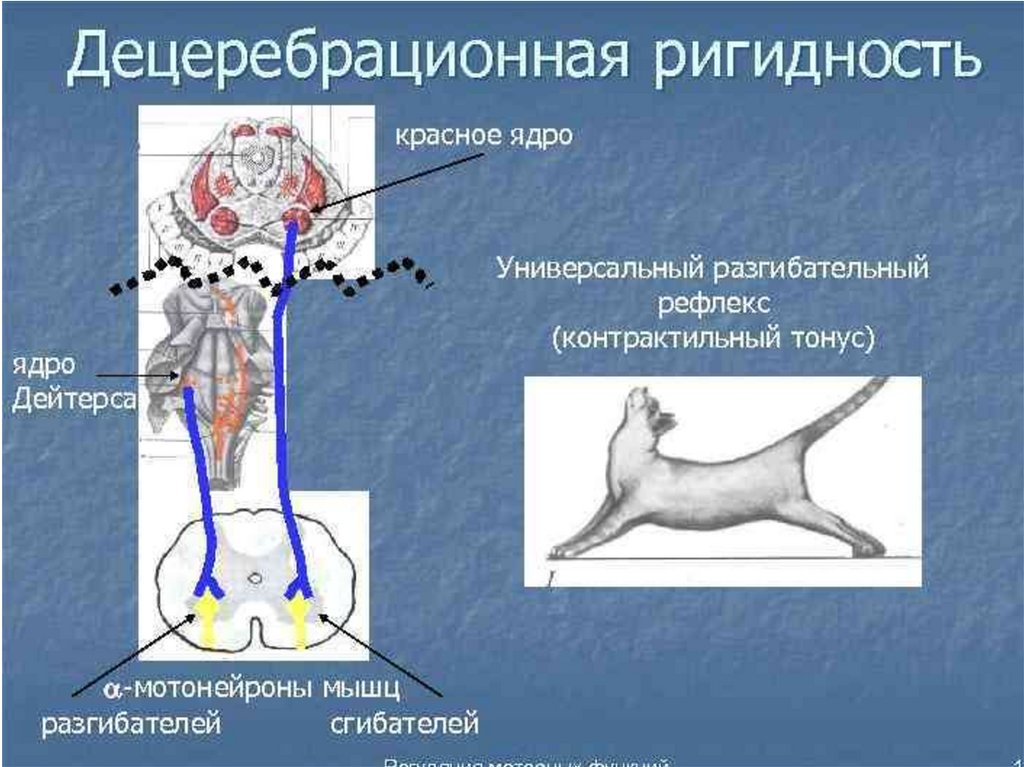
мышц. Установочные рефлексы. Децеребрационная ригидность.
7. Мозжечок и его функции.
8. Функции гипоталамус как высшего центра регуляции вегетативных
реакций.
9. Таламус. Функциональная организация ядер.
2.
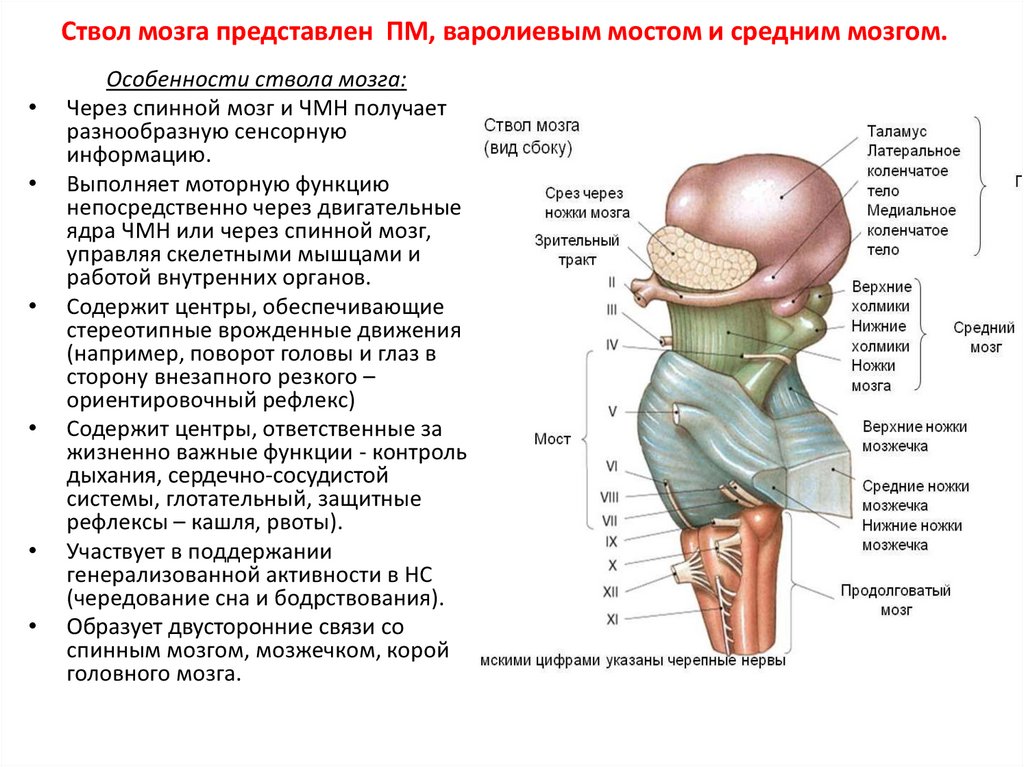
Ствол мозга представлен ПМ, варолиевым мостом и средним мозгом.Особенности ствола мозга:
Через спинной мозг и ЧМН получает
разнообразную сенсорную
информацию.
Выполняет моторную функцию
непосредственно через двигательные
ядра ЧМН или через спинной мозг,
управляя скелетными мышцами и
работой внутренних органов.
Содержит центры, обеспечивающие
стереотипные врожденные движения
(например, поворот головы и глаз в
сторону внезапного резкого –
ориентировочный рефлекс)
Содержит центры, ответственные за
жизненно важные функции - контроль
дыхания, сердечно-сосудистой
системы, глотательный, защитные
рефлексы – кашля, рвоты).
Участвует в поддержании
генерализованной активности в НС
(чередование сна и бодрствования).
Образует двусторонние связи со
спинным мозгом, мозжечком, корой
головного мозга.
3.
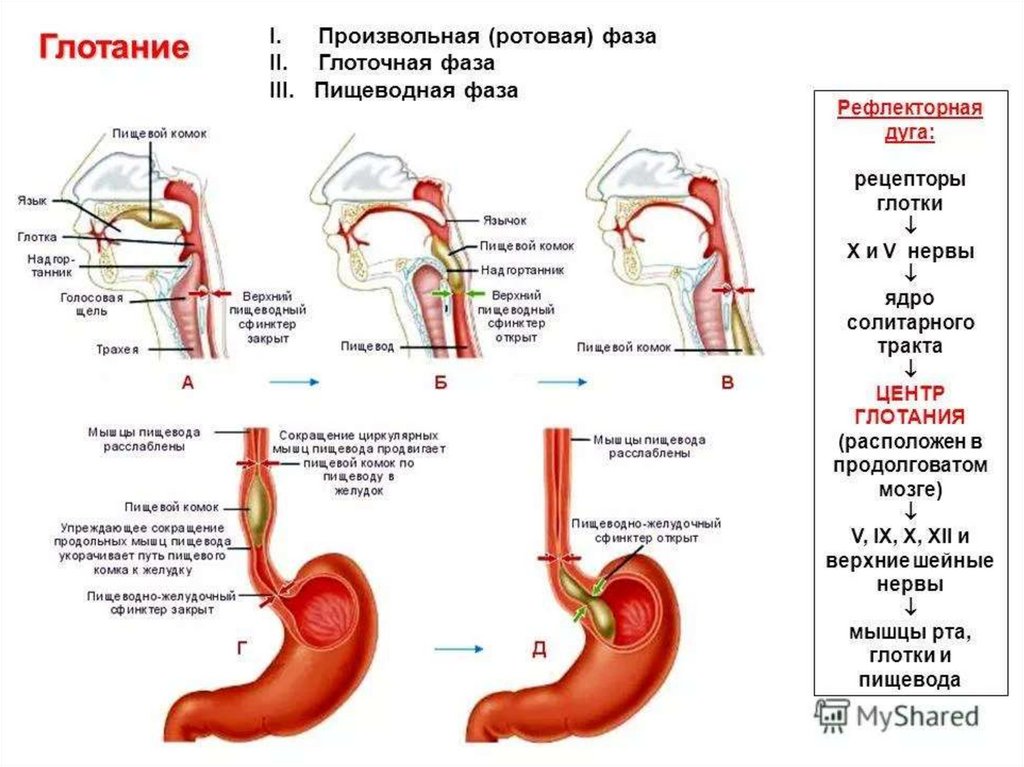
Продолговатый мозг, его функции и центры.• В продолговатом мозге расположены ядра следующих черепномозговых нервов:
• - пара IX —языкоглоточный нерв; его ядро образовано 3 частями —
двигательной, чувствительной и вегетативной. Двигательная часть
участвует в иннервации мышц глотки и полости рта, чувствительная —
получает информацию от рецепторов вкуса задней трети языка;
вегетативная - иннервирует слюнные железы (нижнее
слюноотделительное ядро - парасимпатическое).
• - пара X — блуждающий нерв (главный нерв парасимпатической НС)
имеет 3 ядра: вегетативное иннервирует гортань, пищевод, сердце,
желудок, кишечник, пищеварительные железы; чувствительное получает
информацию от рецепторов альвеол легких и других внутренних органов
и двигательное обеспечивает последовательность сокращения мышц
глотки, гортани при глотании;
!!! Языкоглоточный и блуждающий нерв имеют общее двигательное
(двойное), общее чувствительное (ядро одиночного (солитарного)
тракта).
