


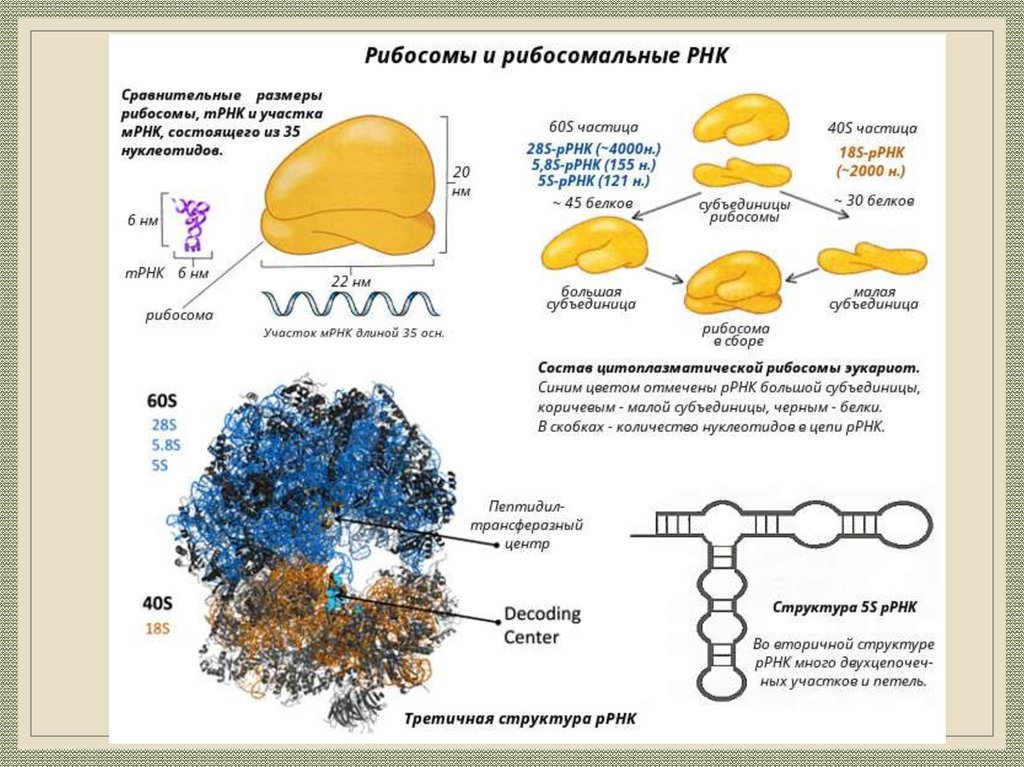

 Биология
БиологияПохожие презентации:

")








Механизм функционирования внутриклеточных систем
1.
ЛекцияМеханизм функционирования внутриклеточных
систем
1.10. Клеточные механизмы считывания генома. Генетический код.
1.10.1. Репликация ДНК.
1.10.2. Репарация и рекомбинация ДНК.
1.11. Матричный синтез: понятие, принципы реализации
генетического материала.
1.11.1. Транскрипция.
1.11.2. Трансляция.
Н.И. Петруняк
2.
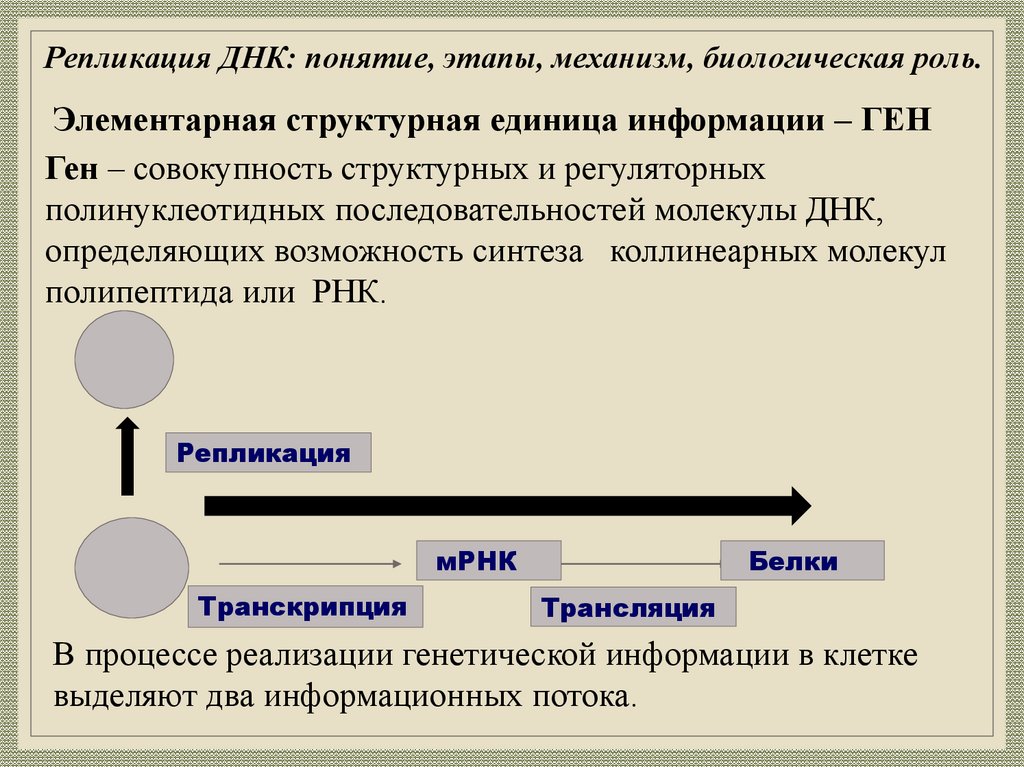
Репликация ДНК: понятие, этапы, механизм, биологическая роль.Элементарная структурная единица информации – ГЕН
Ген – совокупность структурных и регуляторных
полинуклеотидных последовательностей молекулы ДНК,
определяющих возможность синтеза коллинеарных молекул
полипептида или РНК.
Репликация
мРНК
Транскрипция
Белки
Трансляция
В процессе реализации генетической информации в клетке
выделяют два информационных потока.
3.
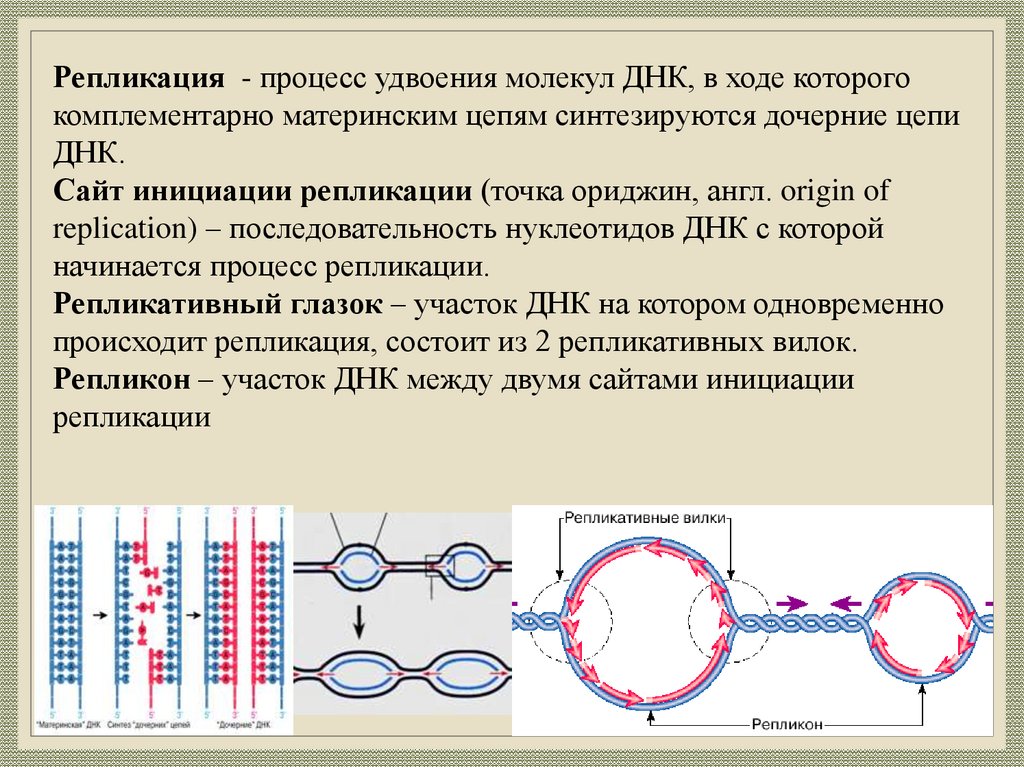
Репликация - процесс удвоения молекул ДНК, в ходе которогокомплементарно материнским цепям синтезируются дочерние цепи
ДНК.
Сайт инициации репликации (точка ориджин, англ. origin of
replication) – последовательность нуклеотидов ДНК с которой
начинается процесс репликации.
Репликативный глазок – участок ДНК на котором одновременно
происходит репликация, состоит из 2 репликативных вилок.
Репликон – участок ДНК между двумя сайтами инициации
репликации
4.
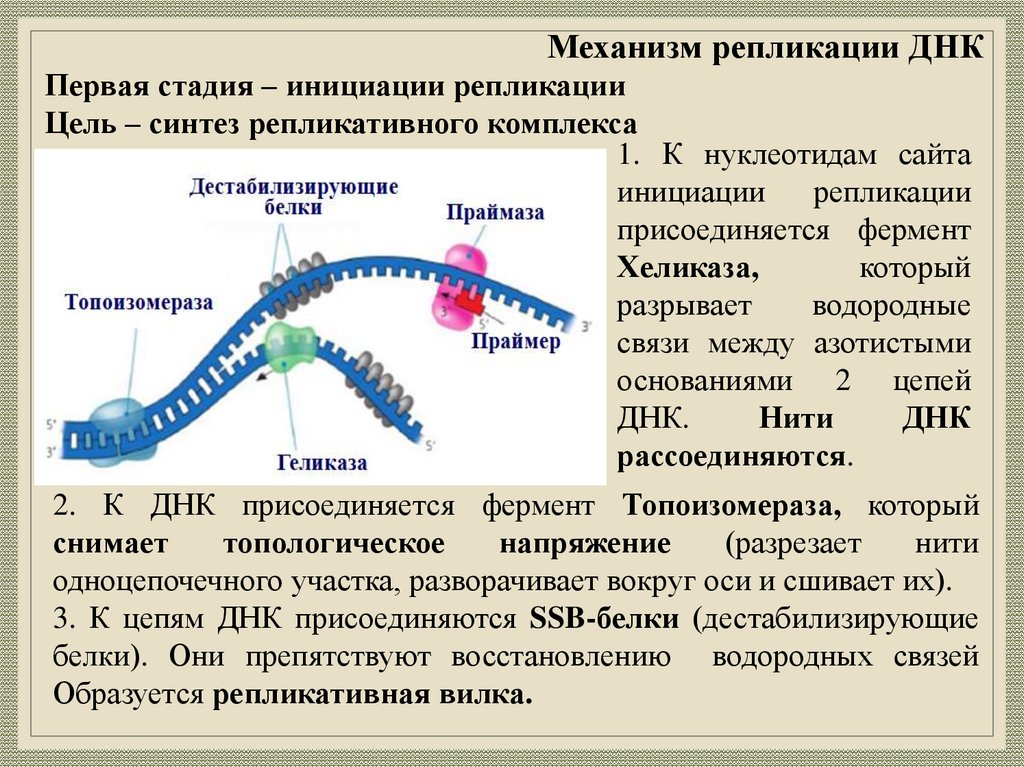
Механизм репликации ДНКПервая стадия – инициации репликации
Цель – синтез репликативного комплекса
1. К нуклеотидам сайта
инициации
репликации
присоединяется фермент
Хеликаза,
который
разрывает
водородные
связи между азотистыми
основаниями 2 цепей
ДНК.
