





")



















")






")






















")

 и длительности мышечного сокращения в ответ на 2 стимула, период между которыми")










")







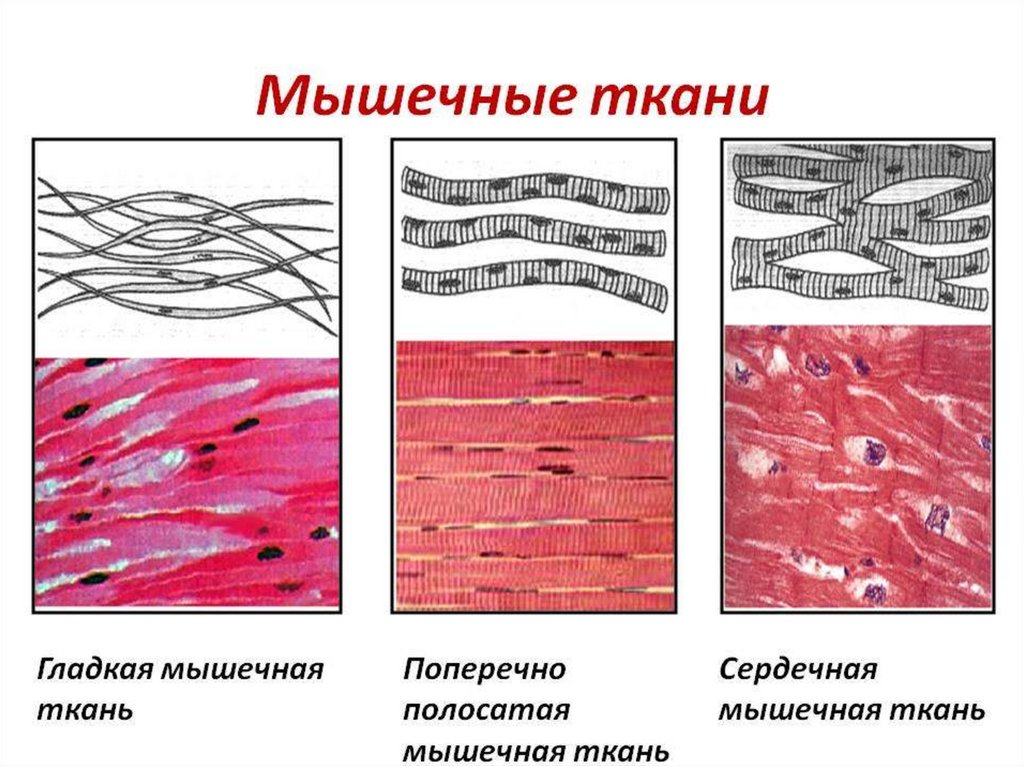
 гладкая мышца")





















 Биология
БиологияПохожие презентации:
")









Физиология мышечной ткани. Три вида мышц человека
1. ФИЗИОЛОГИЯ МЫШЕЧНОЙ ТКАНИ
2. Три вида мышц человека:
• поперечно-полосатыескелетные мышцы;
• сердечная мышца;
• гладкие мышцы внутренних
органов, кожи, сосудов.
3.
4. Физиологические свойства мышц:
1. возбудимость2. сократимость
3. рефрактерность
4. лабильность
5. проводимость
6. автоматизм
7. тонус
5. Возбудимость
• способность приходить всостояние возбуждения при
действии раздражителей.
• Непрямая– импульс
воздействует на нервное
окончание мотонейрона
• Прямая– импульс воздействует
непосредственно на мышцу
6. Сократимость
• способность мышцы изменятьсвою длину и/или напряжение в
ответ на действие
раздражителя.
Проводимость
• способность проводить
возбуждение вдоль и вглубь
мышечного волокна
7. Рефрактерность
• кратковременное снижениевозбудимости мышечной тканей
• Бывает: относительная и
абсолютная
Лабильность (250 Гц)
• функциональная подвижность
ткани, скорость протекания
элементарных циклов
возбуждения в мышечной тканях.
8. Автоматизм (Автоматия)
• способность клетки, ткани илиоргана возбуждаться под
влиянием импульса
возникающего в данном органе
9. Тонус мышц
• Тонус - умеренное напряжениемышц, когда они находятся в
состоянии относительного
покоя.
• Тонус поддерживается за счет
нервных импульсов,
поступающих из центральной
нервной системы от
мотонейронов передних рогов
спинного мозга.
10. ФИЗИЧЕСКИЕ СВОЙСТВА МЫШЦ
• Эластичность;• Пластичность;
• Вязкость;
• Способность развивать силу;
• Способность совершать
работу;
11. Пластичность
• способность мышцы изменятьсвою форму после деформации
Эластичность
• способность мышцы принимать
(восстанавливать) свою
первоначальную форму после
деформации.
12.
• Сила мышцы определяетсямаксимальным грузом, который
мышца в состоянии поднять.
• Работа мышцы
определяется
произведением величины
поднятого груза на высоту
подъема.
13.
Выделяют два диаметра:а) анатомический диаметр –
поперечное сечение мышц.
б) физиологический диаметр –
перпендикулярное
сечение
каждого мышечного волокна.
14. Сила мышц
Зависит от толщины мышцы и еёпоперечного физиологического
сечения
15. Работа мышцы
• Это энергия, затрачиваемая наперемещение тела с определенной
силой на определенное расстояние:
A=FxS
Если F=0, то и работа А=0
Если S=0, то и работа А=0
• Максимальная работа совершается
при средних нагрузках
Amax= Fср x Smax
16. Функции скелетных мышц:
• передвижение тела в пространстве;• перемещение частей тела относительно
друг друга (динамическая функция);
• поддержание позы (статическая
функция);
• передвижение крови и лимфы
(мышечный насос);
• выработка тепла;
• участие в акте вдоха и выдоха;
• Коммуникативная;
• Восприятие окружающего мира за счет
саккадических движений глаз
17. Функции скелетных мышц:
• двигательная активностьважнейший антистрессовый
фактор - «движение - это
жизнь»;
• депонирование воды и солей;
• защита внутренних органов
(пример: органов брюшной
полости).
Мышцы-щупала внешнего мира
(И.П.Павлов)
18.
Сердечная мышечная ткань• Нагнетательная функция
Гладкая мускулатура - образует стенку
полых органов и сосудов.
• поддерживает давление в полых органах;
• поддерживает величину кровяного
давления (тонус сосудов)
• обеспечивает продвижение содержимого
по желудочнокишечному тракту,
мочеточникам
• Родовой акт
• Резкость восприятия оптического
аппарата глаза
19. Мышца
• комплекс мышечных волокон(клеток), связанных
соединительной тканью
20.
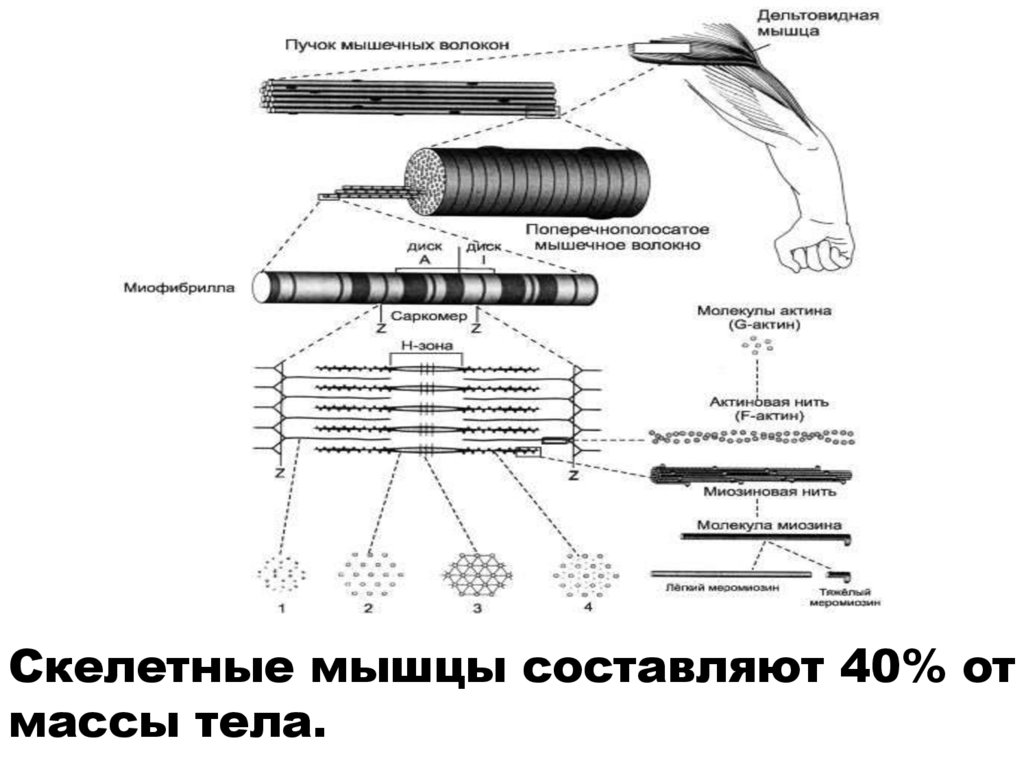
Скелетные мышцы составляют 40% отмассы тела.