• - пара XI — добавочный нерв; его двигательные волокна совместно с
спинномозговыми корешками иннервирует трапециевидную и грудиноключично-сосцевидную мышцы.
• - пара XII — подъязычный нерв иннервирует мышцы языка, участвует в
актах жевания, глотания
4.
Продолговатый мозг организует иреализует ряд защитных рефлексов:
рвоты,
чиханья,
кашля,
слезоотделения, смыкания век. Эти
рефлексы реализуются благодаря
тому, что информация о раздражении
рецепторов слизистой оболочки глаза,
полости рта, гортани, носоглотки
через
чувствительные
ветви
тройничного и языкоглоточного
нервов
попадает
в
ядра
продолговатого мозга. Отсюда идет
команда к двигательным ядрам
тройничного,
блуждающего,
лицевого,
языкоглоточного,
добавочного
или
подъязычного
нервов, в результате реализуется тот
или иной защитный рефлекс. Точно
так же за счет последовательного
включения мышечных групп головы,
шеи, грудной клетки и диафрагмы
организуются рефлексы пищевого
поведения:
сосания,
жевания,
глотания.
5.
Продолговатый мозг регулирует ряд сенсорных функций:• рецепцию кожной
чувствительности лица — в
сенсорном ядре тройничного
нерва;
• первичный анализ рецепции вкуса
— в ядре языкоглоточного нерва;
• рецепцию слуховых раздражений
— в ядре улиткового нерва;
• рецепцию вестибулярных
раздражений — в верхнем
вестибулярном ядре.
• В задневерхних отделах
продолговатого мозга проходят
пути кожной, глубокой,
висцеральной чувствительности,
часть из которых переключается
здесь на второй нейрон (тонкое и
клиновидное ядра).
На уровне продолговатого
мозга перечисленные
сенсорные функции
реализуют первичный анализ
силы и качества раздражения,
далее обработанная
информация передается в
подкорковые структуры для
определения биологической
значимости данного
раздражения.
6.
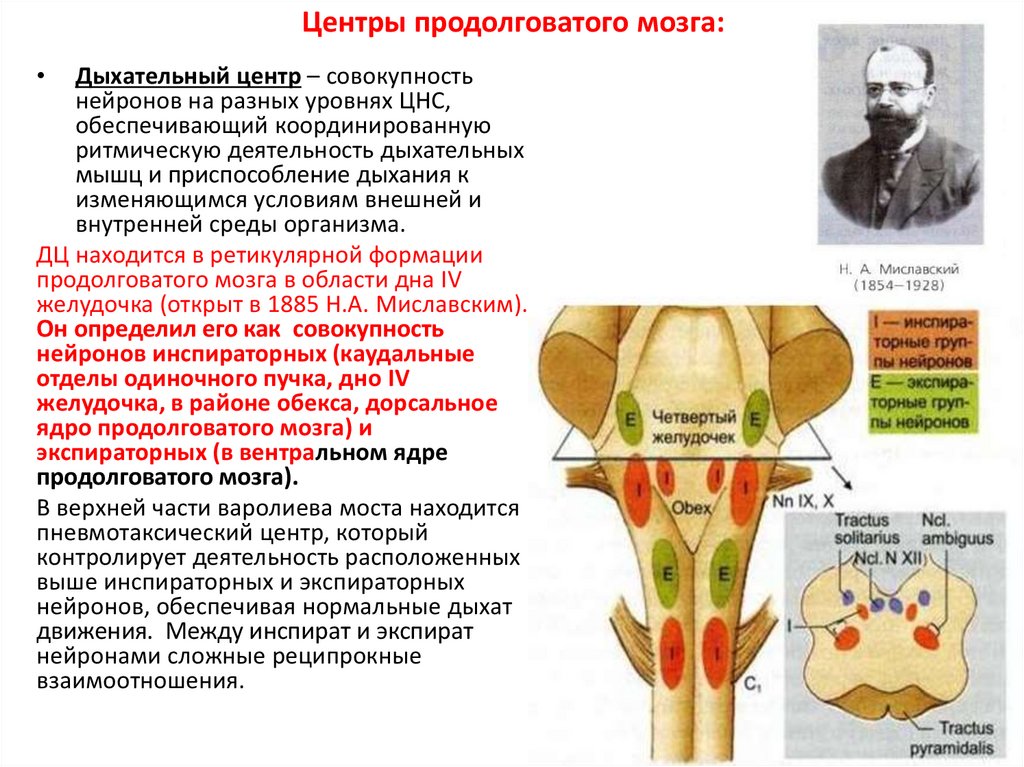
Центры продолговатого мозга:Дыхательный центр – совокупность
нейронов на разных уровнях ЦНС,
обеспечивающий координированную
ритмическую деятельность дыхательных
мышц и приспособление дыхания к
изменяющимся условиям внешней и
внутренней среды организма.
ДЦ находится в ретикулярной формации
продолговатого мозга в области дна IV
желудочка (открыт в 1885 Н.А. Миславским).
Он определил его как совокупность
нейронов инспираторных (каудальные
отделы одиночного пучка, дно IV
желудочка, в районе обекса, дорсальное
ядро продолговатого мозга) и
экспираторных (в вентральном ядре
продолговатого мозга).
В верхней части варолиева моста находится
пневмотаксический центр, который
контролирует деятельность расположенных
выше инспираторных и экспираторных
нейронов, обеспечивая нормальные дыхат
движения. Между инспират и экспират
нейронами сложные реципрокные
взаимоотношения.
7.
• Сосудодвигательный центр. морфофункциональноеобразование продолговатого мозга, играющее ведущую роль в
поддержании тонуса сосудов и рефлекторной регуляции
кровяного давления. СДЦ был открыт в 1871
г.Ф.В.Овсянниковым. Бульбарный отдел сосудодвигательного
центра расположен на уровне нейронов ретикулярной
формации продолговатого мозга. Методом локальной
стимуляции установлено наличие прессорных и депрессорных
нейронов. Они расположены диффузно, но в ростральном
отделе больше прессорных нейронов, а в каудальном депрессорных. В составе бульбарного отдела
сосудодвигательного центра есть кардиоингибирующие
нейроны.
• Ядра ПМ принимают участии в выполнении рефлекторных
актов жевания, сосания, глотания, слюноотделения. Пример:
Жевание - двигательный акт, который может осуществляться
рефлекторно в ответ на раздражение рецепторов ротовой
полости и состоит в перемещении нижней челюсти по
отношению к верхней. Более тонкая регуляция достигается
только при целостности таламуса и моторных зон коры.
8.
9.
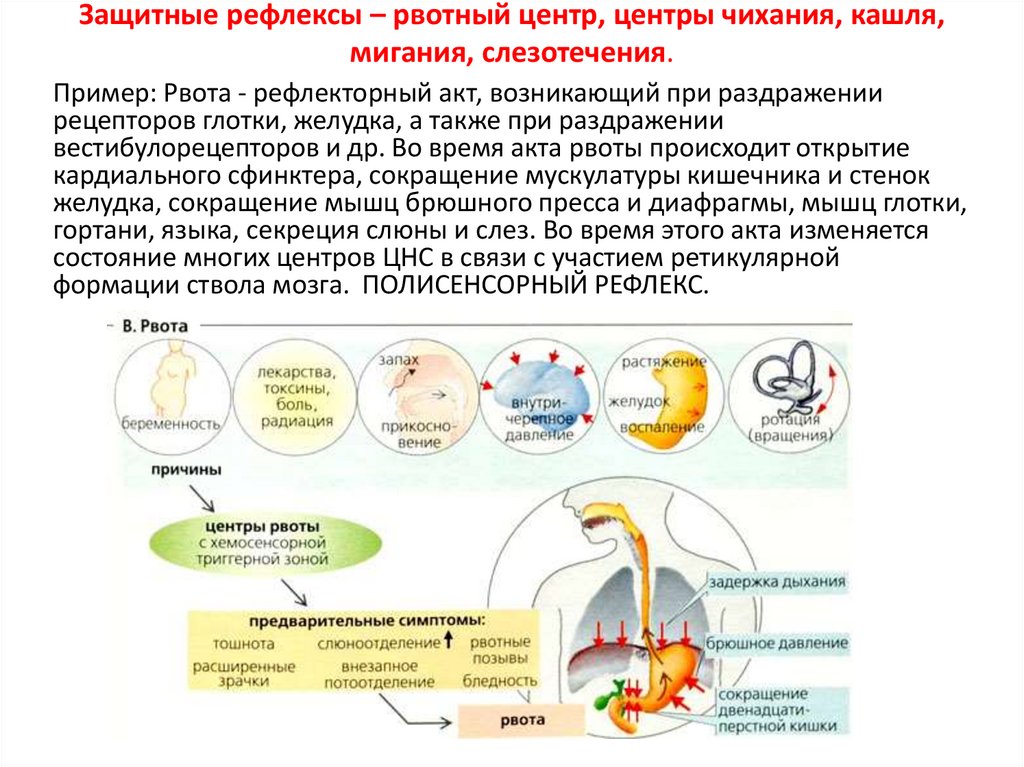
Защитные рефлексы – рвотный центр, центры чихания, кашля,мигания, слезотечения.
