






































 Биология
БиологияПохожие презентации:










Строение генов эукариот
1.
Строение геновэукариот
2.
Гено́м — совокупность наследственного материала, заключённого в клеткеорганизма. Геном содержит биологическую информацию, необходимую для
построения и жизни организма.
Большая часть геномов, в том числе геном человека и геномы всех остальных
клеточных форм жизни, построены из ДНК, однако некоторые вирусы имеют
геномы из РНК.
3.
В геноме человека, так же как и у других млекопитающих, участки ДНК,кодирующие белки - ГЕНЫ, составляют менее 2 % от всей длины
хромосом.
Остальную, большую часть ДНК раньше называли избыточной, но теперь
стало ясно, что она выполняет очень важные регуляторные функции,
определяя, в каких клетках и когда должны функционировать те или иные
гены.
У более просто организованных прокариотических организмов, геном
которых представлен одной кольцевой молекулой ДНК, на кодирующую
часть приходится до 90 % от всего генома.
4.
Ген — это наследственный фактор, который несет информацию о конкретномпризнаке или функции организма и является структурной и функциональной
единицей наследственности.
Основные характеристики гена
Структура: Ген представляет собой участок молекулы ДНК, который кодирует
информацию для синтеза белка или функциональной РНК. Гены могут содержать
экзоны (кодирующие участки) и интроны (некодирующие участки), которые играют
роль в регуляции экспрессии гена.
Функция: Гены определяют физические и биологические признаки организма,
включая цвет глаз, рост и предрасположенность к заболеваниям. Они управляют
синтезом белков, которые выполняют различные функции в клетках и тканях.
Наследственность: Гены передаются от родителей к потомству и являются
основой наследования фенотипических признаков. Большинство признаков
определяется множеством генов, что делает их полигенными.
Изменчивость: Гены могут изменяться в результате мутаций, что приводит к
появлению различных аллелей — вариантов одного и того же гена. Эти аллели
могут кодировать разные версии белка, что может проявляться в фенотипе.
5.
В среднем на один ген в хромосоме человека приходится около 50 тыс.нуклеотидов. Существуют очень короткие гены. Например, белок энкефалин,
который синтезируется в нейронах головного мозга и влияет на формирование
наших положительных эмоций, состоит всего из 5 аминокислот. Следовательно,
ген, отвечающий за его синтез, содержит всего около двух десятков нуклеотидов.
А самый длинный ген, кодирующий один из мышечных белков, состоит из 2,5 млн
нуклеотидов.
6.
Размер генома человека составляет около 3 миллиардов пар оснований, чтосоответствует 23 парам хромосом, включая 22 аутосомы и одну пару половых
хромосом (X и Y).
Структура генома
Хромосомы: Геном человека состоит из 23 пар хромосом, каждая из которых содержит
одну молекулу ДНК. Длина всех молекул ДНК в гаплоидном наборе составляет
примерно 3,3 миллиарда пар оснований.
Гены: В геноме человека содержится около 20 000 - 26 000 генов, которые кодируют
белки. Однако только около 1,5% генома кодирует белки или функциональные РНК.
Митохондриальная ДНК: В геноме также присутствует митохондриальная ДНК,
которая наследуется по материнской линии и составляет менее 1% от всей ДНК.
Повторяющиеся последовательности: Около трети генома составляют повторяющиеся
последовательности различной длины, которые играют важную роль в регуляции
активности генов.
7.
В одной хромосоме человека находится одна молекула ДНК. В 46 хромосомах — 46молекул ДНК.
Длина всех молекул ДНК одной клетки человека — около 2 м.
Тело человека состоит примерно из 5 × 10¹³ клеток. Общая длина ДНК всех клеток — 1
× 10¹¹ км, что почти в тысячу раз больше расстояния от Земли до Солнца.
ДНК состоит из нуклеотидов. Гаплоидный набор ДНК одной клетки содержит 3,2 × 10⁹
пар нуклеотидов.
В одной молекуле ДНК сотни генов.
8.
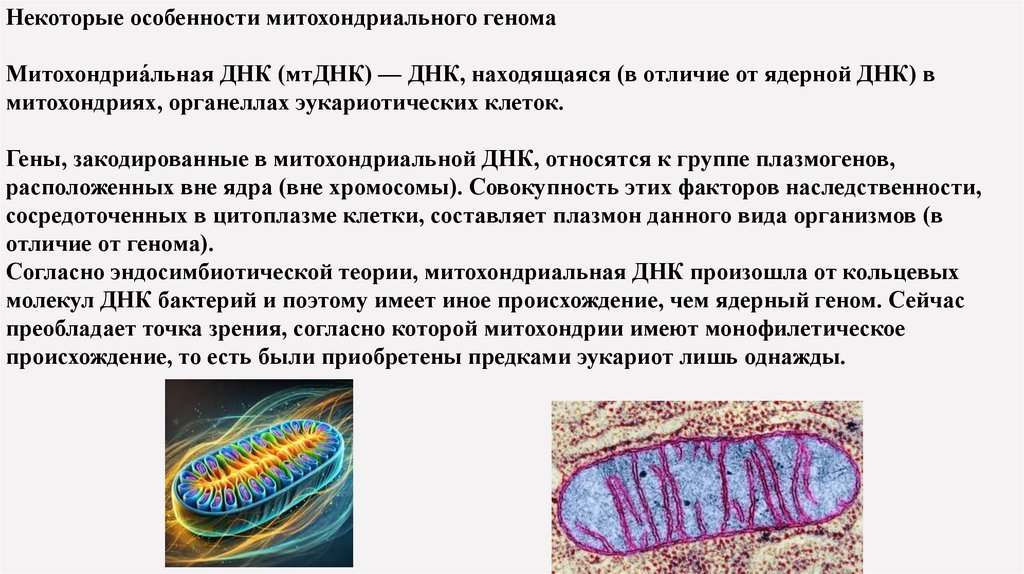
Некоторые особенности митохондриального геномаМитохондриа́льная ДНК (мтДНК) — ДНК, находящаяся (в отличие от ядерной ДНК) в
митохондриях, органеллах эукариотических клеток.