21.
• Отдельное мышечное волокноподчиняется закону «Все или
ничего»
• Целая мышца подчиняется
закону «Силы» ( так как мышца
совокупность мышечных
волокон) волокон
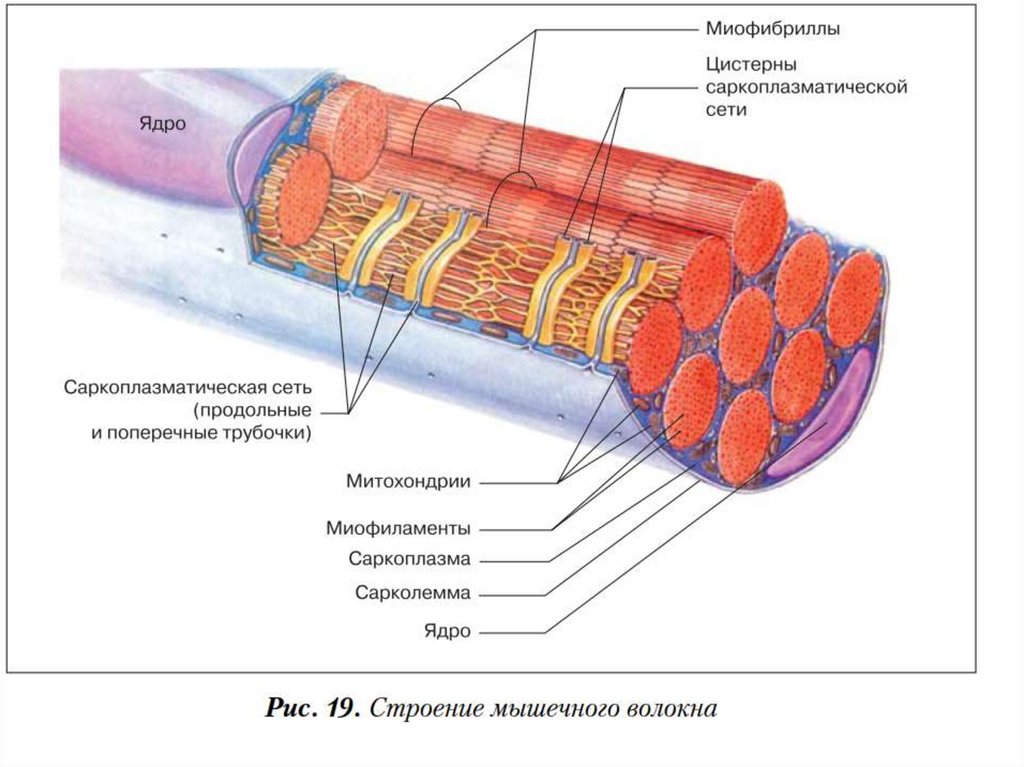
22. Сарколемма
мембрана окружающая мышечноеволокно, сходна по электрическим
свойствам с мембраной нервных
клеток
23. Миофибриллы
• органеллы клетокпоперечнополосатых мышц,
обеспечивающие их сокращение.
• Служат для сокращений мышечных
волокон.
• Миофибриллы создают
характерную поперечную
исчерченность.
• Каждая из миофибрилл состоит из
повторяющихся в продольном
направлении одинаковых
участков саркомеров.
24. Саркомер
• базовая структурнофункциональная сократительнаяединица поперечнополосатой
мышечной ткани, участок
миофибриллы между двумя
соседними Z-линиями (Zмебранами).
• Из саркомеров состоят
миофибрилы.
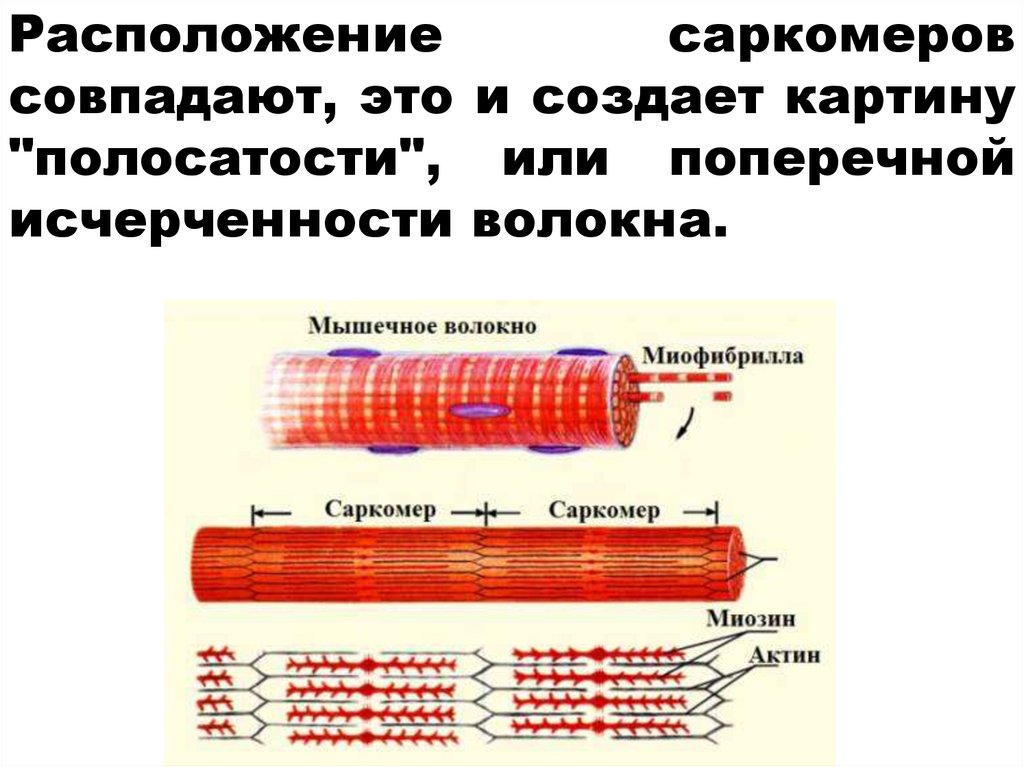
25.
Расположениесаркомеров
совпадают, это и создает картину
"полосатости", или поперечной
исчерченности волокна.
26. Актин
• глобулярный белок, состоящий из376 аминокислотных остатков
• Представлен двойной нитью,
закрученной в спираль с шагом
36,5 нм. Длина 1 мкм, диаметр 6-8
нм
• В состав актинового филамента
входят два вспомогательных
белка - тропомиозин и тропонин
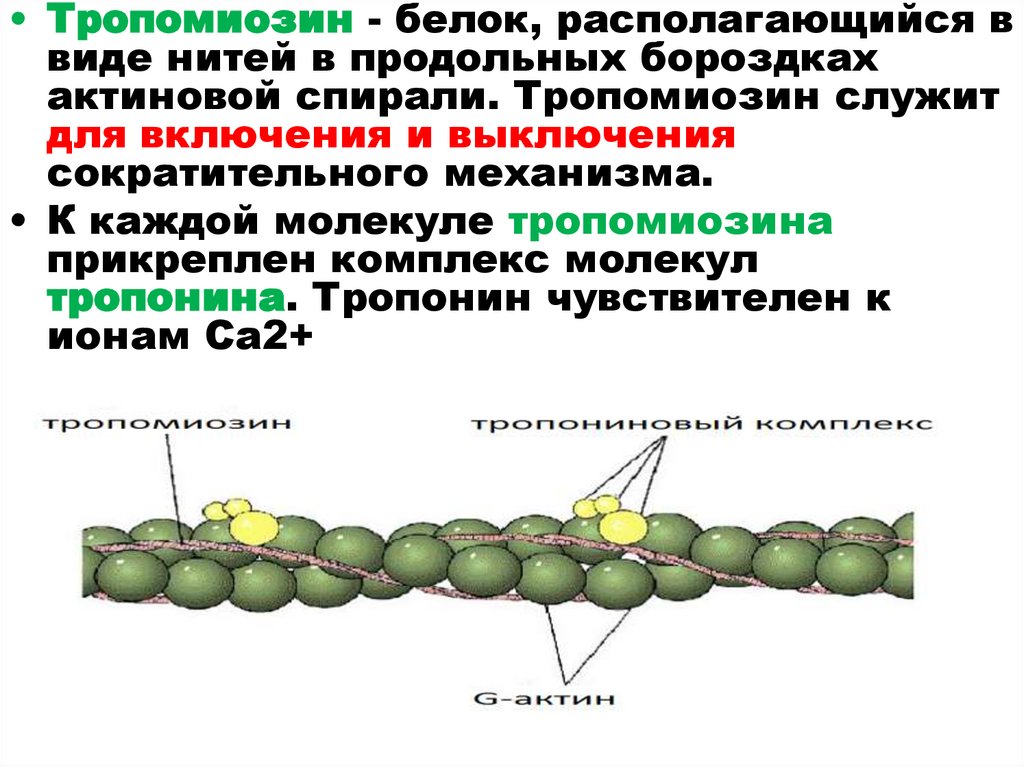
27.
• Тропомиозин - белок, располагающийся ввиде нитей в продольных бороздках
актиновой спирали. Тропомиозин служит
для включения и выключения
сократительного механизма.
• К каждой молекуле тропомиозина
прикреплен комплекс молекул
тропонина. Тропонин чувствителен к
ионам Са2+
28. Миозин
• Миозин составляет 40-60 % общегоколичества мышечных белков.
