






 Биология
БиологияПохожие презентации:
")
")

 Подцарство Одноклеточные, или Простейшие (Protozoa)")
")
. Подтип Жгутиконосцы (Mastigophora)")
. Тип Саркомастигофоры (Sarcomastigophora)")



Protozoa. Сравнение размеров одноклеточного (Actinosphaerium sp., Heliozoea) и многоклеточного (Rotatoria)
1. Protozoa
2. Protozoa
Земля населена множеством одноклеточных организмов. Хотя подавляющее ихбольшинство составляют прокариоты, существуют и одноклеточные эукариоты,
громадное число видов и особей которых едва ли поддается оценке. Они обитают главным образом в воде, причем небольшие размеры позволяют им заселять мельчайшие
пространства, наполненные влагой, такие, например, как поры в почве. Маленькие
размеры - это ещë и существенная причина того, почему среди них есть много
паразитических (особенно эндопаразитических) таксонов.
Размеры самых мелких одноклеточных, таких как спорозоиты Apicomplexa, составляют
лишь несколько микрометров. Размеры самых крупных форм измеряются сантиметрами:
таковы некоторые фораминиферы, образующие раковины, например современные
Cycloclypeus carpenteri (диаметр до 13 см) и ископаемые виды Nummulites (32 см в
диаметре!). Tаким образом, разница в размерах одноклеточных может быть
десятитысячекратной, в то время как для млекопитающих этот показатель составляет лишь
750 раз (синий кит – этрусская карликовая бурозубка).
Илл. 1. Сравнение размеров одноклеточного
(Actinosphaerium sp., Heliozoea)
и многоклеточного (Rotatoria).
3.
Некоторые одноклеточные превосходят по размеру ряд многоклеточных животных(илл.1). Особенно нагляден пример организмов из пор между песчинками морского пляжа: в
одной такой пробе можно найти, например, инфузорий из родов Loxophyllum или (длина до 2
мм) наряду с полихетами из рода Diurodrilus (дли- на 250–450 мкм).
Ещë в 1838 г. Эренберг (1795–1876) в своëм труде «Мельчайшие обитатели настоев
(Infusionsthierchen) как совершенные организмы», не потерявшем значения и до нашего
времени, обосновал мнение о том, что одноклеточные представляют собой миниатюрных
микроскопических животных, имеющих свои прототипы в фауне, видимой невооружëнным
глазом. Он последовательно описал у одноклеточных желудочно-кишечный тракт
(пищеварительные вакуоли), сосудистую систему, слюнные железы (особые вакуоли или
зоохлореллы), яичко (макронуклеус) с семенными пузырьками (сократительными вакуолями)
и яичниками (микронуклеусы).
Зибольд в 1845 г. впервые дал определение Protozoa как животных, «самые разные
системы органов которых разделены нечëтко, а неправильная форма и простая организация
которых могут быть сведены к одной клетке». Сегодня мы знаем, что у существ с
одноклеточной организацией могут выполняться почти все те же функции, для которых
Metazoa необходимы многочисленные клетки в различных дифференцированных тканях и
органах.
4.
Bходе
эволюции
одноклеточных
сложилась
та
дифференцированная клеточная организация, которая нам
известна как эуцит (настоящая эукариотная клетка). У
первоначальных эукариот (Microspora, Archamoeba и
Tetramastigota, называемых также «Archezoa») митохондрий не
было; лишь Metakaryota приобрели их путем эндосимбиоза.
Tем самым, одноклеточные Metakaryota обладают всеми
частями и органеллами типичной эукариотной клетки, а именно, как минимум, одним ядром, митохондриями, иногда
пластидами, эндоплазматической сетью (ЭР), диктиосомами,
лизосомами, пероксисомами, аканто- сомами (coated vesicles),
рибосомами, микрофиламентами и микротрубочками. Как
любая
клетка,
организм
одноклеточного
окружëн
плазматической мембраной. Снаружи на ней часто находится
весьма об- ширный гликокаликс, который может иметь особую
дифференцированную структуру (илл. 2). Сверх того, как
«Archezoa», так и в особенности метакариотические
одноклеточные, имеют ряд органелл, представленных только у
них, главным образом, в тех или иных специфических
структурах: таковы, например, сократительные вакуоли,
экструсомы, аксостиль или парабазальный аппарат.