Гены, закодированные в митохондриальной ДНК, относятся к группе плазмогенов,
расположенных вне ядра (вне хромосомы). Совокупность этих факторов наследственности,
сосредоточенных в цитоплазме клетки, составляет плазмон данного вида организмов (в
отличие от генома).
Согласно эндосимбиотической теории, митохондриальная ДНК произошла от кольцевых
молекул ДНК бактерий и поэтому имеет иное происхождение, чем ядерный геном. Сейчас
преобладает точка зрения, согласно которой митохондрии имеют монофилетическое
происхождение, то есть были приобретены предками эукариот лишь однажды.
9.
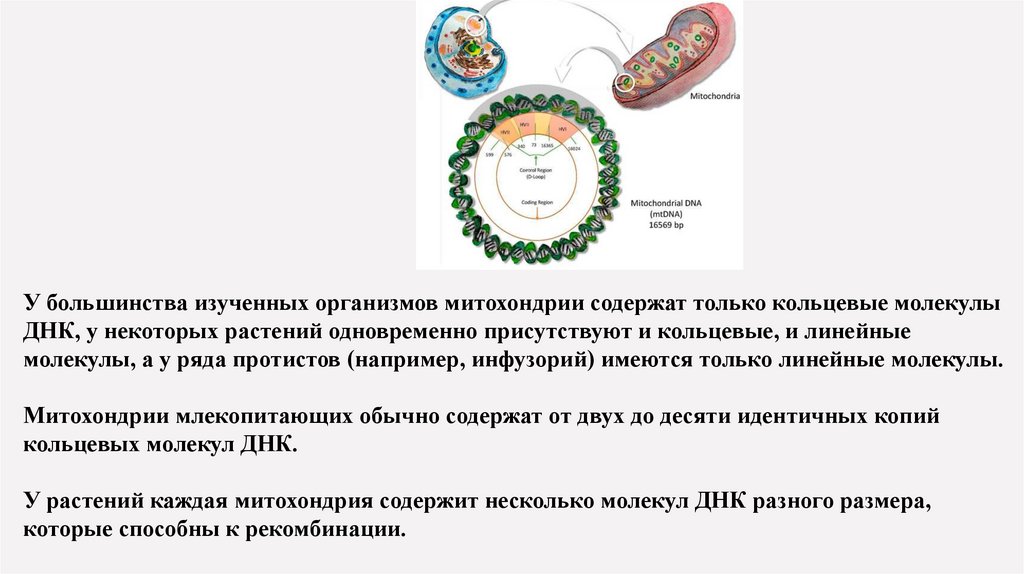
У большинства изученных организмов митохондрии содержат только кольцевые молекулыДНК, у некоторых растений одновременно присутствуют и кольцевые, и линейные
молекулы, а у ряда протистов (например, инфузорий) имеются только линейные молекулы.
Митохондрии млекопитающих обычно содержат от двух до десяти идентичных копий
кольцевых молекул ДНК.
У растений каждая митохондрия содержит несколько молекул ДНК разного размера,
которые способны к рекомбинации.
10.
У большинства многоклеточных организмов митохондриальная ДНК наследуется по материнской линии. Яйцеклеткасодержит на несколько порядков больше копий митохондриальной ДНК, чем сперматозоид. В сперматозоиде обычно
не больше десятка митохондрий (у человека — одна спирально закрученная митохондрия), в небольших яйцеклетках
морского ежа — несколько сотен тысяч, а в крупных ооцитах лягушки — десятки миллионов. Кроме того, обычно
происходит деградация митохондрий сперматозоида после оплодотворения.
При половом размножении митохондрии, как правило, наследуются исключительно по материнской линии,
митохондрии сперматозоида обычно разрушаются после оплодотворения. Большая часть митохондрий
сперматозоида находятся в основании жгутика, которое при оплодотворении иногда теряется.
Было обнаружено, что митохондрии сперматозоидов помечены убиквитином (белком-меткой, которая приводит к
разрушению отцовских митохондрий в зиготе).
Так как митохондриальная ДНК не является высококонсервативной и имеет высокую скорость мутирования, она
является хорошим объектом для изучения филогении (эволюционного родства) живых организмов. Для этого
определяют последовательности митохондриальной ДНК у разных видов и сравнивают их при помощи специальных
компьютерных программ и получают эволюционное древо для изученных видов. Исследование митохондриальных
ДНК собак позволило проследить происхождение собак от диких волков. Исследование митохондриальной ДНК в
популяциях человека позволило вычислить «митохондриальную Еву», гипотетическую прародительницу всех
живущих в настоящее время людей.
11.
Для некоторых видов показана передача митохондриальной ДНК по мужской линии, например, у мидий.Наследование митохондрий по отцовской линии также описано для некоторых насекомых, например, для
дрозофилы,медоносных пчел и цикад.
Существуют также данные о митохондриальном наследовании по мужской линии у млекопитающих. Описаны
случаи такого наследования для мышей,при этом митохондрии, полученные от самца, впоследствии отторгаются.
Такое явление показано для овец и клонированного крупного рогатого скота.
Наследование по отцовской линии у людей
До недавнего времени считалось, что митохондрии человека наследуются только по материнской линии. Был
известен лишь один-единственный случай пациента, у которого в 2002 году достоверно обнаружили отцовскую
митохондриальную ДНК.