• В 2 раза толще нитей актина,
длиной 1,6 мкм
• Имеет поперечные мостики,
состоящие из головки и шейки
• Миозин расщепляет АТФ. Благодаря
АТФ-азной активности миозина,
химическая энергия
макроэргических связей АТФ
превращается в механическую
энергию мышечного сокращения.
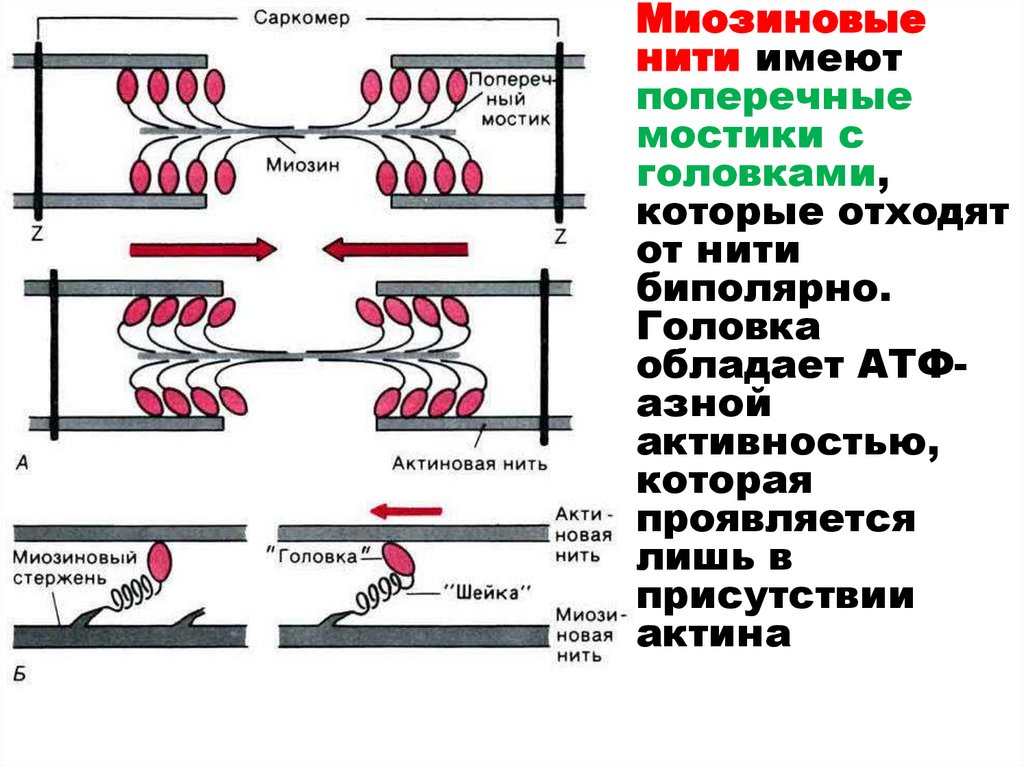
29.
Миозиновыенити имеют
поперечные
мостики с
головками,
которые отходят
от нити
биполярно.
Головка
обладает АТФазной
активностью,
которая
проявляется
лишь в
присутствии
актина
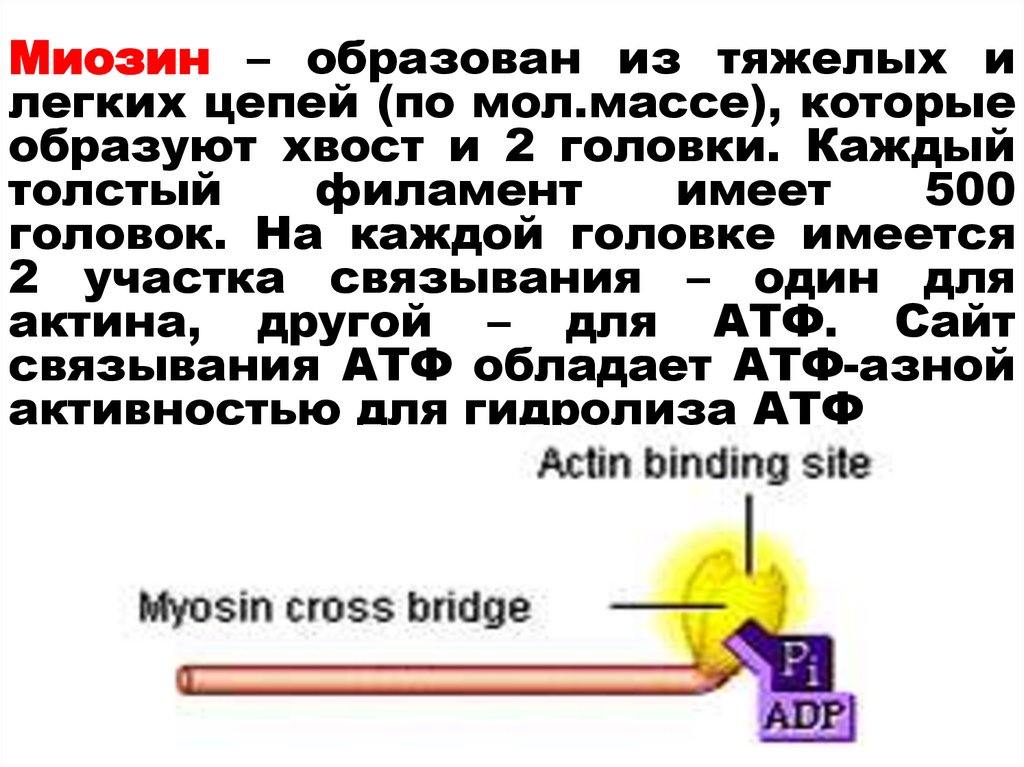
30.
Миозин – образован из тяжелых илегких цепей (по мол.массе), которые
образуют хвост и 2 головки. Каждый
толстый
филамент
имеет
500
головок. На каждой головке имеется
2 участка связывания – один для
актина, другой – для АТФ. Сайт
связывания АТФ обладает АТФ-азной
активностью для гидролиза АТФ
31. Взаимодействие актиновых и миозиновых филаментов
10080
60
Восток
40
Запад
20
Север
0
1 2 3 4
кв кв кв кв
32. Митохондрия
• двумембранный органоидэукариотической клетки, основная
функция которого синтез АТФ –
источника энергии для
жизнедеятельности клетки.
• Большое количество митохондрий
находится между миофибриллами,
что является свидетельством
протекания здесь
высокоэнергетических процессов.
33.
34. Саркоплазматический ретикулум (СР)
• пространство между миофибрилламизаполненное саркоплазмой, в
которой находится сложная система
трубок и цистерн.
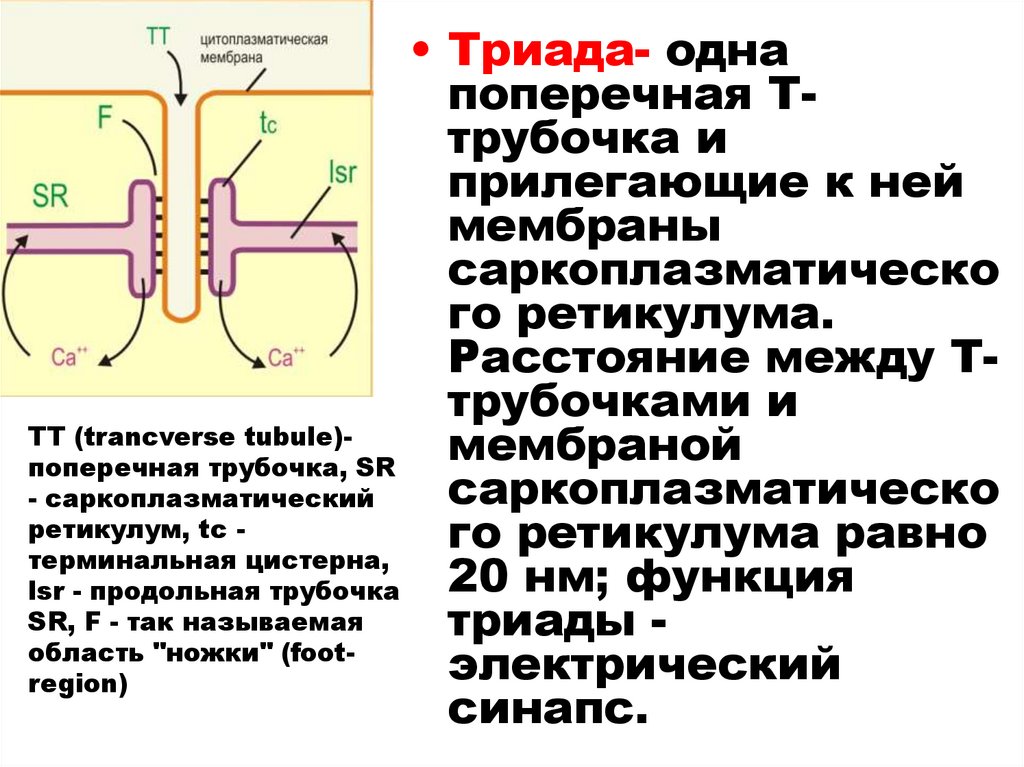
35.
• Триада- однапоперечная Ттрубочка и
прилегающие к ней
мембраны
саркоплазматическо
го ретикулума.
Расстояние между Ттрубочками и
ТТ (trancverse tubule)мембраной
поперечная трубочка, SR
саркоплазматическо
- саркоплазматический
ретикулум, tc го ретикулума равно
терминальная цистерна,
20 нм; функция
lsr - продольная трубочка
SR, F - так называемая
триады область "ножки" (footэлектрический
region)
синапс.