Нити
ДНК
рассоединяются.
2. К ДНК присоединяется фермент Топоизомераза, который
снимает
топологическое
напряжение
(разрезает
нити
одноцепочечного участка, разворачивает вокруг оси и сшивает их).
3. К цепям ДНК присоединяются SSB-белки (дестабилизирующие
белки). Они препятствуют восстановлению водородных связей
Образуется репликативная вилка.
5.
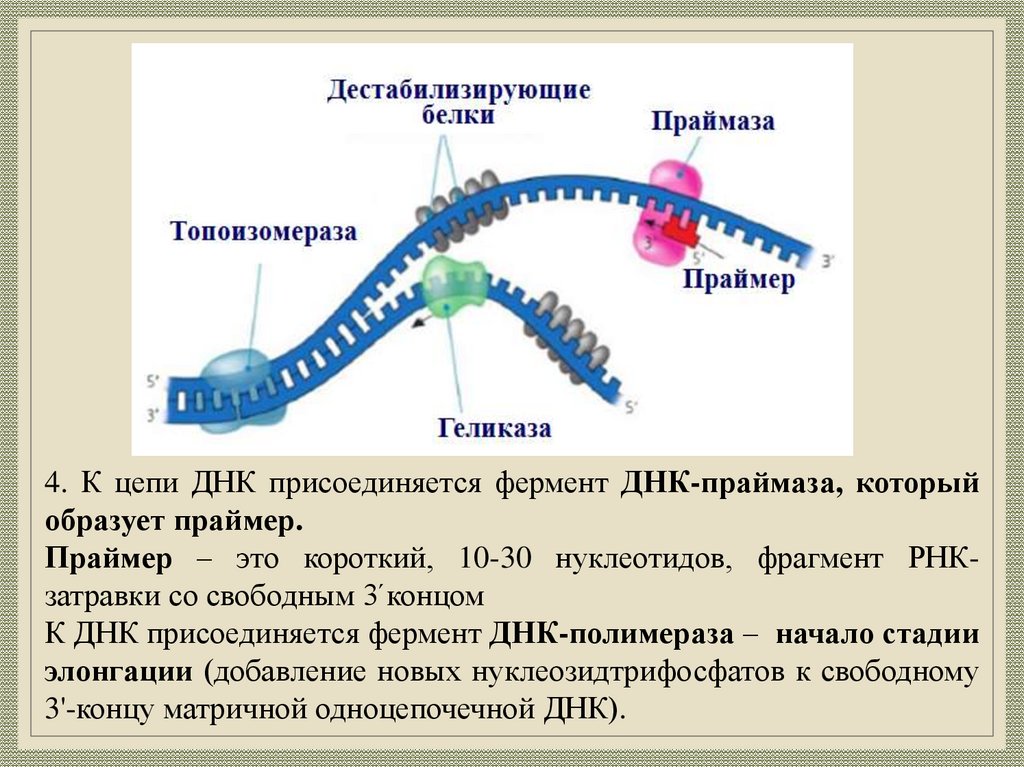
4. К цепи ДНК присоединяется фермент ДНК-праймаза, которыйобразует праймер.
Праймер – это короткий, 10-30 нуклеотидов, фрагмент РНКзатравки со свободным 3́ концом
К ДНК присоединяется фермент ДНК-полимераза – начало стадии
элонгации (добавление новых нуклеозидтрифосфатов к свободному
3'-концу матричной одноцепочечной ДНК).
6.
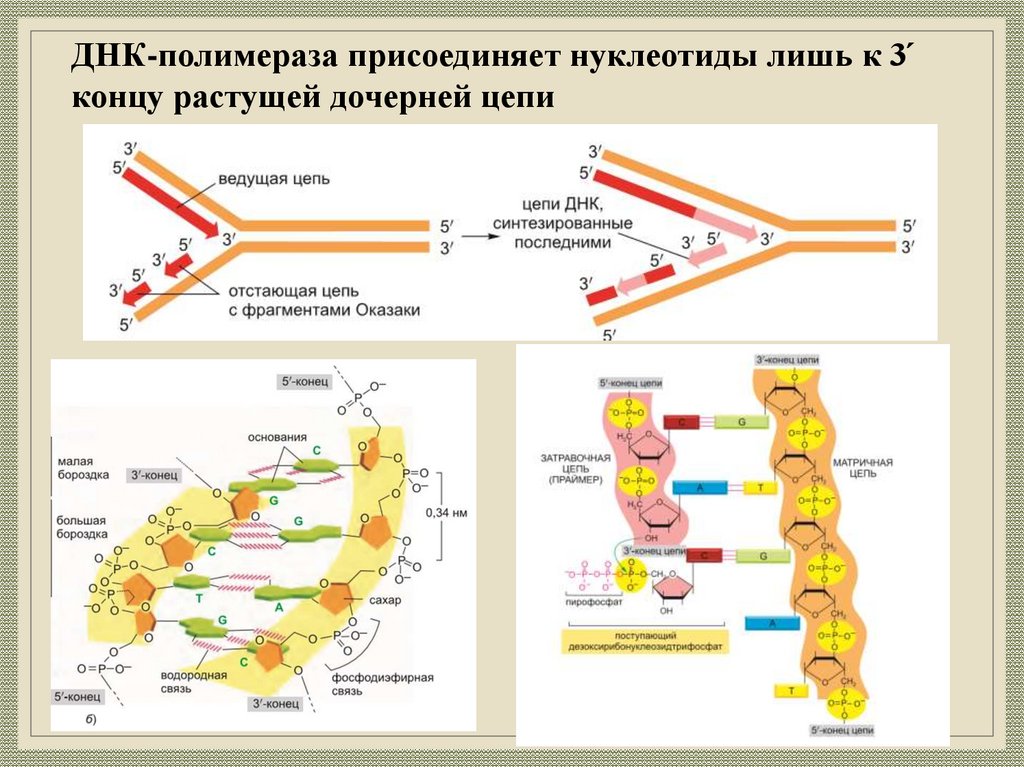
ДНК-полимераза присоединяет нуклеотиды лишь к 3́концу растущей дочерней цепи
7.
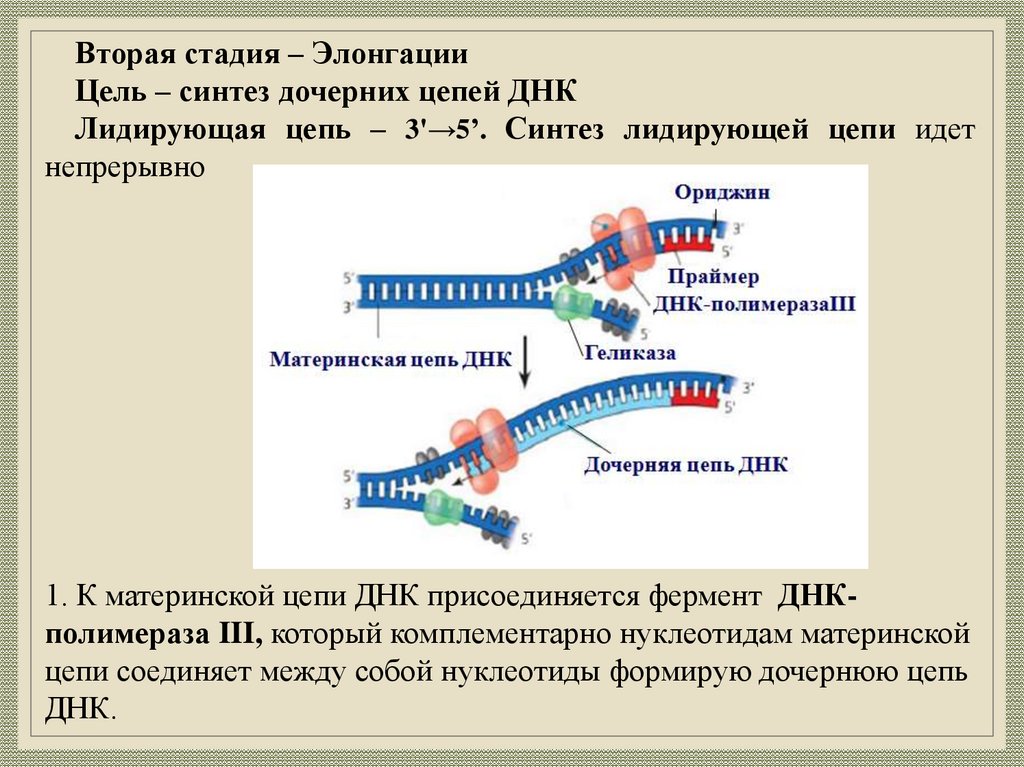
Вторая стадия – ЭлонгацииЦель – синтез дочерних цепей ДНК
Лидирующая цепь – 3'→5’. Синтез лидирующей цепи идет
непрерывно
1. К материнской цепи ДНК присоединяется фермент ДНКполимераза III, который комплементарно нуклеотидам материнской
цепи соединяет между собой нуклеотиды формирую дочернюю цепь
ДНК.
8.
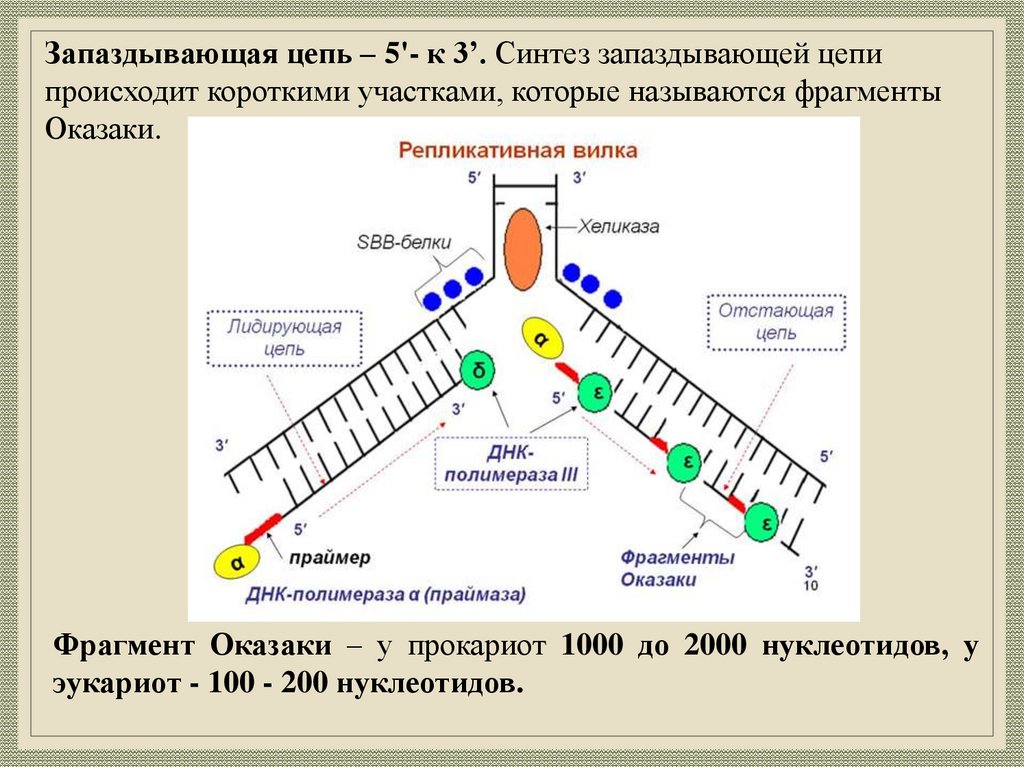
Запаздывающая цепь – 5'- к 3’. Синтез запаздывающей цепипроисходит короткими участками, которые называются фрагменты
Оказаки.
Фрагмент Оказаки – у прокариот 1000 до 2000 нуклеотидов, у
эукариот - 100 - 200 нуклеотидов.
9.