Лишь недавнее исследование 2018 года показало, что митохондриальная ДНК человека иногда всё же может
передаваться и по отцовской линии. Небольшое количество митохондрий отца может попасть в яйцеклетку
матери вместе с цитоплазмой сперматозоида, но, как правило, отцовские митохондрии после этого из зиготы
исчезают. Однако было обнаружено, что у некоторых людей существует «мутация, которая помогает выживать
митохондриям отца».
Кодирующие последовательности (кодоны) митохондриального генома имеют некоторые отличия от
кодирующих последовательностей универсальной ядерной ДНК.
Так, кодон AUA кодирует в митохондриальном геноме метионин (вместо изолейцина в ядерной ДНК), кодоны
AGA и AGG — терминаторные кодоны (в ядерной ДНК кодируют аргинин), кодон UGA в митохондриальном
геноме кодирует триптофан
12.
Международный проект Геном человекаДжеймс Уотсон в 1988 г. инициировал создание международного проекта
«Геном человека».
Цель проекта — выяснить последовательности азотистых оснований и
положения генов (картирование) в каждой молекуле ДНК каждой клетки
человека, что открыло бы причины наследственных заболеваний и пути к
их лечению.
26-го июня 2000 года. На совместной пресс-конференции с участием
президента США и премьер-министра Великобритании представители двух
исследовательских групп — International Human Genome Sequencing
Consortium (IHGSC) и Celera Genomics — объявили о том, что работы по
расшифровке генома человека, начавшиеся ещё в 70-х годах, успешно
завершены, и черновой его вариант составлен. Начался новый эпизод
развития человечества — постгеномная эра.
13.
J. Craig Venter, Mark D. Adams,Eugene W. Myers, Peter W. Li, Richard
J. Mural, et. al.. (2001). The Sequence
of the Human Genome. Science. 291,
1304-1351
Eric S. Lander, Lauren M. Linton,
Bruce Birren, Chad Nusbaum,
Michael C. Zody, et. al.. (2001).
Initial sequencing and analysis of the
human genome. Nature. 409, 860-921
Гонка за генóм», в которой участвовали межгосударственная и частная компании,
формально завершилась «ничьей»: обе группы исследователей опубликовали свои
достижения практически одновременно. Руководитель частной компании Celera
Genomics Крейг Вентер опубликовал свою работу в журнале Science в соавторстве с
~270 учёными, работавшими под его началом. Работа, выполненная международным
консорциумом по секвенированию человеческого генома (IHGSC), опубликована в
журнале Nature, и полный список авторов насчитывает около 2800 человек, работавших
в почти трёх десятках центров по всему миру
14.
Проект состоял из пяти основных этапов:• составление карты, на которой помечены гены, отстоящие
друг от друга не более чем на 2 млн оснований, на языке
специалистов – с разрешением 2 Мб (мегабаза — от англ.
слова «base» — основание);
• завершение физических карт каждой хромосомы с
разрешением 0,1 Мб;
• получение карты всего генома в виде набора описанных по
отдельности клонов (0,005 Мб);
• полное секвенирование ДНК (разрешение 1 основание);
• нанесение на карту с разрешением в 1 Мб основание всех
генов человека.
15.
Международный проект Геном человекаСледует отметить, что это один из самых дорогих научных проектов в истории
изучения генетики. В проекте заняты тысячи специалистов из разных стран мира –
биологи, химики, математики, физики и технические специалисты.
На реализацию проекта было потрачено в 1990 г. 60 млн долл., в 1991 г. — 135 млн, в
1992–1995 гг. — от 165 до 187 млн в год.
Самый значительный вклад в финансирование этого проекта внесли США,
Великобритания, Германия, Франция и Япония. Только США израсходовали в 1996–
1998 гг. соответственно 200, 225 и 253 млн долл.
Ученые разных стран вели исследования, финансируемые из государственных
16.
Международный проект Геном человекаПо мнению специалистов, 85% информации абсолютно достоверны, т.е. последовательность ДНК в
этом объеме перепроверена не один раз, и разночтения больше не выявляются.
Среди наиболее значимых результатов расшифровки генома человека следует выделить следующие:
• определено примерное число генов человека, их оказалось 23 000, а не 80 000, как предполагалось
ранее;
• генетические инструкции по формированию личности занимают меньше двух с половиной
сантиметров на двухметровой ленте ДНК, заключенной практически внутри каждой клетки тела.
Что удивляет самих ученых – насколько малая часть человеческого генома напрямую участвует в
построении организма;
• количество генов, несущих эти инструкции, – всего в пять раз больше, чем нужно для взращивания
мухи;
• из 3 млрд генетических букв, составляющих человеческие гены, которые образуют ДНК, 99,9%
одни и те же. Всего одна десятая процента и есть наша индивидуальность, что делает нас теми, кто
мы есть – красивыми и не очень, здоровыми или больными, умными или глупыми, добрыми или,
наоборот, жестокими;
• женская яйцеклетка является и главным источником эволюционных новаций;
• основную ответственность за генетические ошибки несет мужская сперма, в которой содержится
вдвое больше мутаций, чем в женской яйцеклетке.
17.
Международный проект Геном человекаКроме того, реализация международного проекта «Геном человека» дала толчок развитию
передовых технологий в самых разных отраслях, привела к появлению новых подходов в
изучении вирусологии, иммунологии, фармакологии и медицине.
Появилась новая отрасль - фармокогенетика.
Достижения генетиков могут с успехом применяться в криминалистике и судебной
медицине для идентификации личности. Разработан метод «генетической дактилоскопии».
По последовательностям ДНК можно устанавливать степень родства людей, а по
митохондриальной ДНК – точно устанавливать родство по материнской линии.