36.
Нейромоторная ( двигательная) единица совокупность нейрона и группымышечных волокон, иннервируемых
аксоном этого нейрона.
В состав нейромоторной единицы входят:
• нервная клетка - в основном
мотонейроны, тела которых лежат в
передних рогах спинного мозга;
• группа мышечных волокон - в
зависимости от вида деятельности
количества волокон различно. Если
тонкая работа 2-4, если грубая - до
нескольких тысяч.
Число мышечных волокон в различных ДЕ
• Глаза менее 10
• Пальцы рук 1-25
• Камбаловидная мышца 2000
37.
Схемадвигательной
единицы:
а – мотонейрон;
б–нервномышечные
синапсы;
в – мышечные
волокна.
Концевые веточки заканчиваются на
мышечных волокнах, образуя
нервно-мышечный синапс,
названный моторной пластинкой.
38.
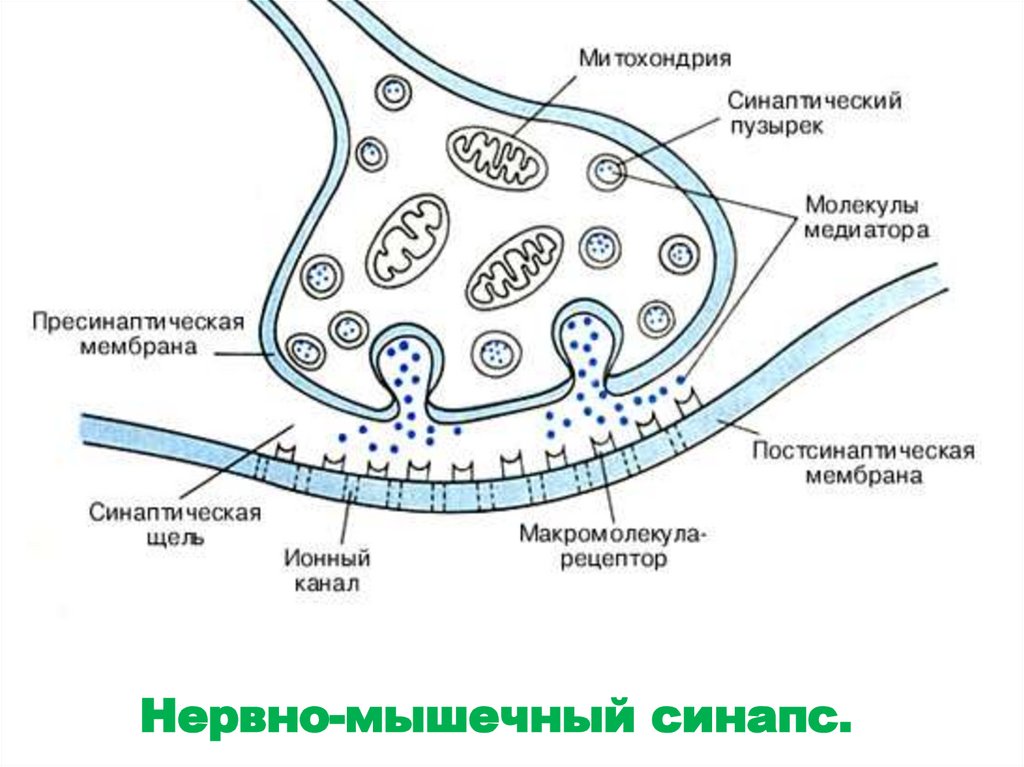
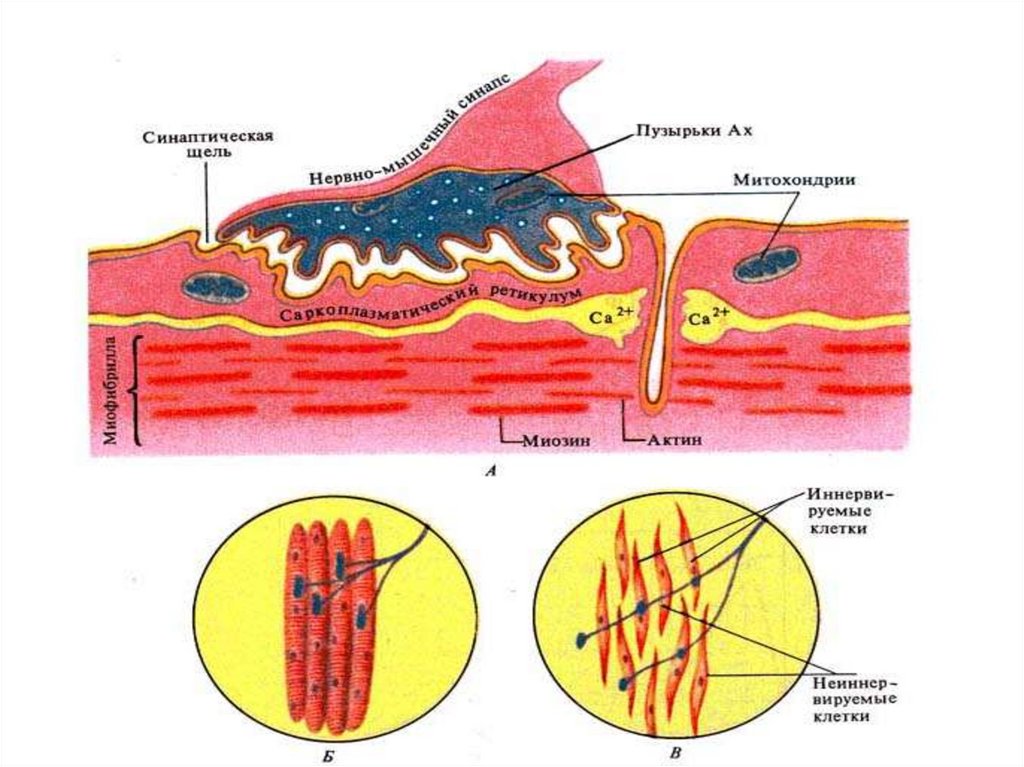
Нервно-мышечный синапс.39.
40. Нервно-мышечный синапс
• Нейромедиатором в этом синапсеявляется ацетилхолин.
• В синапсе нервный импульс
превращается в механическое движение
мышечной ткани.
• Внутри нервного окончания имеется
большое количество (до 300.000)
синаптических пузырьков (диаметром
около 50 нм), содержащих химическое
соединение ацетилхолин (АХ).
• Это химический передатчик возбуждения
- медиатор.
• Каждый пузырек содержит «квант»
медиатора - около 104 молекул АХ.
41. Передача возбуждения в нервно-мышечном синапсе.
Передача возбуждения в нервномышечном синапсе.• Нервный импульс достигает концевой
веточки аксона и вызывает выделение
медиатора ацетилхолина в
синаптическую щель.
• Запасов ацетилхолина в нервном
окончании достаточно для проведения
примерно 10 тысяч импульсов.
• Медиатор диффундирует к
постсинаптической мембране и образует
на ней комплекс с холинорецептором.
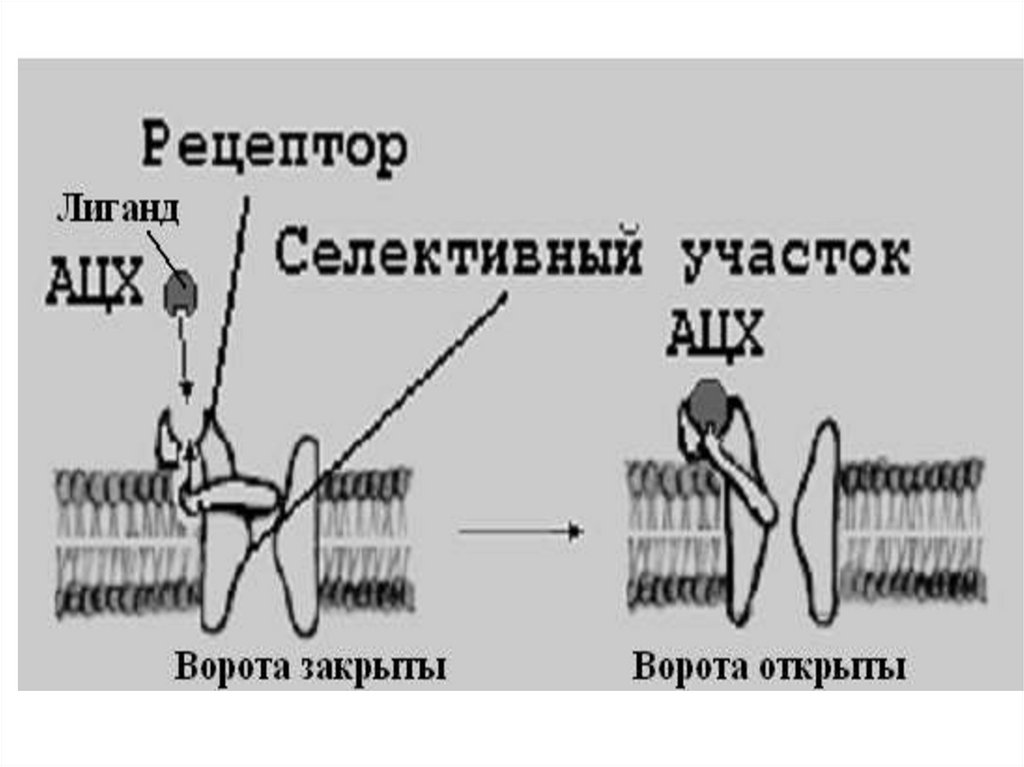
42. Взаимодействие медиатора с постсинаптической мембраной
• Результатом взаимодействия АХ с ХРявляется открытие хемовозбудимых
ионных каналов. В норме превалирует
поток ионов натрия. Они по
концентрационному градиенту из
синаптической щели поступают внутрь
мышечного волокна и деполяризуют
постсинаптическую мембрану.