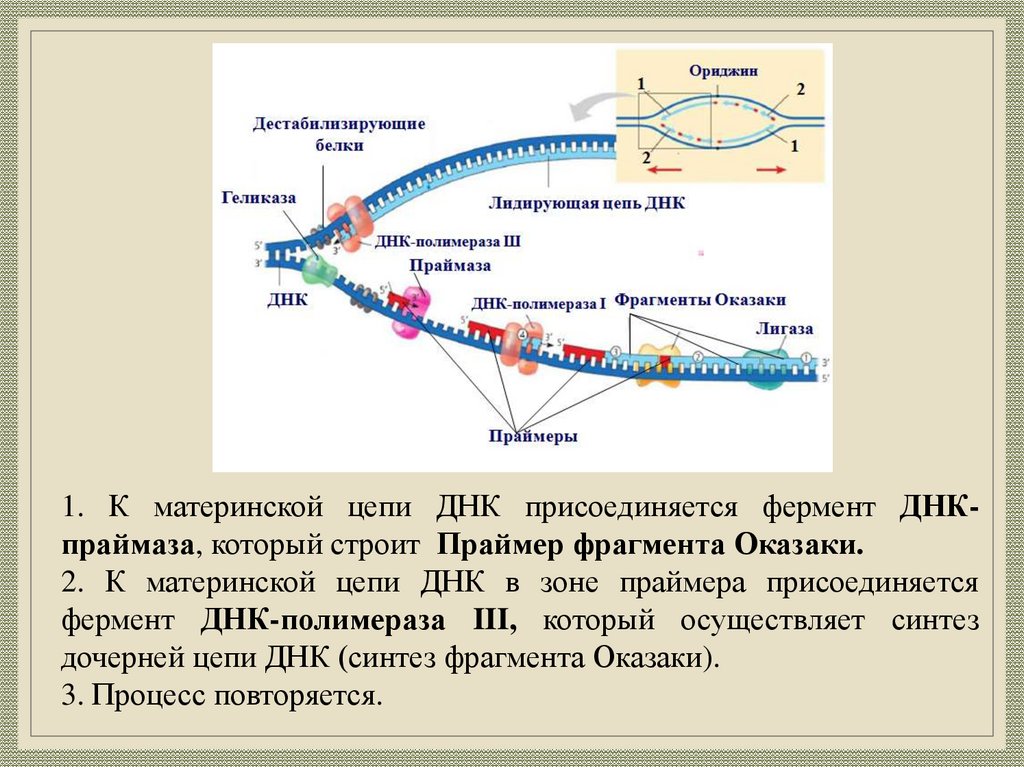
1. К материнской цепи ДНК присоединяется фермент ДНКпраймаза, который строит Праймер фрагмента Оказаки.2. К материнской цепи ДНК в зоне праймера присоединяется
фермент ДНК-полимераза III, который осуществляет синтез
дочерней цепи ДНК (синтез фрагмента Оказаки).
3. Процесс повторяется.
10.
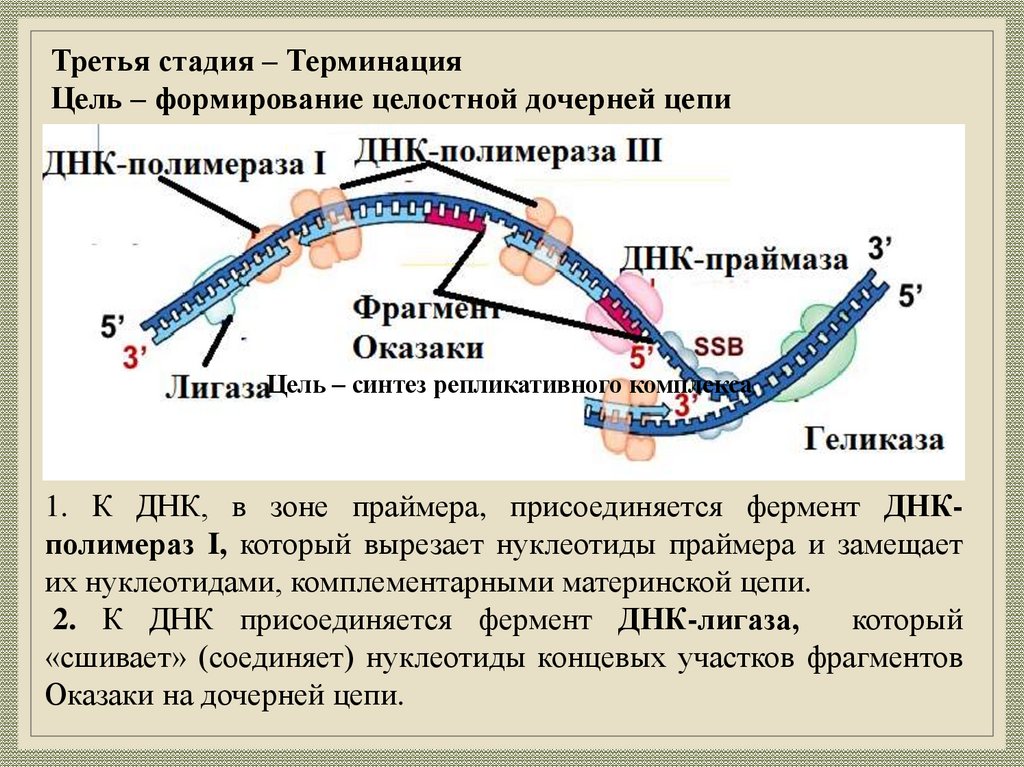
Третья стадия – ТерминацияЦель – формирование целостной дочерней цепи
Цель – синтез репликативного комплекса
1. К ДНК, в зоне праймера, присоединяется фермент ДНКполимераз I, который вырезает нуклеотиды праймера и замещает
их нуклеотидами, комплементарными материнской цепи.
2. К ДНК присоединяется фермент ДНК-лигаза,
который
«сшивает» (соединяет) нуклеотиды концевых участков фрагментов
Оказаки на дочерней цепи.
11.
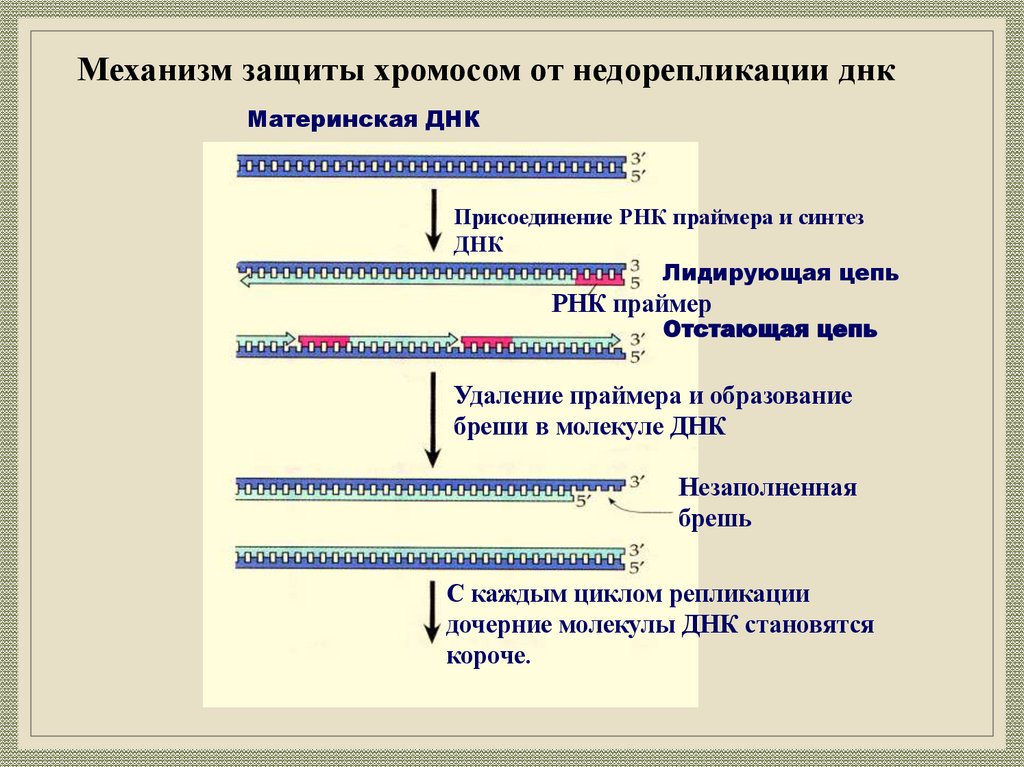
Механизм защиты хромосом от недорепликации днкМатеринская ДНК
Присоединение РНК праймера и синтез
ДНК
Лидирующая цепь
РНК праймер
Отстающая цепь
Удаление праймера и образование
бреши в молекуле ДНК
Незаполненная
брешь
С каждым циклом репликации
дочерние молекулы ДНК становятся
короче.
12.
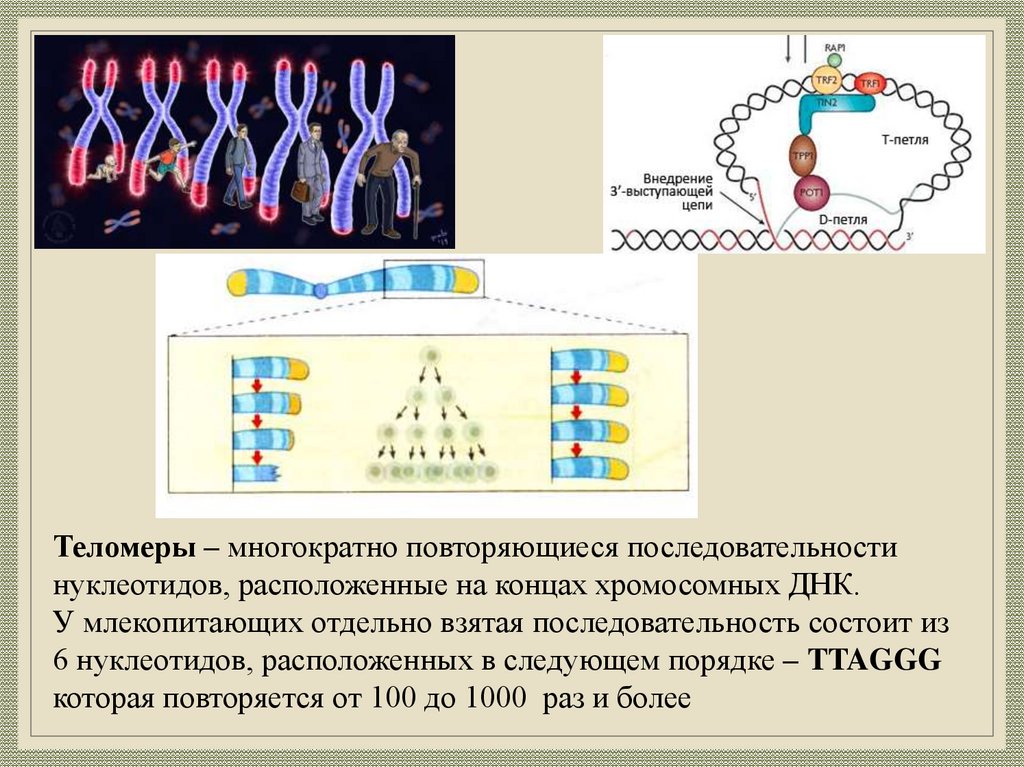
Теломеры – многократно повторяющиеся последовательностинуклеотидов, расположенные на концах хромосомных ДНК.
У млекопитающих отдельно взятая последовательность состоит из
6 нуклеотидов, расположенных в следующем порядке – TTAGGG
которая повторяется от 100 до 1000 раз и более
13.