Илл. 2.
Гликокаликс
Vannella simplex
(Amoebozoa).
Различные
типы
филаментов (F1, F2) на
ячеистом
основном
слое (Gw); под ним
плазмалемма (Pl). Из
Hausmann и Stockem
(1972).
5.
B комплект даже самого простого эукариотного одноклеточного входят хромосомы и аппаратмитотического деления, так что у них имеется митоз как предпосылка бесполого (вегетативного)
размножения. Половые же процессы — образование гаплоидных клеток путем мейоза и их слияние —
представляют собой изначальный признак одноклеточных эукариот; он обнаружен уже у Micospora.
Cтроение
Большинство одноклеточных имеют свою специфическую форму, которая поддерживается внутриили внеклеточными скелетными элементами Такую опорную функцию выполняют, в частности,
микротрубочки. Она становится явной, когда они объединяются в пучки и служат опорой для сложных структур, таких, например, как аксоподии солнечников, аксостили Axostylata или опорный
палочковый аппарат эвглен и инфузорий; кроме того, они поддерживают в определенном положении
системы органоидов, на- пример, сократительных вакуолей у инфоузорий.
Стабилизация формы достигается также благо- даря полосам пелликулы эвглен или эпиплазме
некоторых инфузорий. У ряда групп скелет-ную функцию выполняют внутриклеточные элементы,
образованные из неорганических веществ (на- пример, скелеты радиолярий из кремнезëма).
Особняком стоит скелет из сульфата стронция, представленный только у Acantharea.
Форма псевдоподий.
A — лопастная, полиподиальная (Amoeba, Lobosea). Б —
коническая полиподиальная (Mayorella, Lobosea). B — лопастная,
моноподиаль- ная (Saccamoeba, Lobosea). Г — вееровидная
(Vannella, Lobosea). Д — нитевидная (Nuclearia, Filosea). Е —
сетевидная (Allogromia, Forami- niferea). Ж — лопастная,
организм покрыт оболочкой (Nebela, Lobo- sea). З — нитевидная,
организм покрыт оболочкой (Cyphoderia, Fi- losea). И — лучистое
расположение (Actinophrys, Heliozoea). К — разветвлëнная
(Stereomyxa, Acarpomyxea). Из Hausmann (1985).
6.
Весьма обычны и внеклеточные чешуйки, оболочки или иные покровы из органического инеорганического материала, участвующие в поддержании формы; они образуются, главным
образом, в эндоплазматической с ети или в диктиосомах и выводятся наружу посредством
экзоцитоза.
Органеллы движения и локомоция
Большинство одноклеточных организмов могут активно передвигаться. В настоящее время известно
два механизма, имеющих самое общее распространение и обеспечивающих подвижность: это (1)
движение цитоплазмы и (2) движение жгутиков и ресничек.
Движение протоплазмы (движение посредством псевдоподий), особенно отчëтливо выраженное у
амебоидных клеток, создается сократительной системой, молекулярную основу которой образуют
белки актин и миозин. У амëбоидных одноклеточных эти белки упорядочены в трëхмерную сеть в
периферических областях клетки (илл. 5). При локальном и согласованном сокращении этой сети
возни- кает ток цитоплазмы, обусловливающий передвижение клетки. Различные типы псевдоподий
(ложноножек) показаны на илл. 4.
Основной принцип сокращения, вероятно, тот же, что и в поперечнополосатой мускулатуре
млекопитающих: благодаря активности миозиновых головок, актиновые и миоцентриолей (илл. 7, 8).
Не каждое базальное тело несëт аксонему. Часто базальные тела представляют собой окончания
корешковых структур микротубулярной или микрофиламентозной природы, которые могут быть
втянуты глубоко в клетку.
Ил. 5 Сеть кортикальных
филаментов у Amoeba proteus
(Amoebozoa); у верхнего края
рисунка косо срезанная
плазматическая мембрана.
Илл. 7.
Кинетиды инфузорий.
A — продольный срез. Б —
поперечный срез. A — из
Margulis,
McKann
и
Olendzenski (1993); Б — из
Lynn (1995).
7.