• Возникает ПКП (потенциал концевой
пластинки)
Концевая пластинка – участок мышечной
мембраны, прилегающий к аксону и
содержащий
большое
количество
натриевых каналов
43.
44.
Медиатор разрушается поддействием фермента холинэстеразы на холин и
уксусную кислоту.
Большая часть продуктов гидролиза
ацетилхолина возвращается в
синаптическую бляшку, где
участвует в ресинтезе новых
молекул медиатора, который
поступает во вновь формирующиеся
везикулы.
45. Теория скольжения нитей (Хаксли)
• В 1954 г. А. Хаксли прииспользовании микроскопа
установил, что актиновые и
миозиновые филаменты не
изменяют своей длины как при
укорочении или удлинении
саркомера.
• В момент изменения длины
саркомера происходит
перекрывание актиновых и
миозиновых филаментов, в
результате их скольжения.
46. Механизмы сокращения мышечного волокна
• В покое мышечные волокнанаходятся в расслабленном
состоянии.
• Концентрация ионов Са2+ в
межфибриллярном пространстве
меньше 10-8.
• Белок тропомиозин блокирует
участки белка актина, способные
взаимодействовать с
поперечными мостиками миозина.
47.
• При возбуждении мышечноговолокна потенциал действия,
возникающий в области концевой
пластинки, распространяется по
Т-системе вглубь волокна,
вызывает активацию мембран
цистерн саркоплазматического
ретикулума и освобождает из них
ионы Са2+.
• Ионы Са2+ при их концентрации в
межфибриллярном пространстве
10-6 запускают процесс
сокращения.
48. Теория мышечного сокращения
Рецепторыдигидропиридина
(ДГП)
Ca2+
release
Рецептор
рианодина
49. В состоянии покоя
• В условиях покояголовка миозина не
прикреплена к
активной зоне
актиновой нити мешает тропомиозин.
• Белок тропомиозин
блокирует активные
участки белка актина,
способные
взаимодействовать с
поперечными
мостиками миозина
Тропомиозин
G-актин
50. Шаг 1. Ионы кальция, выходя из СР запускают процесс сокращения.
• Под влияниемШаг 1.
ионов Са2+
молекула
Ионы кальция, выходя
тропонина
из СР запускают
изменяет свою
процесс сокращения.
форму и
выталкивает
тропомиозин в
желобок между
двумя нитями
актина,
освобождая
активные
участки для
прикрепления
миозиновых
головок
(мостиков) к
актину.
51. Шаг 2: Головка миозина прикрепляется к активному участку актина
52. Шаг 3: Фермент миозиновая АТФ-аза гидролизует АТФ, расположенный на головке миозина, что обеспечивает энергией поперечные
мостики.Мостики
совершают
движение к
центру
саркомера.
Происходит
укорочение
саркомера.
53. Шаг 4: Освобождающиеся при гидролизе АТФ молекула АДФ и неорганический фосфат используются для последующего ресинтеза АТФ
На миозиновомпоперечном
мостике
образуется
новая молекула
АТФ. При этом
происходит
разъединение
поперечного
мостика с
нитью актина.
54. Шаг 5:
Повторноеприкрепление и
отсоединение
мостиков
продолжается до
тех пор, пока
концентрация
ионов Са2+ внутри
миофибрилл не
снизится до
подпороговой
величины.
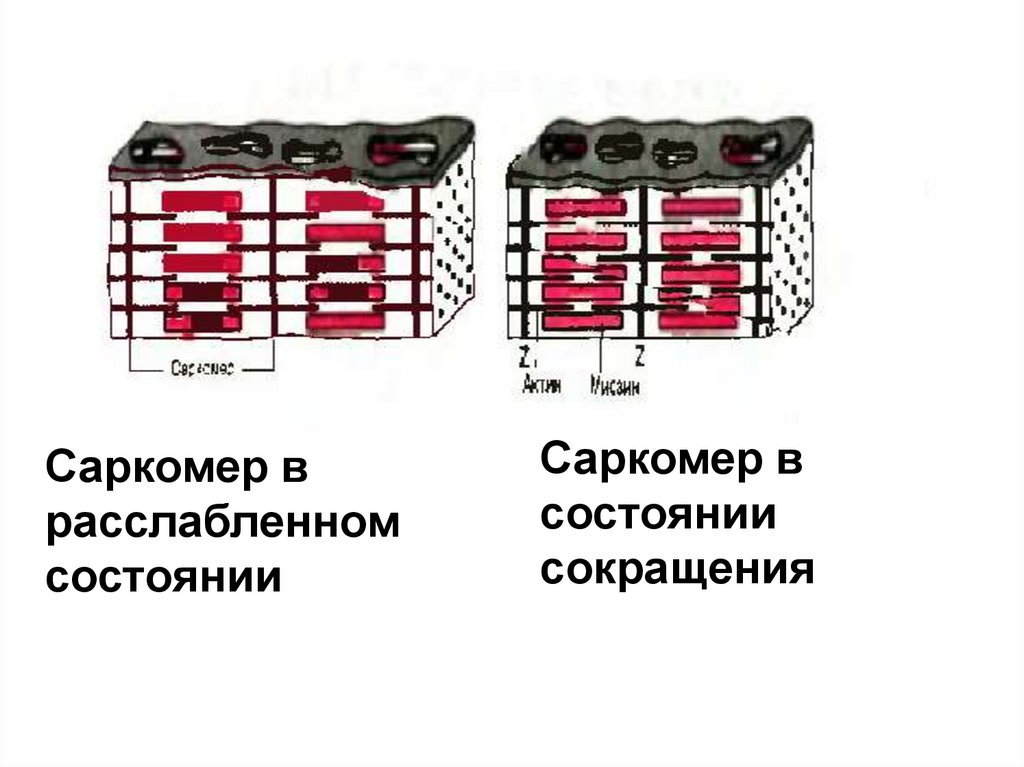
55.
Саркомер врасслабленном
состоянии
Саркомер в
состоянии
сокращения
56.
Откачивание ионов Са назад вСПР
Кальций в саркоплазме активирует
Са-АТФ-азу, кальциевый насос
осуществляет активный перенос
этого иона в СПР.
57.
В настоящее время расслаблениесчитается активным процессом,
причем расход энергии больше, чем
при сокращении. Источник
сокращения - АТФ,
расщепляющийся при сокращении.
Доказательства:
• появление мышечных контрактур
при утомлении (т.е. при деятельном
сокращении);
• явления трупного окоченения
(отсутствие энергии).
58. Роль кальция в мышечном сокращении
1 – Выброс медиатора всинаптическую щель.
2 – Освобождение активного
центра актина.
3 – Расслабление мышцы (разрыв
связи миозина с актином – АТФаза кальциевая).
59. Роль АТФ в мышечном сокращении
а) сокращения (образованиямостиков);
б) расслабления (разрыва мостиков);
в) работы Са-насоса (2 АТФ и 1 ион
Са);
г) работы Nа,К-насоса.
Однако в саркоплазме мышцы АТФ
относительно немного. Ее хватит лишь на
несколько мышечных сокращений
(примерно 8 одиночных сокращений).
60. Пути ресинтеза АТФ
1) креатинфосфокиназный (КФ):АДФ + КФ <==> АТФ + К
(3,6 моль АТФ/мин)
2) гликолитический (1,2 моль
АТФ/мин),
З) аэробное окисление
(при окислении глюкозы - 0,8 моль
АТФ/мин, жиров - 0,4 моль АТФ/мин)
61.
Энергетические потребностиМышечное сокращение требует
значительных
энергетических
затрат.
Основной
источник
энергии — гидролиз макроэрга
АТФ. В митохондриях в процессе
цикла трикарбоновых кислот и
окислительного
фосфорилирования генерируется
АТФ.
62.
Гликоген запасaется всaркоплазме в виде включений.
Анаэробный гликолиз сопряжён
с синтезом АТФ.
Креатинфосфокиназа,
связанная в области М-линии,
катализирует перенос фосфата
от фосфокреатина на АДФ с
образованием креатина и АТФ.
63.
Миоглобин, как и Hb, обратимосвязывает
кислород.
Запасы
кислорода
необходимы
для
синтеза АТФ при длительной
непрерывной работе мышцы. На
один рабочий цикл затрачивается
1
молекула
АТФ.
В
МВ
концентрация
АТФ
равна
4 ммоль/л. Такого запаса энергии
достаточно
для
поддержания
сокращения не более 1–2 сек.
64.
Утомление–
временное
снижение
работоспособности,
наступающее
в
процессе
выполнения мышечной работы и
исчезающее после отдыха.
Причины утомления:
1.
Накопление
продуктов
обмена (молочная кислота) в
мышцах, что ведет к угнетению
генерации потенциала действия.
2. Кислородное голодание, т.е. к
мышце не успевает доставляться
кислород.
65.
3. Истощение энергии.4. Центрально-нервная теория
утомления.
По
этой
теории
утомление
нервных
клеток
наступает быстрее, чем мышц.
5. Утомление синапсов, через
которые импульсы передаются к
мышцам.
66. Режимы мышечного сокращения
1. Изотонический — волокнамышцы изменяют длину без
изменения их напряжения
2. Изометрический - мышца
развивает напряжение без
изменения своей длины.
3. Ауксотонический - мышца
развивает напряжение и
изменяет длину
( эксцентрический и
концентрический)
67. Типы мышечных сокращений
Типы мышечных сокращенийзависит от частоты импульсации
стимулов:
• Одиночное
• Суммация
• Тетанус
68. Одиночное сокращение
механический ответ мышечноговолокна на одиночный стимул
(0,1с)
Выделяют три фазы сокращения
1. латентная (срытая) фаза возбуждения
это время, прошедшее от нанесения
раздражения до момента начала
механической реакции мышцы (0,01с)
2. фаза напряжения или укорочения (0,04
с)
3. фаза расслабления или удлинения (0,05
с)
В естественных условиях характерно
только для сердечной мышцы
69. Фазы одиночного мышечного сокращения (ОМС)
70. Соотношение фаз ПД с фазами сокращения и возбудимости
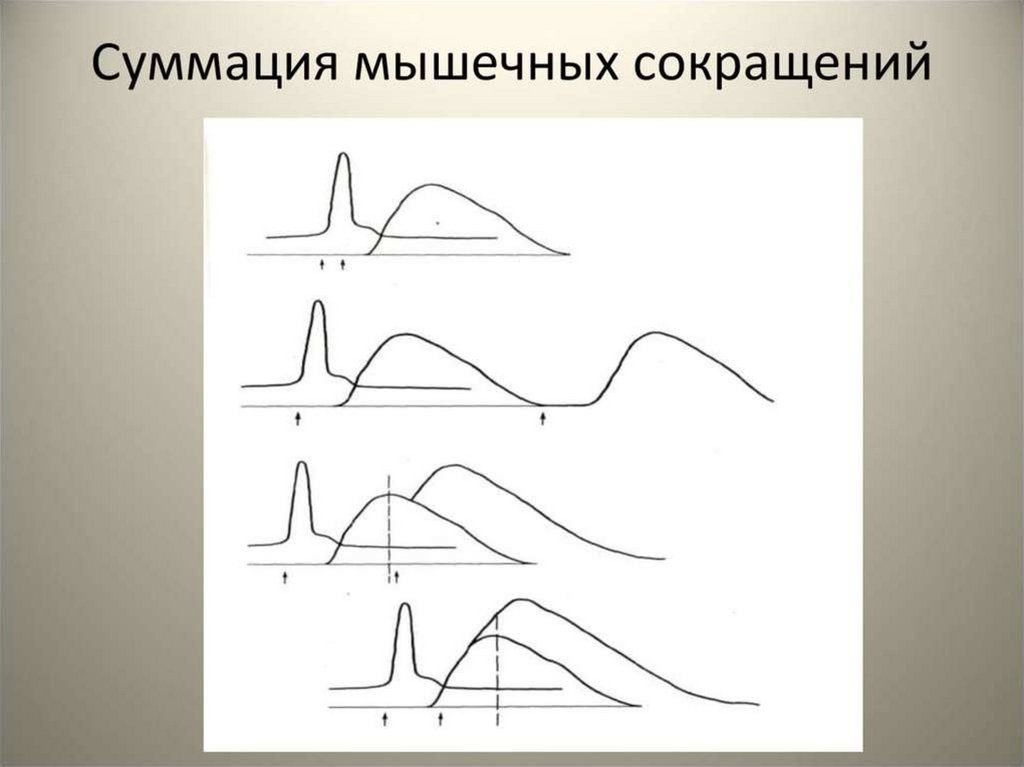
71. Суммация сокращений увеличение амплитуды(силы) и длительности мышечного сокращения в ответ на 2 стимула, период между которыми
меньшеОМС, но больше латентного периода
72.
73. Суммация и тетанус мышечных сокращений
74. Тетанус
Длительное, сильное сокращениемышцы в ответ на многочастотный
импульс
• К тетаническому сокращению
способны только скелетные
мышцы.
• Мышца сердца не способны к
тетаническому сокращению,
вследствие наличия
продолжительного рефрактерного
периода.
75. Тетанус
• Тетанус возникает вследствиемножественной суммации.
• Для того, чтобы возник тетанус,
необходимо действие повторных
раздражителей (или нервных
импульсов) на мышцу еще до того,
как закончится ее одиночное
сокращение.
76.
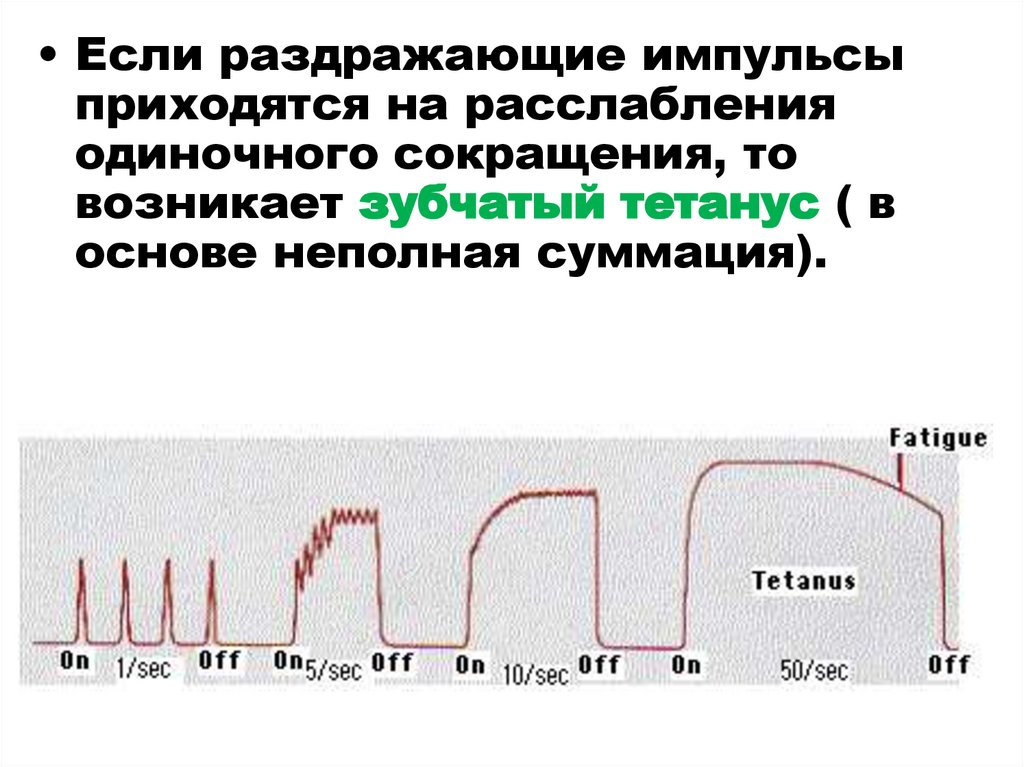
• Если раздражающие импульсыприходятся на расслабления
одиночного сокращения, то
возникает зубчатый тетанус ( в
основе неполная суммация).
77.
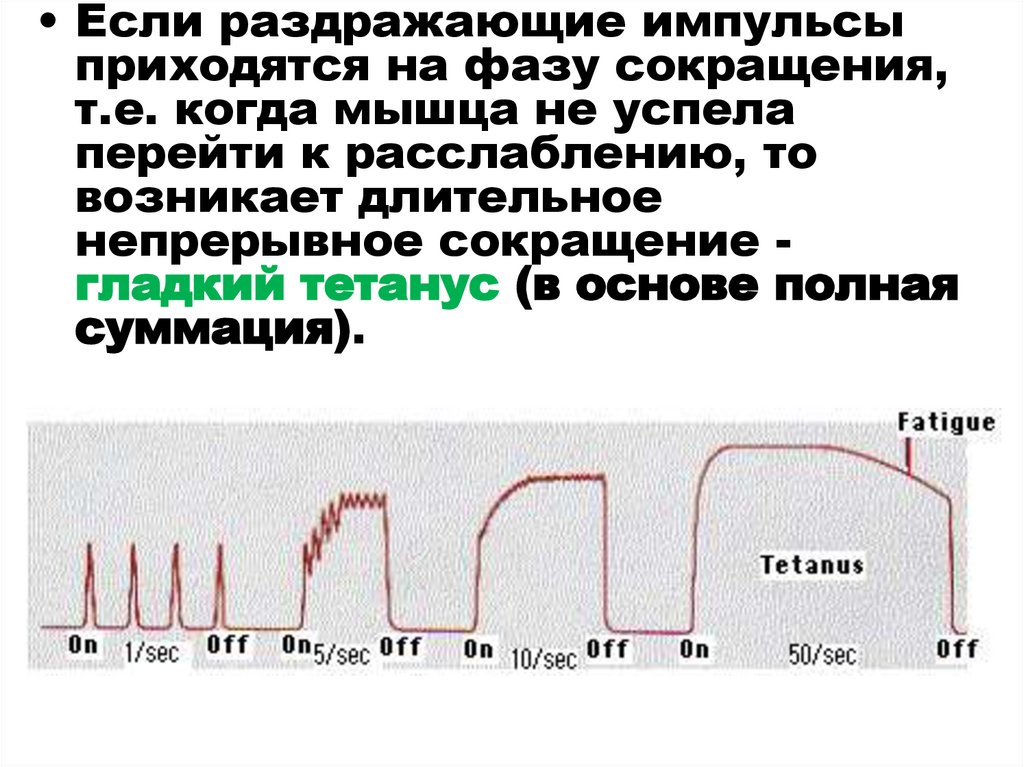
• Если раздражающие импульсыприходятся на фазу сокращения,
т.е. когда мышца не успела
перейти к расслаблению, то
возникает длительное
непрерывное сокращение гладкий тетанус (в основе полная
суммация).
78.
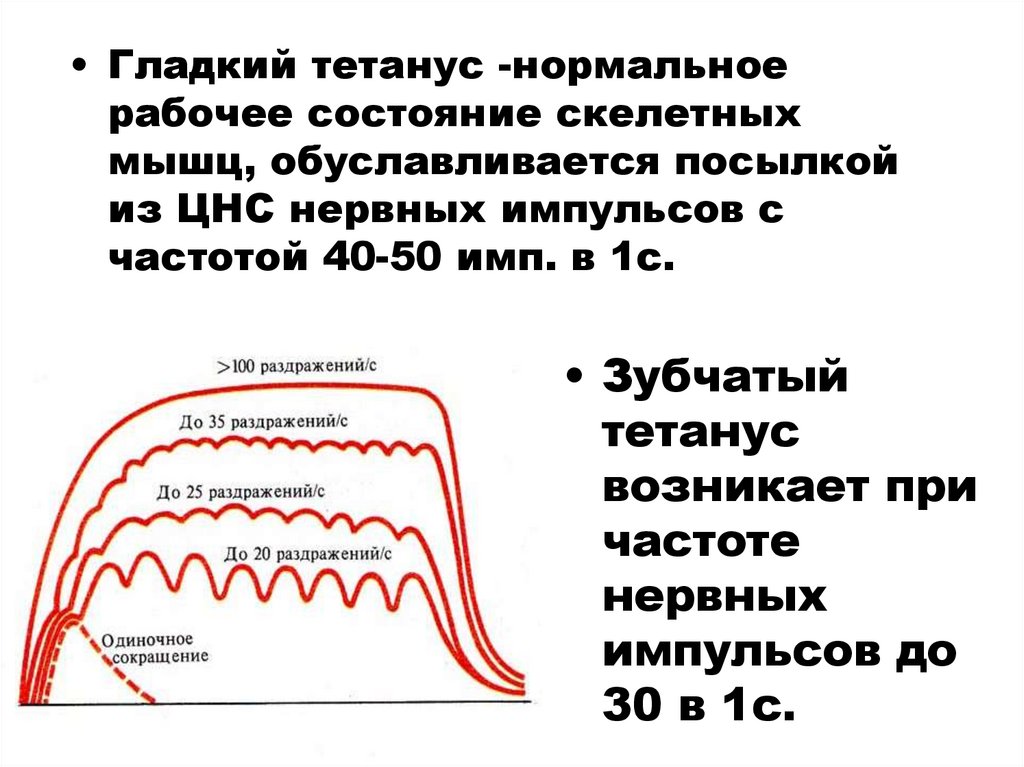
• Гладкий тетанус -нормальноерабочее состояние скелетных
мышц, обуславливается посылкой
из ЦНС нервных импульсов с
частотой 40-50 имп. в 1с.
• Зубчатый
тетанус
возникает при
частоте
нервных
импульсов до
30 в 1с.
79. При анализе графиков, обращает на себя внимание следующая особенность: амплитуда зубчатого и гладкого тетануса выше, чем
амплитудаодиночного мышечного сокращения.
Введенский
показал, что
увеличение
амплитуды
тетануса связано
с тем, что каждый
последующий
нервный импульс
попадает в стадию
экзальтации.
80.
• Оптимум - частота раздражения, прикоторой наблюдается суммарное
сокращение (тетанус) наибольшей
амплитуды. При этом каждый
последующий импульс попадает в
период экзальтации. Наилучшие
условия для сокращения мышцы
• Пессимум - частота раздражения, при
которой не наблюдается суммации
сокращения
При этом каждый последующий
импульс попадает в период
рефрактерности. Наихудшие условия
для сокращения мышцы
81.
82. ФИЗИОЛОГИЯ ГЛАДКИХ МЫШЦ
• гладкая (непроизвольная)мускулатура находятся в
стенках внутренних
органах, сосудах и коже
• обеспечивают функцию
полых органов: изгнание
содержимого из мочевого
пузыря, кишки, желудка,
желчного пузыря, матки
…
83. Гладкие мышцы
• Структурной единицей их являетсявытянутой формы клетка: длиной
20-400 мкм, толщиной 2-10 мкм.
• На мембране гладкомышечных
клеток, в отличие от скелетных,
имеются не только натриевые и
калиевые каналы, но и большое
количество кальциевых каналов.
84.
Гладкомышечная ткань• Тканевым
элементом
является
гладкомышечная клетка - миоцит
веретеновидной
или
звездчатой
формы.
• слабо
развиты
тропонинтропомиозиновый
комплекс
и
эндоплазматический
ретикулум,
(необходимый для инициирования
мышечного сокращения)
85.
• миофиламенты содержат актин имиозин, которые распределены менее
упорядочено, чем в волокнах скелетной
мышцы
• Плазматические мембраны очень
тесно примыкают друг к другу, образуя
между собой плотные контакты нексусы, через которые возбуждение
электротонически распространяется от
клетки к клетке.
• функционируют как синцитий —
функциональное образование, в
котором возбуждение способно
беспрепятственно передаваться с
одной клетки на другую.
86. Гладкомышечная ткань (физиологические особенности)
• Потенциал покоя -30 до -70 мВ• Потенциал действия двух
типов: пикоподобные и плато.
Ионный механизм ПД
определяется Са2+
87. Разновидности деполяризации гладко-мышечных клеток
Разновидности деполяризации гладкомышечных клеток• Один из них (а) напоминает ПД
скелетной мышцы, отличаясь от
нее большей продолжительностью
(10-50 мс). Этот ПД возникает при
воздействии на клетку многих
раздражителей: нервного
импульса, гормонов,
электрического тока.
• В мышцах стенки желудочнокишечного тракта изменение
заряда мембраны происходит по
типу (Б). В них развивается
спонтанная (без действия какихлибо посторонних факторов)
медленная деполяризация.
• В сердечной мышце ПД имеет
замедление
реполяризации(плато) за счет
входа ионов Са2+ в клетку (В)
88. Свойства гладких мышц
• сила сокращения зависит от степенирастяжения мышцы (прямопропорционально)
• пластичность - способность сохранять
приданную им при растяжении длину без
напряжения.
• автоматия - спонтанные сокращения ГМК,
благодаря наличию водителей ритма
(пейсмекеры). ПД пейсмекерных клеток
создает миогенный тонус гладких мышц.
• сокращение индуцируется вегетативной
нервной системой с помощью
медиаторов, гормонов, БАВ
89.
Важной особенностью висцеральной гладкоймышцы многих полых органов является ее
способность
изменять
напряжение
без
закономерной связи с ее длиной. Например,
внезапное
увеличение
объема
жидкости
в
полости
мочевого
пузыря,
сопровождается
растяжением гладкой мышцы его стенки, что
обязательно
приводит
к
увеличению
внутрипузырного
давления.
Однако
в
последующие от 15 сек до нескольких минут,
несмотря
на
постоянно
действующую
растягивающую силу внутрипузырное давление
возвращается к почти исходному значению и
сокращение не наступает. Такую способность
сохранения при растяжении длину, без изменения
напряжения называют пластичностью ГМ.
90.
Классификациямышц:
гладких
Мультиунитарные,
входят
в
состав цилиарной мышцы, мышц
радужки
глаза,
мышцы
поднимающей волос.
Унитарные
(висцеральная),
находятся во всех внутренних
органах,
протоках
пищеварительных
желез,
кровеносных и лимфатических
сосудах, коже.
91.
Мультиунитарная гладкая мышца.состоит
из
отдельных
гладкомышечных клеток, каждая из
которых, находится независимо друг от
друга;
имеет большую плотность иннервации;
как и поперечнополосатые мышечные
волокна, снаружи покрыты веществом,
напоминающим базальную мембрану, в
состав которого входят, изолирующие
клетки друг от друга, коллагеновые и
гликопротеиновые волокна;
каждая
мышечная
клетка
может
сокращаться отдельно и ее активность
регулируется нервными импульсами;
92. Мультиунитарная гладкая мышца
93.
Унитарная гладкая мышца (висцеральная).представляет собой пласт или пучок, а
сарколеммы отдельных миоцитов имеют
множественные точки соприкосновения.
мембраны рядом расположенных клеток
образуют множественные плотные контакты
(gap junctions), через которые ионы имеют
возможность свободно передвигаться из
одной клетки в другу
потенциал действия, возникающий на
мембране
гладкомышечной
клетки,
и
ионные потоки могут распространяться по
мышечному
волокну,
обеспечивая
возможность одновременного сокращения
большого количества отдельных клеток.
Данный тип взаимодействия известен как
функциональный синцитий.
94. Унитарная (висцеральная) гладкая мышца
Gapjunctions
95. Пейсмекеры
• Среди гладкомышечных клеток, образующихфункциональный синцитий, имеются такие,
которые обладают пейсмекерными свойствами
(от англ. рacemaker - задающий темп). Их
мембрана обладает высокой спонтанной
проницаемостью к ионам (в первую очередь к
кальцию), поэтому у них фактически отсутствует
мембранный потенциал покоя. После
предшествующей реполяризации
самопроизвольно, без действия раздражителя,
благодаря проникновению внутрь ионов кальция
начинается постепенная деполяризация
мембраны. При достижении критического
уровня этот препотенциал переходит в
потенциал действия. Данный потенциал с
помощью нексусов передается соседним
клеткам.