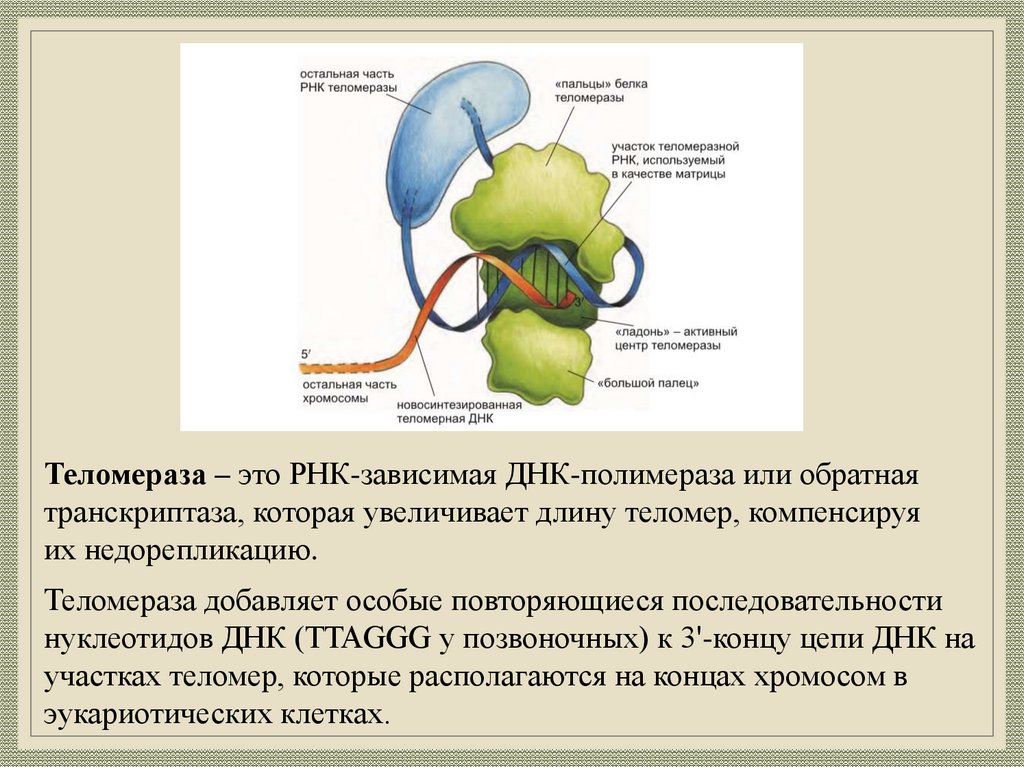
Теломераза – это РНК-зависимая ДНК-полимераза или обратнаятранскриптаза, которая увеличивает длину теломер, компенсируя
их недорепликацию.
Теломераза добавляет особые повторяющиеся последовательности
нуклеотидов ДНК (TTAGGG у позвоночных) к 3'-концу цепи ДНК на
участках теломер, которые располагаются на концах хромосом в
эукариотических клетках.
14.
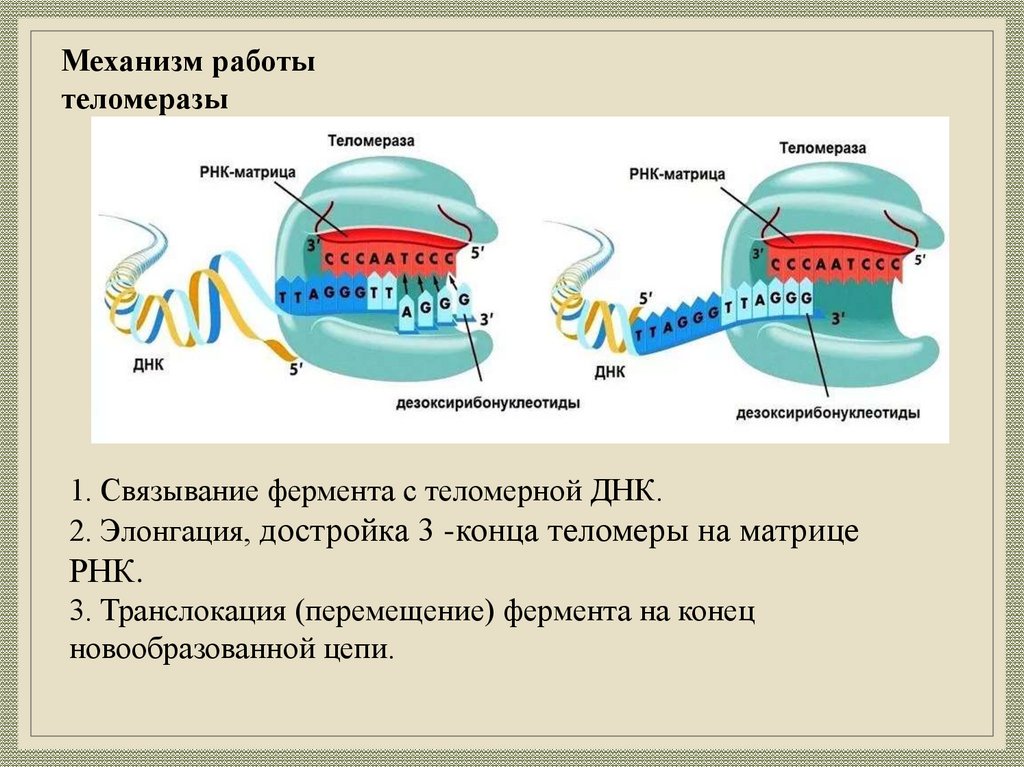
Механизм работытеломеразы
1. Связывание фермента с теломерной ДНК.
2. Элонгация, достройка 3 -конца теломеры на матрице
РНК.
3. Транслокация (перемещение) фермента на конец
новообразованной цепи.
15.
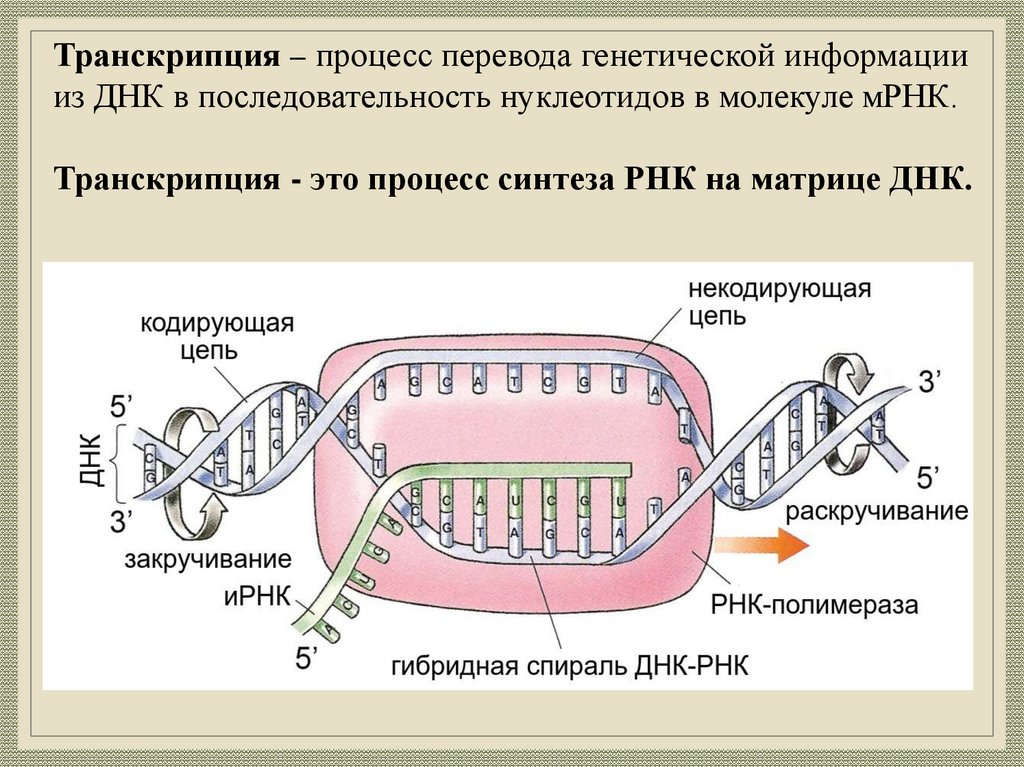
Транскрипция – процесс перевода генетической информациииз ДНК в последовательность нуклеотидов в молекуле мРНК.
Транскрипция - это процесс синтеза РНК на матрице ДНК.
16.
Первая стадия – инициация транскрипцииI. Инициация транскрипции прокариот
1. Сигма-фактор (σ-фактор) движется вдоль молекулы ДНК,
распознает регуляторную последовательность (CAP site),
которая указывает на начало промотора.
2. Сигма-фактор присоединяет к промотору фермент РНКполимеразу I и отсоединяется от ДНК.
17.
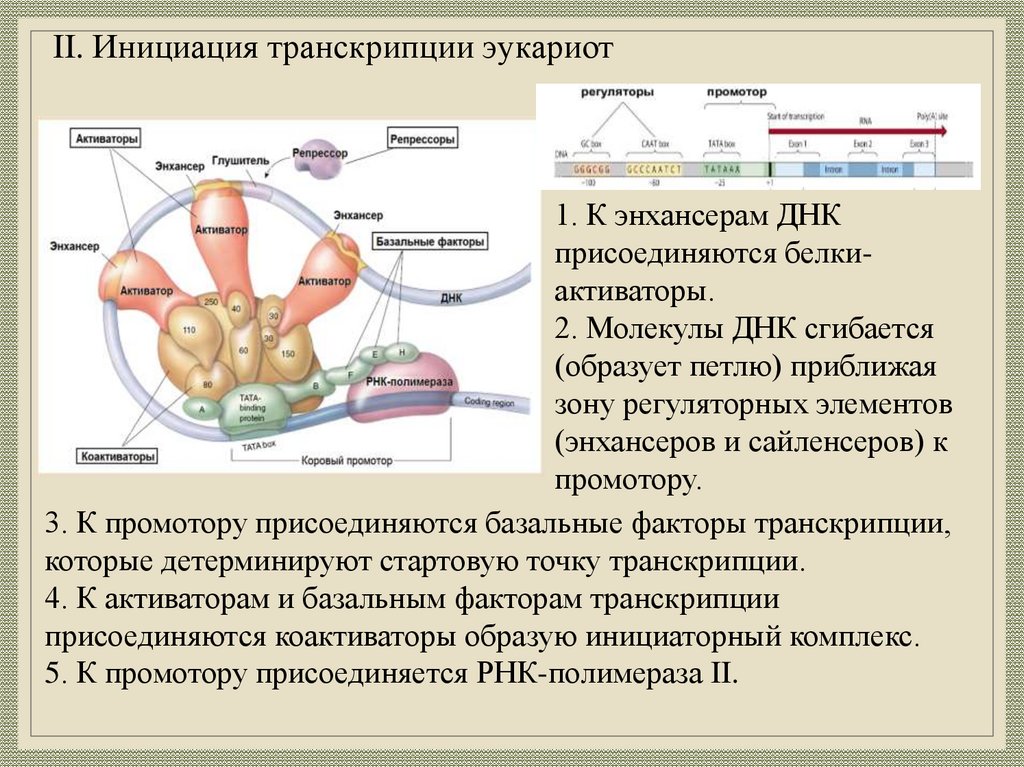
II. Инициация транскрипции эукариот1. К энхансерам ДНК
присоединяются белкиактиваторы.
2. Молекулы ДНК сгибается
(образует петлю) приближая
зону регуляторных элементов
(энхансеров и сайленсеров) к
промотору.
3. К промотору присоединяются базальные факторы транскрипции,
которые детерминируют стартовую точку транскрипции.
4. К активаторам и базальным факторам транскрипции
присоединяются коактиваторы образую инициаторный комплекс.
5. К промотору присоединяется РНК-полимераза II.
18.
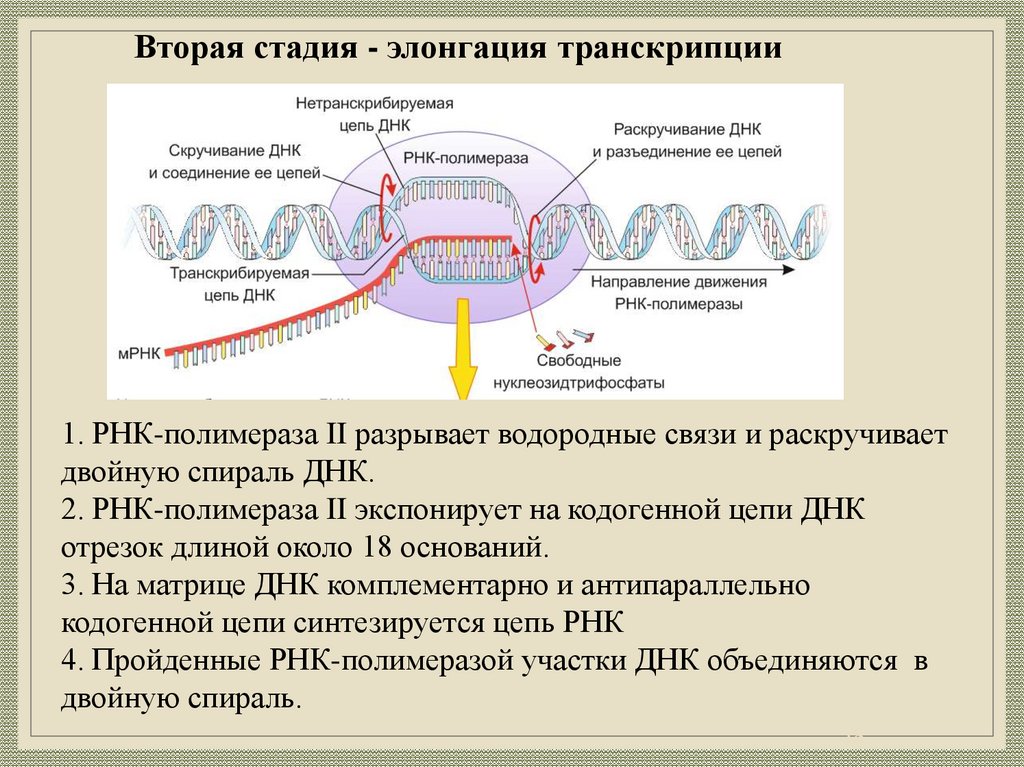
Вторая стадия - элонгация транскрипции1. РНК-полимераза II разрывает водородные связи и раскручивает
двойную спираль ДНК.
2. РНК-полимераза II экспонирует на кодогенной цепи ДНК
отрезок длиной около 18 оснований.
3. На матрице ДНК комплементарно и антипараллельно
кодогенной цепи синтезируется цепь РНК
4. Пройденные РНК-полимеразой участки ДНК объединяются в
двойную спираль.
18
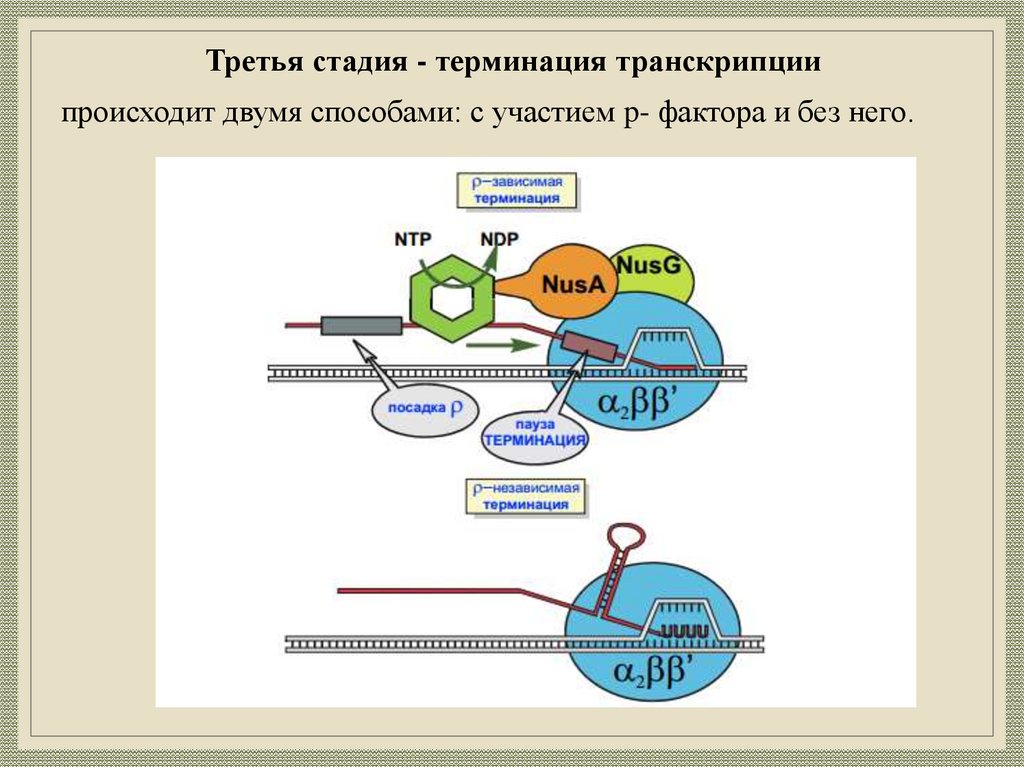
19.
Третья стадия - терминация транскрипциипроисходит двумя способами: с участием p- фактора и без него.
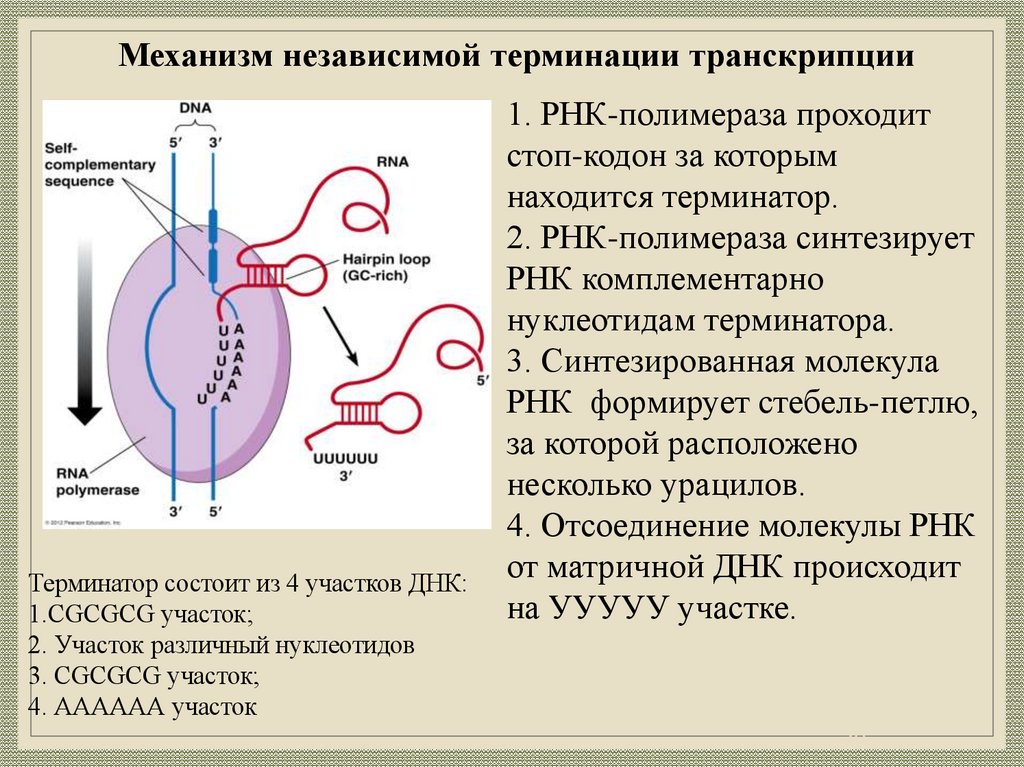
20.
Механизм независимой терминации транскрипцииТерминатор состоит из 4 участков ДНК:
1.CGCGCG участок;
2. Участок различный нуклеотидов
3. CGCGCG участок;
4. АААААА участок
1. РНК-полимераза проходит
стоп-кодон за которым
находится терминатор.
2. РНК-полимераза синтезирует
РНК комплементарно
нуклеотидам терминатора.
3. Синтезированная молекула
РНК формирует стебель-петлю,
за которой расположено
несколько урацилов.
4. Отсоединение молекулы РНК
от матричной ДНК происходит
на УУУУУ участке.
20
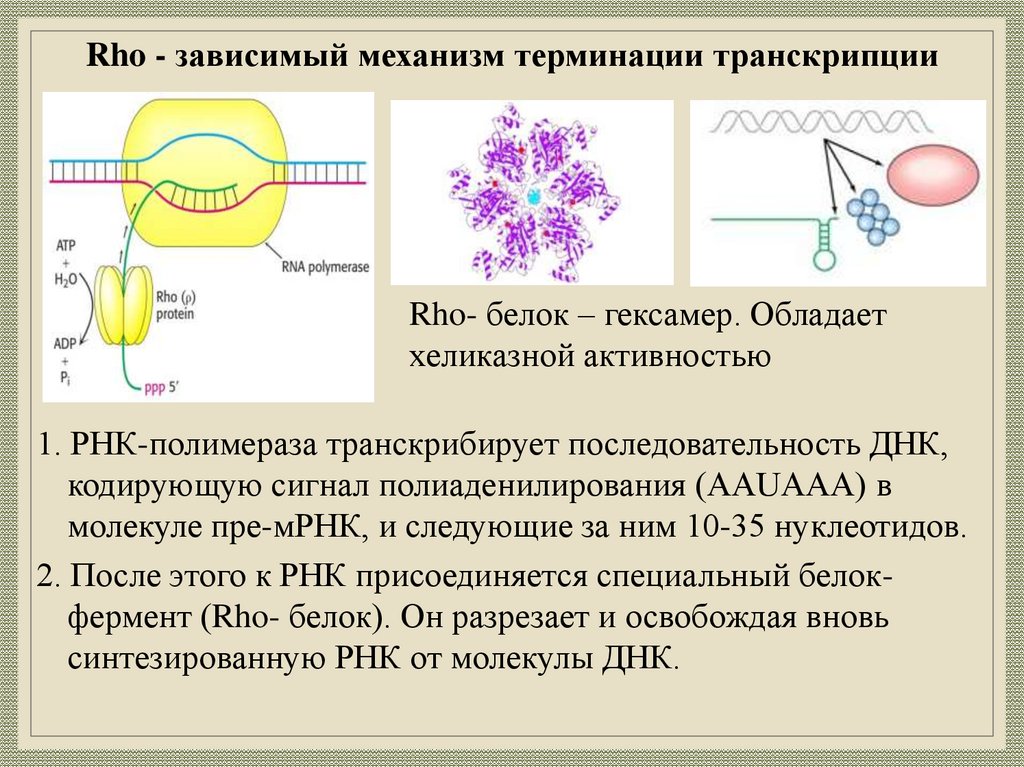
21.
Rho - зависимый механизм терминации транскрипцииRho- белок – гексамер. Обладает
хеликазной активностью
1. РНК-полимераза транскрибирует последовательность ДНК,
кодирующую сигнал полиаденилирования (AAUAAA) в
молекуле пре-мРНК, и следующие за ним 10-35 нуклеотидов.
2. После этого к РНК присоединяется специальный белокфермент (Rho- белок). Он разрезает и освобождая вновь
синтезированную РНК от молекулы ДНК.
22.
Процессинг – стадия созревания РНКЭукариотические мРНК синтезируются в виде
предшественников и проходят в своем биогенезе стадию
довольно сложного созревания, или процессинга.