Параллельно с расшифровкой генома человека на базе тех же современных методов были
полностью прочитаны геномы таких классических генетических объектов изучения, как
муха дрозофила и круглый червь нематода.
Тем самым положено начало созданию единого геномного информационного поля, что
чрезвычайно важно как для изучения функции тех или иных генов, так и для понимания
механизма эволюции.
18.
Международный проект Геном человекаОказалось, что человек незначительно отличается по
сложности от червя, имеющего в своем геноме 20 тыс.
генов. Гены, выполняющие сходные функции и у
дрозофилы, и у червя, и у человека, имеют много общего.
Техника расшифровки структуры генома позволила
прочитать генетические коды более 30 патогенных
микроорганизмов, в том числе возбудителей чумы, холеры,
других вирусов. Найден ген, мутация которого может
защитить человека от заражения вирусом иммунодефицита.
Однако следует подчеркнуть, что исследования по
определению последовательности нуклеотидов в ДНК,
которые были объявлены как завершенные, это еще не
расшифровка генома.
19.
Все десятки тысяч генов не работают одновременно в каждой клетке многоклеточногоорганизма, этого не требуется.
Существующая специализация между клетками определяется избирательным
функционированием определённых генов. Мышечной клетке не надо синтезировать
кератин, а нервной – мышечные белки.
Хотя надо отметить, что существует довольно большая группа генов, которые работают
практически постоянно во всех клетках.
Это гены, в которых закодирована информация о белках, необходимых для осуществления
жизненно важных функций клетки, таких как редупликация, транскрипция, синтез АТФ и
многие другие. (гены домашнего хозяйства).
20.
21.
22.
23.
СВОЙСТВА ГЕНОВ:1. Дискретность – имеет определенный размер и позицию (локус)
2. Лабильность (может мутировать)
3. Стабильность (однако мутирует достаточно редко)
4. Специфичность (ген кодирует конкретный белок или близкородственные белки)
5. Аллельность – (в результате мутаций возникают варианты – аллели)
6. Плейотропность – (множественность действия)
7. Дозированность – (чем больше экземпляров гена в генотипе (доз), тем сильнее эффект гена).
8. Ген имеет сложную структуру;
9. Информация записана в гене посредством универсального генетического кода;
10. Наименьшая мутирующая единицей гена – пара нуклеотидов (SNP, single nucleotide
polymorphisms).
11. В геноме имеются разные типы (группы) генов;
24.
Существует несколько подходов к классификации генов, каждый из которых отражает особенностиих функционирования в процессе онтогенеза.
Гены, как единицы функции наследственного материала, разделяются на:
- структурные,
- регуляторные,
- гены-модуляторы.
Структурные гены содержат информацию о структуре белка (полипептидов) и рибонуклеиновых
кислот (рибосомальной и транспортной), при этом генетическая информация реализуется в процессе
транскрипции и трансляции или только транскрипции.У человека насчитывается около 30 000
структурных генов, но только часть из них экспрессируются, то есть реализуют генетическую
информацию, закодированную в них.
Регуляторные гены (ген - регулятор лактозного оперона, ген TFM и др.) координируют активность
структурных генов на уровне клетки, а также дерепрессию и репрессию генов на уровне организма.
Наряду с регуляторными генами, имеются регуляторные последовательности (промотор, оператор,
терминатор, энхансеры, сайленсеры, элемент перед промотором), функция которых выявляется во
взаимодействии со специфическими белками.
Гены-модуляторы усиливают или ослабляют действие структурных генов, изменяя их
функциональную активность.
25.
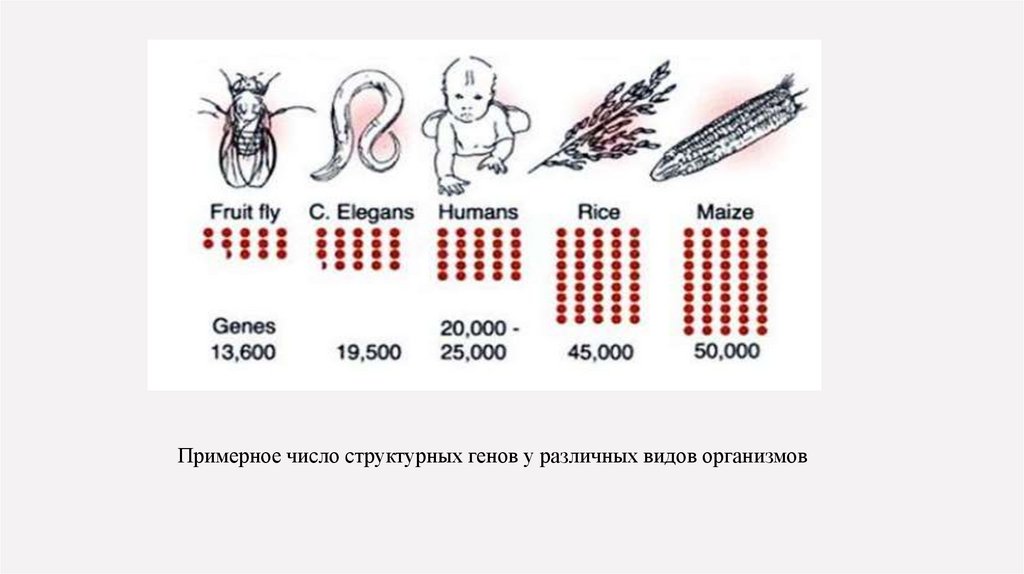
Примерное число структурных генов у различных видов организмов26.
У эукариот структурные гены, функция которых связана с регуляторными, могут бытьорганизованы как:
- независимые гены,
- повторяющиеся гены,
- кластеры генов.