96.
Секреция – процессобразования внутри клетки
(гландулоцита) из веществ,
поступающих в неё, и
выделения из клетки
специфического продукта
(секрета) определённого
функционального
назначения
97.
• Гландулоциты выделяют различныепо химической природе продукты:
белки, липиды, мукополисахариды,
растворы солей, оснований, кислот.
• Собственный секрет - продукт
метаболизма клетки,
• экскрет – продукт её катаболизма,
• рекрет – вещество,которое клека
поглотила и выделила в
неизменённом виде.
• Эндосекреция – выделение секрета
ч/з базальную мембрану
непосредственно в кровь или лимфу.
98.
Для синаптических окончаний нагландулоцитах характерны широкие
синаптические щели, заполненные
интерстициальной жидкостью.
Сюда из окончаний нейронов
поступают медиаторы: из крови –
гормоны, из соседних эндокринных
клеток – парагормоны, от самих
гландулоцитов – продукты их
деятетельности.
99.
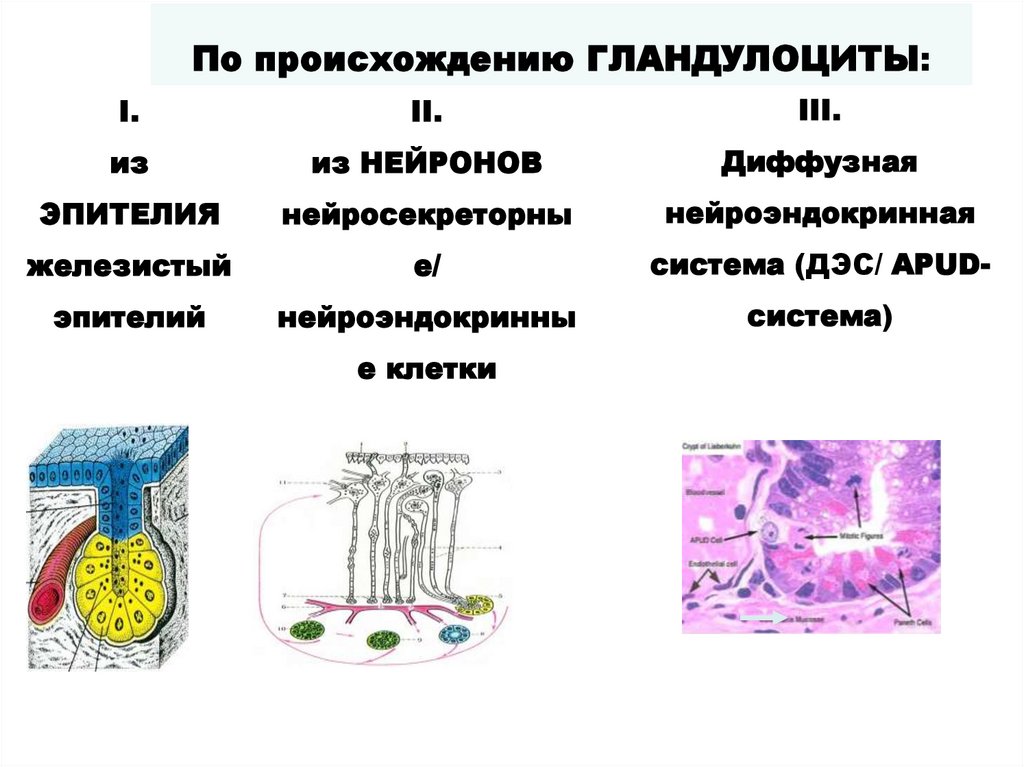
По происхождению ГЛАНДУЛОЦИТЫ:I.
II.
III.
из
из НЕЙРОНОВ
Диффузная
ЭПИТЕЛИЯ
нейросекреторны
нейроэндокринная
железистый
е/
система (ДЭС/ APUD-
эпителий
нейроэндокринны
система)
е клетки
100.
APUD-система (АПУД-система, диффузнаянейроэндокринная система) — система
клеток, имеющих общего эмбрионального
предшественника и обладающих
способностью синтезировать, накапливать и
секретировать биогенные амины и/или
пептидные гормоны.
.
101.
В настоящее время идентифицировано около 60 типовклеток APUD-системы (апудоциты), которые
встречаются в:
• ЦНС: гипоталамусе, мозжечке;
• симпатических ганглиях;
• железах внутренней секреции: аденогипофизе,
шишковидном теле, щитовидной железе, тимусе,
островках поджелудочной железы, надпочечниках,
яичниках;
• желудочно-кишечном тракте;
• эпителии дыхательных путей и легких;
• почках;
• коже;
• мочевых путях;
• плаценте
102. Биопотенциалы гландулоцитов
Биопотенциалы секреторных клетокимеют ряд особенностей в покое и
при секреции:
• низкую величину и скорость
изменения
• градуальность
• различную поляризованность
базальной и апикальной мембран.
103. Биопотенциалы гландулоцитов
• Мембранный потенциал гландулоцитовразличных экзокринных желез в
состоянии относительного покоя равен от
—30 до —75 мВ.
• Стимуляция секреции меняет
мембранный потенциал. Это изменение
поляризованности мембраны называется
секреторным потенциалом.
• Оптимальной для возникновения
секреторных потенциалов считается
поляризованность мембран, равная —50
мВ.
104.
Секреция железконтролируется:
1. Нервными
механизмами,
2. Гуморальными
механизмами,
3. Паракринными
механизмами.
В результате действия этих
механизмов происходит:
• УСИЛЕНИЕ СЕКРЕЦИИ
или
• ТОРМОЖЕНИЕ
СЕКРЕЦИИ.
105.
Особенностимышц у
новорожденных
и детей грудного
возраста
106.
У новорожденных анатомическиимеются все скелетные мышцы.
Количество мышечных волокон с
возрастом в них не увеличивается.
Рост мышечной массы происходит
за счет увеличения размеров
миофибрилл, которые оттесняют
ядра на периферию. Количество
ядер уменьшается, а их размеры
возрастают. Одновременно
утолщается сарколемма. Размеры
мышечных волокон
стабилизируются к 20-ти годам.
107.
Количество волокон в мышцеустанавливается через 4-5
месяцев после рождения и
затем практически не
меняется на протяжении
всей жизни. При рождении
толщина их составляет
примерно 1/5 толщины
волокон у взрослых людей.
108.
Возбудимость мышц новорожденногоочень низка. Показателем этого
является высокая хронаксия и
большой порог деполяризации. У
новорожденного потенциал покоя
миоцитов составляет -20-40 мВ.
Низкий уровень мембранного
потенциал покоя в раннем возрасте
связан с меньшей активностью
Na+/K+-АТФ-азы, в результате чего
внутриклеточное содержание ионов
Na+ выше, а ионов К+ – ниже, чем у
взрослых.
109.
Трансмембранная разность ионов К+и Nа+ невысока, поэтому и
величина потенциала действия
небольшая. Отмечается большая
длительность фаз абсолютной и
относительной рефрактерности. В
процессе роста проницаемость
мембраны уменьшается, работа
ионных насосов улучшается и
увеличивается потенциал покоя и
потенциал действия.
110.
Лабильность у детей ниже, чем увзрослых вследствие большой
длительности рефрактерных фаз. В
связи с этим скелетные мышцы
новорожденных не способны
развивать тетанус, мышечные
сокращения носят тонический
характер. С возрастом происходит
укорочение фаз абсолютной и
относительной рефрактерности и,
как следствие, возрастание
скорости проведения возбуждения и
увеличение быстроты движений.
111.
Проводимость у новорожденныхнизкая. Скорость проведения
потенциала действия с возрастом
увеличивается, что является
следствием увеличения толщины
мышечного волокна и амплитуды
потенциала действия, т. к.
снижается сопротивление
ионному току и быстрее
развивается возбуждение на
соседнем участке мембраны
112.
Сократимость у новорожденныхпонижена. Фазы укорочения и
расслабления одиночного мышечного
сокращения новорожденного замедлены
и характеризуется большим временем
сокращения. В мышцах детей быстрее
накапливаются продукты метаболизма,
поэтому тетанус имеет пологое начало и
постепенное расслабление. Мышцы
отвечают тоническим сокращением на
раздражители любой частоты и
сокращаются без пессимального
торможения все время действия
раздражителя, что обусловлено
недостаточностью зрелости
мионевральных синапсов.
113.
Эластичность мышц у детейвысокая, они сильнее
укорачиваются при
сокращении и удлиняются в
момент растяжения.
Эластичность мышц с
возрастом уменьшается, а
упругость и прочность,
наоборот, увеличиваются.
114.
В 4-5 лет быстрее развиваютсямышцы предплечья. Усиленное
развитие мышц кисти происходит в
6-7 лет. При этом скорость развития
сгибателей превышает созревание
разгибателей. В раннем детском
возрасте повышен тонус мышц
сгибателей кистей рук и
разгибателей бедра. В дальнейшем
распределение тонуса
нормализуется.
115.
У новорожденныхотсутствует
дифференциация моторных
единиц на быстрые и
медленные, но уже с первых
дней жизни у ребенка
начинается постепенная
дифференцировка,
характерная для взрослых.