Этапы процессинга:
- присоединения к 5'-концу пре-мРНК 7-метил
гуанозинтрифосфата («колпачка» или кэпа );
- присоединение к 3'- концу пре-мРНК
нуклеотидной последовательности, насчитывающей
100 — 200 остатков адениловой кислоты;
- сплайсинг: вырезание интронов и «сшивание»
экзонов
22
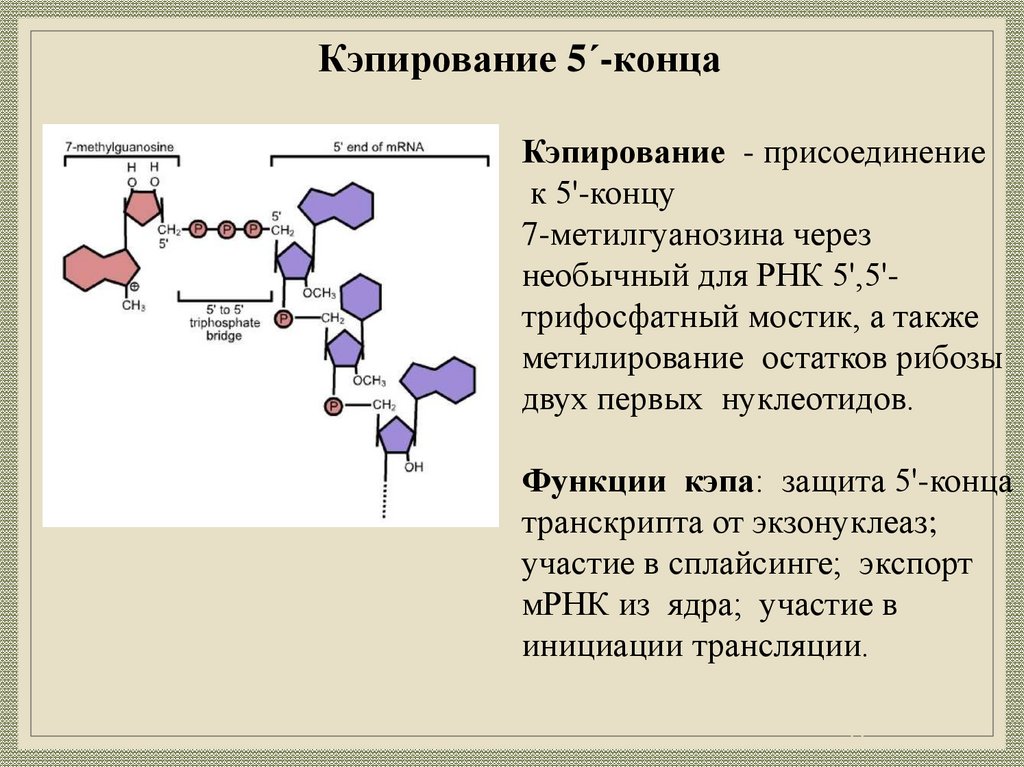
23.
Кэпирование 5ʹ-концаКэпирование - присоединение
к 5'-концу
7-метилгуанозина через
необычный для РНК 5',5'трифосфатный мостик, а также
метилирование остатков рибозы
двух первых нуклеотидов.
Функции кэпа: защита 5'-конца
транскрипта от экзонуклеаз;
участие в сплайсинге; экспорт
мРНК из ядра; участие в
инициации трансляции.
23
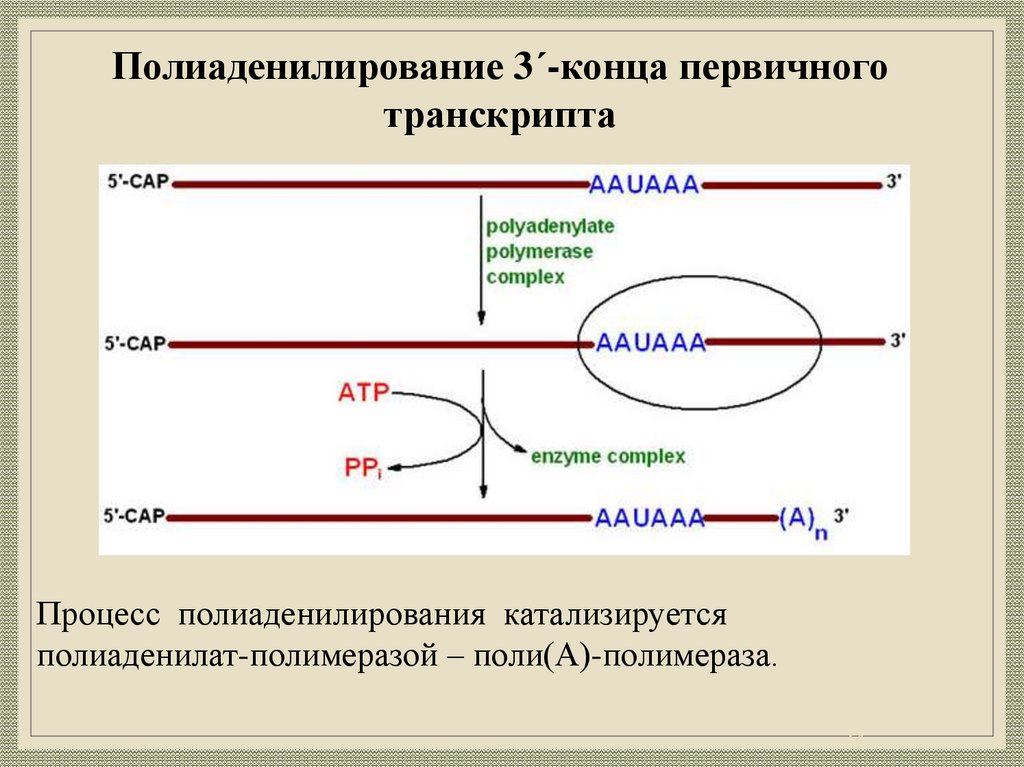
24.
Полиаденилирование 3ʹ-конца первичноготранскрипта
Процесс полиаденилирования катализируется
полиаденилат-полимеразой – поли(А)-полимераза.
24
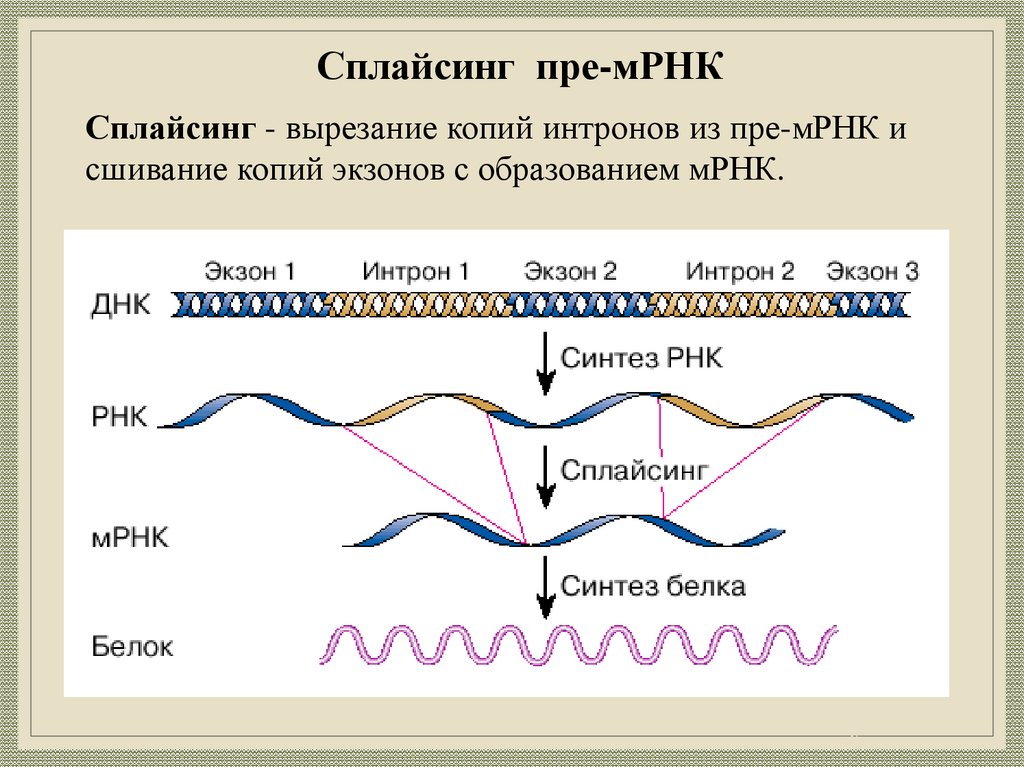
25.
Сплайсинг пре-мРНКСплайсинг - вырезание копий интронов из пре-мРНК и
сшивание копий экзонов с образованием мРНК.
25
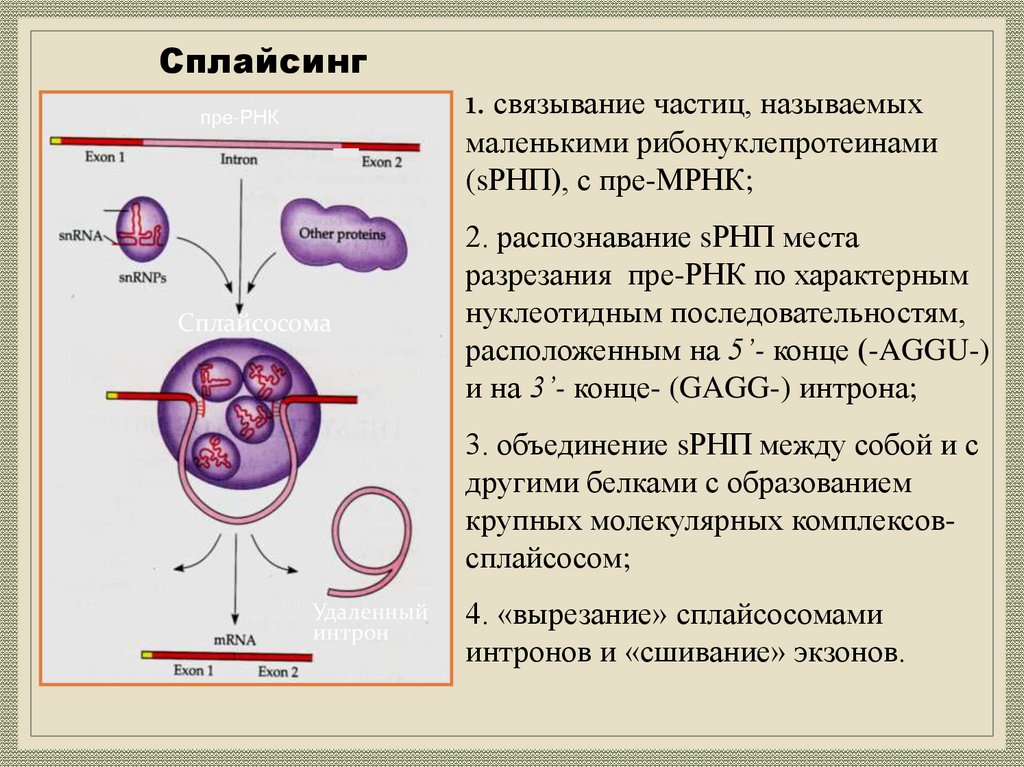
26.
Сплайсинг1. связывание частиц, называемых
пре-РНК
маленькими рибонуклепротеинами
(sРНП), с пре-МРНК;
Белок
Сплайсосома
2. распознавание sРНП места
разрезания пре-РНК по характерным
нуклеотидным последовательностям,
расположенным на 5’- конце (-AGGU-)
и на 3’- конце- (GAGG-) интрона;
3. объединение sРНП между собой и с
другими белками с образованием
крупных молекулярных комплексовсплайсосом;
Удаленный
интрон
4. «вырезание» сплайсосомами
интронов и «сшивание» экзонов.
27.
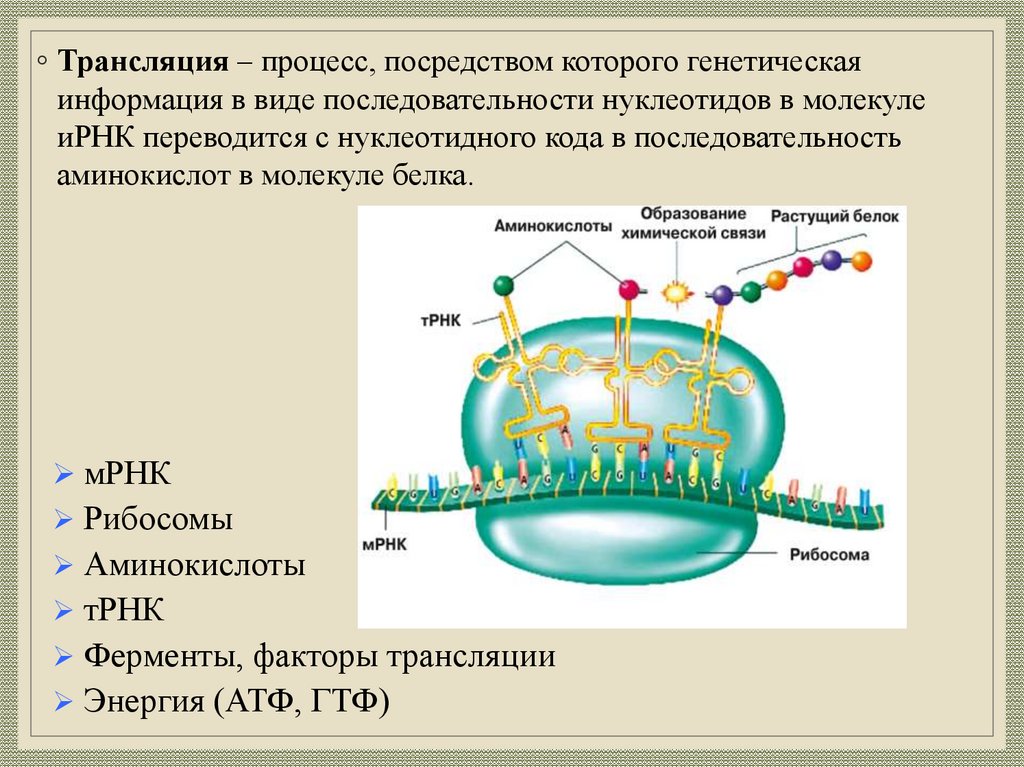
◦ Трансляция – процесс, посредством которого генетическаяинформация в виде последовательности нуклеотидов в молекуле
иРНК переводится с нуклеотидного кода в последовательность
аминокислот в молекуле белка.
мРНК
Рибосомы
Аминокислоты
тРНК
Ферменты, факторы трансляции
Энергия (АТФ, ГТФ)
28.
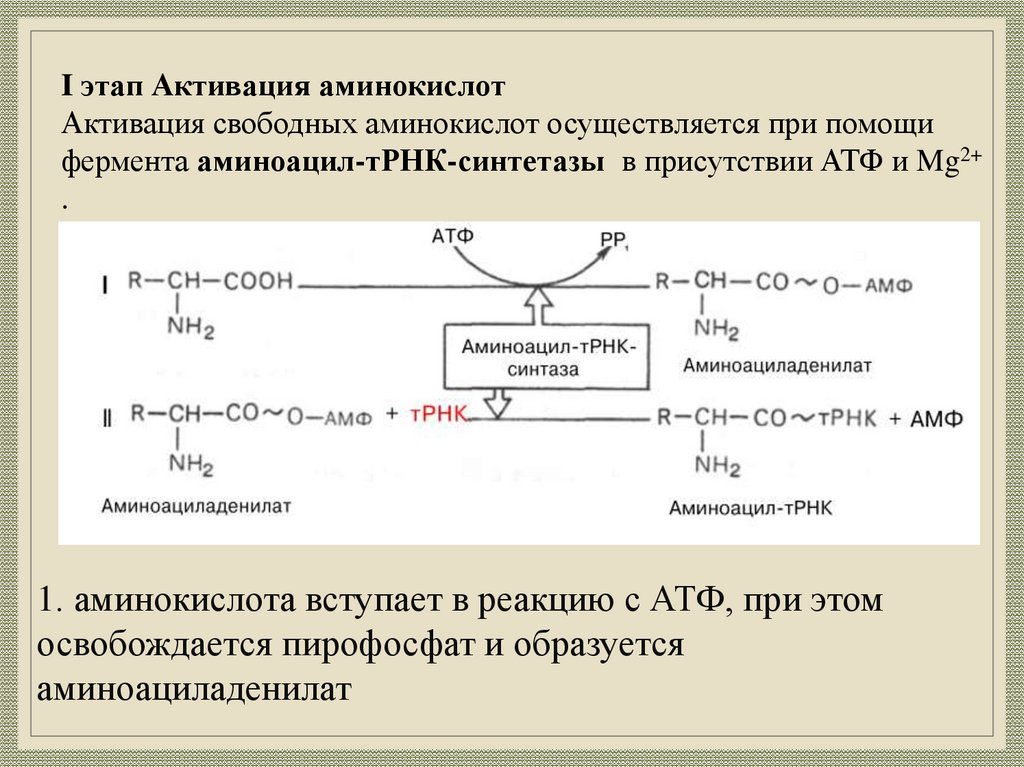
29.
I этап Активация аминокислотАктивация свободных аминокислот осуществляется при помощи
фермента аминоацил-тРНК-синтетазы в присутствии АТФ и Mg2+
.
1. аминокислота вступает в реакцию с АТФ, при этом
освобождается пирофосфат и образуется
аминоациладенилат
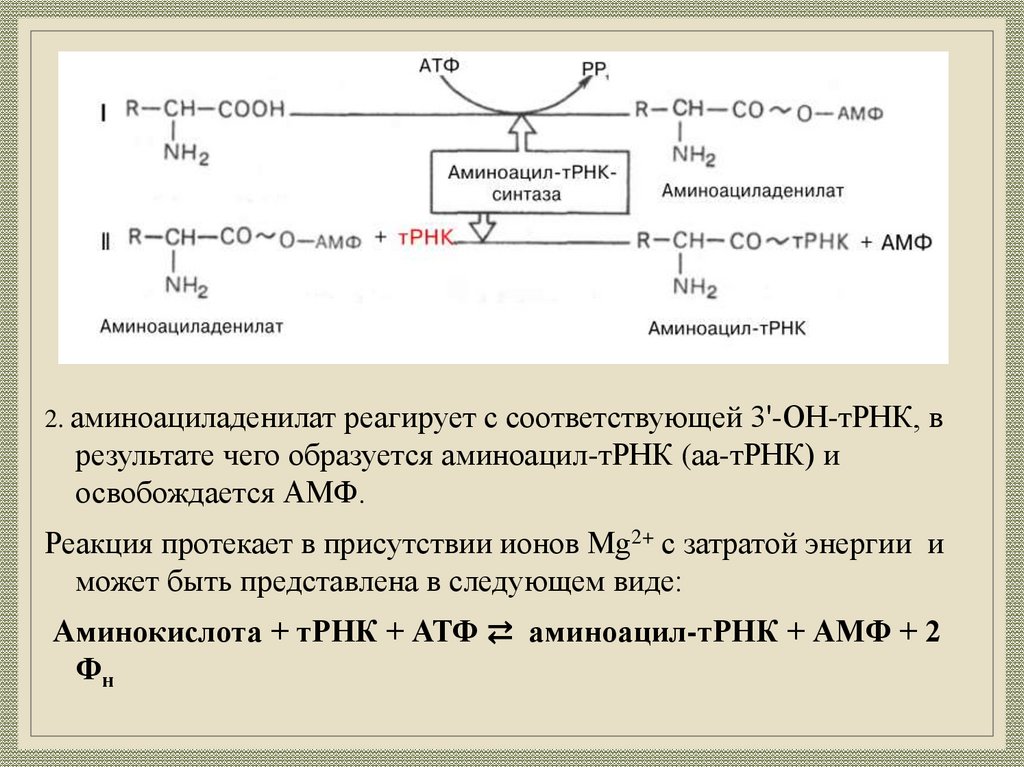
30.
2. аминоациладенилат реагирует с соответствующей 3'-ОН-тРНК, врезультате чего образуется аминоацил-тРНК (аа-тРНК) и
освобождается АМФ.
Реакция протекает в присутствии ионов Mg2+ с затратой энергии и
может быть представлена в следующем виде:
Аминокислота + тРНК + АТФ ⇄ аминоацил-тРНК + АМФ + 2
Фн
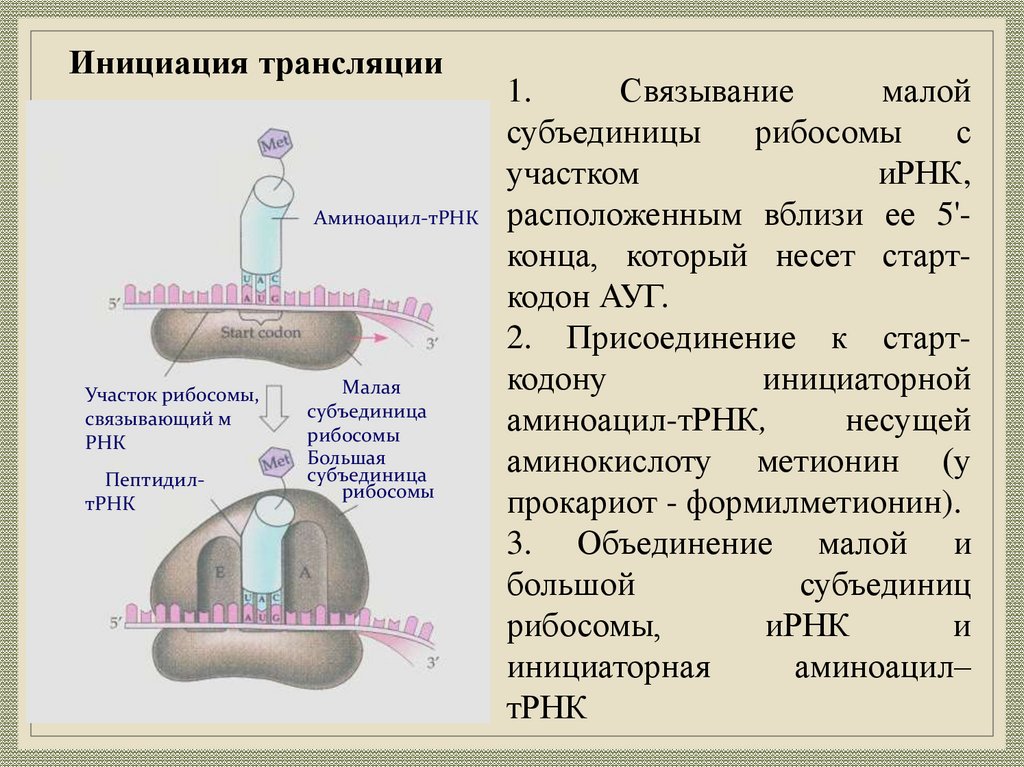
31.
Инициация трансляцииАминоацил-тРНК
Участок рибосомы,
связывающий м
РНК
ПептидилтРНК
Малая
субъединица
рибосомы
Большая
субъединица
рибосомы
1.
Связывание
малой
субъединицы
рибосомы
с
участком
иРНК,
расположенным вблизи ее 5'конца, который несет старткодон АУГ.
2. Присоединение к старткодону
инициаторной
аминоацил-тРНК,
несущей
аминокислоту метионин (у
прокариот - формилметионин).
3. Объединение малой и
большой
субъединиц
рибосомы,
иРНК
и
инициаторная
аминоацил–
тРНК
32.
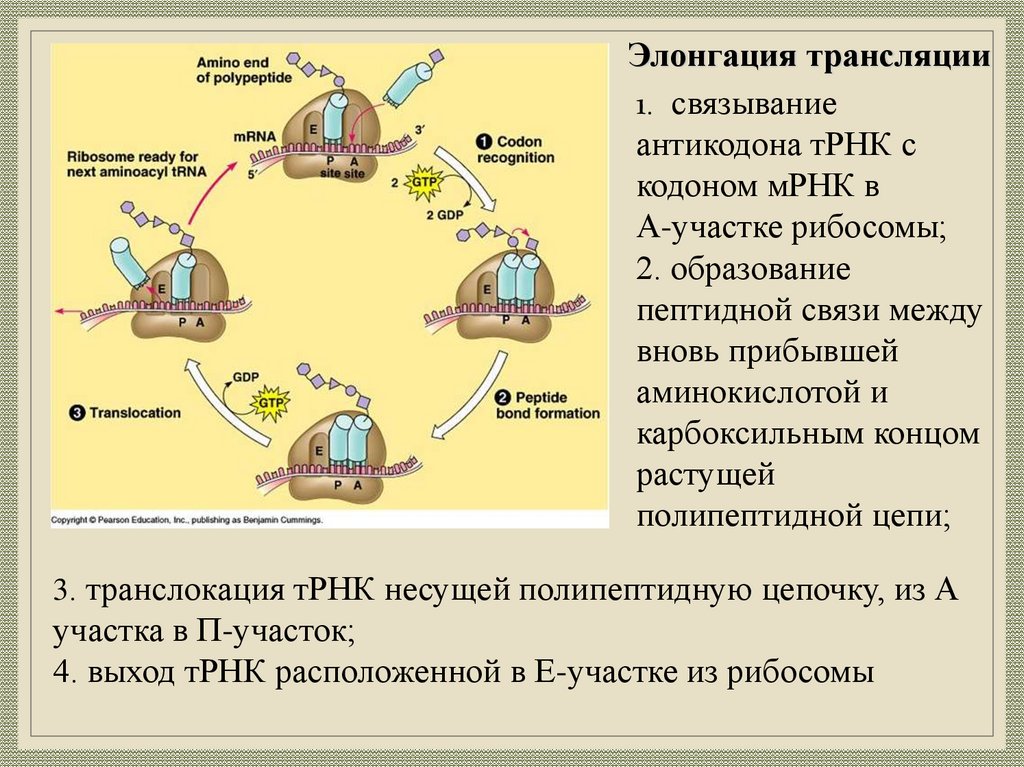
Элонгация трансляции1. связывание
антикодона тРНК с
кодоном мРНК в
А-участке рибосомы;
2. образование
пептидной связи между
вновь прибывшей
аминокислотой и
карбоксильным концом
растущей
полипептидной цепи;
3. транслокация тРНК несущей полипептидную цепочку, из А
участка в П-участок;
4. выход тРНК расположенной в Е-участке из рибосомы
33.
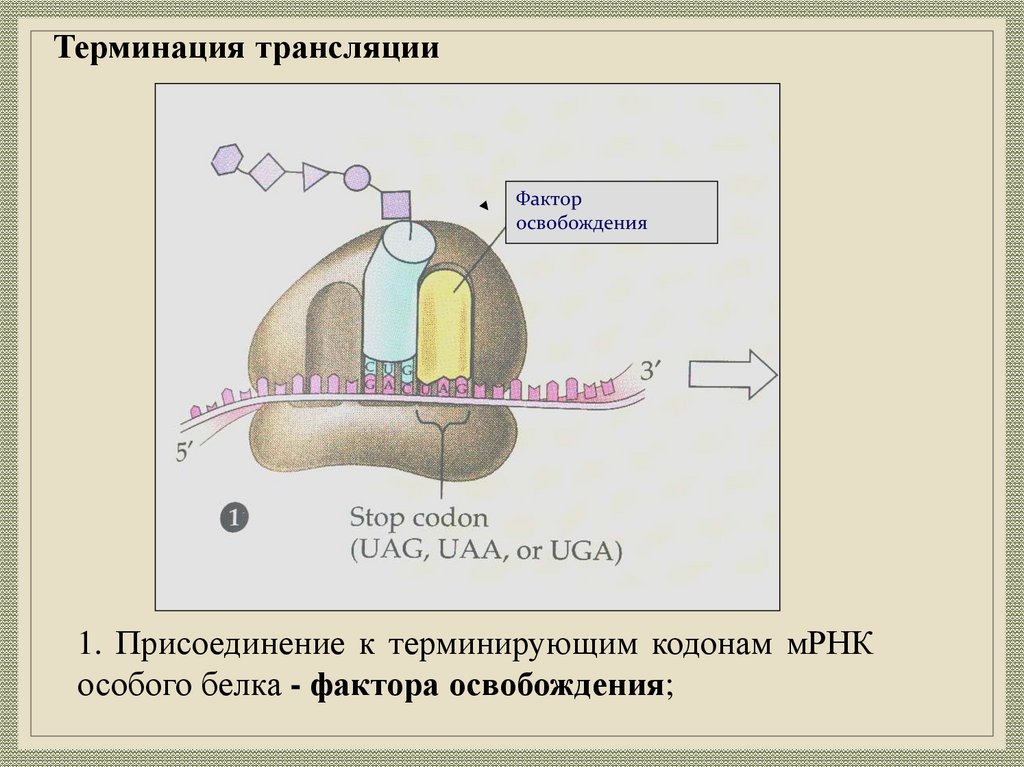
Терминация трансляцииФактор
освобождения
1. Присоединение к терминирующим кодонам мРНК
особого белка - фактора освобождения;
34.
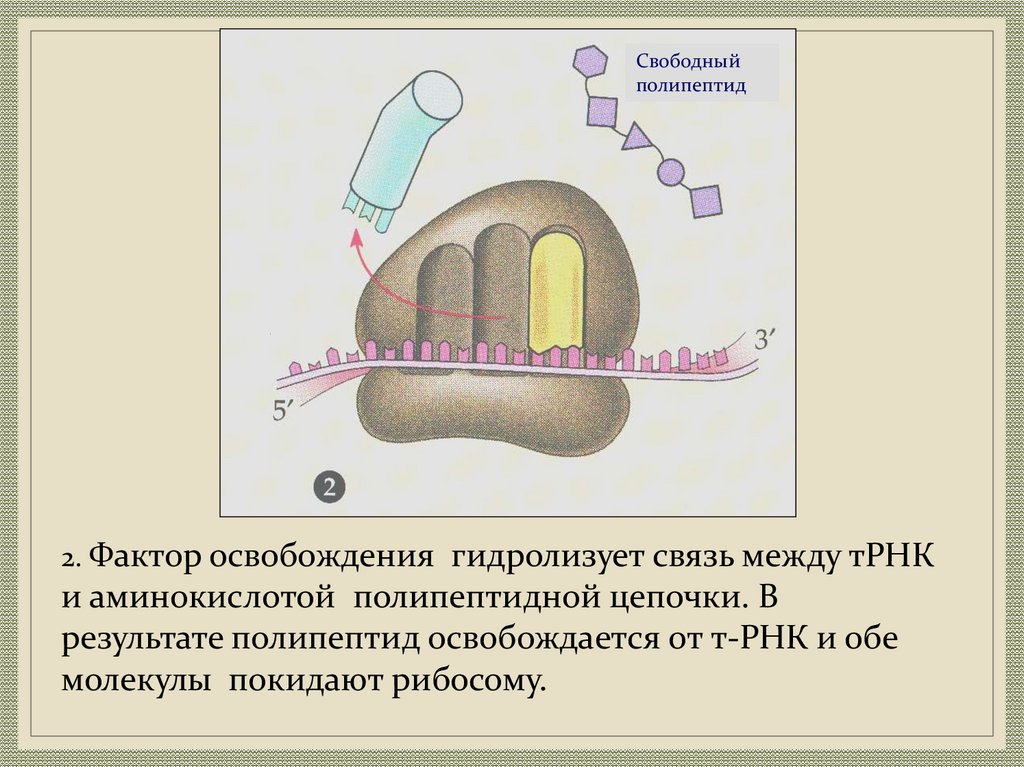
Свободныйполипептид
2. Фактор освобождения гидролизует связь между тРНК
и аминокислотой полипептидной цепочки. В
результате полипептид освобождается от т-РНК и обе
молекулы покидают рибосому.
35.
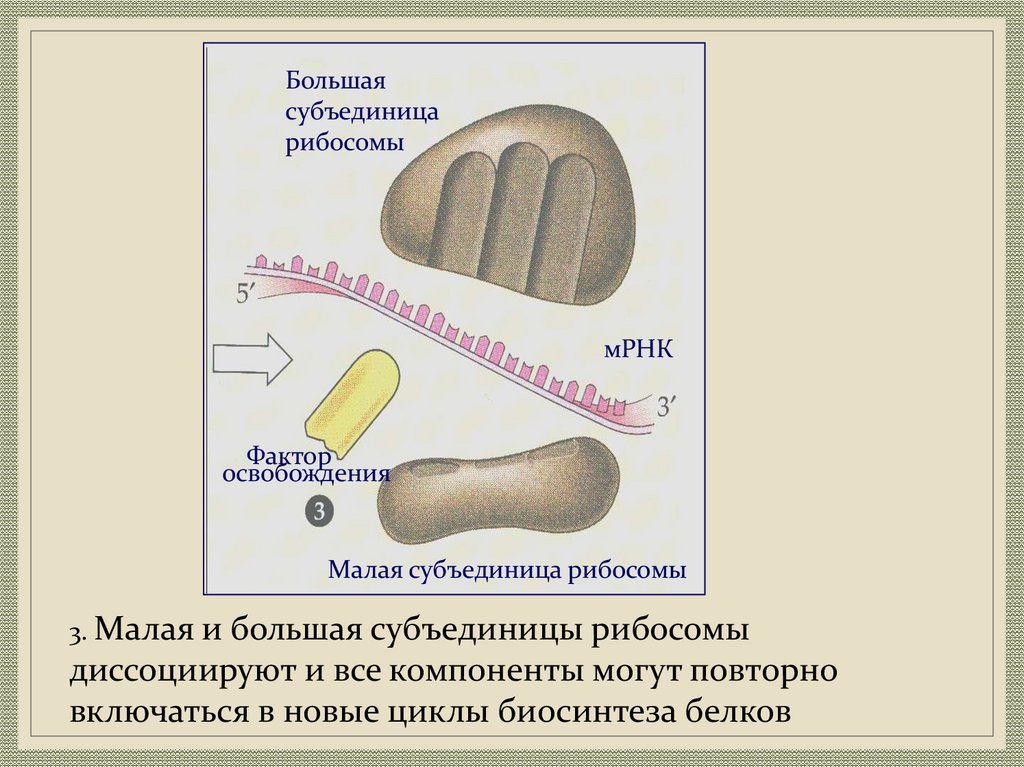
Большаясубъединица
рибосомы
мРНК
Фактор
освобождения
Малая субъединица рибосомы
3. Малая и большая субъединицы рибосомы
диссоциируют и все компоненты могут повторно
включаться в новые циклы биосинтеза белков
36.
Основные положения теории генетической информации.1. Носителем наследственной информации являются нуклеиновые
кислоты (главным образом ДНК, исключение – РНК вирусы).
2. Единицей наследственности является ген (участок ДНК,
характеризующийся определенной последовательностью
нуклеотидов).
3. Способность ДНК, как химической основы гена, к репликации
обеспечивает передачу наследственной информации из поколения в
поколение.
4. Генетическая информация о первичной структуре белка
кодируется с помощью определенной последовательности
нуклеотидов в цепи ДНК.
5. Биосинтез белка является процессом реализации наследственной
информации на матрице мРНК.
6. Образовавшиеся белки ферменты вступают в цепь
биохимических реакций, конечным результатом которых является
формирование фенотипического выражения признаков.