Пример: Рвота - рефлекторный акт, возникающий при раздражении
рецепторов глотки, желудка, а также при раздражении
вестибулорецепторов и др. Во время акта рвоты происходит открытие
кардиального сфинктера, сокращение мускулатуры кишечника и стенок
желудка, сокращение мышц брюшного пресса и диафрагмы, мышц глотки,
гортани, языка, секреция слюны и слез. Во время этого акта изменяется
состояние многих центров ЦНС в связи с участием ретикулярной
формации ствола мозга. ПОЛИСЕНСОРНЫЙ РЕФЛЕКС.
10.
Варолиев мост• V пара черепных нервов – n. trigeminus – тройничный
нерв.По функции – смешанный нерв. Содержит
двигательные, чувствительные и парасимпатические
волокна. Иннервирует все жевательные мышцы, кожу
лица, зубы, железы полости рта.
• VI пара черепных нервов – n.abducens – отводящий
нерв. По функции – двигательный. Иннервирует
наружную прямую мышцу глаза.
• VII пара черепных нервов – n. facialis – лицевой нерв.
Нерв смешанный, преимущественно двигательный для
мимической мускулатуры лица. Имеет три ядра:
двигательное, чувствительное, вегетативное.
• VIII пара черепных нервов —преддверно-улитковый
нерв - n. vestibulocochlearis - состоит из улитковой и
преддверной частей. Улитковое ядро лежит в ПМ. Нерв
чувствительный. Волокна идут от органа слуха и
равновесия.
11.
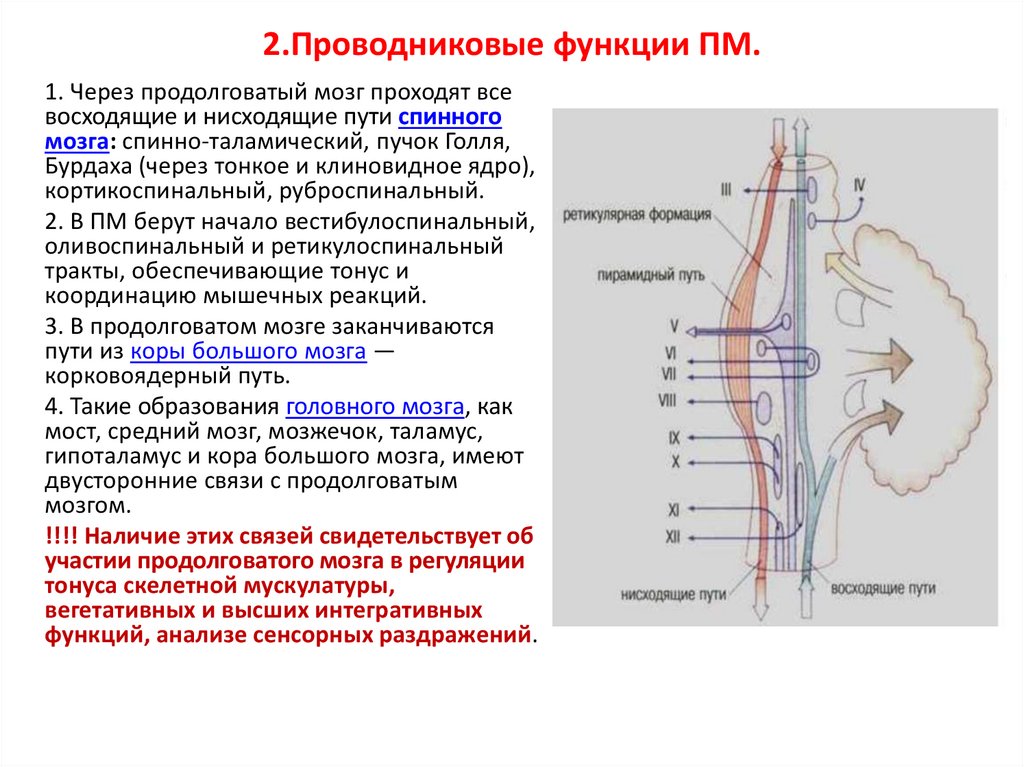
2.Проводниковые функции ПМ.1. Через продолговатый мозг проходят все
восходящие и нисходящие пути спинного
мозга: спинно-таламический, пучок Голля,
Бурдаха (через тонкое и клиновидное ядро),
кортикоспинальный, руброспинальный.
2. В ПМ берут начало вестибулоспинальный,
оливоспинальный и ретикулоспинальный
тракты, обеспечивающие тонус и
координацию мышечных реакций.
3. В продолговатом мозге заканчиваются
пути из коры большого мозга —
корковоядерный путь.
4. Такие образования головного мозга, как
мост, средний мозг, мозжечок, таламус,
гипоталамус и кора большого мозга, имеют
двусторонние связи с продолговатым
мозгом.
!!!! Наличие этих связей свидетельствует об
участии продолговатого мозга в регуляции
тонуса скелетной мускулатуры,
вегетативных и высших интегративных
функций, анализе сенсорных раздражений.
12.
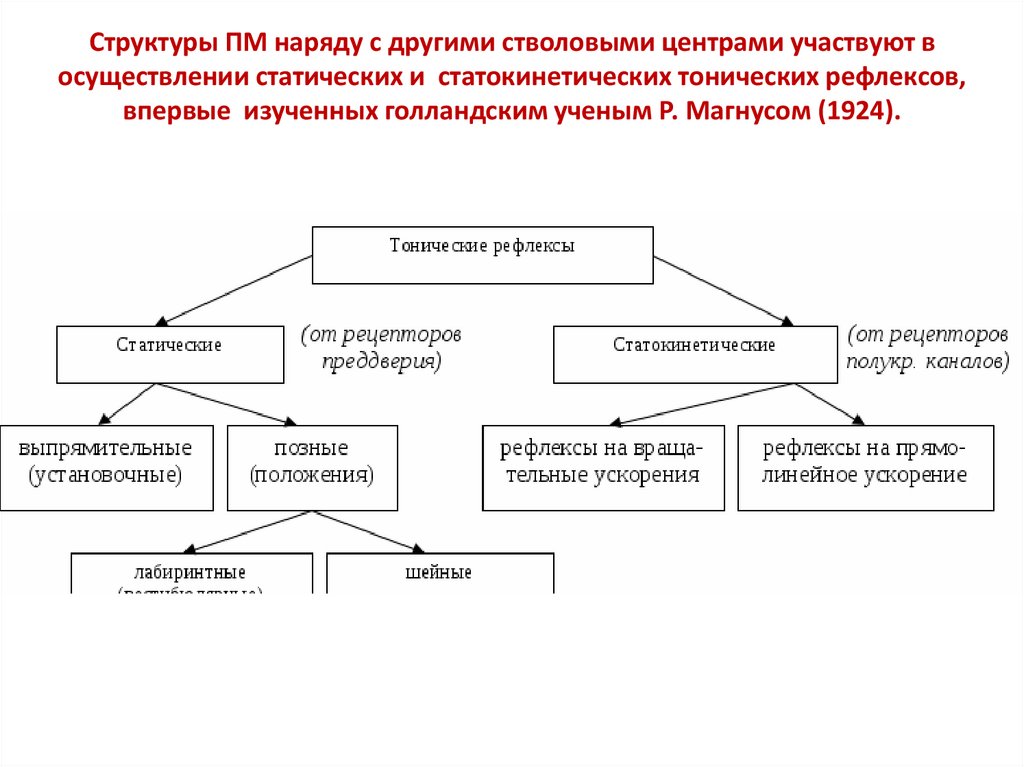
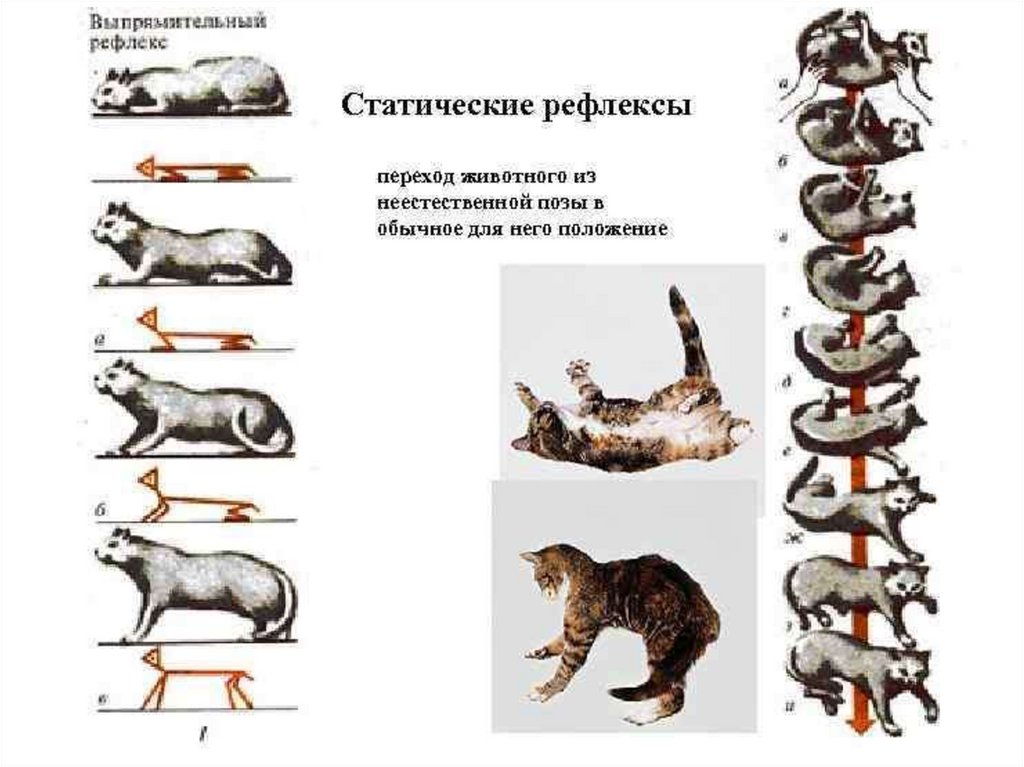
Структуры ПМ наряду с другими стволовыми центрами участвуют восуществлении статических и статокинетических тонических рефлексов,
впервые изученных голландским ученым Р. Магнусом (1924).
13.
А. Статические рефлексы обусловливают положение тела и его равновесие в покое.Магнус разделил их на рефлексы позы (положения тела), выпрямительные
рефлексы и рефлексы компенсаторного положения глаз.
1. Рефлексы позы
• возникают при изменениях
положения головы (сдвиг центра
равновесия) и направлены на
создание удобной позы.
Афферентные импульсы поступают
с рецепторов отолитового
аппарата и проприорецепторов
мышц шеи.
• При вертикальной позе человека
(голова вверх, шея наклонена
несколько вперед) происходит
повышение тонуса разгибателей
нижних конечностей и сгибателей
верхних конечностей. При этом
шейные и лабиринтные рефлексы
усиливают друг друга.
14.
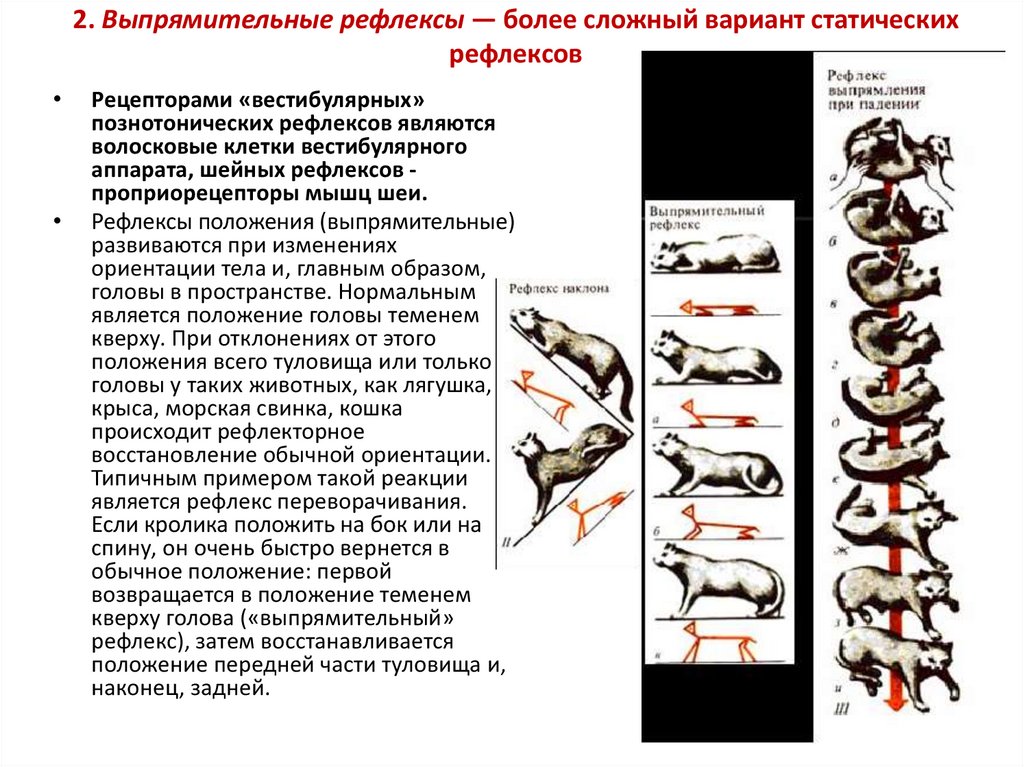
2. Выпрямительные рефлексы — более сложный вариант статическихрефлексов
Рецепторами «вестибулярных»
познотонических рефлексов являются
волосковые клетки вестибулярного
аппарата, шейных рефлексов проприорецепторы мышц шеи.
Рефлексы положения (выпрямительные)
развиваются при изменениях
ориентации тела и, главным образом,
головы в пространстве. Нормальным
является положение головы теменем
кверху. При отклонениях от этого
положения всего туловища или только
головы у таких животных, как лягушка,
крыса, морская свинка, кошка
происходит рефлекторное
восстановление обычной ориентации.
Типичным примером такой реакции
является рефлекс переворачивания.
Если кролика положить на бок или на
спину, он очень быстро вернется в
обычное положение: первой
возвращается в положение теменем
кверху голова («выпрямительный»
рефлекс), затем восстанавливается
положение передней части туловища и,
наконец, задней.
15.
16.
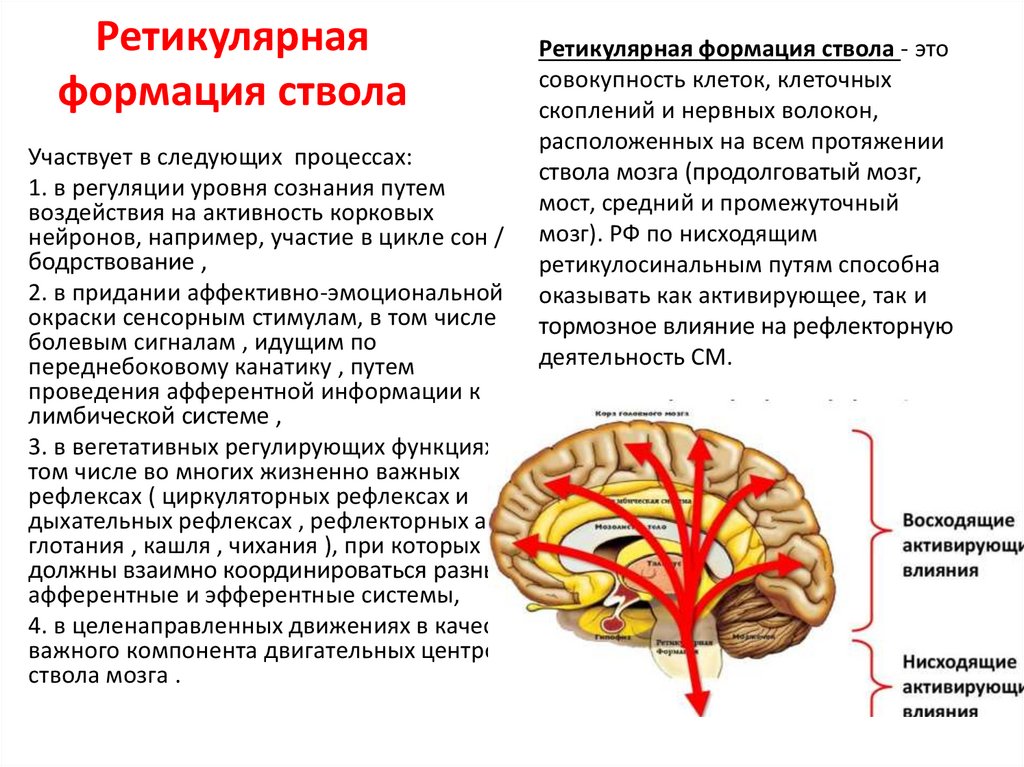
Ретикулярнаяформация ствола
Участвует в следующих процессах:
1. в регуляции уровня сознания путем
воздействия на активность корковых
нейронов, например, участие в цикле сон /
бодрствование ,
2. в придании аффективно-эмоциональной
окраски сенсорным стимулам, в том числе
болевым сигналам , идущим по
переднебоковому канатику , путем
проведения афферентной информации к
лимбической системе ,
3. в вегетативных регулирующих функциях, в
том числе во многих жизненно важных
рефлексах ( циркуляторных рефлексах и
дыхательных рефлексах , рефлекторных актах
глотания , кашля , чихания ), при которых
должны взаимно координироваться разные
афферентные и эфферентные системы,
4. в целенаправленных движениях в качестве
важного компонента двигательных центров
ствола мозга .
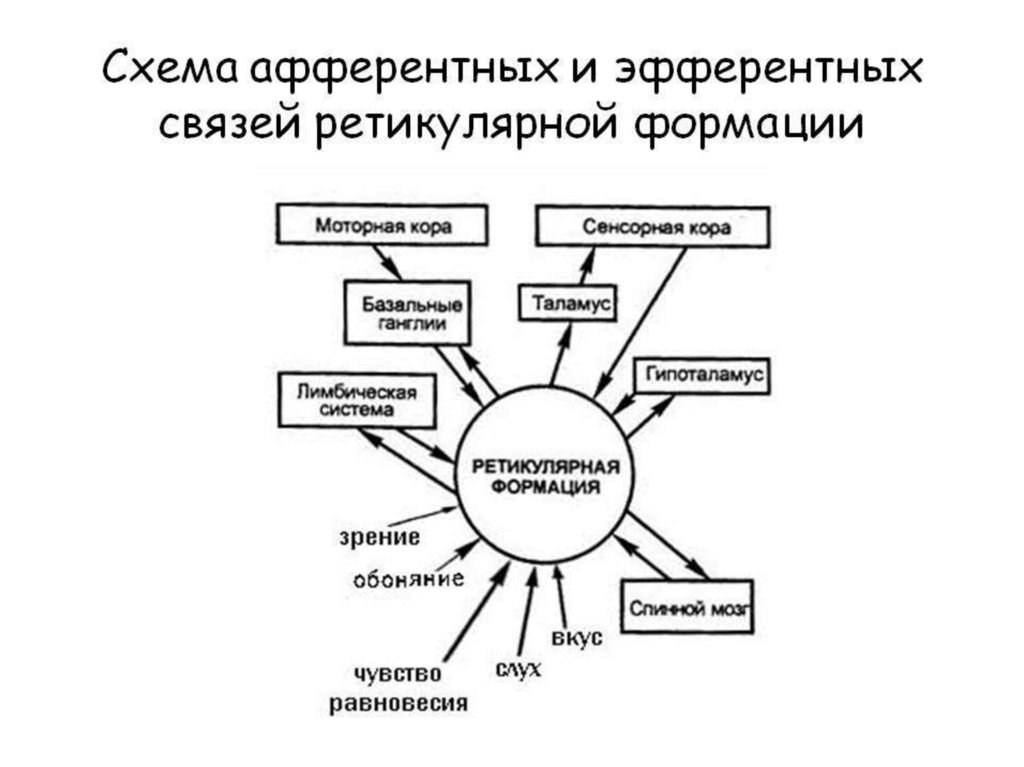
Ретикулярная формация ствола - это
совокупность клеток, клеточных
скоплений и нервных волокон,
расположенных на всем протяжении
ствола мозга (продолговатый мозг,
мост, средний и промежуточный
мозг). РФ по нисходящим
ретикулосинальным путям способна
оказывать как активирующее, так и
тормозное влияние на рефлекторную
деятельность СМ.
17.
18.
19.
Средний мозг содержит ядра четверохолмия, расположенные надсильвиевым водопроводом, красное ядро, ядра III и IV пары ЧМН,
черную субстанцию.
• III пара черепных нервов – n. oculomotorius – глазодвигательный
нерв. По функции – смешанный, но преимущественно двигательный
для мышц глаза. Имеет двигательное и парасимпатическое ядра –
(nucleus accessorius).
• IV пара черепных нервов – n. trochlearis – блоковый нерв. По функции
– двигательный. Иннервирует верхнюю косую мышцу глаза. Черная
субстанция (основание среднего мозга).
• Четверохолмие – часть стволового отдела зрительной (верхние
холмики) и слуховой (нижние холмики) сенсорных систем. Верхние
бугры четверохолмия – первичные зрительные центры ,при их
участии реализуются рефлексы в ответ на световые раздражения. С
участием ядер глазодвигат и блокового нервов + Зрачковый рефлекс.
• Нижние бугры четверохолмия - первичные слуховые центры.
Ориентировочные слуховые рефлексы - рефлекс настораживания,
старт-рефлекс, сторожевой рефлекс – значение которого подготовить
организм к реакции на новое внезапное раздражение. За эту реакцию
отвечает нисходящий тектоспинальный путь. Существенным здесь
является перераспределение мышечного тонуса в сторону усиления
тонуса мышц-сгибателей,что способствует бегству или нападению.
Человек с нарушениями в области четверохолмия не способен быстро
реагировать на неожиданный раздражитель.
20.
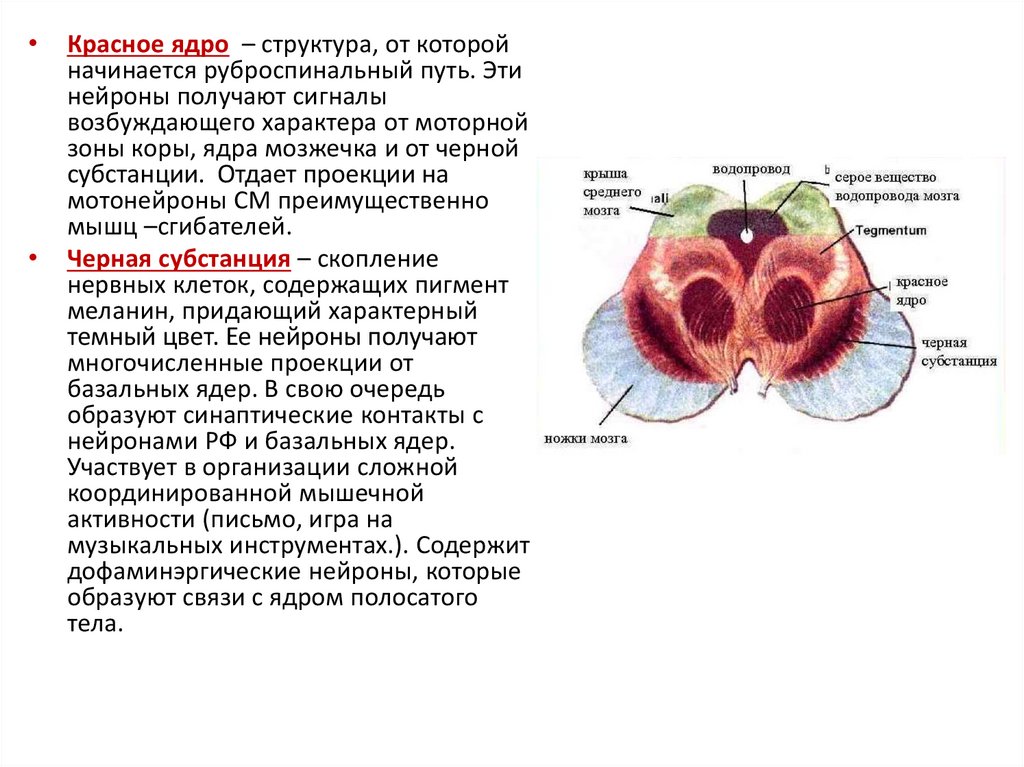
Красное ядро – структура, от которой
начинается руброспинальный путь. Эти
нейроны получают сигналы
возбуждающего характера от моторной
зоны коры, ядра мозжечка и от черной
субстанции. Отдает проекции на
мотонейроны СМ преимущественно
мышц –сгибателей.
Черная субстанция – скопление
нервных клеток, содержащих пигмент
меланин, придающий характерный
темный цвет. Ее нейроны получают
многочисленные проекции от
базальных ядер. В свою очередь
образуют синаптические контакты с
нейронами РФ и базальных ядер.
Участвует в организации сложной
координированной мышечной
активности (письмо, игра на
музыкальных инструментах.). Содержит
дофаминэргические нейроны, которые
образуют связи с ядром полосатого
тела.
21.
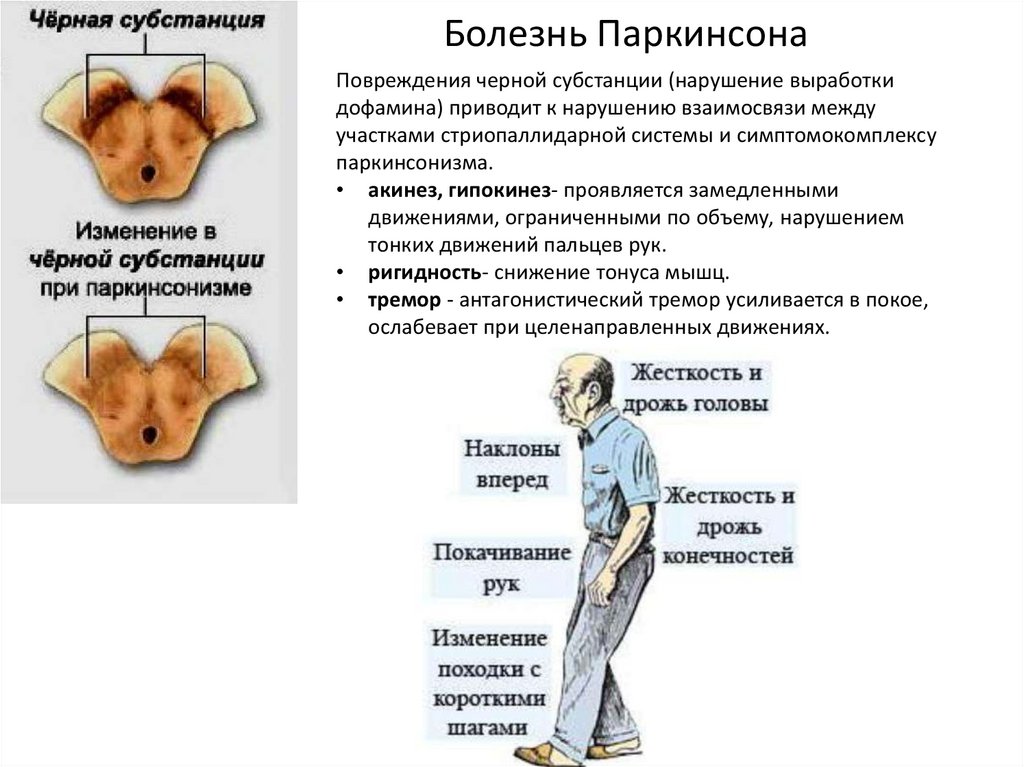
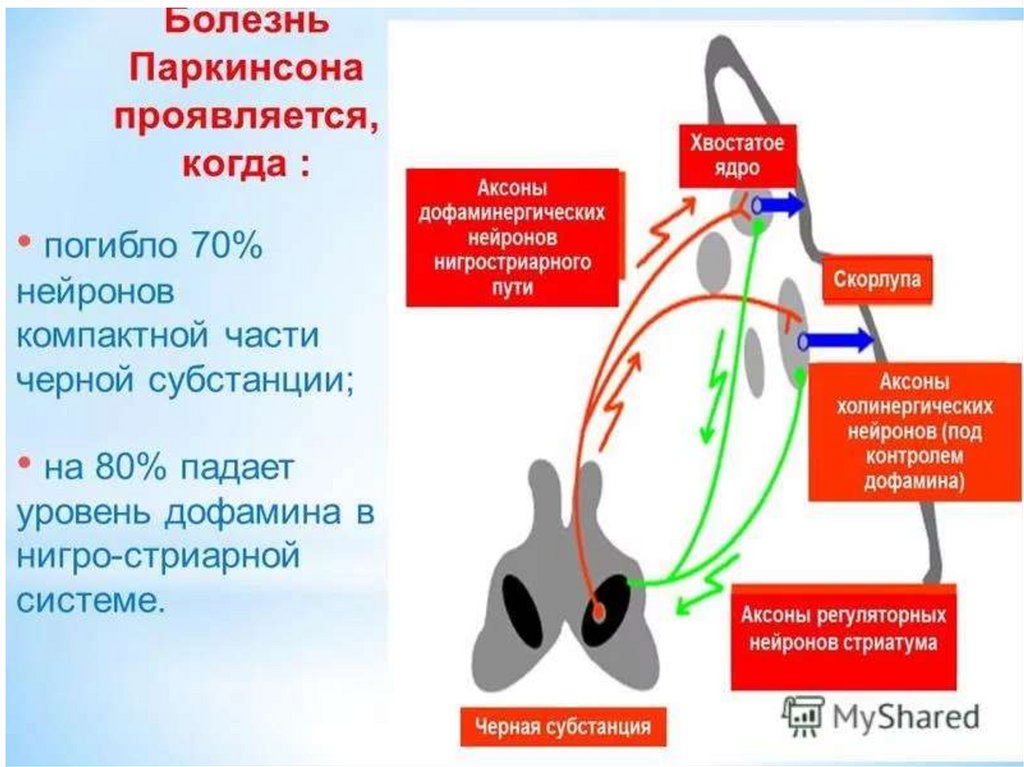
Болезнь ПаркинсонаПовреждения черной субстанции (нарушение выработки
дофамина) приводит к нарушению взаимосвязи между
участками стриопаллидарной системы и симптомокомплексу
паркинсонизма.
• акинез, гипокинез- проявляется замедленными
движениями, ограниченными по объему, нарушением
тонких движений пальцев рук.
• ригидность- снижение тонуса мышц.
• тремор - антагонистический тремор усиливается в покое,
ослабевает при целенаправленных движениях.
22.
23.
Статокинетические рефлексы могут осуществлятьсятолько при целостности структур среднего мозга.
• возникают при ускорениях прямолинейного и
вращательного движений организма.
• Сокращения мыщц при этом направлены на
преодоление действующих на человека ускорений,
сохранение нормальной позы, равновесия и
ориентации в пространстве.
• Для их осуществления необходимо сохранение
функции моторных центров ствола мозга не ниже
уровня среднего мозга.
• Эти рефлексы запускаются с рецепторов
вестибулярного аппарата: с рецепторов
отолитового аппарата возникают рефлексы
прямолинейного ускорения, с рецепторов ампул
полукружных каналов — рефлексы вращения.
24.
25.
26.
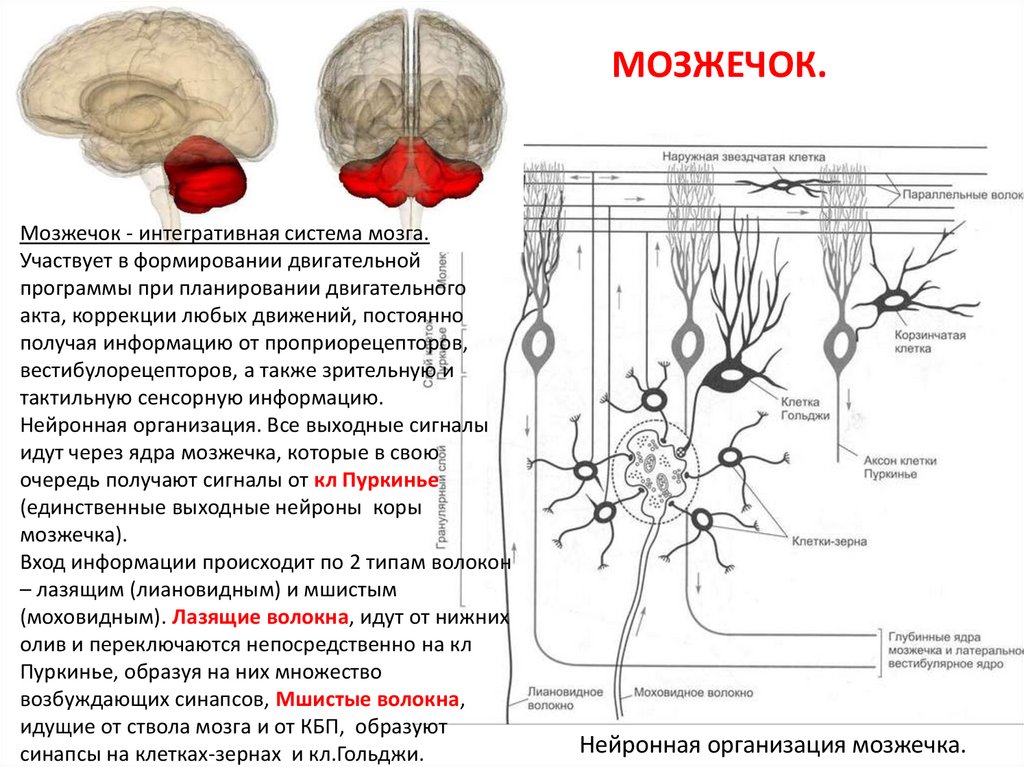
МОЗЖЕЧОК.Мозжечок - интегративная система мозга.
Участвует в формировании двигательной
программы при планировании двигательного
акта, коррекции любых движений, постоянно
получая информацию от проприорецепторов,
вестибулорецепторов, а также зрительную и
тактильную сенсорную информацию.
Нейронная организация. Все выходные сигналы
идут через ядра мозжечка, которые в свою
очередь получают сигналы от кл Пуркинье
(единственные выходные нейроны коры
мозжечка).
Вход информации происходит по 2 типам волокон
– лазящим (лиановидным) и мшистым
(моховидным). Лазящие волокна, идут от нижних
олив и переключаются непосредственно на кл
Пуркинье, образуя на них множество
возбуждающих синапсов, Мшистые волокна,
идущие от ствола мозга и от КБП, образуют
синапсы на клетках-зернах и кл.Гольджи.
Нейронная организация мозжечка.
27.
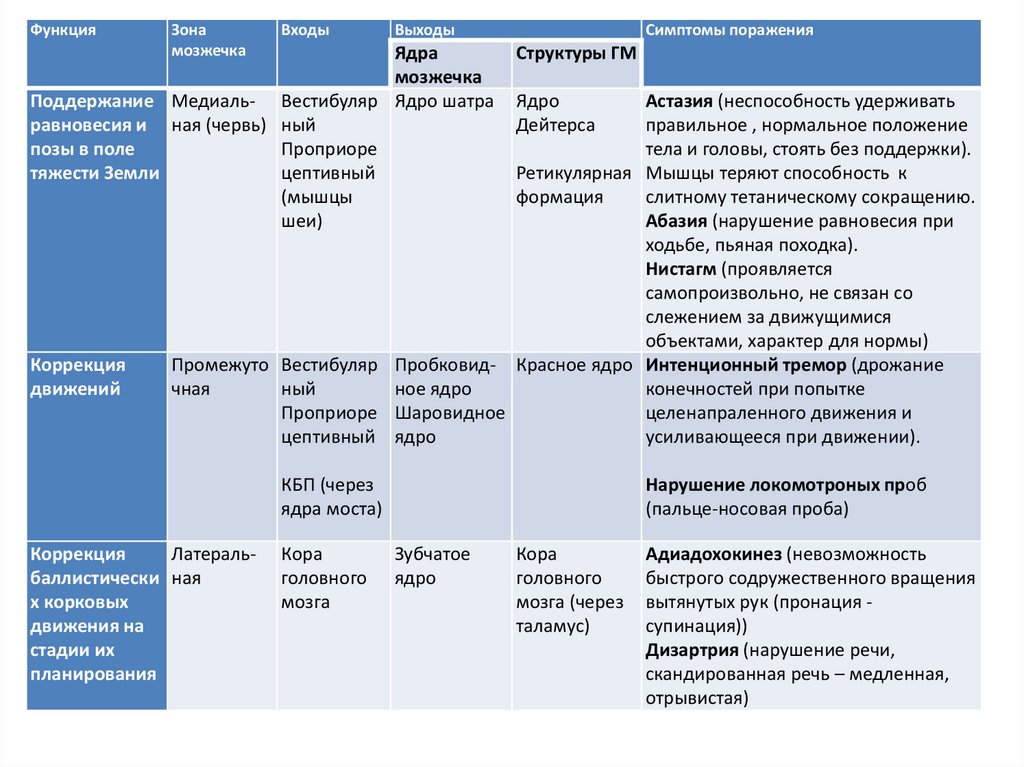
ФункцияЗона
мозжечка
Входы
Выходы
Ядра
мозжечка
Поддержание Медиаль- Вестибуляр Ядро шатра
равновесия и ная (червь) ный
позы в поле
Проприоре
тяжести Земли
цептивный
(мышцы
шеи)
Коррекция
движений
Симптомы поражения
Структуры ГМ
Ядро
Дейтерса
Астазия (неспособность удерживать
правильное , нормальное положение
тела и головы, стоять без поддержки).
Ретикулярная Мышцы теряют способность к
формация
слитному тетаническому сокращению.
Абазия (нарушение равновесия при
ходьбе, пьяная походка).
Нистагм (проявляется
самопроизвольно, не связан со
слежением за движущимися
объектами, характер для нормы)
Промежуто Вестибуляр Пробковид- Красное ядро Интенционный тремор (дрожание
чная
ный
ное ядро
конечностей при попытке
Проприоре Шаровидное
целенапраленного движения и
цептивный ядро
усиливающееся при движении).
КБП (через
ядра моста)
Коррекция
Латеральбаллистически ная
х корковых
движения на
стадии их
планирования
Кора
головного
мозга
Нарушение локомотроных проб
(пальце-носовая проба)
Зубчатое
ядро
Кора
головного
мозга (через
таламус)
Адиадохокинез (невозможность
быстрого содружественного вращения
вытянутых рук (пронация супинация))
Дизартрия (нарушение речи,
скандированная речь – медленная,
отрывистая)
28.
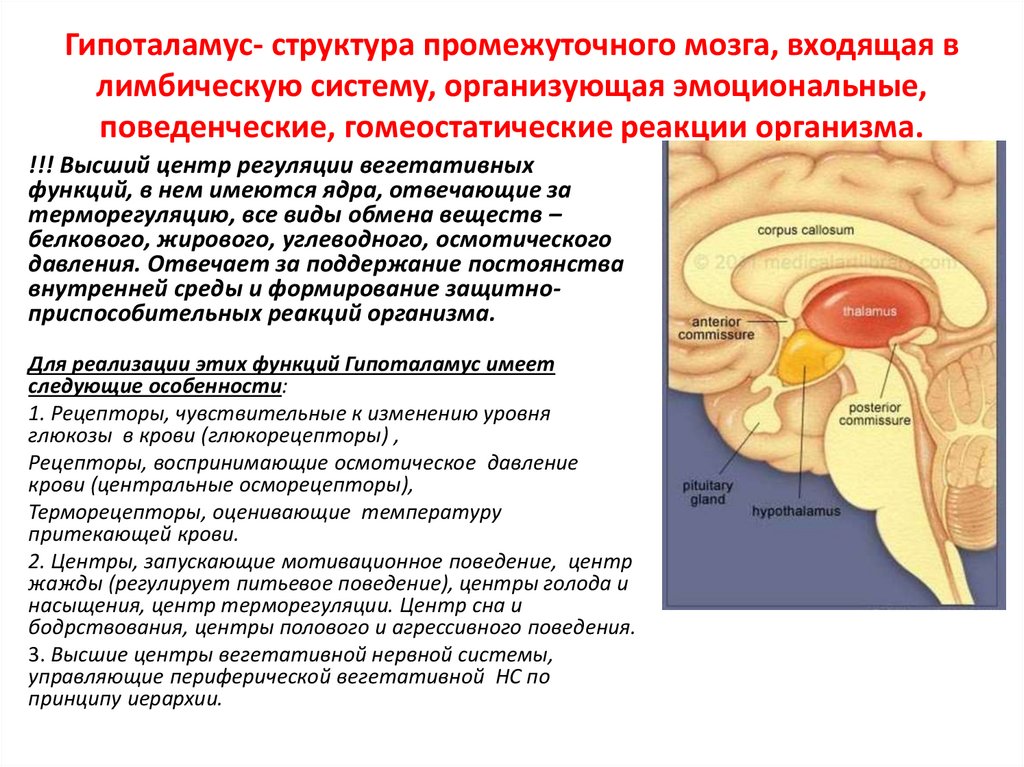
Гипоталамус- структура промежуточного мозга, входящая влимбическую систему, организующая эмоциональные,
поведенческие, гомеостатические реакции организма.
!!! Высший центр регуляции вегетативных
функций, в нем имеются ядра, отвечающие за
терморегуляцию, все виды обмена веществ –
белкового, жирового, углеводного, осмотического
давления. Отвечает за поддержание постоянства
внутренней среды и формирование защитноприспособительных реакций организма.
Для реализации этих функций Гипоталамус имеет
следующие особенности:
1. Рецепторы, чувствительные к изменению уровня
глюкозы в крови (глюкорецепторы) ,
Рецепторы, воспринимающие осмотическое давление
крови (центральные осморецепторы),
Терморецепторы, оценивающие температуру
притекающей крови.
2. Центры, запускающие мотивационное поведение, центр
жажды (регулирует питьевое поведение), центры голода и
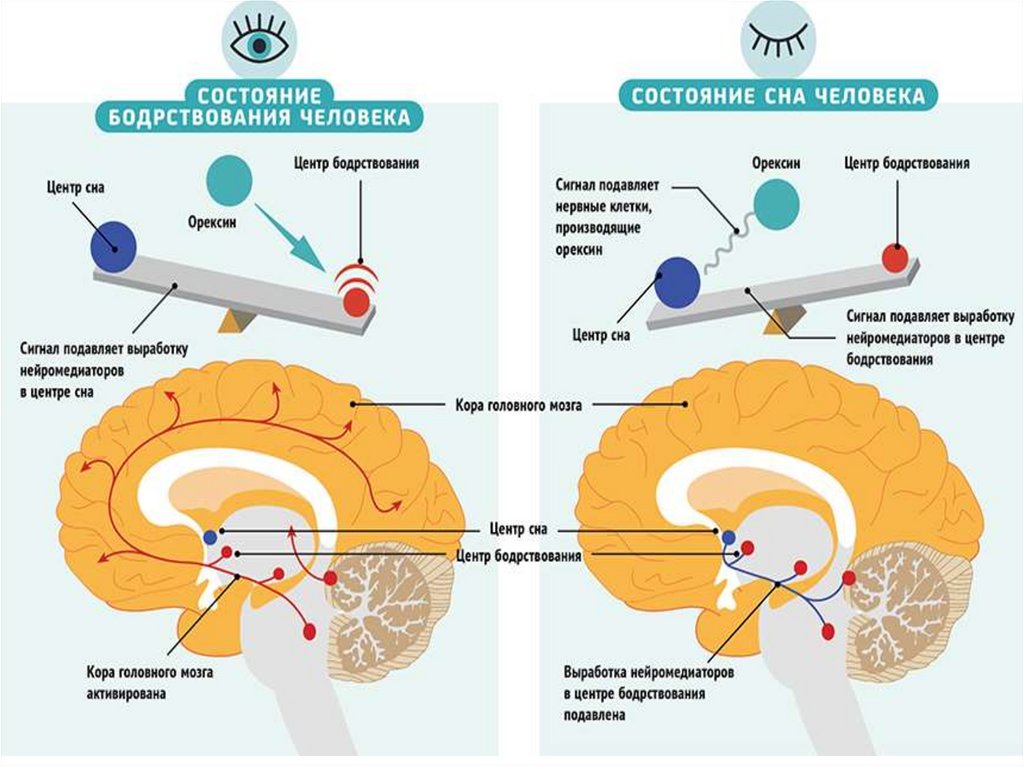
насыщения, центр терморегуляции. Центр сна и
бодрствования, центры полового и агрессивного поведения.
3. Высшие центры вегетативной нервной системы,
управляющие периферической вегетативной НС по
принципу иерархии.
29.
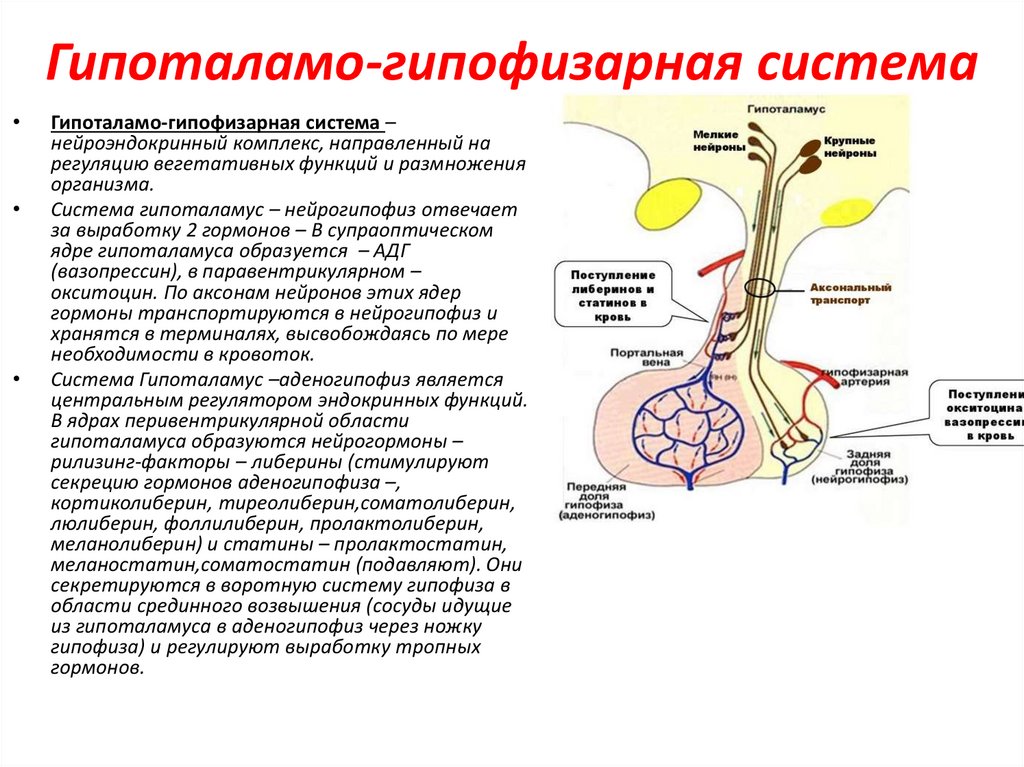
Гипоталамо-гипофизарная системаГипоталамо-гипофизарная система –
нейроэндокринный комплекс, направленный на
регуляцию вегетативных функций и размножения
организма.
Система гипоталамус – нейрогипофиз отвечает
за выработку 2 гормонов – В супраоптическом
ядре гипоталамуса образуется – АДГ
(вазопрессин), в паравентрикулярном –
окситоцин. По аксонам нейронов этих ядер
гормоны транспортируются в нейрогипофиз и
хранятся в терминалях, высвобождаясь по мере
необходимости в кровоток.
Система Гипоталамус –аденогипофиз является
центральным регулятором эндокринных функций.
В ядрах перивентрикулярной области
гипоталамуса образуются нейрогормоны –
рилизинг-факторы – либерины (стимулируют
секрецию гормонов аденогипофиза –,
кортиколиберин, тиреолиберин,соматолиберин,
люлиберин, фоллилиберин, пролактолиберин,
меланолиберин) и статины – пролактостатин,
меланостатин,соматостатин (подавляют). Они
секретируются в воротную систему гипофиза в
области срединного возвышения (сосуды идущие
из гипоталамуса в аденогипофиз через ножку
гипофиза) и регулируют выработку тропных
гормонов.
30.
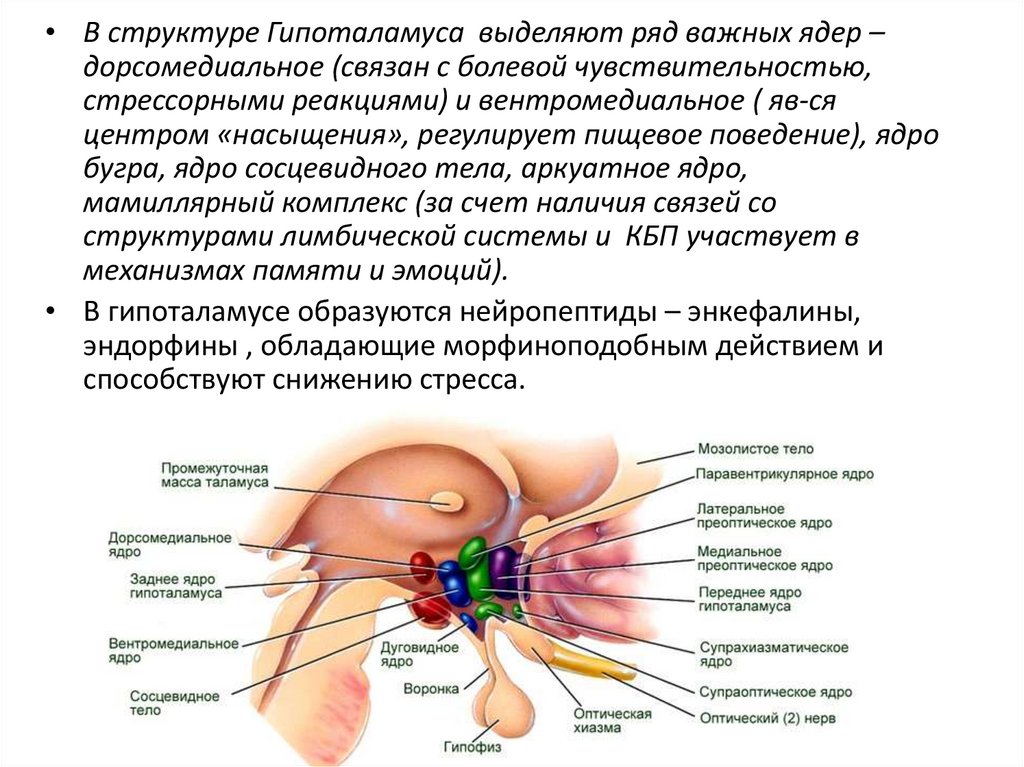
• В структуре Гипоталамуса выделяют ряд важных ядер –дорсомедиальное (связан с болевой чувствительностью,
стрессорными реакциями) и вентромедиальное ( яв-ся
центром «насыщения», регулирует пищевое поведение), ядро
бугра, ядро сосцевидного тела, аркуатное ядро,
мамиллярный комплекс (за счет наличия связей со
структурами лимбической системы и КБП участвует в
механизмах памяти и эмоций).
• В гипоталамусе образуются нейропептиды – энкефалины,
эндорфины , обладающие морфиноподобным действием и
способствуют снижению стресса.
31.
ТАЛАМУСТаламус – универсальные ворота к коре головного мозга: вся информация поступающая в
КБП проходит через таламуса: исключение –основная часть обонятельных путей.
Считается , что функция таламуса –сенсорная фильтрация, пропускается только
значимая информация.
• Таламус — парное образование, составляющее основную массу промежуточного мозга,
получающий импульсы всех видов чувствительности, кроме обонятельных. В таламусе
выделяют до 120 ядер серого вещества. По месту их расположения ядра делят на передние,
латеральные и медиальные группы. В задней части латеральной группы ядер таламуса
выделяют подушку, медиальное и латеральное коленчатые тела.
• Одной из важнейших функций таламуса является анализ, отбор и передача в кору головного
мозга сенсорных сигналов, поступающих к нему из большинства сенсорных систем ЦНС.
• По выполняемым функциям ядра таламуса делятся на специфические, ассоциативные и
неспецифические.
1. Специфические ядра получают сигналы от вторых нейронов длинных восходящих
афферентных путей, проводящих в кору мозга соматосенсорные, зрительные, слуховые сигналы.
Эти ядра, передают обработанные сигналы в проекционные (хорошо очерченные) области коры
— соматосенсорную, слуховую, зрительную сенсорные области, а также в премоторную и
первичную моторные области коры. С нейронами этих областей коры специфические ядра
таламуса имеют реципрокные связи..
К специфическим ядрам таламуса подходят волокна проводящих путей от
коры, ретикулярной формации и ядер ствола мозга. По этим путям могут
передаваться как возбуждающие, так и тормозные влияния на активность
нейронов ядер. Благодаря таким связям кора мозга может регулировать
потоки идущей к ней информации и отбирать наиболее значимую в данный
момент. При этом кора может блокировать передачу сигналов одной
модальности и облегчать передачу другой.
32.
Функциональная организация ядер таламуса:1. Среди специфических ядер таламуса имеются также
несенсорные ядра. Они обеспечивают обработку и переключение
сигналов не от чувствительных восходящих путей, а от других
областей мозга. К нейронам таких ядер поступают сигналы от
красного ядра, базальных ганглиев, лимбической системы,
зубчатого ядра мозжечка, которые после их обработки проводятся
к нейронам моторной коры.
• Ядра передней группы таламуса участвуют в передаче сигналов
от мамиллярных тел к лимбической системе, обеспечивая
круговую циркуляцию нервных импульсов по кольцу:
лимбическая кора — гиппокамп — гипоталамус —
миндалевидное тело — таламус — лимбическая кора.
Нейронную сеть, сформированную этими структурами,
называют кругом (кольцом) Пайпеца. Циркуляция сигналов по
структурам этого круга связана с запоминанием новой
информации и формированием эмоций — эмоциональное
кольцо Пайпеца.
33.
2. Ассоциативные ядра таламуса расположены преимущественномедиодорсально, латерально и в ядре подушки. Они отличаются
от специфических тем, что к их нейронам не поступают сигналы из
чувствительных восходящих путей, а поступают сигналы уже
обработанные в других нервных центрах и ядрах таламуса.
Ассоциативность нейронов этих ядер выражается в том, что на
один и тот же нейрон ядра приходят сигналы разных
модальностей. Изменение активности нейронов ядер может быть
связано (ассоциировано) с поступлением разнородных сигналов
из разных источников (например, от центров, обеспечивающих
зрительную, тактильную и болевую чувствительность).
• Нейроны ассоциативных ядер являются полисенсорными и
обеспечивают возможность осуществления интегративных
процессов, в результате которых формируются обобщенные
сигналы, передающиеся в ассоциативные области коры
лобной, теменной и височной долей коры. Потоки этих
сигналов способствуют осуществлению корой таких
психических процессов, как узнавание предметов и явлений,
согласование речевых, зрительных и двигательных функций,
формирование представления о позе тела, трехмерности
пространства и положении в нем тела человека.
34.
3. Неспецифические ядра таламуса представленыпреимущественно интраламинарными, центральными и
ретикулярными группами ядер таламуса. Они состоят из мелких
нейронов, к которым по многочисленным синаптическим связям
поступают сигналы от нейронов других ядер таламуса,
лимбической системы, базальных ядер, гипоталамуса, ствола
мозга. По чувствительным восходящим путям к неспецифическим
ядрам поступает сигнализация от болевых и температурных
рецепторов, а по сетям нейронов ретикулярной формации —
сигнализация практически от всех других сенсорных систем ЦНС.
• Эфферентные пути от неспецифических ядер идут ко всем
зонам коры как непосредственно, так и через другие
таламические и ретикулярные ядра. От неспецифических ядер
таламуса начинаются также нисходящие пути к стволу мозга.
При повышении активности неспецифических ядер таламуса
(например, при электрической стимуляции в эксперименте)
регистрируется диффузное повышение нейронной активности
практически во всех областях коры больших полушарий.
• Они оказывают модулирующее влияние на состояние
активности нервных центров, создают условия для их
оптимальной настройки на выполнение работы.