


























 Биология
БиологияПохожие презентации:










Поверхностный аппарат клетки
1. Практическое занятие 3 Поверхностный аппарат клетки
ЦитологияПрактическое занятие 3
Поверхностный аппарат
клетки
Кафедра биологии
Военно-медицинская академия им. С.М.Кирова
2. Учебные вопросы
1.Общий план строения плазмалеммы. Свойства и функцииплазмалеммы
2. Белки плазмалеммы: интегральные, полуинтегральные,
периферические
3. Липиды плазмалеммы: фосфолипиды, гликолипиды, липоиды
4. Типы транспорта веществ через плазмалемму
5. Надмембранный комплекс разных клеток
3.
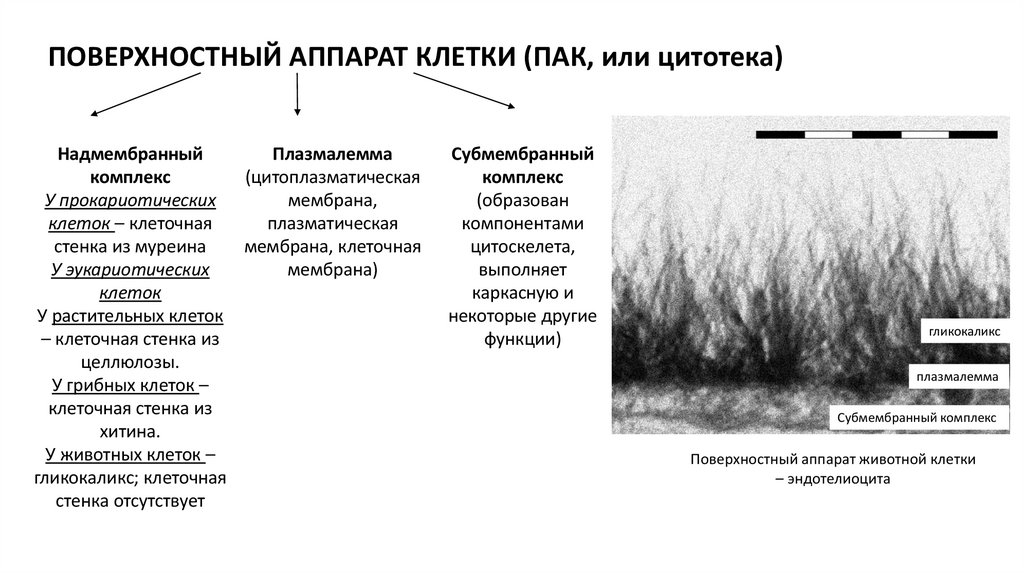
ПОВЕРХНОСТНЫЙ АППАРАТ КЛЕТКИ (ПАК, или цитотека)Надмембранный
Плазмалемма
комплекс
(цитоплазматическая
У прокариотических
мембрана,
клеток – клеточная
плазматическая
стенка из муреина
мембрана, клеточная
У эукариотических
мембрана)
клеток
У растительных клеток
– клеточная стенка из
целлюлозы.
У грибных клеток –
клеточная стенка из
хитина.
У животных клеток –
гликокаликс; клеточная
стенка отсутствует
Субмембранный
комплекс
(образован
компонентами
цитоскелета,
выполняет
каркасную и
некоторые другие
функции)
гликокаликс
плазмалемма
Субмембранный комплекс
Поверхностный аппарат животной клетки
– эндотелиоцита
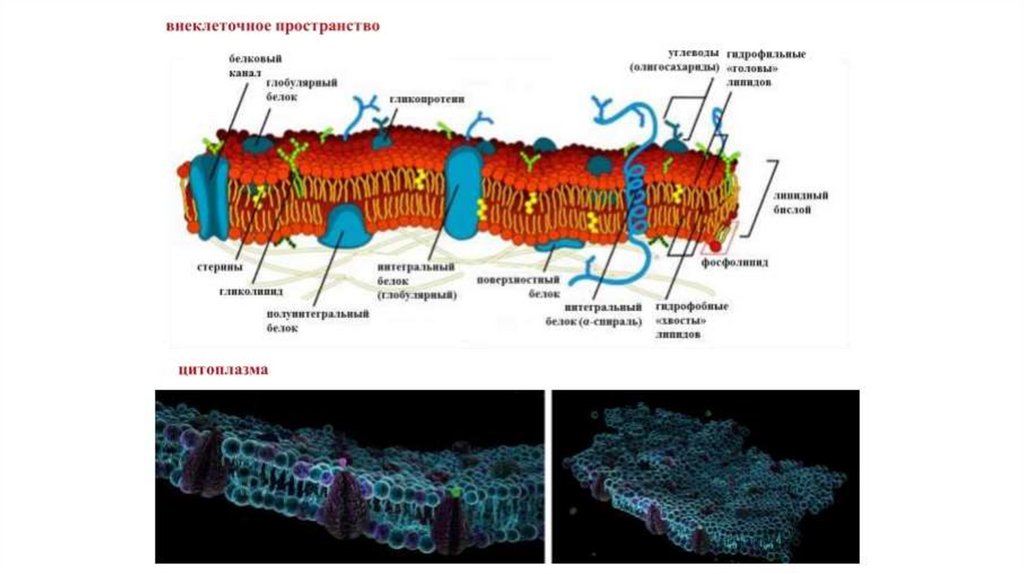
4. Строение поверхностного аппарата животной клетки
91 — липидный бислой
плазмалеммы;
2,8 — трансмембранные
белки плазмалеммы;
3 — гидрофобные
«хвосты»
фосфолипидов;
4 — интегральные белки
(гликопротеиды)
плазмалеммы;
5,7 — структурные
элементы (лучше –
компоненты)
субмембранного
комплекса;
6 — гидрофильные
«головки» фосфолипидов
9 – углеводные радикалы
гликолипидов и
гликопротеидов
5. Вопрос 1. Общий план строения плазмалеммы. Свойства и функции плазмалеммы
ПЛАЗМАЛЕММА –сферически замкнутая
биологическая мембрана,
располагающаяся вокруг
содержимого клетки и
выполняющая
рецепторно-барьернотранспортную функцию
6.
7.
МОДЕЛИ СТРОЕНИЯ ПЛАЗМАЛЕММЫ1) Сэндвич-модель (1935,Н. Даусон и Р. Даниэлли): плазмалемма
состоит из липидного бислоя, окруженного двумя слоями белков.
Модель была признана несостоятельной в связи с тем, что она не
объясняла:
• Транспортную функцию мембраны (транспорту гидрофильных веществ препятствует сплошной липидный бислой,
транспорту гидрофобных (липофильных) веществ – периферические слои белков).
• Рецепторную функцию мембраны (ее могут выполнять интегральные белки, которые в данной модели отсутствуют).
• Смену мембранного потенциала покоя мембранным потенциалом действия (отсутствуют интегральные белки – ионные
каналы)
• Способность мембраны к самосборке
8.
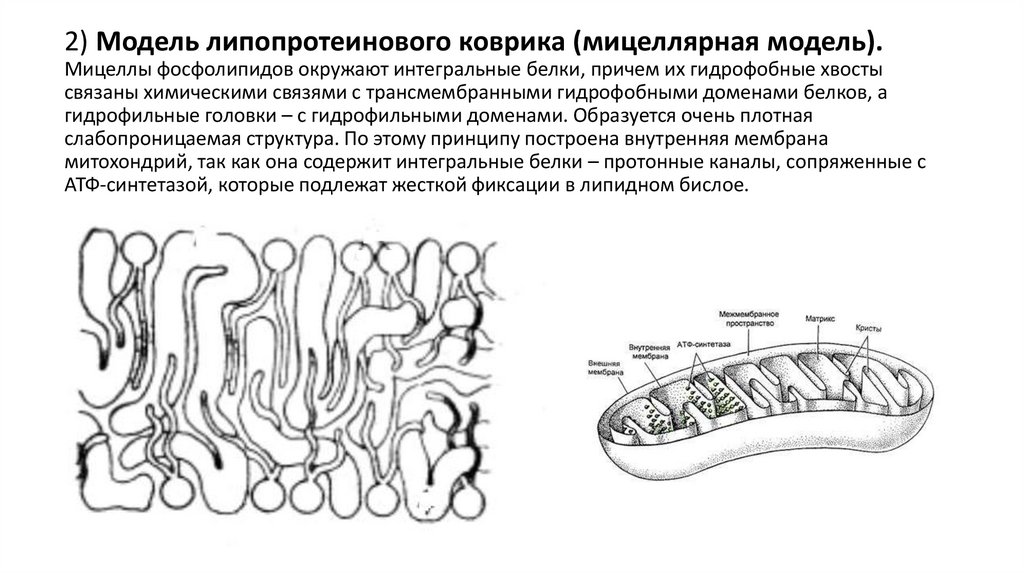
2) Модель липопротеинового коврика (мицеллярная модель).Мицеллы фосфолипидов окружают интегральные белки, причем их гидрофобные хвосты
связаны химическими связями с трансмембранными гидрофобными доменами белков, а
гидрофильные головки – с гидрофильными доменами. Образуется очень плотная
слабопроницаемая структура. По этому принципу построена внутренняя мембрана
митохондрий, так как она содержит интегральные белки – протонные каналы, сопряженные с
АТФ-синтетазой, которые подлежат жесткой фиксации в липидном бислое.
9.
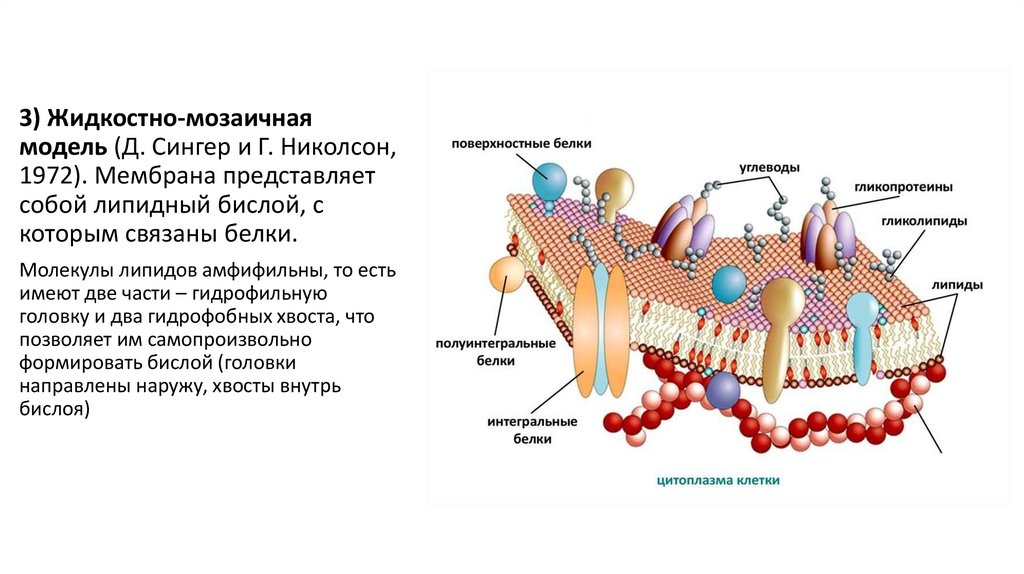
3) Жидкостно-мозаичнаямодель (Д. Сингер и Г. Николсон,
1972). Мембрана представляет
собой липидный бислой, с
которым связаны белки.
Молекулы липидов амфифильны, то есть
имеют две части – гидрофильную
головку и два гидрофобных хвоста, что
позволяет им самопроизвольно
формировать бислой (головки
направлены наружу, хвосты внутрь
бислоя)
10.
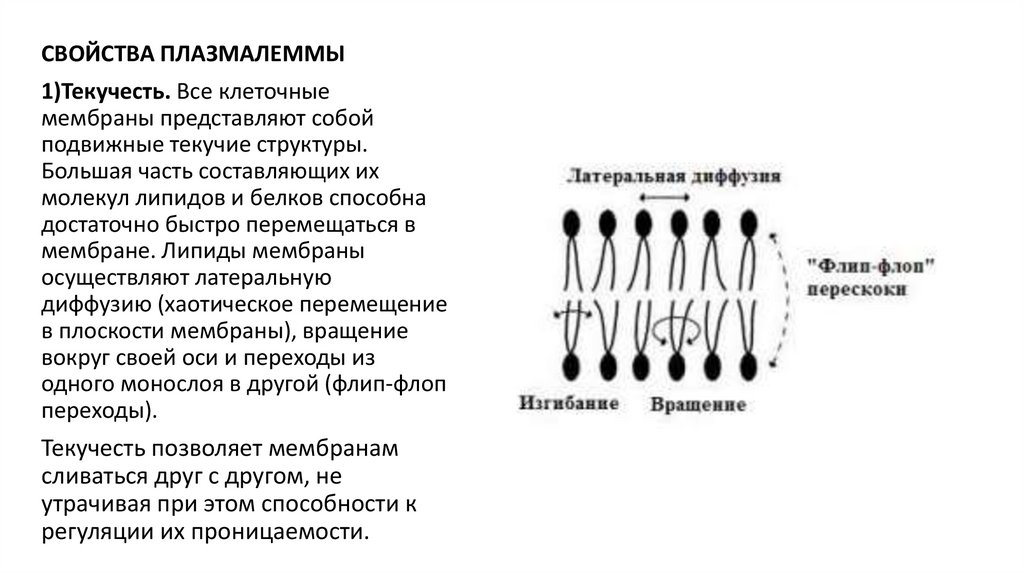
СВОЙСТВА ПЛАЗМАЛЕММЫ1)Текучесть. Все клеточные
мембраны представляют собой
подвижные текучие структуры.
Большая часть составляющих их
молекул липидов и белков способна
достаточно быстро перемещаться в
мембране. Липиды мембраны
осуществляют латеральную
диффузию (хаотическое перемещение
в плоскости мембраны), вращение
вокруг своей оси и переходы из
одного монослоя в другой (флип-флоп
переходы).
Текучесть позволяет мембранам
сливаться друг с другом, не
утрачивая при этом способности к
регуляции их проницаемости.
11.
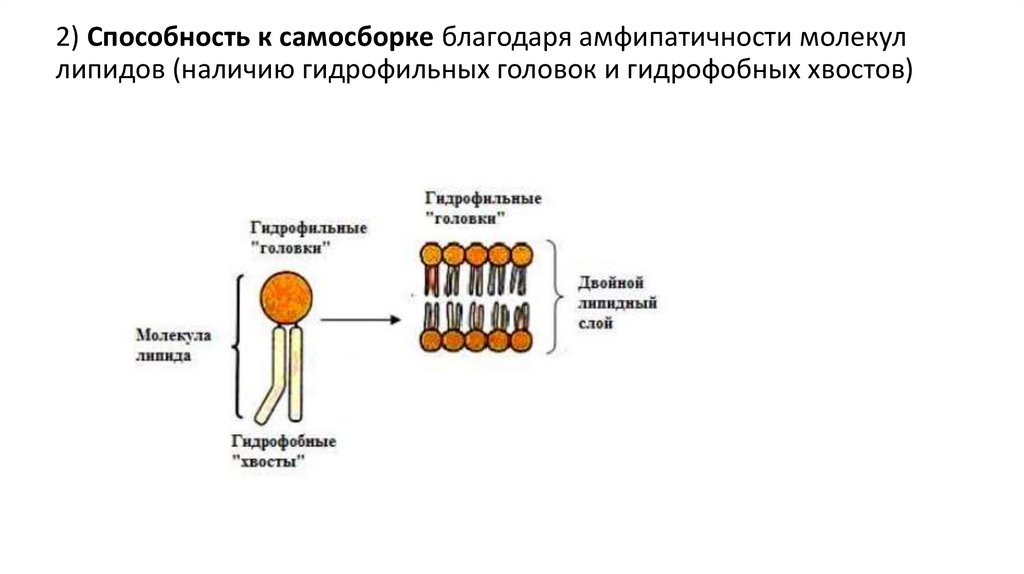
2) Способность к самосборке благодаря амфипатичности молекуллипидов (наличию гидрофильных головок и гидрофобных хвостов)
12.
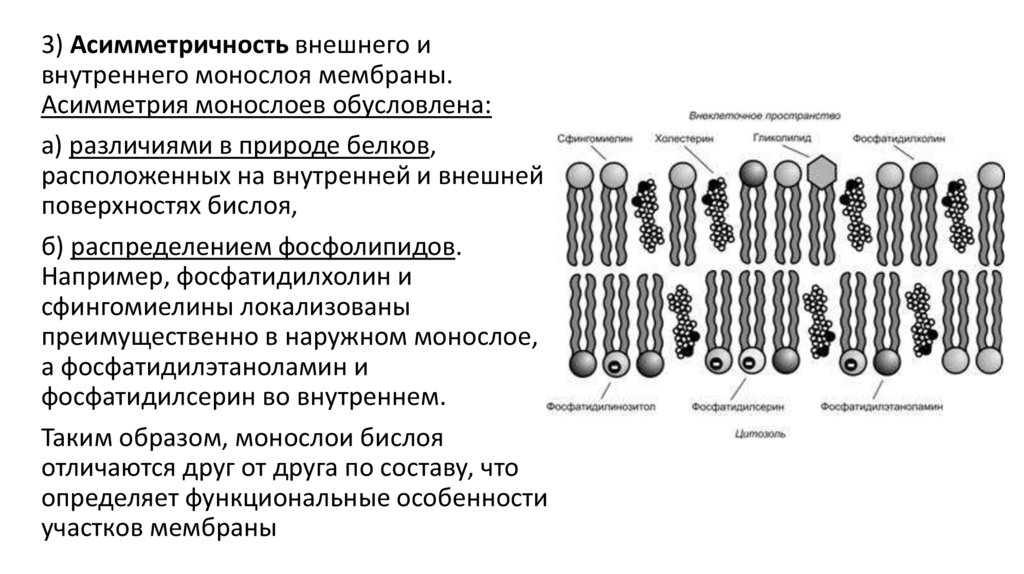
3) Асимметричность внешнего ивнутреннего монослоя мембраны.
Асимметрия монослоев обусловлена:
а) различиями в природе белков,
расположенных на внутренней и внешней
поверхностях бислоя,
б) распределением фосфолипидов.
Например, фосфатидилхолин и
сфингомиелины локализованы
преимущественно в наружном монослое,
а фосфатидилэтаноламин и
фосфатидилсерин во внутреннем.
Таким образом, монослои бислоя
отличаются друг от друга по составу, что
определяет функциональные особенности
участков мембраны
13.
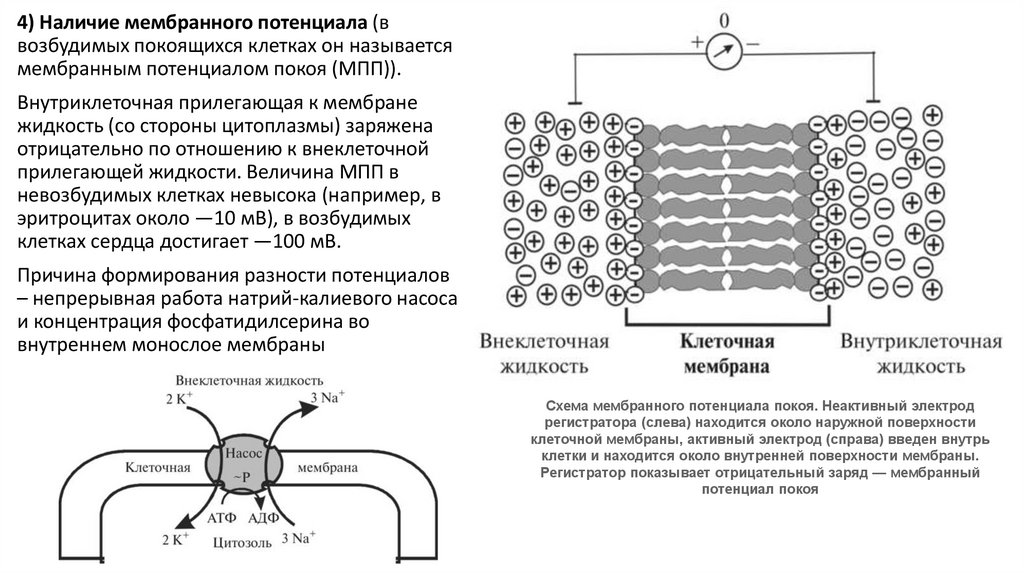
4) Наличие мембранного потенциала (ввозбудимых покоящихся клетках он называется
мембранным потенциалом покоя (МПП)).
Внутриклеточная прилегающая к мембране
жидкость (со стороны цитоплазмы) заряжена
отрицательно по отношению к внеклеточной
прилегающей жидкости. Величина МПП в
невозбудимых клетках невысока (например, в
эритроцитах около —10 мВ), в возбудимых
клетках сердца достигает —100 мВ.
Причина формирования разности потенциалов
– непрерывная работа натрий-калиевого насоса
и концентрация фосфатидилсерина во
внутреннем монослое мембраны
Схема мембранного потенциала покоя. Неактивный электрод
регистратора (слева) находится около наружной поверхности
клеточной мембраны, активный электрод (справа) введен внутрь
клетки и находится около внутренней поверхности мембраны.
Регистратор показывает отрицательный заряд — мембранный
потенциал покоя
14.
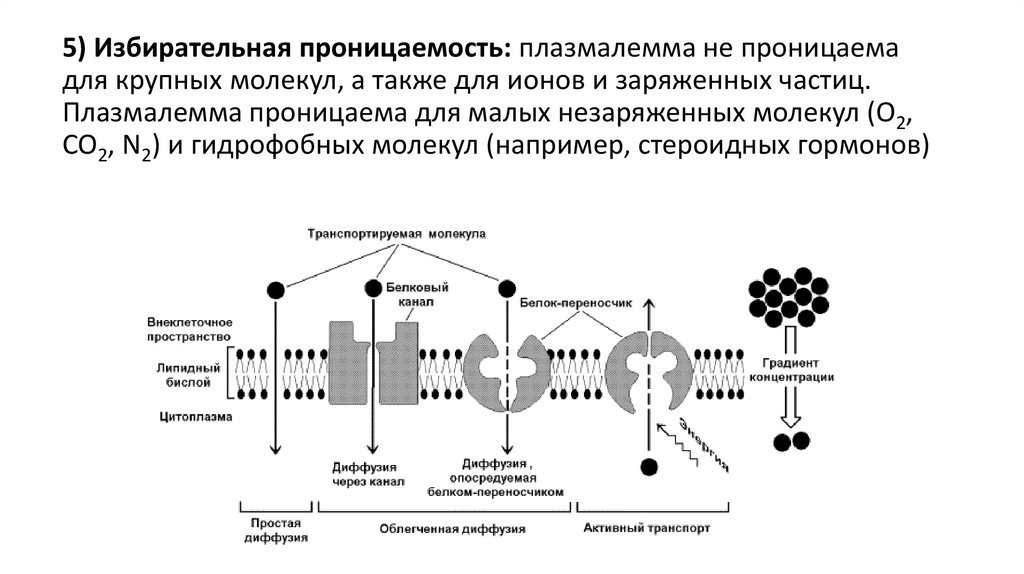
5) Избирательная проницаемость: плазмалемма не проницаемадля крупных молекул, а также для ионов и заряженных частиц.
Плазмалемма проницаема для малых незаряженных молекул (O2,
CO2, N2) и гидрофобных молекул (например, стероидных гормонов)
15.
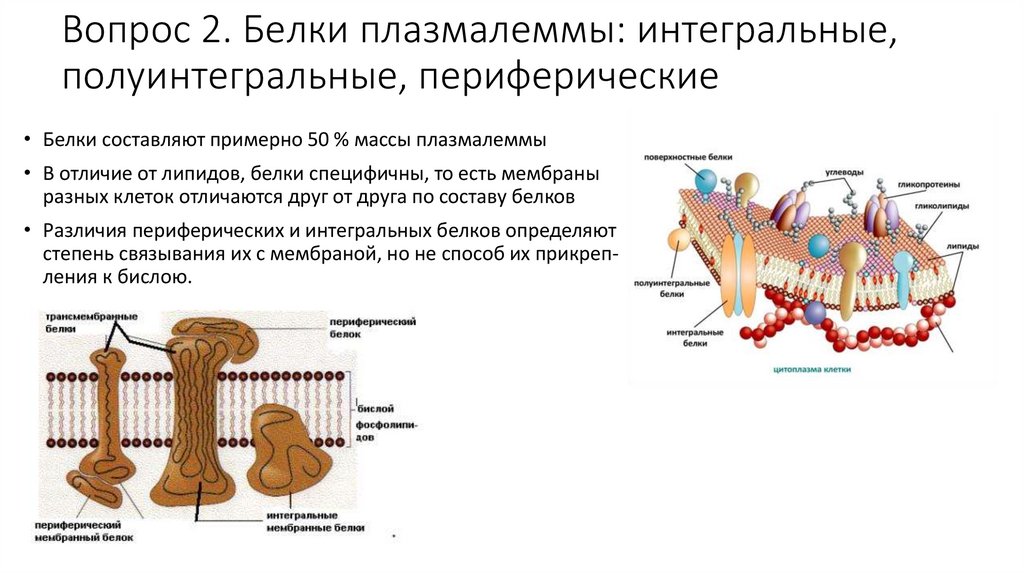
Вопрос 2. Белки плазмалеммы: интегральные,полуинтегральные, периферические
• Белки составляют примерно 50 % массы плазмалеммы
• В отличие от липидов, белки специфичны, то есть мембраны
разных клеток отличаются друг от друга по составу белков
• Различия периферических и интегральных белков определяют
степень связывания их с мембраной, но не способ их прикрепления к бислою.
16.
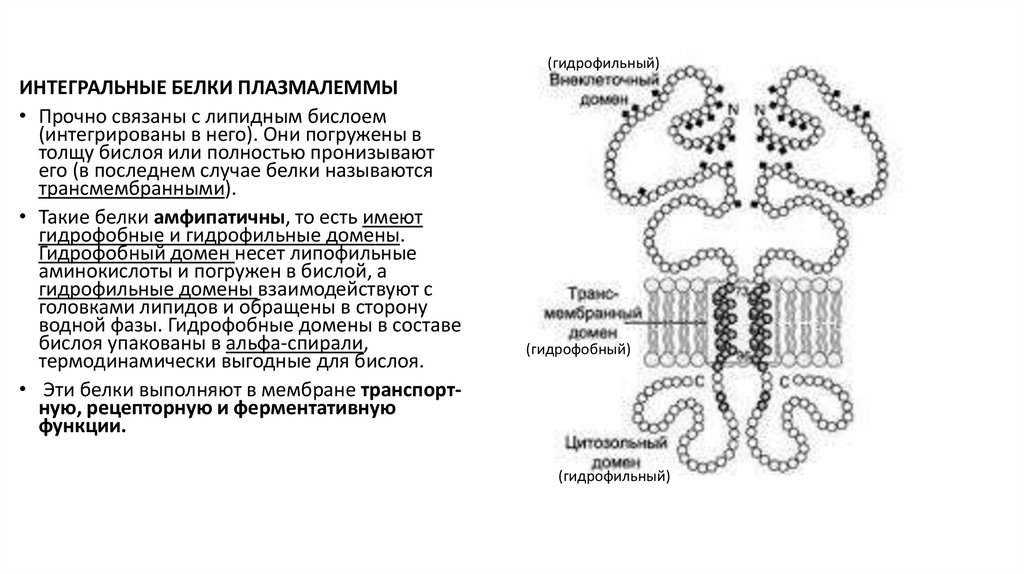
(гидрофильный)ИНТЕГРАЛЬНЫЕ БЕЛКИ ПЛАЗМАЛЕММЫ
• Прочно связаны с липидным бислоем
(интегрированы в него). Они погружены в
толщу бислоя или полностью пронизывают
его (в последнем случае белки называются
трансмембранными).
• Такие белки амфипатичны, то есть имеют
гидрофобные и гидрофильные домены.
Гидрофобный домен несет липофильные
аминокислоты и погружен в бислой, а
гидрофильные домены взаимодействуют с
головками липидов и обращены в сторону
водной фазы. Гидрофобные домены в составе
бислоя упакованы в альфа-спирали,
термодинамически выгодные для бислоя.
• Эти белки выполняют в мембране транспортную, рецепторную и ферментативную
функции.
(гидрофобный)
(гидрофильный)
17.
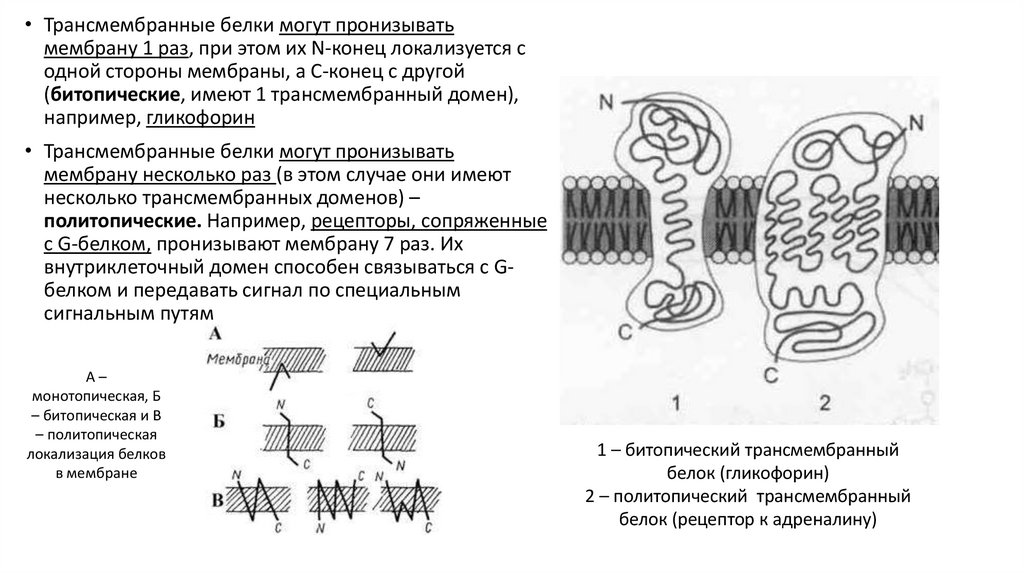
• Трансмембранные белки могут пронизыватьмембрану 1 раз, при этом их N-конец локализуется с
одной стороны мембраны, а С-конец с другой
(битопические, имеют 1 трансмембранный домен),
например, гликофорин
• Трансмембранные белки могут пронизывать
мембрану несколько раз (в этом случае они имеют
несколько трансмембранных доменов) –
политопические. Например, рецепторы, сопряженные
с G-белком, пронизывают мембрану 7 раз. Их
внутриклеточный домен способен связываться с Gбелком и передавать сигнал по специальным
сигнальным путям
А–
монотопическая, Б
– битопическая и В
– политопическая
локализация белков
в мембране
1 – битопический трансмембранный
белок (гликофорин)
2 – политопический трансмембранный
белок (рецептор к адреналину)
18.
• ПОЛУИНТЕГРАЛЬНЫЕ БЕЛКИ• Погружены в бислой частично, располагаются либо во внешнем,
либо во внутреннем монослое.
• Гидрофобные домены располагаются только на одном полюсе
белка, а на противоположном –гидрофильные
• Могут пронизывать монослой несколько раз, например, фермент
простагландинсинтетаза
19.
ПЕРИФЕРИЧЕСКИЕ БЕЛКИ• Не имеют гидрофобных доменов,
располагаются на внешней или
внутренней поверхности
мембраны
• Легко выделяются из состава
мембраны даже при изменении
солевого баланса
• Могут быть связаны ионными
взаимодействиями либо с
головками липидов, либо с
интегральными белками
1
2
Типы поверхностных белков:
1- белки, связанные с интегральными белками
(сукцинатдегидрогеназа)
2 - белки, присоединенные к полярным «головкам»
липидного слоя (протеинкиназа С)
20.
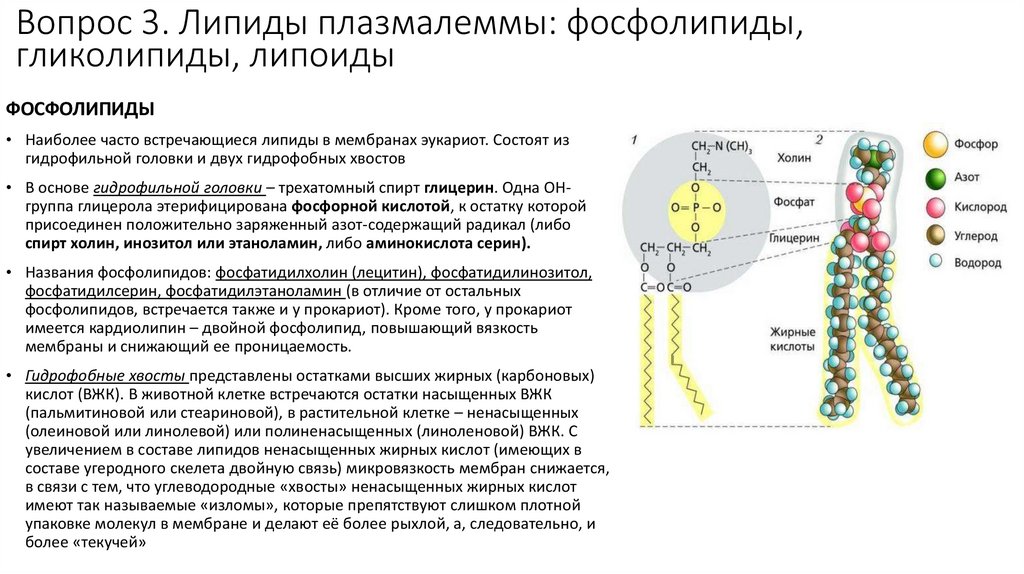
Вопрос 3. Липиды плазмалеммы: фосфолипиды,гликолипиды, липоиды
ФОСФОЛИПИДЫ
• Наиболее часто встречающиеся липиды в мембранах эукариот. Состоят из
гидрофильной головки и двух гидрофобных хвостов
• В основе гидрофильной головки – трехатомный спирт глицерин. Одна ОНгруппа глицерола этерифицирована фосфорной кислотой, к остатку которой
присоединен положительно заряженный азот-содержащий радикал (либо
спирт холин, инозитол или этаноламин, либо аминокислота серин).
• Названия фосфолипидов: фосфатидилхолин (лецитин), фосфатидилинозитол,
фосфатидилсерин, фосфатидилэтаноламин (в отличие от остальных
фосфолипидов, встречается также и у прокариот). Кроме того, у прокариот
имеется кардиолипин – двойной фосфолипид, повышающий вязкость
мембраны и снижающий ее проницаемость.
• Гидрофобные хвосты представлены остатками высших жирных (карбоновых)
кислот (ВЖК). В животной клетке встречаются остатки насыщенных ВЖК
(пальмитиновой или стеариновой), в растительной клетке – ненасыщенных
(олеиновой или линолевой) или полиненасыщенных (линоленовой) ВЖК. С
увеличением в составе липидов ненасыщенных жирных кислот (имеющих в
составе угеродного скелета двойную связь) микровязкость мембран снижается,
в связи с тем, что углеводородные «хвосты» ненасыщенных жирных кислот
имеют так называемые «изломы», которые препятствуют слишком плотной
упаковке молекул в мембране и делают её более рыхлой, а, следовательно, и
более «текучей»
21.
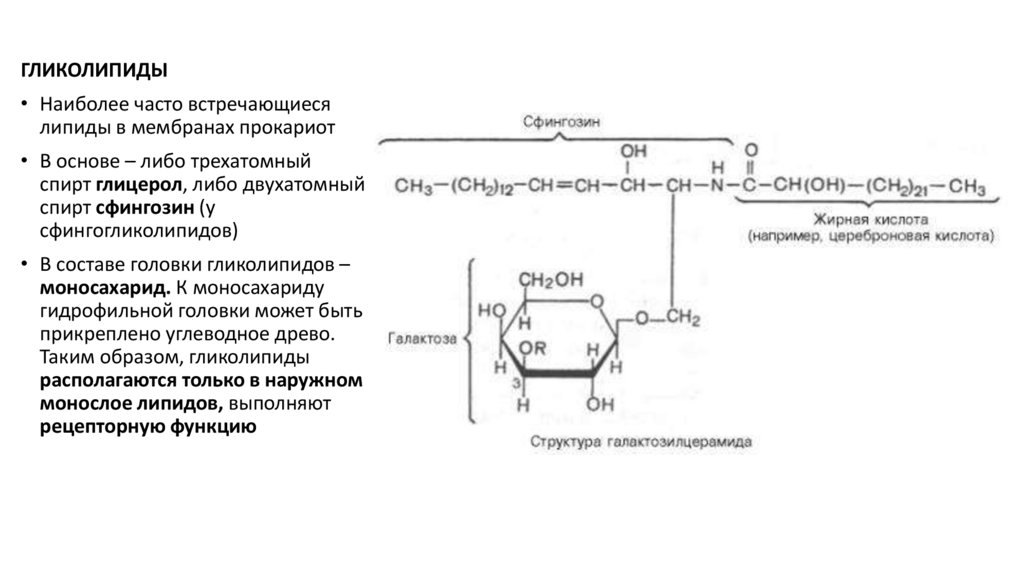
ГЛИКОЛИПИДЫ• Наиболее часто встречающиеся
липиды в мембранах прокариот
• В основе – либо трехатомный
спирт глицерол, либо двухатомный
спирт сфингозин (у
сфингогликолипидов)
• В составе головки гликолипидов –
моносахарид. К моносахариду
гидрофильной головки может быть
прикреплено углеводное древо.
Таким образом, гликолипиды
располагаются только в наружном
монослое липидов, выполняют
рецепторную функцию
22.
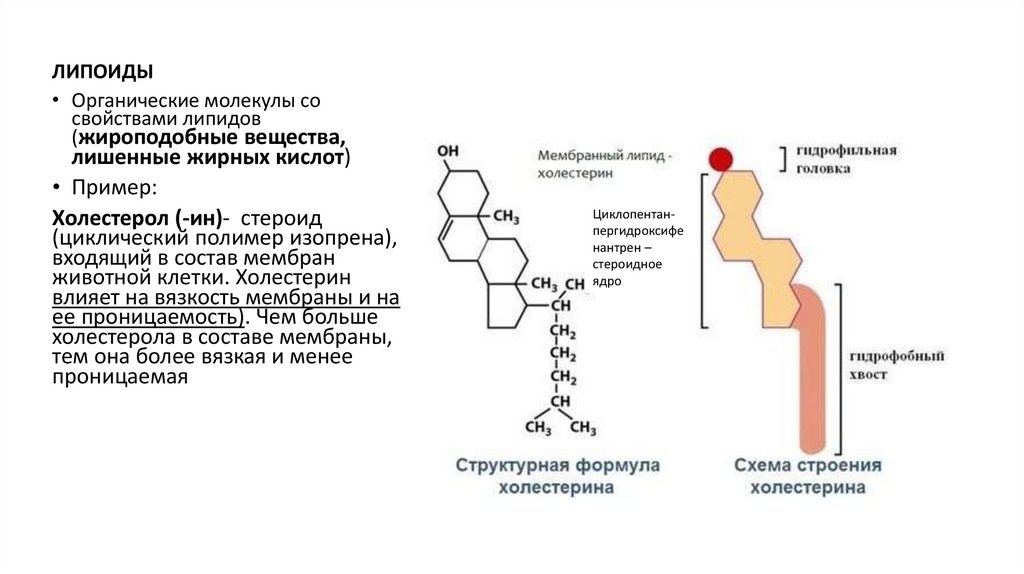
ЛИПОИДЫ• Органические молекулы со
свойствами липидов
(жироподобные вещества,
лишенные жирных кислот)
• Пример:
Холестерол (-ин)- стероид
(циклический полимер изопрена),
входящий в состав мембран
животной клетки. Холестерин
влияет на вязкость мембраны и на
ее проницаемость). Чем больше
холестерола в составе мембраны,
тем она более вязкая и менее
проницаемая
Циклопентанпергидроксифе
нантрен –
стероидное
ядро
23. Вопрос 4. Типы транспорта веществ через плазмалемму
ТРАНСПОРТ• А) КЛАССИФИКАЦИЯ ТИПОВ ТРАНСПОРТА ПО
ГРАДИЕНТУ КОНЦЕНТРАЦИИ, ЭНЕРГОЗАТРАТАМ
И НАЛИЧИЮ СПЕЦИФИЧЕСКИХ ПЕРЕНОСЧИКОВ
Активный
Пассивный
Простая диффузия Облегченная диффузия
24.
1) ПРОСТАЯ ДИФФУЗИЯ• Осуществляется по градиенту концентрации, без
энергозатрат и не требует наличия специальных
переносчиков. К ней относится осмос и транспорт малых
неполярных молекул через мембранный бислой
• Осмос – простая диффузия воды через полупроницаемую
мембрану. В изотоническом растворе содержание солей
соответствует содержимому солей внутри клетки,
следовательно, движение воды не происходит. В
гипертоническом растворе содержание солей выше, чем
внутри клетки, поэтому вода выходит из клетки, что приводит
ее сжатию (плазмолиз). В гипотоническом растворе
содержание солей ниже, чем внутри клетки, в связи с чем
вода проникает в клетку, что приводит к ее расширению
(деплазмолиз).
• Липофильные молекулы, способные осуществлять простую
диффузию: витамин А (ретинол), витамин D (кальциферол),
витамин Е (токоферол), витамин К (филлохинон)
25.
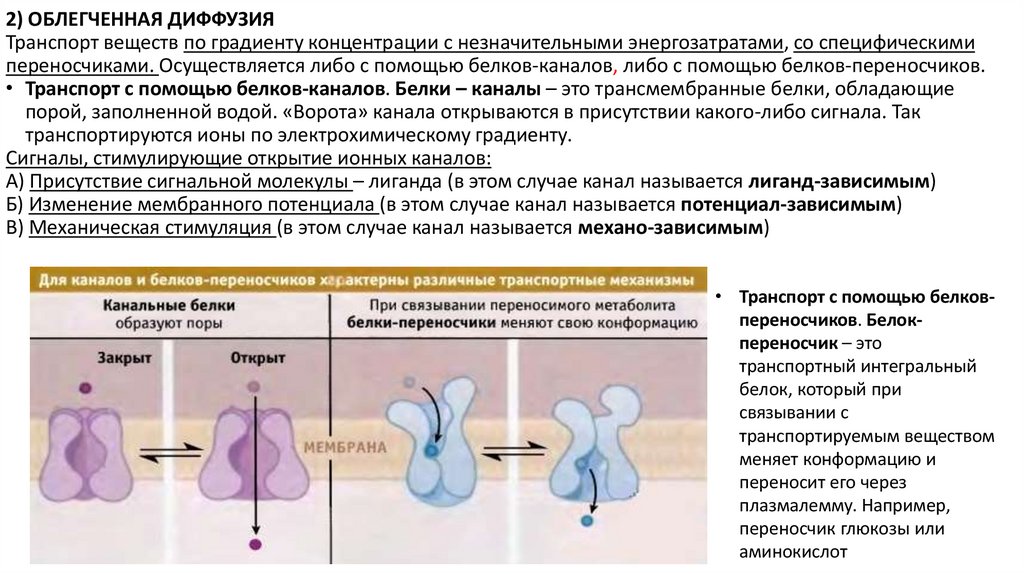
2) ОБЛЕГЧЕННАЯ ДИФФУЗИЯТранспорт веществ по градиенту концентрации с незначительными энергозатратами, со специфическими
переносчиками. Осуществляется либо с помощью белков-каналов, либо с помощью белков-переносчиков.
• Транспорт с помощью белков-каналов. Белки – каналы – это трансмембранные белки, обладающие
порой, заполненной водой. «Ворота» канала открываются в присутствии какого-либо сигнала. Так
транспортируются ионы по электрохимическому градиенту.
Сигналы, стимулирующие открытие ионных каналов:
А) Присутствие сигнальной молекулы – лиганда (в этом случае канал называется лиганд-зависимым)
Б) Изменение мембранного потенциала (в этом случае канал называется потенциал-зависимым)
В) Механическая стимуляция (в этом случае канал называется механо-зависимым)
• Транспорт с помощью белковпереносчиков. Белокпереносчик – это
транспортный интегральный
белок, который при
связывании с
транспортируемым веществом
меняет конформацию и
переносит его через
плазмалемму. Например,
переносчик глюкозы или
аминокислот
26.
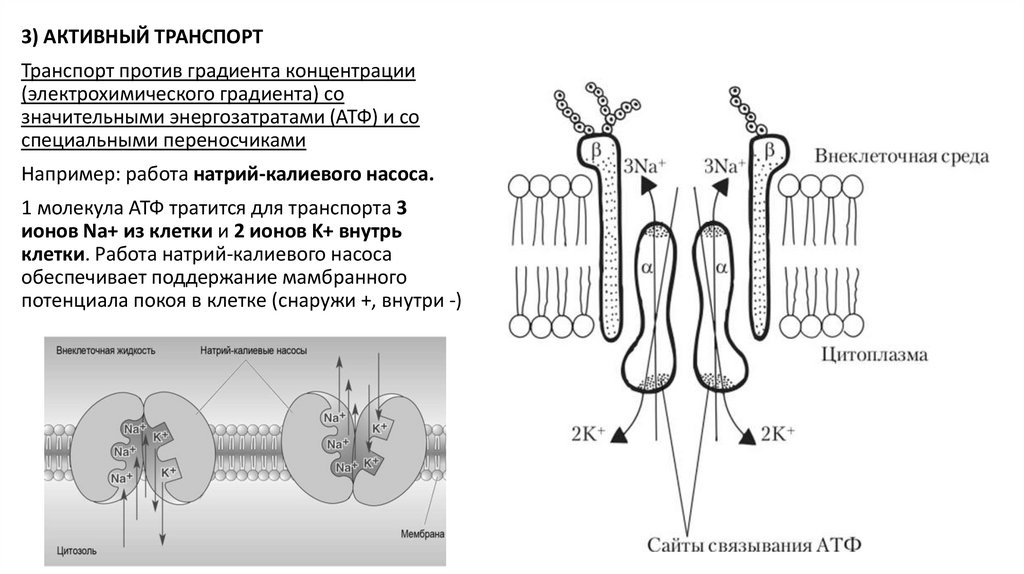
3) АКТИВНЫЙ ТРАНСПОРТТранспорт против градиента концентрации
(электрохимического градиента) со
значительными энергозатратами (АТФ) и со
специальными переносчиками
Например: работа натрий-калиевого насоса.
1 молекула АТФ тратится для транспорта 3
ионов Na+ из клетки и 2 ионов K+ внутрь
клетки. Работа натрий-калиевого насоса
обеспечивает поддержание мамбранного
потенциала покоя в клетке (снаружи +, внутри -)
27.
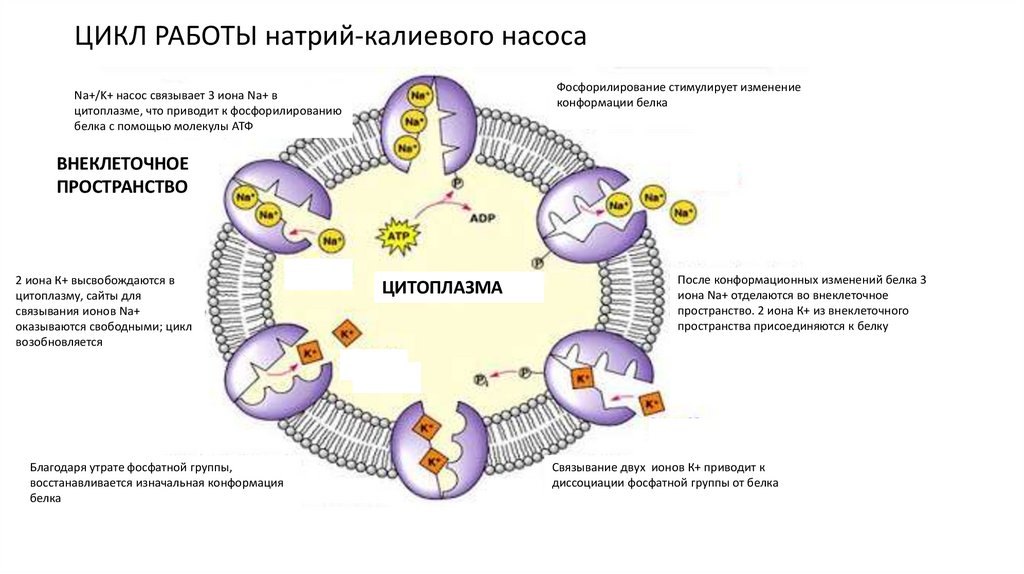
ЦИКЛ РАБОТЫ натрий-калиевого насосаФосфорилирование стимулирует изменение
конформации белка
Na+/K+ насос связывает 3 иона Na+ в
цитоплазме, что приводит к фосфорилированию
белка с помощью молекулы АТФ
ВНЕКЛЕТОЧНОЕ
ПРОСТРАНСТВО
2 иона К+ высвобождаются в
цитоплазму, сайты для
связывания ионов Na+
оказываются свободными; цикл
возобновляется
Благодаря утрате фосфатной группы,
восстанавливается изначальная конформация
белка
ЦИТОПЛАЗМА
После конформационных изменений белка 3
иона Na+ отделаются во внеклеточное
пространство. 2 иона К+ из внеклеточного
пространства присоединяются к белку
Связывание двух ионов К+ приводит к
диссоциации фосфатной группы от белка
28.
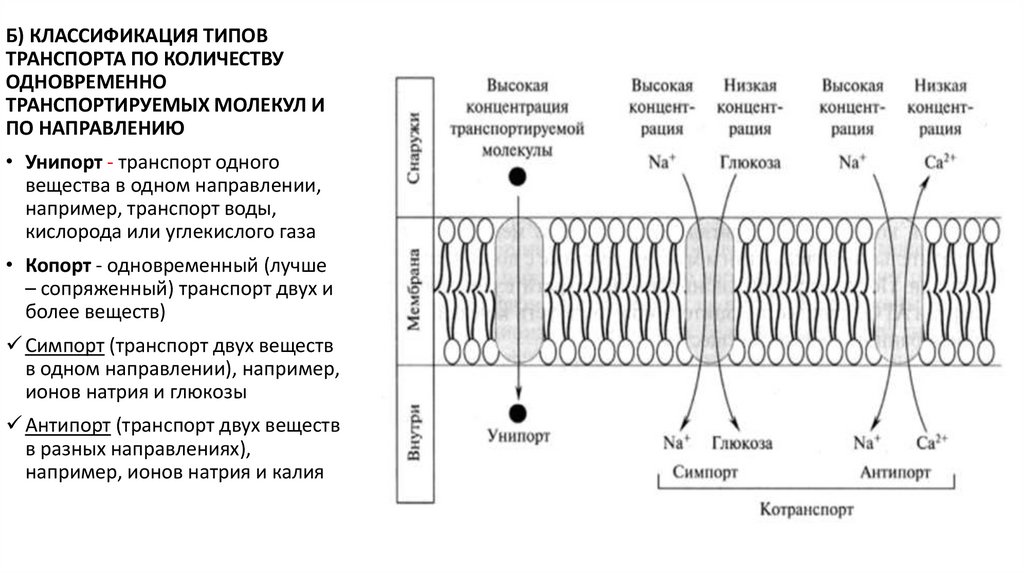
Б) КЛАССИФИКАЦИЯ ТИПОВТРАНСПОРТА ПО КОЛИЧЕСТВУ
ОДНОВРЕМЕННО
ТРАНСПОРТИРУЕМЫХ МОЛЕКУЛ И
ПО НАПРАВЛЕНИЮ
• Унипорт - транспорт одного
вещества в одном направлении,
например, транспорт воды,
кислорода или углекислого газа
• Копорт - одновременный (лучше
– сопряженный) транспорт двух и
более веществ)
Симпорт (транспорт двух веществ
в одном направлении), например,
ионов натрия и глюкозы
Антипорт (транспорт двух веществ
в разных направлениях),
например, ионов натрия и калия
29.
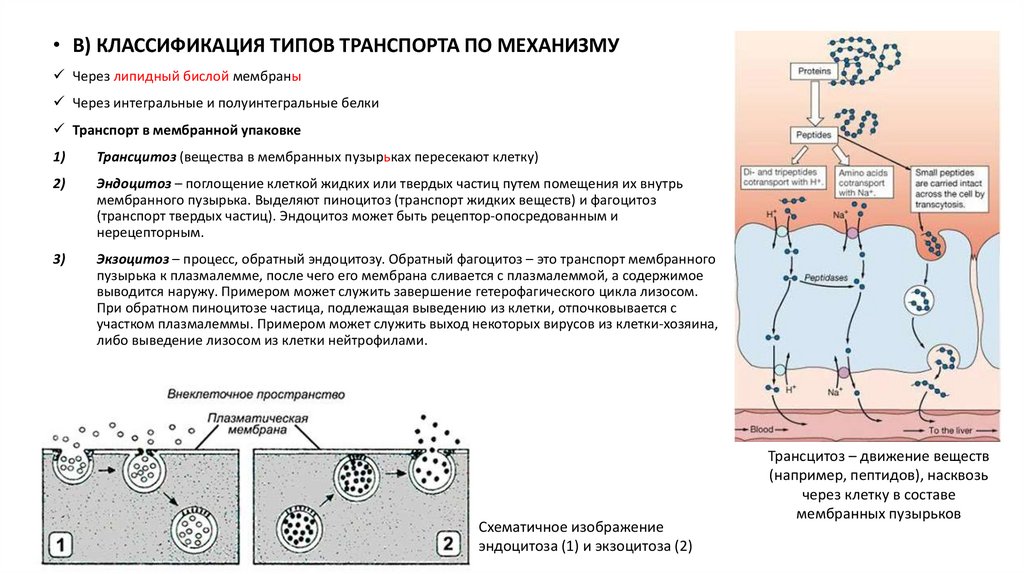
• В) КЛАССИФИКАЦИЯ ТИПОВ ТРАНСПОРТА ПО МЕХАНИЗМУЧерез липидный бислой мембраны
Через интегральные и полуинтегральные белки
Транспорт в мембранной упаковке
1)
Трансцитоз (вещества в мембранных пузырьках пересекают клетку)
2)
Эндоцитоз – поглощение клеткой жидких или твердых частиц путем помещения их внутрь
мембранного пузырька. Выделяют пиноцитоз (транспорт жидких веществ) и фагоцитоз
(транспорт твердых частиц). Эндоцитоз может быть рецептор-опосредованным и
нерецепторным.
3)
Экзоцитоз – процесс, обратный эндоцитозу. Обратный фагоцитоз – это транспорт мембранного
пузырька к плазмалемме, после чего его мембрана сливается с плазмалеммой, а содержимое
выводится наружу. Примером может служить завершение гетерофагического цикла лизосом.
При обратном пиноцитозе частица, подлежащая выведению из клетки, отпочковывается с
участком плазмалеммы. Примером может служить выход некоторых вирусов из клетки-хозяина,
либо выведение лизосом из клетки нейтрофилами.
Схематичное изображение
эндоцитоза (1) и экзоцитоза (2)
Трансцитоз – движение веществ
(например, пептидов), насквозь
через клетку в составе
мембранных пузырьков
30.
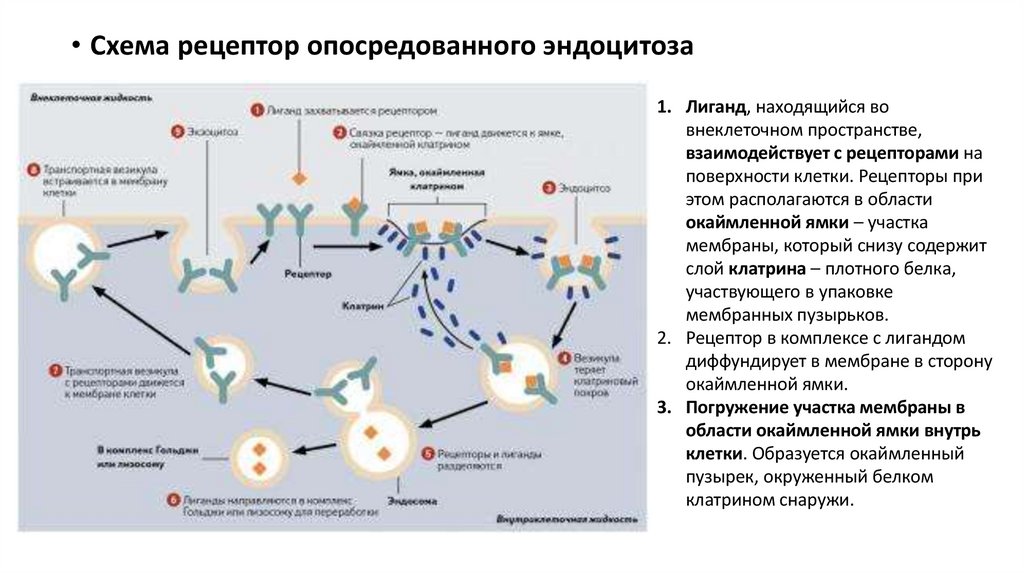
• Схема рецептор опосредованного эндоцитоза1. Лиганд, находящийся во
внеклеточном пространстве,
взаимодействует с рецепторами на
поверхности клетки. Рецепторы при
этом располагаются в области
окаймленной ямки – участка
мембраны, который снизу содержит
слой клатрина – плотного белка,
участвующего в упаковке
мембранных пузырьков.
2. Рецептор в комплексе с лигандом
диффундирует в мембране в сторону
окаймленной ямки.
3. Погружение участка мембраны в
области окаймленной ямки внутрь
клетки. Образуется окаймленный
пузырек, окруженный белком
клатрином снаружи.
31.
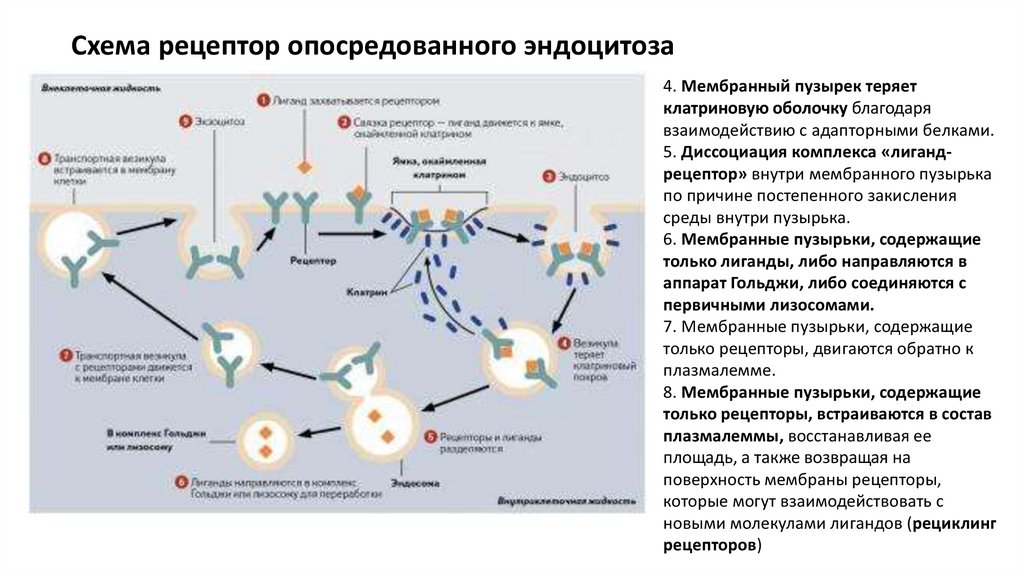
Схема рецептор опосредованного эндоцитоза4. Мембранный пузырек теряет
клатриновую оболочку благодаря
взаимодействию с адапторными белками.
5. Диссоциация комплекса «лигандрецептор» внутри мембранного пузырька
по причине постепенного закисления
среды внутри пузырька.
6. Мембранные пузырьки, содержащие
только лиганды, либо направляются в
аппарат Гольджи, либо соединяются с
первичными лизосомами.
7. Мембранные пузырьки, содержащие
только рецепторы, двигаются обратно к
плазмалемме.
8. Мембранные пузырьки, содержащие
только рецепторы, встраиваются в состав
плазмалеммы, восстанавливая ее
площадь, а также возвращая на
поверхность мембраны рецепторы,
которые могут взаимодействовать с
новыми молекулами лигандов (рециклинг
рецепторов)
32.
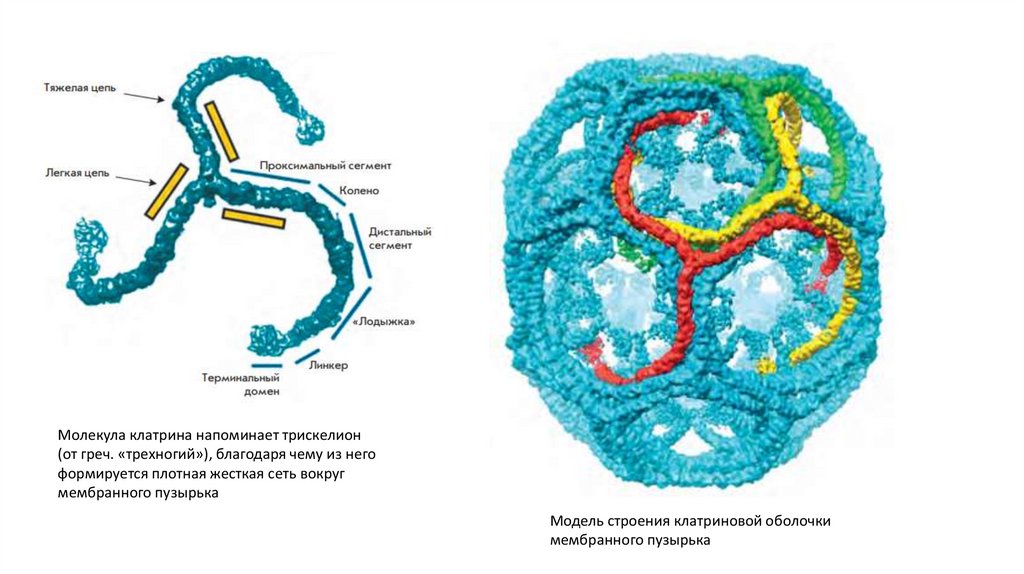
Молекула клатрина напоминает трискелион(от греч. «трехногий»), благодаря чему из него
формируется плотная жесткая сеть вокруг
мембранного пузырька
Модель строения клатриновой оболочки
мембранного пузырька
33. Вопрос 5. Надмембранный комплекс разных клеток
НАДМЕМБРАННЫЙ КОМПЛЕКС ЖИВОТНЫХ КЛЕТОК –ГЛИКОКАЛИКС
Химический состав:
• Гликолипиды (их углеводные радикалы).
• Гликопротеиды (углеводные радикалы надмембранных
доменов рецепторных белков мембраны).
• Белки клеточной адгезии (САМ – cell adgesion molecule),
участвующие в формировании межклеточных контактов
• Белки субстратной адгезии (SАМ - substratе adgesion
molecule) участвующие в формировании временных
контактов клетки с субстратом
• Белки-маркеры (антигены).
• Некоторые углеводы, ех. ацетилгликозамин,
ацетилгалактозамин.
• Ферменты примембранного пищеварения в
эпителиоцитах тонкой кишки. Ех, липаза - расщепляет
ТАГ (триацилглицериды) до глицерина и ВЖК,
гликозидазы - расщепляют олиго-и полисахариды до
моносахаридов, пептидазы расщепляют олиго- и
полипептиды до аминокислот, нуклеазы -расщепляют
нуклеиновые кислоты до нуклеотидов
34.
ФУНКЦИИ ГЛИКОКАЛИКСА:• Антигенная. Белки-маркеры
обеспечивают индивидуальную и видовую
специфичность клетки.
• Рецепторная. С углеводными
радикалами рецепторных белков
связываются сигнальные молекулы (ех.
гормоны или медиаторы), после чего
конформация рецептора изменяется и
сигнал поступает в клетку.
• Формирование межклеточных
контактов (белки САМ).
• Формирование временных контактов
клетки с субстратом, ех. при фагоцитозе
(белки SАМ).
• 5. Примембранное пищеварение
(ферменты -гидролазы, адсорбированные в
гликокаликсе эпителиоцитов тонкой кишки)
35.
НАДМЕМБРАННЫЙ КОМПЛЕКС РАСТИТЕЛЬНЫХ КЛЕТОК - Целлюлозная клеточная стенка (КС)* В клетках грибов - хитиновая клеточная стенка (хитин-полимер ацетилгликозамина)
Основные компоненты целлюлозной КС:
• Целлюлоза- полимер Β-ГЛЮКОЗЫ
• Гемицеллюлоза. 3.Протопектин.
Компоненты видоизмененных клеточных стенок
1.
Лигнин - вызывает лигнинизацию (одревеснение) клеток. Встречается в клетках
• а)
ксилемы - проводящей ткани, обеспечивающей восходящий ток воды и минеральных солей от
корня к другим органам (ех, сосуды и трахеиды);
• б)
склеренхимы - механической ткани, обеспечивающей прочность древесного ствола.
2. Суберин - воскоподобное вещество,
вызывающее суберинизацию (опробковение)
клеток. Встречается в клетках пробки покровной ткани древесного ствола.
3. Кутин - воскоподобное вещество,
вызывающее кутинизацию клеток.
Встречается в клетках эпидермы – покровной
ткани листьев, зеленых стеблей, плодов и др.
4. Соли кальция. Вызывают минерализацию
клеточных стенок. Встречаются в клетках
стеблей злаков.
36.
ФУНКЦИИ КЛЕТОЧНОЙ СТЕНКИ РАСТЕНИЙ:• Барьерная. Препятствует проникновению некоторых веществ в клетку. Растительные
клетки питаются только осмотически, фагоцитоз невозможен.
• Каркасная. КС обеспечивает экзоскелет клетки.
• Защитная. КС обеспечивает механическую защиту клетки, а также защиту от
проникновения бактерий и вирусов.
• Формирование межклеточных
контактов. Ех, цитоплазматические
мостики - плазмодесмы.
• Участие в цитокинезе – делении
цитоплазмы. В телофазе митоза
цитокинез происходит за счет
формирования клеточных стенок
между дочерними клетками.
37.
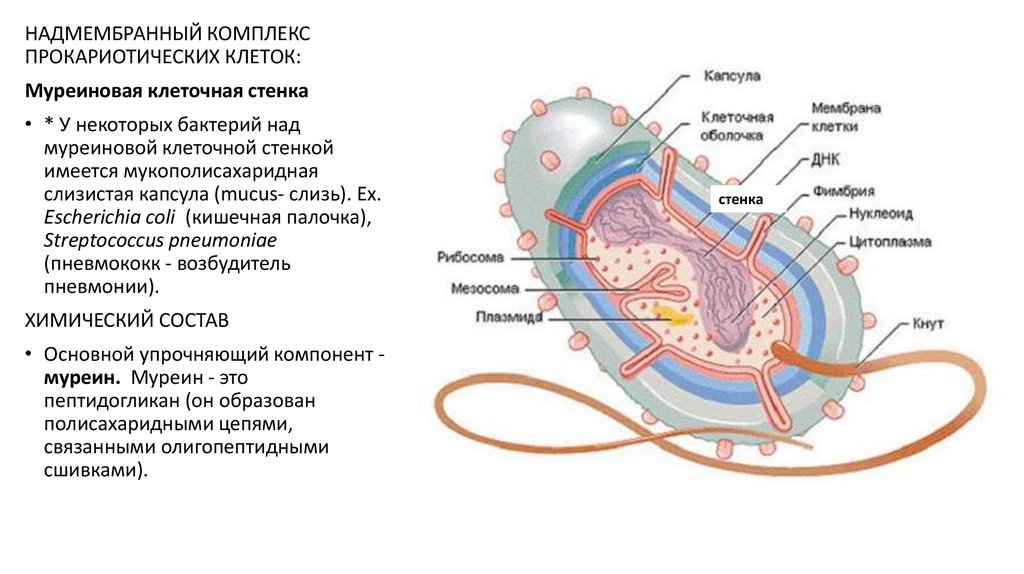
НАДМЕМБРАННЫЙ КОМПЛЕКСПРОКАРИОТИЧЕСКИХ КЛЕТОК:
Муреиновая клеточная стенка
• * У некоторых бактерий над
муреиновой клеточной стенкой
имеется мукополисахаридная
слизистая капсула (mucus- слизь). Ех.
Escherichia coli (кишечная палочка),
Streptococcus pneumoniae
(пневмококк - возбудитель
пневмонии).
ХИМИЧЕСКИЙ СОСТАВ
• Основной упрочняющий компонент муреин. Муреин - это
пептидогликан (он образован
полисахаридными цепями,
связанными олигопептидными
сшивками).
стенка
38.
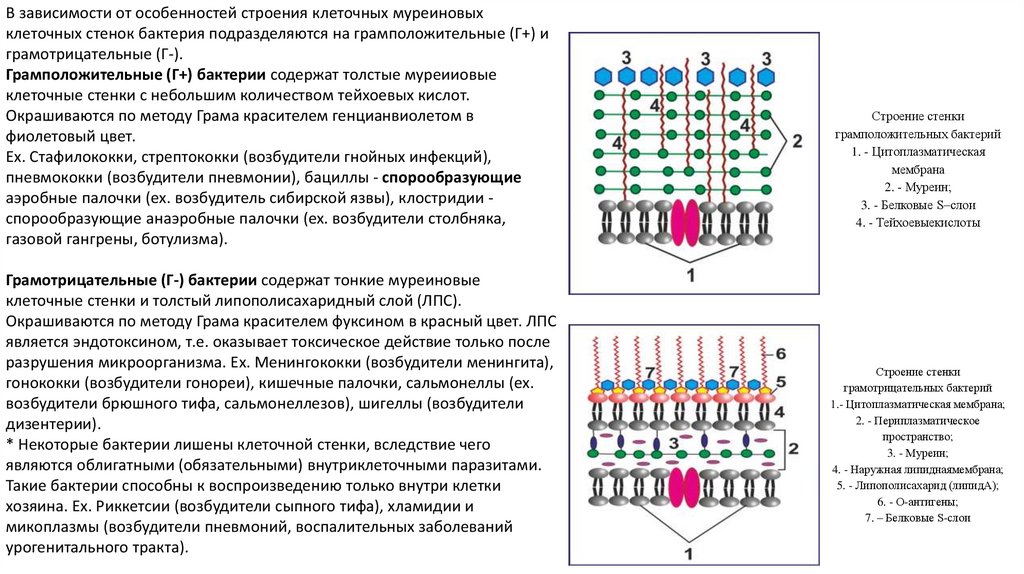
В зависимости от особенностей строения клеточных муреиновыхклеточных стенок бактерия подразделяются на грамположительные (Г+) и
грамотрицательные (Г-).
Грамположительные (Г+) бактерии содержат толстые муреииовые
клеточные стенки с небольшим количеством тейхоевых кислот.
Окрашиваются по методу Грама красителем генцианвиолетом в
фиолетовый цвет.
Ех. Стафилококки, стрептококки (возбудители гнойных инфекций),
пневмококки (возбудители пневмонии), бациллы - спорообразующие
аэробные палочки (ех. возбудитель сибирской язвы), клостридии спорообразующие анаэробные палочки (ех. возбудители столбняка,
газовой гангрены, ботулизма).
Грамотрицательные (Г-) бактерии содержат тонкие муреиновые
клеточные стенки и толстый липополисахаридный слой (ЛПС).
Окрашиваются по методу Грама красителем фуксином в красный цвет. ЛПС
является эндотоксином, т.е. оказывает токсическое действие только после
разрушения микроорганизма. Ех. Менингококки (возбудители менингита),
гонококки (возбудители гонореи), кишечные палочки, сальмонеллы (ех.
возбудители брюшного тифа, сальмонеллезов), шигеллы (возбудители
дизентерии).
* Некоторые бактерии лишены клеточной стенки, вследствие чего
являются облигатными (обязательными) внутриклеточными паразитами.
Такие бактерии способны к воспроизведению только внутри клетки
хозяина. Ех. Риккетсии (возбудители сыпного тифа), хламидии и
микоплазмы (возбудители пневмоний, воспалительных заболеваний
урогенитального тракта).
Строение стенки
грамположительных бактерий
1. - Цитоплазматическая
мембрана
2. - Муреин;
3. - Белковые S–слои
4. - Тейхоевыекислоты
Строение стенки
грамотрицательных бактерий
1.- Цитоплазматическая мембрана;
2. - Периплазматическое
пространство;
3. - Муреин;
4. - Наружная липиднаямембрана;
5. - Липополисахарид (липидА);
6. - О-антигены;
7. – Белковые S-слои
39.
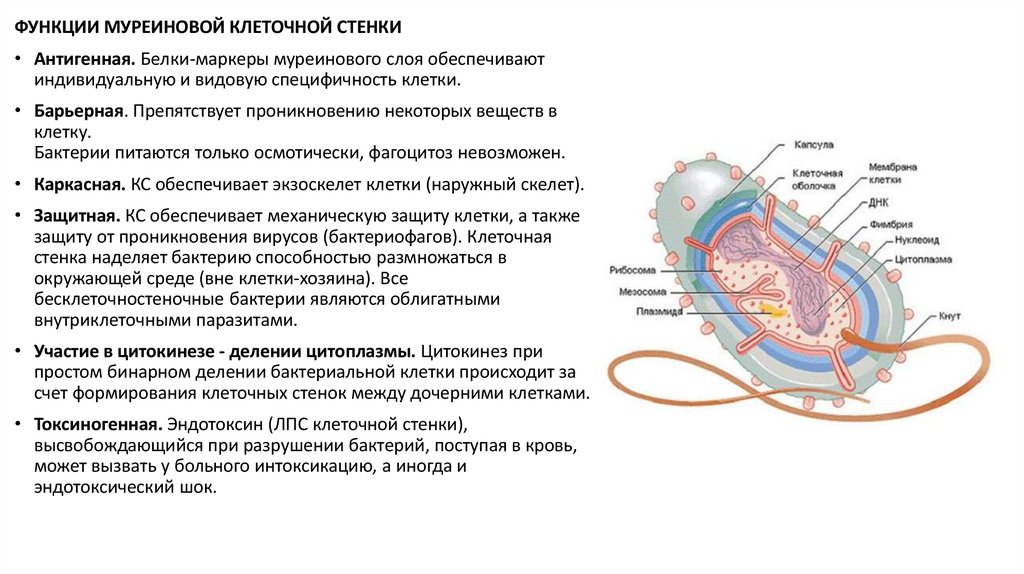
ФУНКЦИИ МУРЕИНОВОЙ КЛЕТОЧНОЙ СТЕНКИ• Антигенная. Белки-маркеры муреинового слоя обеспечивают
индивидуальную и видовую специфичность клетки.
• Барьерная. Препятствует проникновению некоторых веществ в
клетку.
Бактерии питаются только осмотически, фагоцитоз невозможен.
• Каркасная. КС обеспечивает экзоскелет клетки (наружный скелет).
• Защитная. КС обеспечивает механическую защиту клетки, а также
защиту от проникновения вирусов (бактериофагов). Клеточная
стенка наделяет бактерию способностью размножаться в
окружающей среде (вне клетки-хозяина). Все
бесклеточностеночные бактерии являются облигатными
внутриклеточными паразитами.
• Участие в цитокинезе - делении цитоплазмы. Цитокинез при
простом бинарном делении бактериальной клетки происходит за
счет формирования клеточных стенок между дочерними клетками.
• Токсиногенная. Эндотоксин (ЛПС клеточной стенки),
высвобождающийся при разрушении бактерий, поступая в кровь,
может вызвать у больного интоксикацию, а иногда и
эндотоксический шок.
40.
Контрольно-обучающие вопросы по теме занятия:• Задание 1. Дайте характеристику рецепторно-барьернотранспортной системы клетки и ее биологического значения.
• Задание 2. Сформулируйте современные представления о
строении и свойствах биологических мембран клетки и
плазмалеммы.
• Здание 3. Охарактеризуйте структурно-функциональную
организацию поверхностного аппарата клетки.
• Задание 4. Опишите основные виды трансмембранного
транспорта и дайте характеристику их биологического значения.
• Задание 5. Расскажите об основных типах межклеточных
контактов.
41. Контрольно-обучающие тесты по теме занятия:
1. Текучесть плазмалеммы и ее способность к самосборке обусловлены свойствами входящих в еесостав молекул:
а) липидов; б) белков; в) полисахаридов; г) всех указанных соединений
2. Плазмалемма непроницаема для
а) ионов; б) малых незаряженных молекул; в) малых гидрофобных молекул;
г) молекул воды; д) всех указанных соединений.
3. Только в наружном слое плазмалеммы животной клетки располагаются:
а) липиды; б) гликопротеиды; в) липопротеиды; г) все указанные соединения.
4. Толщина гликокаликса в животных клетках составляет:
а) 1-2 нм;
б) 3-4 нм;
в) 10-15 нм;
г) 20-30 нм;
д) 35 -50 нм.
5. Транспорт, при котором белок-переносчик функционирует только в отношении молекул или
ионов одного вида
а) унипорт;
б) копорт;
в) симпорт;
г) антипорт.
42.
Практическая работа на занятии43. Клетки листа элодеи – E. сanadensis: 1 - клеточная стенка; 2 - цитоплазма; 3 - хлоропласты.
• Задание 1. Изучение временного препарата листаэлодеи канадской - Elodea canadensis.
• Учебный временный препарат листа элодеи
канадской - E. сanadensis. Препарат изучается под
малым и большим увеличением микроскопа.
• Лист отделите от побега элодеи, предварительно
выдержанного в течение 1-2 часов в теплой воде при
ярком освещении. На предметное стекло нанесите
каплю теплой воды, в которую поместите кусочек
листа элодеи наружной стороной вверх. Содержимое
капли накройте покровным стеклом.
• При большом увеличении микроскопа найдите
клетки листа элодеи. Они имеют прямоугольную
форму с хорошо выраженной толстой бесцветной
оболочкой. В цитоплазме клеток отчетливо видны
многочисленные тельца зеленого цвета округлой и
овальной формы – хлоропласты.
• Ядра в неокрашенных клетках не видны. