






















 Биология
БиологияПохожие презентации:

. Дәріс 2")








ДНҚ репликациясы. Нәруыз биосинтезі
1.
ӨмірбекДилда
Өмірбекқызы
Білімге құштарлық табысқа
апаратын жол
2.
ДНҚ репликациясы.Нәруыз биосинтезі
3.
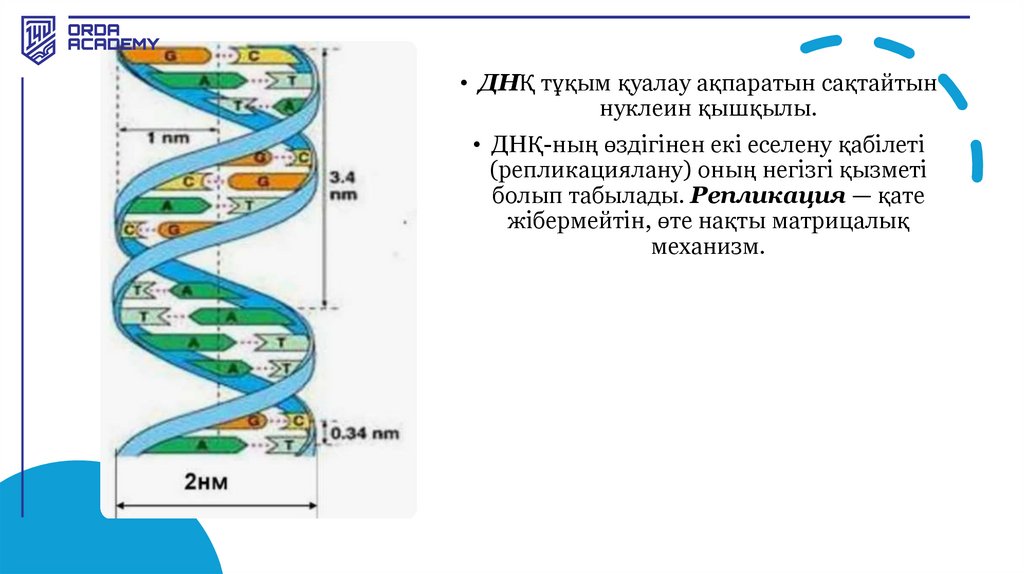
• ДНҚ тұқым қуалау ақпаратын сақтайтыннуклеин қышқылы.
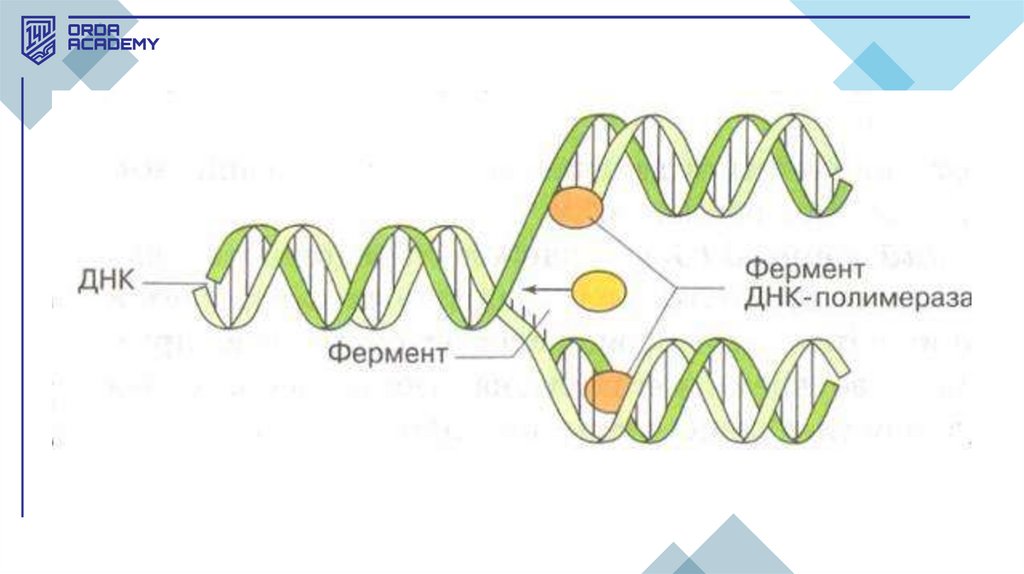
• ДНҚ-ның өздігінен екі еселену қабілеті
(репликациялану) оның негізгі қызметі
болып табылады. Репликация — қате
жібермейтін, өте нақты матрицалық
механизм.
4.
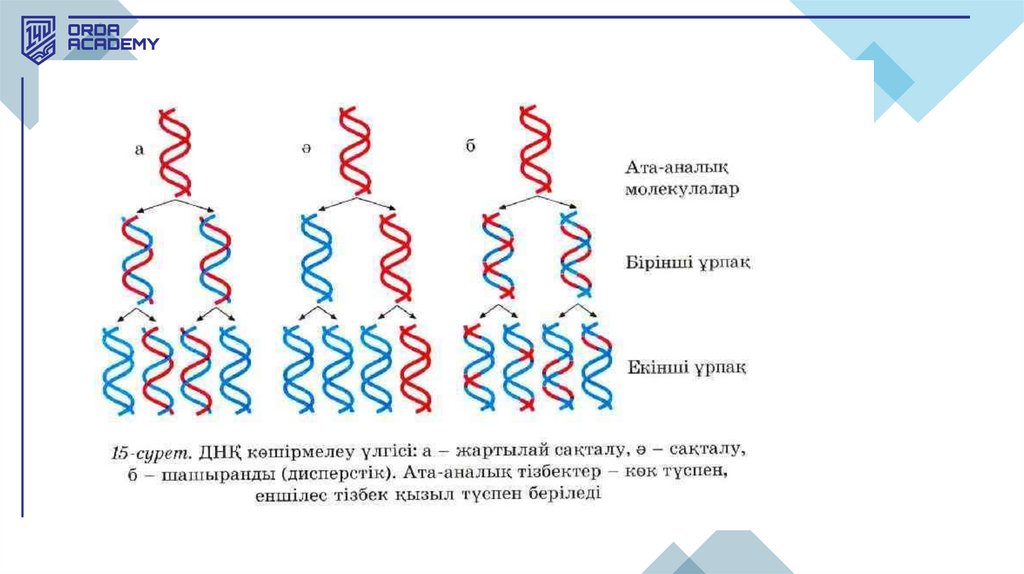
• ДНҚ репликациясы жартылай консервативті механизм арқылыжүзеге асады (Мезельсон мен Сталь дәлелдеді).
• ДНҚ репликациясының жартылай консервативті механизмінде әр тізбек
матрица (шаблон) қызметін атқарады, ал жаңа синтезделген молекулалар
гибридті болады (бір ескі және бір жаңа тізбектен тұрады). Мұндай
жартылай сақтала екі еселену механизмі жартылай консервативті
репликация деп аталады. Басқа ұсынылған модельдер консервативті
(толығымен сақтала екі еселену) және дисперсивті (бытыраңқы екі
еселену) болды.
5.
6.
7.
8.
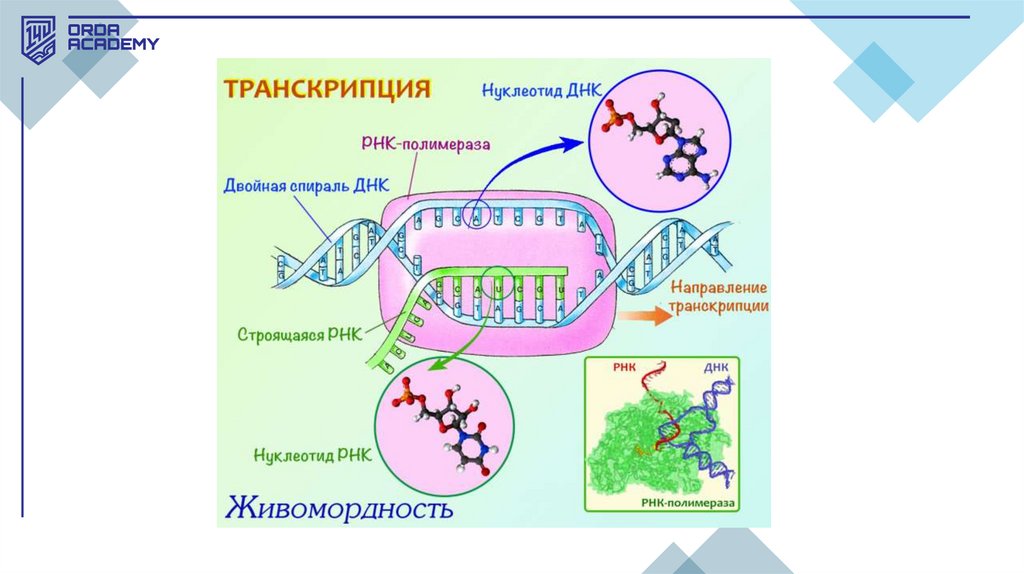
Транскрипция• Транскрипция (РНҚ синтезі) – ДНҚ-дан нәруыз синтезіне
қажетті ақпаратты алып, оны нәруыз синтезі жүретін орынға
жеткізу процесі. Бұл процесті ақпараттық РНҚ (м-РНҚ) жүзеге
асырады. Сонымен қатар, жасушада тасымалдаушы РНҚ (тРНҚ) мен рибосомалық РНҚ (р-РНҚ) синтезі қажет, себебі олар
нәруыз синтезіне тікелей қатысады. Транскрипция процесі
генетикалық ақпаратты РНҚ-ға көшіріп жазу ретінде
орындалады, ол ДНҚ тізбегінде жүреді.
9.
• Транскрипция процесіне келесі негізгі элементтер қатысады:1.Матрица – ДНҚ тізбегі, яғни РНҚ-полимераза арқылы көшірілуге тиіс ДНҚ
аймағы.
2.РНҚ синтезіне қажетті субстрат – рибонуклеотидтер (УТФ, ГТФ, ЦТФ, АТФ).
3.Энергия көзі – АТФ, ГТФ, ЦТФ, УТФ, олар субстрат ретінде де, энергия көзі
ретінде де пайдаланылады.
4.Ферменттер – РНҚ-полимеразалар, транскрипцияның нәруызды факторлары.
5.Өсуші тізбек – транскрипция кезінде пайда болатын РНҚ молекуласы.
10.
Транскрипцияның принциптері:Комплементарлық: м-РНҚ ДНҚ матрицасына комплементарлы
болады.
Антипараллельдік: РНҚ синтезі ДНҚ-ның қарама-қарсы бағытында
жүреді.
Униполярлылық және бастамасыздық: РНҚ-полимеразаға
праймер қажет емес.
Асимметриялылық: тек бір ДНҚ тізбегі қызмет етеді.
11.
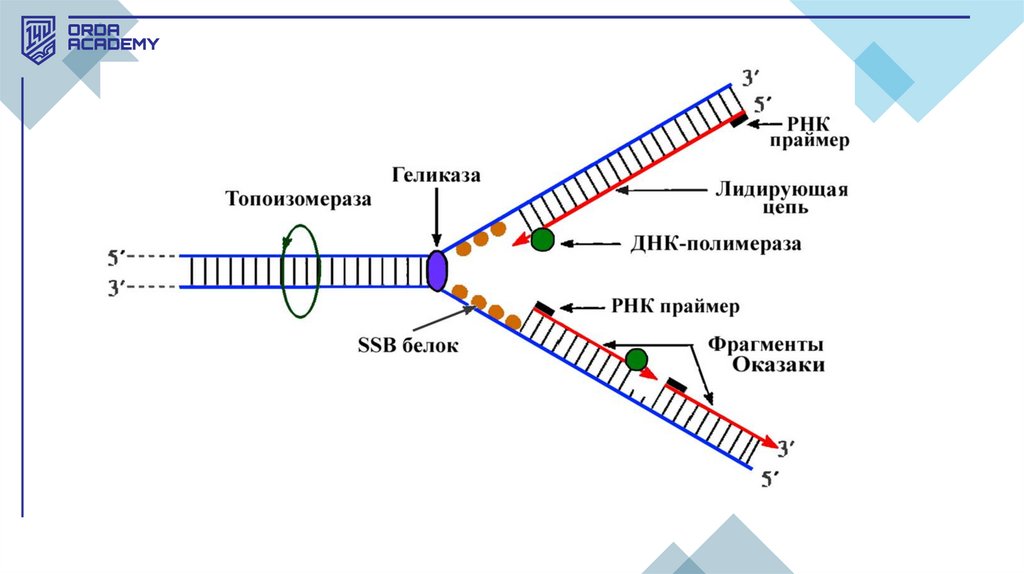
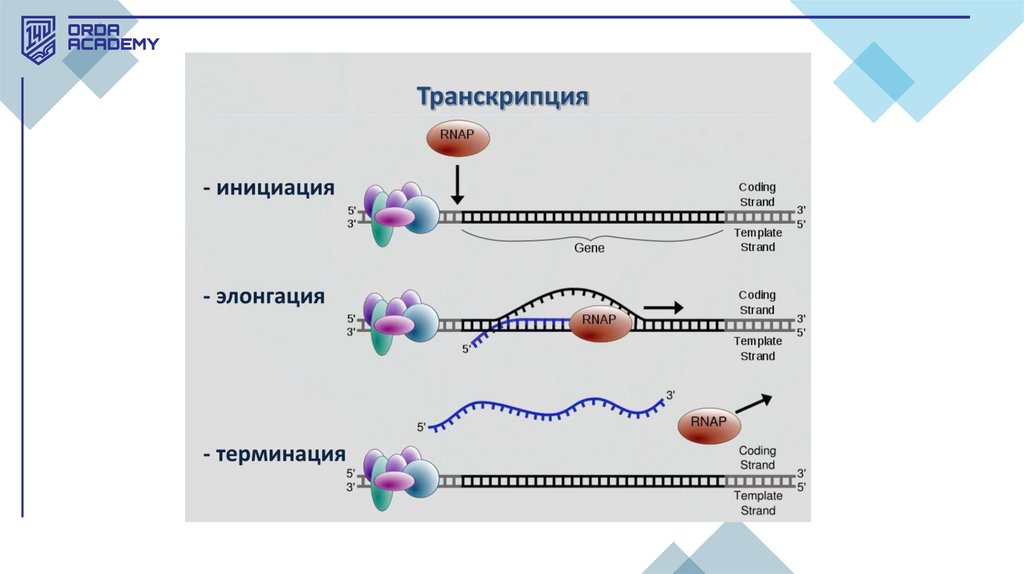
• Транскрипция кезеңдері:1.Инициация (бастапқы кезең): Бұл кезеңде РНҚ-полимераза ДНҚның промотор деп аталатын арнайы аймағымен байланысып, ДНҚ
тізбегін ажыратады. Инициация фазасында РНҚ-полимераза
транскрипцияны бастайтын нүктені таниды, ол жерде м-РНҚ-ның
синтезі басталады. Эукариоттарда инициация барысында қосымша
нәруыздық факторлар қажет.
2.Элонгация (тізбек ұзарту): Бұл кезеңде РНҚ полимераза ДНҚ
матрицалық тізбегінің бойымен қозғалып, комплементарлық ережеге
сай жаңа РНҚ тізбегін синтездейді. ДНҚ-ның бір тізбегі матрица
қызметін атқарады, оған сәйкес рибонуклеотидтер қосылады.
Элонгация кезеңі ДНҚ-ның 5' -> 3' бағытында жүзеге асады.
3.Терминация (аяқталу кезеңі): РНҚ-полимераза ДНҚ-ның
терминатор деп аталатын нүктесіне жеткенде транскрипция процесі
тоқтайды. Терминация кезінде синтезделген РНҚ тізбегі матрицадан
ажырайды.
12.
13.
14.
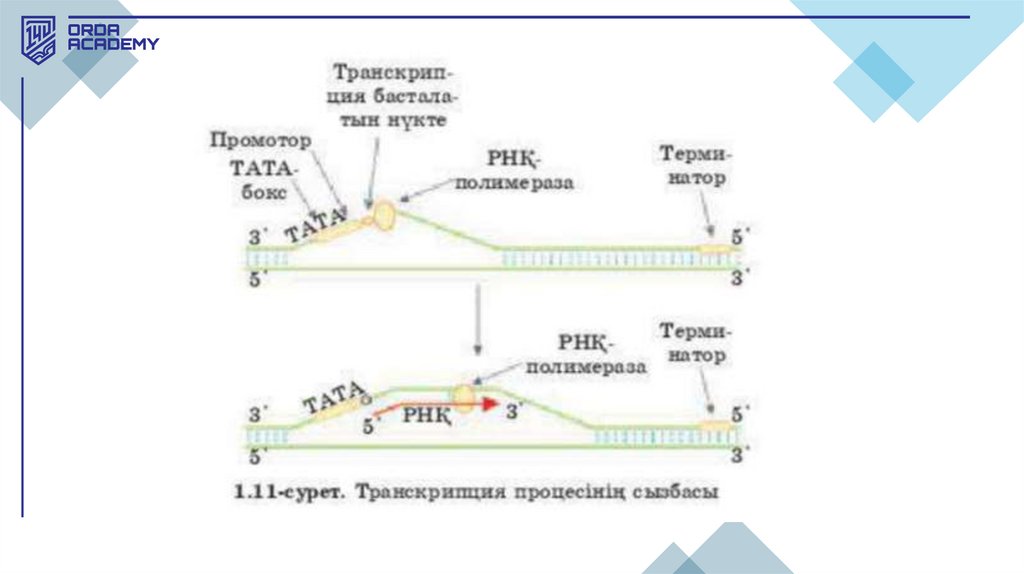
ТАТА-бокс• Промоторда (ДНҚ нуклеотидтерінің реттілігі) транскрипцияның старттық
белгісі ТАТА-бокс болады. ТАТА-бокс инициацияның бірінші факторы
деп ТАТА-факторды байланыстырып алатын ДНҚ нуклеотидтерінің
белгілі бір реттілігін атайды.
• Бұл ТАТА-фактор транскрипцияда үлгі ретінде пайдаланылатын РНҚполимеразаның ДНҚ жіпшесіне (ДНҚ-ның матрицалық жіпшесі) қосылуын
қамтамасыз етеді. Промотор ассиметриялы ("ТАТА") болғандықтан, ол РНҚполимеразаны тек бір бағытта байланыстырады, бұл транскрипцияның 5'үшынан 3'-үшына қарай (5'->3') бағытын анықтайды. РНҚ-полимеразаның
промотормен байланысуы үшін инициацияның тағы бір факторы — сигмафактор керек, бірақ РНҚ-ның бастапқы кесіндісі (ұзындығы 8-10
рибонуклеотид болатын) синтезінен кейін сигма-фактор бірден ферменттен
ажырайды.
15.
16.
• Эукариоттарда транскрипциядан кейін пайда болған м-РНҚ әлі толық қызмет атқаруғадайын емес және бірқатар посттранскрипциялық модификацияларға ұшырайды:
• Кэптүзу (5'-ұшына гуанозин қалдығы қосылады) – бұл процесс м-РНҚ-ның
тұрақтылығын арттырады және оны деградациядан қорғайды.
• Полиаденилдену – м-РНҚ-ның 3'-ұшына поли (А) тізбегі қосылады, бұл да м-РНҚ
тұрақтылығын сақтауға көмектеседі.
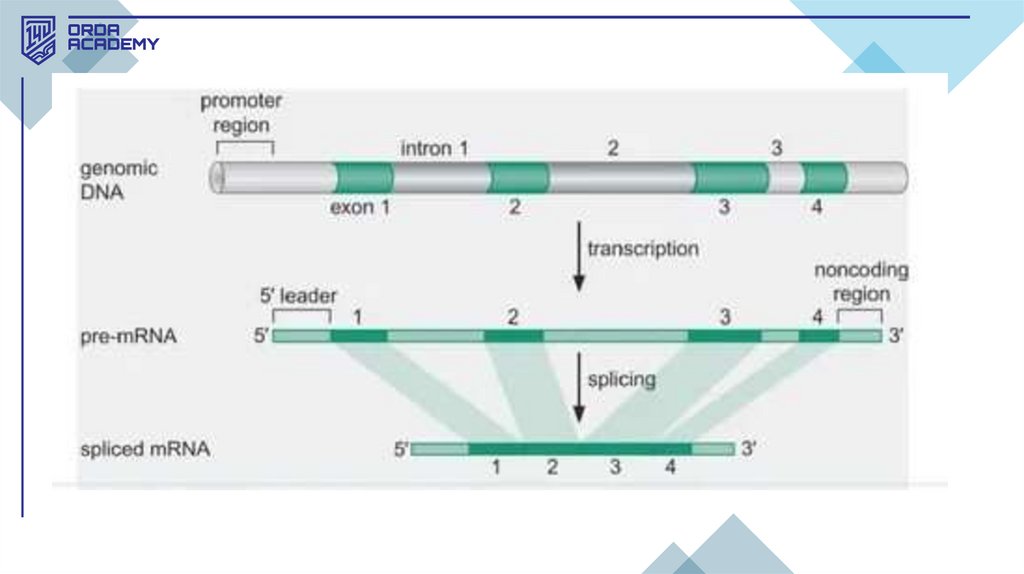
• Сплайсинг – транскрипция кезінде м-РНҚ құрамына интрондар мен экзондар кіреді.
Интрондар – ақпараты жоқ бөліктер, ал экзондар – маңызды генетикалық
ақпаратты қамтиды. Сплайсинг кезінде интрондар кесіліп алынып, экзондар
біріктіріледі, нәтижесінде үздіксіз кодталатын м-РНҚ түзіледі.
17.
18.
Трансляция• Трансляция (лат. translatio – ауысу, аудару) — бұл рибосомада
жүзеге асатын ақпараттық РНҚ (м-РНҚ) матрицасында
аминқышқылдарынан нәруызды синтездеу процесі. Нәруыз синтезі
жасуша тіршілігінің негізі болып саналады. Барлық тірі
организмдердің жасушаларында бұл процесті арнайы органеллалар —
рибосомалар жүзеге асырады.
• Аминқышқылдарды тану үшін арнайы «адаптерлер» —
тасымалдаушы РНҚ (т-РНҚ) қажет. Бұл молекулалар м-РНҚ
кодонына комплементарлы аймақпен (антикодонмен) және сәйкес
аминқышқыл қосылатын аймақпен жабдықталған.
Аминқышқылдардың т-РНҚ-ға қосылуы аминоацил-т-РНҚсинтетазалар деп аталатын энерготәуелді ферменттер арқылы
жүзеге асады. Пайда болған молекула аминоацил-т-РНҚ деп
аталады. Трансляцияның арнайылығы м-РНҚ кодоны мен т-РНҚ
антикодоны арасындағы комплементарлы өзара әрекеттесу арқылы
анықталады.
19.
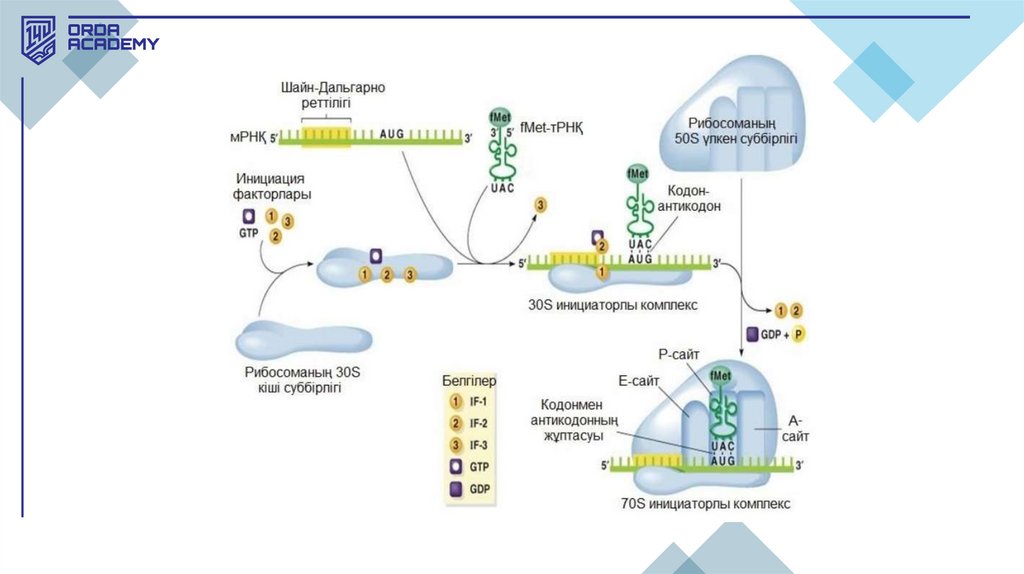
Трансляцияның кезеңдері:1. Инициация:
1. Трансляция метионин аминқышқылын кодтайтын АУГ-кодоннан басталады.
2. Рибосоманың кіші суббірлігі старттық кодонды таниды және метионинді
тасымалдайтын т-РНҚ осы кодонға қосылады. Одан кейін рибосоманың үлкен суббірлігі
қосылып, нәруыз синтезі басталады.
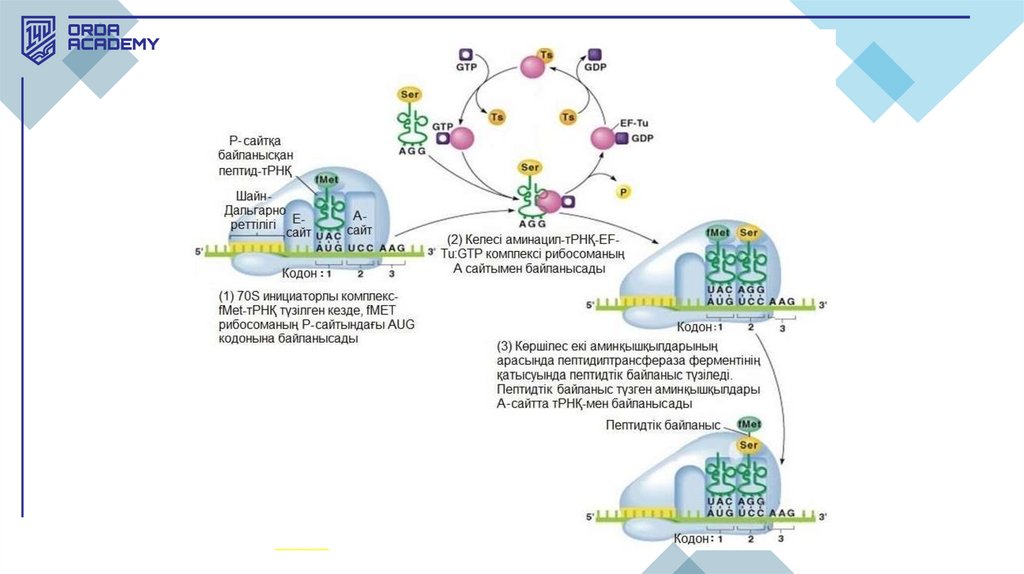
2.Элонгация:
1. Рибосома м-РНҚ-ның бойымен жылжып, т-РНҚ арқылы аминқышқылдарды
кодондарға сәйкес қосып отырады.
2. Пептидтік байланыстар арқылы аминқышқылдары бірігеді, нәтижесінде нәруыздың
тізбегі ұзартылады.
3. Әр циклде рибосома м-РНҚ бойымен бір триплетке жылжып, жаңа кодонды оқып
шығады.
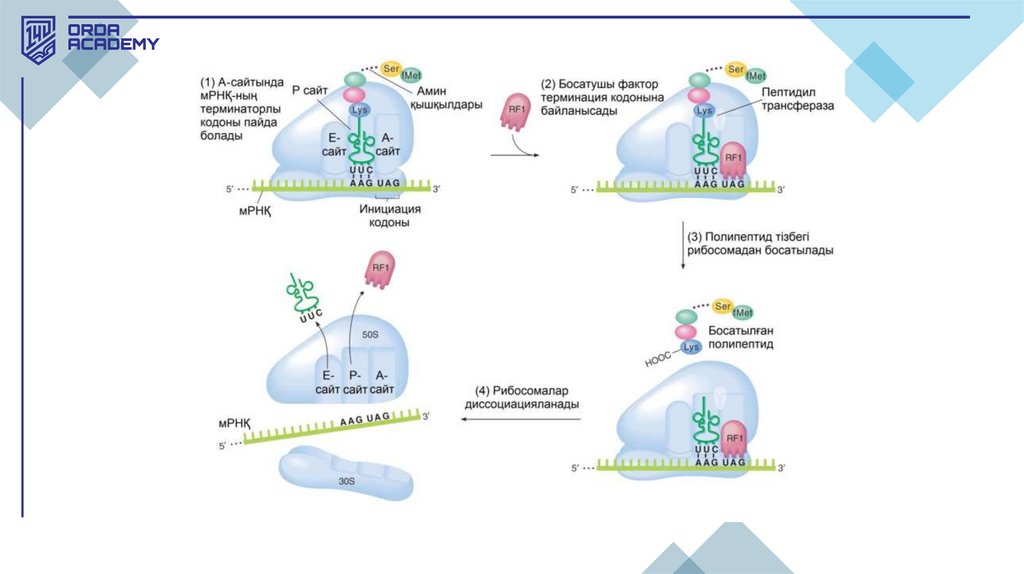
3.Терминация:
1. Терминация рибосома м-РНҚ тізбегінде стоп-кодон (UAA, UAG, UGA) кездескен кезде
жүреді.
2. Бұл кодондарға сәйкес т-РНҚ жоқ болғандықтан, релиз факторлары полипептид
тізбегінің босатылуын катализдейді.
20.
21.
• Элонгация. Полипептидті тізбекті өсіру процесіне элонгацияның екі нәруызфакторы қатысады. Бірінші нәруыз факторы (эукариоттарда — EF-a,
прокариоттарда — EF-Tu) аминоацилденген ("зарядталған" аминқышқылы) тРНҚ-ны рибосомаға көшіреді. Рибосома т-РНҚ-мен байланысқан пептидтің
тасымалын және ондағы аминқышқылы қалдығымен пептидті байланыс түзуін
катализдейді. Осылайша өсіп келе жатқан пептид тағы бір аминқышқылы
қалдығына ұзарады. Содан соң екінші нәруыз транслокацияны катализдейді.
Транслокация рибосоманың м-РНҚ бойымен бір триплетке жылжуы, осыдан
кейін рибосома элонгацияның жаңа цикліне дайын болады.
22.
23.
• Нәруыз синтезінің аяқталуы рибосоманың А-сайтында УАГ, УАА, УГА — стопкодондарының (триплеттердің) бірі болғанда жүзеге асырылады. Бұлкодондарға сәйкес т-РНҚ жоқ болғандықтан, пептидил-т-РНҚ рибосоманың Рсайтымен байланысқан күйі қалады. Бұл жерде полипептидті тізбектің м-РНҚдан ажырауын катализдейтін RF1 немесе RF2, сонымен қатар рибосомадан мРНҚ-ның диссоциациялануын тудыратын RF3 арнайы нәруыздары әрекетке
түседі. RF1 A-аймақта УАА немесе УАГ таниды; ал RF2 УАА немесе УГА таниды.
Басқа стоп-кодондарға қарағанда, УАА-мен терминация тиімдірек болады.
24.
25.
26.
• Оқу шеңберлері. Әр кодон үш нуклеотидтен тұратын болғандықтан, бір ғанагенетикалық мәтінді үш әртүрлі тәсілмен оқуға болады (бірінші, екінші, үшінші
нуклеотидтерден бастап), яғни оқуды үш шеңберде жүргізуге болады.
• Қазақстандағы молекулалық биологияның негізін қалаушы, академик,
профессор Мұрат Әбенұлы Айтхожин әлемдік ғылымда алғашқылардың
бірі болып жоғары сатылы организмдерде, соның ішінде өсімдіктердегі нәруыз
синтездеуші аппараттарға салыстырмалы зерттеу жүргізді және оның зерттеу
тобы жоғары молекулалық (рибосомалық емес) рибонуклеин қышқылынан
тұратын бөлшектер өсімдіктер информасома класын ашты.
27.
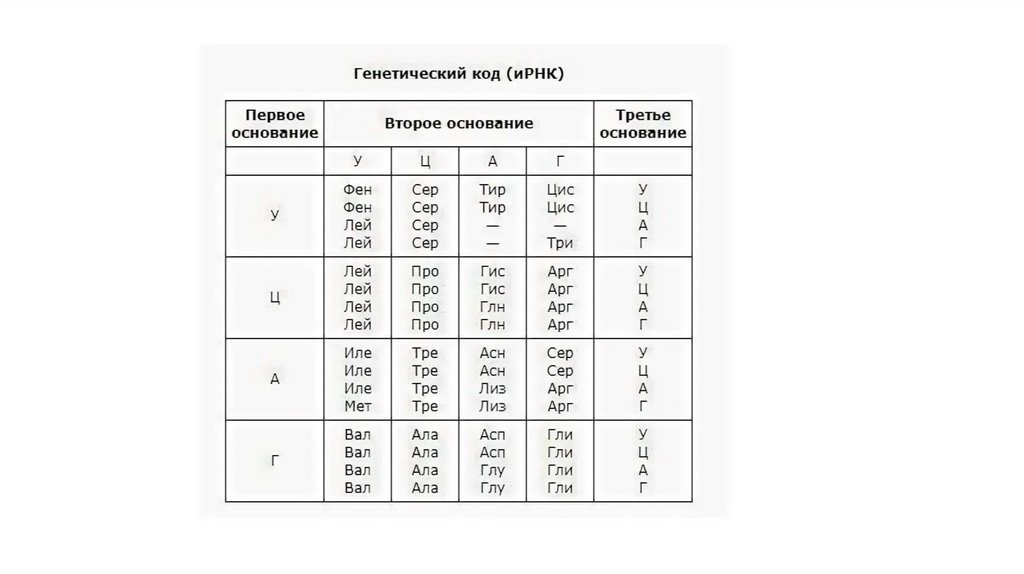
Генетикалық код• Ген — тұқым қуалау ақпаратының бірлігі. Адамда шамамен 25-30 мың ген
болады.
• ГЕН:
Реттеуші: Ақпараттың оқылуын белсендіреді немесе басады.
Құрылымдық: Нәруыздың біріншілік құрылымын кодтайды, рРНҚ және тРНҚ.
• Нәруыздың біріншілік құрылымы туралы ақпарат ДНҚ молекуласының
аймағында генде нуклеотидтердің реттілігі түрінде кодталған. Әрбір ДНҚ
молекуласында көптеген гендер болады. Организмдегі барлық гендер
жиынтығы генотипті құрайды. Тұқым қуалау ақпаратының кодталуы
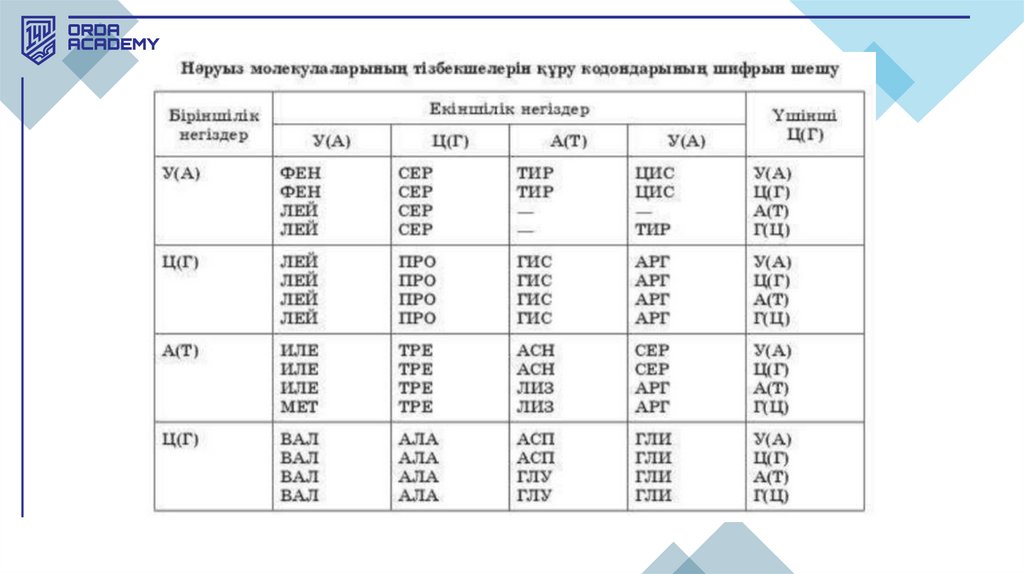
генетикалық код көмегімен жүзеге асырылады.
28.
• 1961 жылы Ниренберг және Маттеи ең алғаш рет код терминінтүсіндірген. Олар ішек таяқшасынан нәруыз синтезіне қажетті рибосомалар
және басқа да заттары бар экстракт алды. Ортаға қажетті м-РНҚ қосқанда,
нәруыз синтезі үшін қажет жасушасыз жүйе пайда болды. Тек урацилдерден
тұратын ортаға синтетикалық РНҚ-ны қосқанда, олар тек фенилаланиннен
(полифенилаланин) тұратын нәруыз пайда болғанын анықтады. Осылайша
УУУ нуклеотидтер триплеті (кодон) фенилаланинге сәйкес келетіні
анықталды.
29.
30.
31.
32.
Генетикалық кодтың қасиеттері.1.Үшөрімділік (триплеттілік) - әр аминқышқылына нуклеотидтердің үштігі сәйкес
келеді. 4^3 = 64 кодон бар екенін есептеу қиынға соқпайды. Олардың 61-інің мәні бар,
ал 3-еуі мәнсіз (терминирациялаушы, стоп-кодондар) болып келеді.
2.Гендердің арасындағы тыныс белгілері - стоп-кодондар деп аталатын
триплеттер болады. Олар бір полипептидті тізбектің синтезі аяқталғаны жөнінде белгі
беріп отырады.
3.Үзіліссіздік (нуклеотидтер арасында бөлгіш белгілер жоқ) - генішілік тыныс
белгілерінің болмауы, ген ішінде әр нуклеотид белгіленетін кодонның құрамына
кіреді. 1961 жылы Сеймур Бензер мен Френсис Крик эксперимент жүзінде кодтың
триплеттілігін және оның үзіліссіз (ықшамдылық) екенін дәлелдеді.
4.Коллинеарлылық - м-РНҚ кодондары мен нәруыздағы аминқышқылдарының
сызықтық реттілігінің сәйкестігі.
33.
Генетикалық кодтың қасиеттері.5. Арнайылылық - әр аминқышқылына тек белгілі бір, басқа аминқышқылы үшін
қолдана алмайтын кодондар сәйкес келеді.
6. Бірбағыттылық - кодоны бір бағытта - бірінші нуклеотидтен бастап келесілерге
қарай оқылады.
7. Артықтық - бір аминқышқылын бірнеше триплет кодтай алады (аминқышқылы 20, мүмкін болатын триплеттер - 64, оның ішінде 61 мәні бар, яғни орташа алғанда әр
аминқышқылына 3 кодон сәйкес келеді); тек метионин (Met) мен триптофан (Trp)
ережеден тыс болады.
8. Әмбебаптық – генетикалық кодтың жоғарыда аталған қасиеттерінің барлығы тірі
организмдерге тән болып келеді.
34.
Эпигенетика• Эпигенетика — биология ғылымының салыстырмалы түрде жаңа бағыты,
генетика сияқты кең танылмаған. Бұл сала организмнің дамуы немесе
жасушаның бөлінуі кезінде гендердің белсенділігінің тұқым қуалайтын
өзгерістерін зерттейді. Эпигенетикалық өзгерістер кезінде ДНҚ
нуклеотидтерінің реттілігі өзгермейді.
• 1942 жылы ағылшын генетигі Waddington "эпигенетика" терминін
енгізді. Генетикалық және эпигенетикалық тұқым қуалау механизмдерінің
айырмашылығы олардың әсерінің ұзақтығына және тұрақтылығына
байланысты. Генетикалық мутациялар тұрақты және шексіз қайталанады, ал
эпигенетикалық модификациялар уақытша және бірнеше ұрпақ ішінде ғана
сақталады.
35.
36.
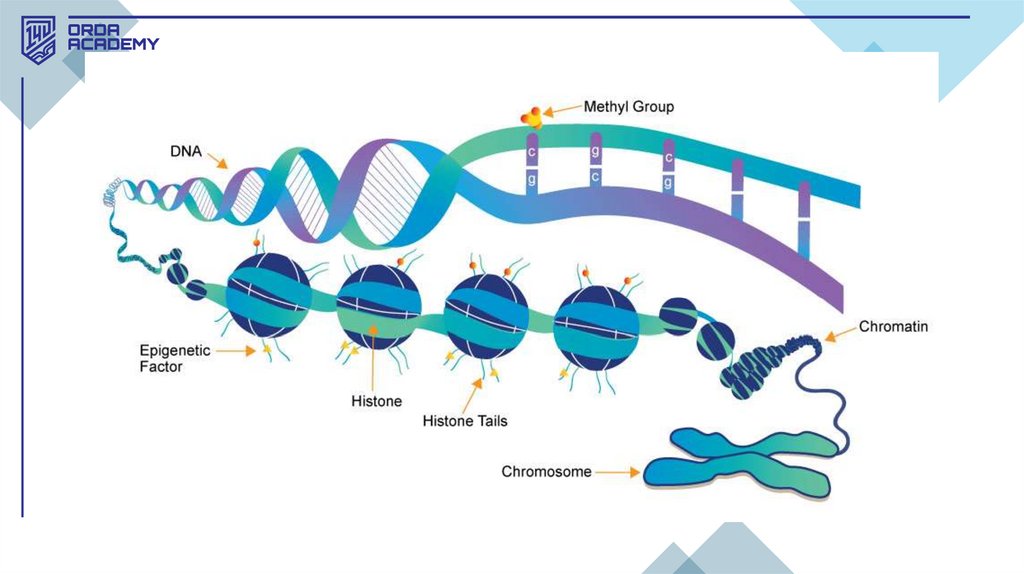
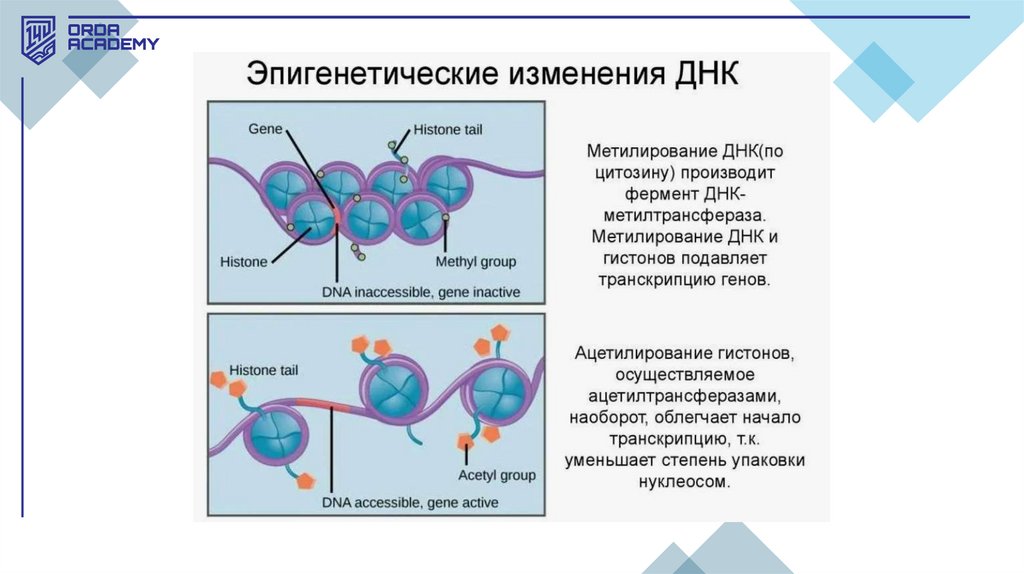
• Эпигенетикалық механизмдер хроматиннің қызметін реттеуде маңыздырөл атқарады, бұл гистондардың модификациясы және ДНҚ метилденуі
арқылы жүзеге асады. Соңғы екі онжылдықта эукариоттардағы транскрипция
механизмдері туралы түсінік өзгерді. Гистондардың трансляцияны реттеудегі
рөлі анықталып, "гистон коды" ашылды, мұнда гистон модификациясы
гендерді белсендендіру немесе репрессияға ұшырату мүмкін.
• Эпигенетикалық маркерлердің жиынтығы эпиген деп аталады, және олар
фенотипке әсер ете алады.
• Эпигенетика тәжірибелік медицинада да кеңінен қолданылады.
Эпигенетикалық реттелудің бұзылуы көптеген аурулардың себебі болуы
мүмкін. Мұндай аурулар гендік және хромосомдық импринтингке байланысты
24 түрлі нозологияға бөлінеді. Гендік импринтинг ауруларына, мысалы,
Прадера-Вилли синдромы, Ангельман синдромы және БеквиттВидеман синдромы жатады.
37.
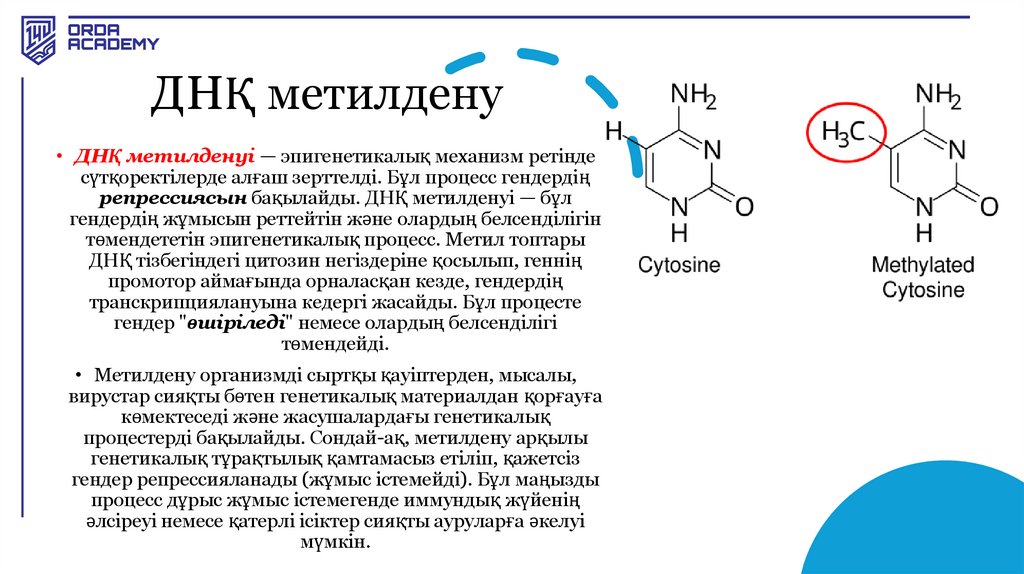
ДНҚ метилдену• ДНҚ метилденуі — эпигенетикалық механизм ретінде
сүтқоректілерде алғаш зерттелді. Бұл процесс гендердің
репрессиясын бақылайды. ДНҚ метилденуі — бұл
гендердің жұмысын реттейтін және олардың белсенділігін
төмендететін эпигенетикалық процесс. Метил топтары
ДНҚ тізбегіндегі цитозин негіздеріне қосылып, геннің
промотор аймағында орналасқан кезде, гендердің
транскрипциялануына кедергі жасайды. Бұл процесте
гендер "өшіріледі" немесе олардың белсенділігі
төмендейді.
• Метилдену организмді сыртқы қауіптерден, мысалы,
вирустар сияқты бөтен генетикалық материалдан қорғауға
көмектеседі және жасушалардағы генетикалық
процестерді бақылайды. Сондай-ақ, метилдену арқылы
генетикалық тұрақтылық қамтамасыз етіліп, қажетсіз
гендер репрессияланады (жұмыс істемейді). Бұл маңызды
процесс дұрыс жұмыс істемегенде иммундық жүйенің
әлсіреуі немесе қатерлі ісіктер сияқты ауруларға әкелуі
мүмкін.
38.
• Гистондардың ацетилденуі — бұл ДНҚ мен гистондардың байланысынәлсіретіп, гендердің белсенді болуына мүмкіндік беретін процесс. Бұл өзгеріс
гистондарға ацетил топтарының қосылуы арқылы жүреді, осылайша ДНҚ-ның
кейбір бөліктері ашылып, олармен жұмыс істеуге мүмкіндік береді. Гистондық
модификациялар қысқа мерзімді болуы мүмкін және жасушаның қажеттілігіне
қарай өзгереді.
• Эпигенетикалық өзгерістер — бұл ДНҚ-ның реттілігі өзгермей, гендердің
белсенділігі өзгерген кезде болатын құбылыс. Бұл өзгерістер ұрпақтан-ұрпаққа
берілуі мүмкін, бірақ бірнеше ұрпақтан кейін жоғалып кетуі де ықтимал.