Независимые гены, как правило, располагаются индивидуально, их транскрипция не связана с
транскрипцией других генов. Активность некоторых из них регулируется гормонами.
Повторяющиеся гены присутствуют в хромосоме в виде повторов (копий) одного гена - гены
гистонов, тРНК, рРНК. Причина повторяемости гистоновых генов определяется необходимостью
синтезировать большое количество гистонов, которые являются основными структурными белками
ядра (суммарная масса гистонов равна массе ДНК).
Кластер генов - это группа различных генов с родственными функциями, локализованных в
определённых участках хромосом. В состав кластера входят активно функционирующие гены и
псевдогены. Нуклеотидные последовательности псевдогенов сходны с последовательностями
функционально активных генов, но псевдогены не экспрессируются и не образуют белок. Часто
кластеры являются семейством генов, произошедшим от некоего гена-предка.
27.
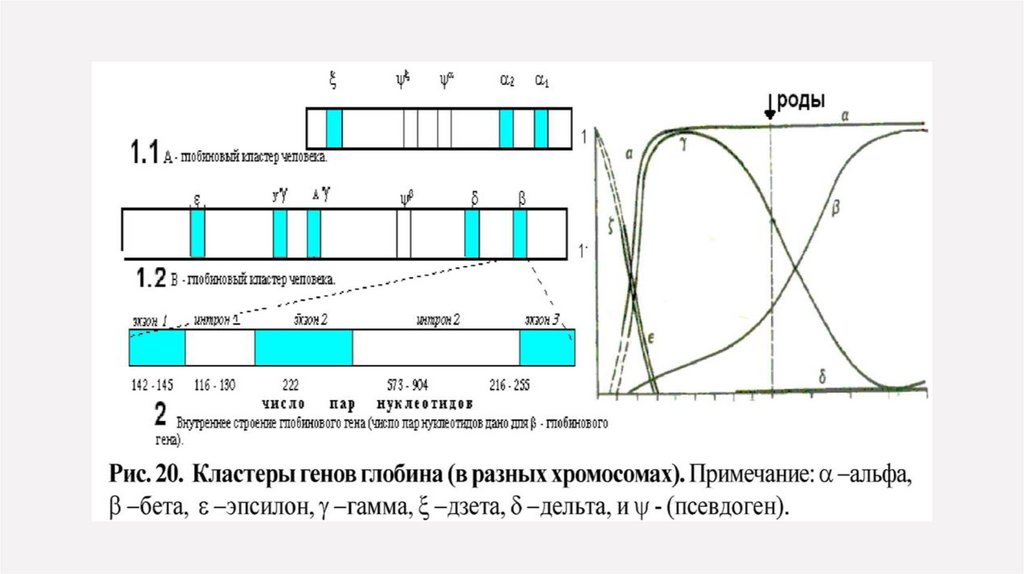
Классическим примером кластерных генов являются гены глобина в составе А и В кластеров.Гемоглобин представлен гемом и белком тетрамером-глобином.
Глобиновый тетрамер состоит из двух идентичных цепей и двух идентичных цепей.
Аминокислотная последовательность каждой глобиновой цепи и кодируется своим собственным
геном, входящим соответственно в состав А или В кластера.
У человека А кластер располагается в 16 хромосоме, а В кластер в 11 хромосоме.
Кластер В занимает участок ДНК размером 50 тысяч пар нуклеотидов и включает в себя пять
функционально активных генов и один псевдоген: ген (эпсилон); два гена (гамма); псевдоген (бэта);
ген (дельта) и ген (бэта).
Кластер А располагается более компактно и занимает участок ДНК размером более 28 тысяч пар
нуклеотидов и включает в себя активный ген (дзета), псевдоген (дзета), псевдоген (альфа) и гены
(альфа) два и (альфа) один, кодирующие идентичные белки. Гены глобина по внутреннему
строению являются мозаичными.
Повторяющиеся гены и кластеры глобиновых генов относятся к мультигенным семействам.
28.
29.
Основные принципы регуляции транскрипции генов у прокариотических иэукариотических организмов сходны: регуляция осуществляется через специфические
взаимодействия белков и нуклеиновых кислот друг с другом, а также между собой.
Однако, в связи с особенностями строения генов и организации генома в целом,
механизмы регуляции генной активности у эукариот намного разнообразнее и
сложнее по сравнению с прокариотами. Прежде всего, это связано с необходимостью
поддержания координированной экспрессии эукариотических генов в сложно
организованной генетической системе, которая имеет свои особенности:
эукариоты превосходят прокариот по размерам генома, клетки и организма в
целом;
организм эукариот может состоять из разных типов клеток, жизненный цикл
которых более сложен и продолжителен;
ядерная ДНК находится не в одной молекуле, а представлена несколькими
разными молекулами, формирующими хромосомы;
кроме ядерной ДНК у эукариот имеется ДНК митохондрий и хлоропластов,
которая также находится под контролем ядра;
ввиду наличия ядра процессы транскрипции и трансляции генов у эукариот
разделены в пространстве и во времени.
30.
В организме человека гистологическиразличают более 100 типов клеток,
формирующих различные органы и
ткани. Все этапы жизнедеятельности
клеток
высших
организмов
обеспечиваются функционированием
их генов. В связи с этим у
многоклеточных организмов в клетках
разных тканей и в разных типах
клеток одной ткани экспрессируются
различные гены, то есть для эукариот
характерно
явление
тканеспецифической
экспрессии,
обусловленное сложной системой
регуляторных
механизмов
генетической активности.
31.
Необходимо отметить, что в отличие от прокариот, у эукариотбольшинство генов репрессировано (выключено), поэтому
регуляция направлена в основном на их включение.
Например, в клетках эукариотического организма постоянно
транскрибируется не более 7–10% генов, большинство из которых
относится к группе «генов домашнего хозяйства» (housekeeping
genes), обеспечивающих процессы гликолиза, катаболизма белков,
биосинтеза протеинов общего назначения (рибосомальные белки,
гистоны, белки, формирующие веретено деления и т.п.),
транспортных и рибосомальных РНК.
Гены, относящиеся к другой группе – «гены роскоши»
(luxury genes) – экспрессируются лишь в определенных типах
клеток при определенных метаболических условиях либо на
отдельных этапах клеточной дифференцировки в зависимости от
ее тканевой принадлежности, периода жизненного цикла и стадии
индивидуального развития всего организма.
32.
В соответствии с современными научными представлениями ген эукариотическихклеток, кодирующий определённый белок (СТРУКТУРНЫЙ ГЕН), всегда состоит
из нескольких обязательных элементов. Как правило, в начале и в конце гена
располагаются специальные регуляторные участки; они определяют, когда, в какие
периоды онтогенеза и в каких тканях будет экспрессироваться этот ген.
Подобные регуляторные участки дополнительно могут находиться и вне гена,
располагаясь достаточно далеко, но тем не менее активно участвуя регуляции его
функционирования.
Кроме регуляторных зон существует структурная часть гена, которая, собственно, и
содержит информацию о первичной структуре соответствующего белка. У
большинства генов эукариот она существенно короче регуляторной зоны.
33.
Регуляторные области, как участок первого экзонов гена (промотор), так идистальные (энхансер и сайленсер) НЕ ТРАНСКРИБИРУЮТСЯ!
34.
Промотор — нуклеотидная последовательность ДНК длиной до 80—100 парнуклеотидов, которую узнает молекула фермента РНК-полимеразы и соединяется с
ней, чтобы начать транскрипцию данного гена. При этом промотор определяет,
какая из цепей двойной спирали ДНК присоединит к себе РНК-полимеразу и тем
самым будет служить матрицей для синтеза и-РНК.
Каждый структурный ген имеет свой промотор (в отличие от гена прокариот, где
один промотор может регулировать экспрессию нескольких генов, находящихся в
составе оперона).
35.
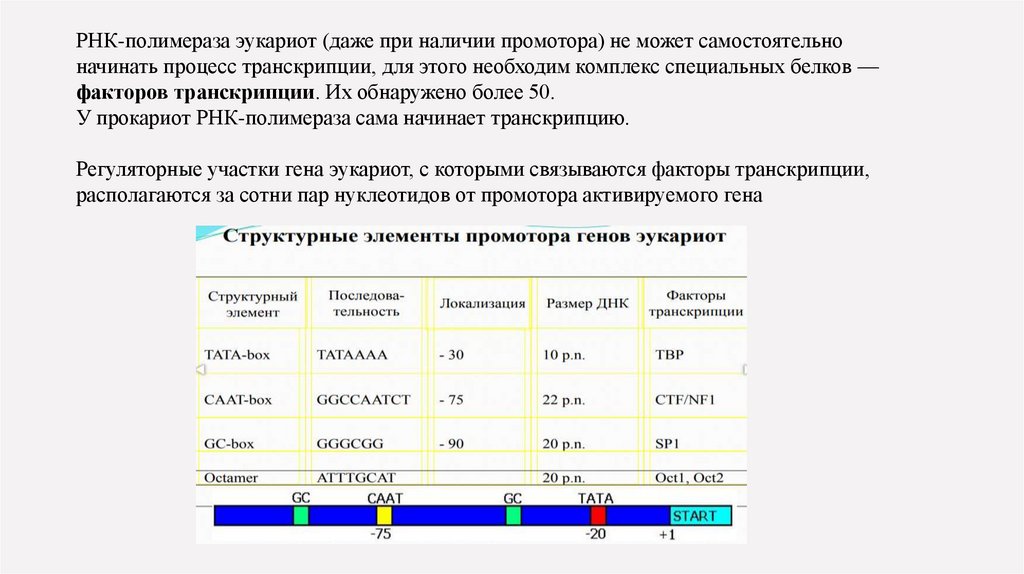
РНК-полимераза эукариот (даже при наличии промотора) не может самостоятельноначинать процесс транскрипции, для этого необходим комплекс специальных белков —
факторов транскрипции. Их обнаружено более 50.
У прокариот РНК-полимераза сама начинает транскрипцию.
Регуляторные участки гена эукариот, с которыми связываются факторы транскрипции,
располагаются за сотни пар нуклеотидов от промотора активируемого гена
36.
Для нормальной работы гена очень важны также участки, расположенные в концекодирующей последовательности и служащие сигналом остановки транскрипции терминаторы.
37.
У эукариот обнаружены специальные участки, которые могут бытьрасположены за несколько тысяч пар нуклеотидов от промотора, но
способные при связывании со специальными регуляторными белками
активировать или подавлять процесс транскрипции.
Открытые в 1976 г., эти участки получили название энхансеры (от
англ. enchance — усиливать). Энхансеры способны связываться с РНКполимеразой и факторами транскрипции, что приводит к изменению
конформации ДНК и началу процесса транскрипции.
Также в нескольких тысячах пар нуклеотидов от промотора
эукариотического гена расположены сайленсеры (от англ. silence —
заглушать) — это последовательности ДНК, и оказывающие дистанционное
влияние на его транскрипцию, снижая активность гена.
Каждый ген или группа генов отделены друг от друга специальными
участками ДНК, которые называют спейсерами.
38.
39.
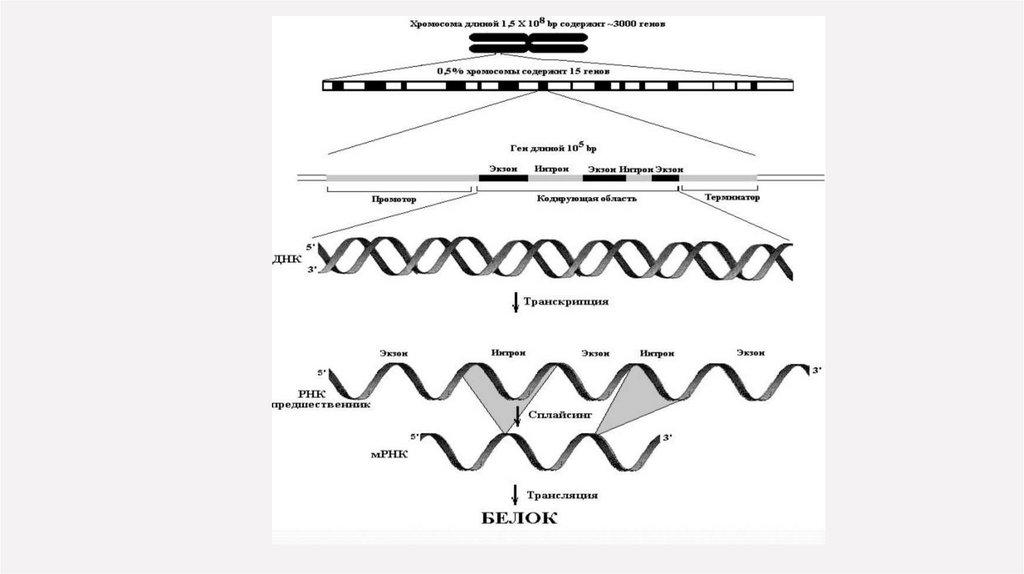
Число интронов может сильно варьировать: от 2 (в генах гемоглобина) до нескольких десятков(в гене коллагена у человека).
Размер интронов колеблется от 100 п.н. до 1000 п.н. или 1 kb и более.
Таким образом из всего мозаичного гена (длиной обычно от 3 до 10 kb) лишь 10-20 % ДНК несет
информацию о составе кодируемого им продукта.
40.
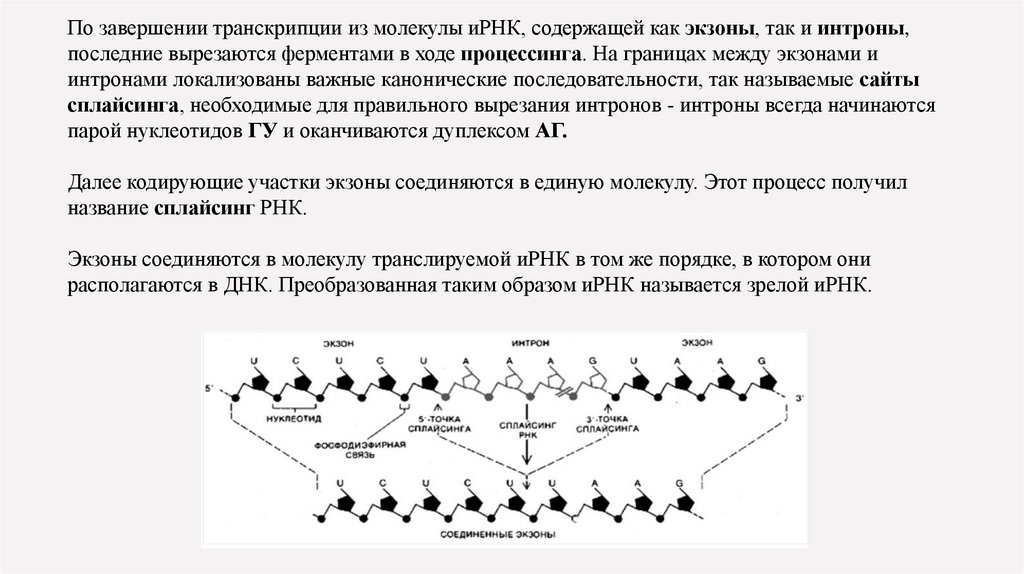
По завершении транскрипции из молекулы иРНК, содержащей как экзоны, так и интроны,последние вырезаются ферментами в ходе процессинга. На границах между экзонами и
интронами локализованы важные канонические последовательности, так называемые сайты
сплайсинга, необходимые для правильного вырезания интронов - интроны всегда начинаются
парой нуклеотидов ГУ и оканчиваются дуплексом АГ.
Далее кодирующие участки экзоны соединяются в единую молекулу. Этот процесс получил
название сплайсинг РНК.
Экзоны соединяются в молекулу транслируемой иРНК в том же порядке, в котором они
располагаются в ДНК. Преобразованная таким образом иРНК называется зрелой иРНК.
41.
На сегодня известно несколько механизмов сплайсинга1. Один из них описан в 1980 г. Абельсоном, обнаружившим ферменты, вырезающие интроны из
молекул – предшественников транспортной РНКу дрожжей. Точный разрез по границе экзон-интрон
обеспечивается тем, что молекула интрона обладает способностью складываться таким образом, что
оба конца его оказываются рядом. В этом процессе принимают участие малые РНК,
комплиментарные участкам интронов, прилегающим к терминирующим дуплексам, получившие
название – «лидерные последовательности». Терминирующие дуплексы являются сигналом,
позволяющим молекулярному комплексу, обеспечивающему сплайсинг опознать место
прикрепления лРНК, которая, связываясь с комплиментарными участками интрона, способствует
образованию им петли.
2. Второй тип сплайсинга был обнаружен Чех в 1982 при изучении биосинтеза рибосомальной РНК.
Оказалось, что в данном случае сама нуклеотидная последовательность интрона обладает
псевдоферментной активностью, т.е. интрон вырезает себя сам (аутокаталитический сплайсинг).
42.
3. Но наиболее интересным оказался феномен, выявленный Кошко и Ламуром (1978). Длямитохондриального гена, кодирующего синтез цитохрома b, было показано, что интрон может
вырезаться ферментом, который кодируется нуклеотидными последовательностями самого
интрона. Этот фермент получил название мРНК-матуразы (от франц. maturation - созревание).
Было показано, что матуразы играют регуляторную в координированной экспрессии генов
(Слонимски, Даншен; 1981). В ряде случаев они являются белками, ускоряющими
аутокаталитическое вырезание интронов.
4. Однако наиболее значимым результатом этих исследований, явилось открытие роли
интронов в хранении генетической информации, что позволило Дашену и Слонимски (1987)
выдвинуть принцип – «Один ген может кодировать несколько белков».
Существует, по крайней мере, два механизма реализации схемы "один ген – несколько белков",
которые несколько видоизменяют классическую концепцию гена, но не опровергают ее: вопервых, кодирование белков интронами; во-вторых, синтез разных белков с одной матрицы в
результате модификации сплайсинга (альтернативный сплайсинг).
43.
44.
Нетранслируемые области (НТО (НТП), англ. untranslated regions, UTR) — особые участки ДНК,не выступающие в качестве матрицы для синтеза белка и прилегающие с обеих сторон к
транслируемой области (то есть той, на матрице которой синтезируется белок). Таких области две:
5'-нетранслируемая область (5'-НТО) и 3'-нетранслируемая область (3'-НТО), располагающиеся на
5'- и 3'-конце ДНК соответственно. Такое же название имеют участки мРНК, соответствующие 5'НТО и 3'-НТО транскрипта.
Нетранслируемые области вовлечены в регуляцию локализации, трансляции и деградации того
транскрипта, в составе которого они находятся. Для них характерно наличие шпилек, внутренних
инициаторных кодонов и открытых рамок считывания, сайтов связывания рибосомы, различных
цис-регуляторных элементов, связывающихся с РНК-связывающими белками.
Схематическое изображение строения типичной зрелой человеческой мРНК. Масштаб соблюдён
только приблизительно: кэп представляет собой лишь один модифицированный нуклеотид, в то
время как длина 5'-UTR и 3'-UTR составляет 170 и 700 нуклеотидов соответственно.
45.
Для нетранслируемых областей характерны следующие элементы:Шпилька (англ. stem-loop, hairpin) — в молекулярной биологии элемент
вторичной структуры РНК, а также одноцепочечной ДНК. Шпилька
образуется в том случае, когда две последовательности одной и той же
цепи комплементарны друг другу и соединяются друг с другом,
перегибаясь одна к другой и образуя на конце неспаренный участок —
петлю. Такие комплементарные последовательности нередко
представляют собой палиндромные последовательности.
У некоторых видов РНК шпильки имеют важное функциональное
значение. Шпильки часто встречаются в 5'-нетранслируемой области
прокариот. Эти структуры нередко связываются с белками и отвечают за
аттенюацию (это регулируемая терминация, которая происходит лишь в
том случае, если строящаяся мРНК приобретает определенную
вторичную структуру), тем самым участвуя в регуляции транскрипции.
46.
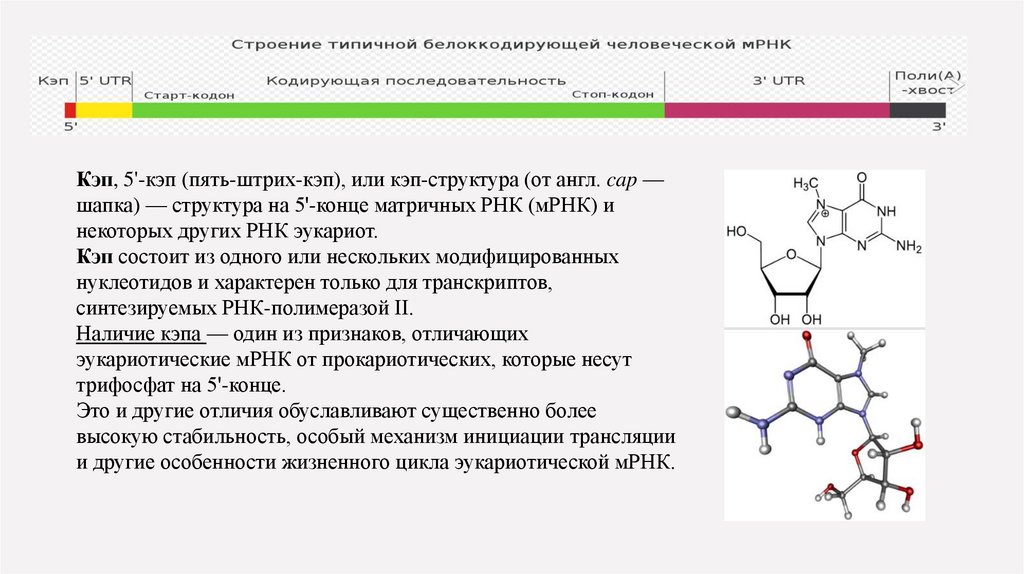
Кэп, 5'-кэп (пять-штрих-кэп), или кэп-структура (от англ. cap —шапка) — структура на 5'-конце матричных РНК (мРНК) и
некоторых других РНК эукариот.
Кэп состоит из одного или нескольких модифицированных
нуклеотидов и характерен только для транскриптов,
синтезируемых РНК-полимеразой II.
Наличие кэпа — один из признаков, отличающих
эукариотические мРНК от прокариотических, которые несут
трифосфат на 5'-конце.
Это и другие отличия обуславливают существенно более
высокую стабильность, особый механизм инициации трансляции
и другие особенности жизненного цикла эукариотической мРНК.