Их строение и функции различны у разных таксонов. Особое таксономическоезначение имеют структуры, связанные с базальными телами жгутиков и ресничек (илл. 7,
8). Корешковые структуры заякоривают базальные тела, испытывающие механическое
усилие при ударе реснички или жгутика. B определëнных пределах они могут отвечать и
за поддержание формы клетки. Наконец, при делении клетки базальные тела, в качестве
центриолей, могут служить центрами образования многочисленных микротрубочек, из
которых собирается веретено митотического деления. Далеко не всегда все жгутики либо
реснички одной клетки устроены совершенно идентично: в этих случаях имеет место
анизоконтия (гетероконтия). У Ochromonas, например, один, более длинный, жгутик
вытянут вперед, тогда как другой изогнут и направлен назад (илл. 9); при этом у
короткого жгу- тика есть базальное утолщение, а длинный несëт мастигонемы —
волосовидные придатки, расчленëнные на три отдела (илл. 10). Эти два жгутика предназначены для различных задач: длинный служит для передвижения и, у фаготрофных
видов, для захвата пищи, в то время как короткий может, например, участвовать в
фототаксисе. Строение и расположение жгутиков может быть использовано для целей
систематики.
Илл. 8. Базальные тельца (кинетосомы)
жгутиков (А, Б) и ресничек (B) и
корешковые структуры, связанные с ними.
A, Б — Chilomonas paramecium
(Cryptomonada). B — Climacostomum virens
(Ciliophora). А – х 22 000, B - х77 000, С –
х 66 000. А,
Б — оригиналы K. Hausmann, Берлин; B —
оригинал D. Fischer-Defoy, Дармштадт.
8.
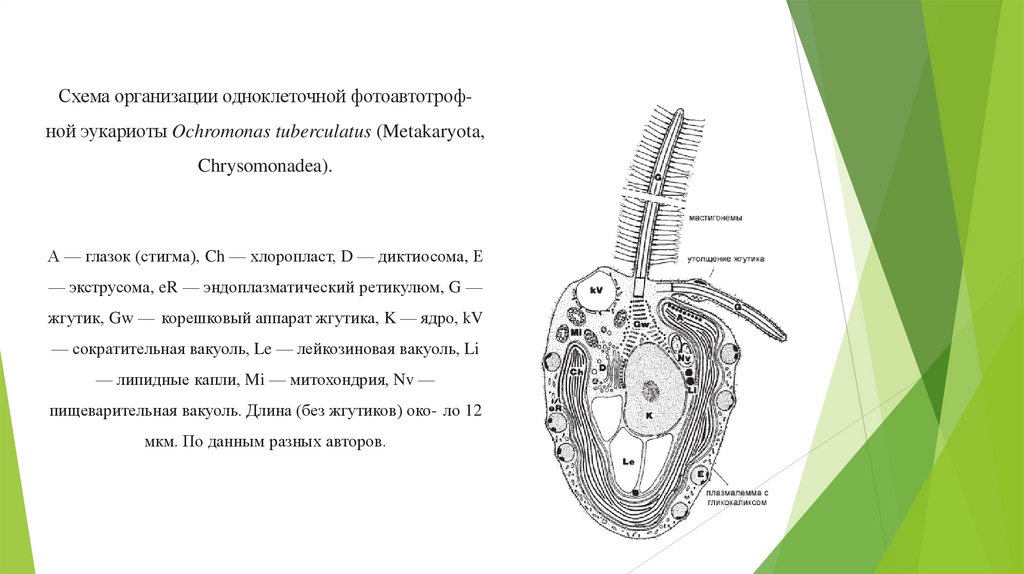
Схема организации одноклеточной фотоавтотрофной эукариоты Ochromonas tuberculatus (Metakaryota,Chrysomonadea).
А — глазок (стигма), Ch — хлоропласт, D — диктиосома, Е
— экструсома, eR — эндоплазматический ретикулюм, G —
жгутик, Gw — корешковый аппарат жгутика, K — ядро, kV
— сократительная вакуоль, Le — лейкозиновая вакуоль, Li
— липидные капли, Mi — митохондрия, Nv —
пищеварительная вакуоль. Длина (без жгутиков) око- ло 12
мкм. По данным разных авторов.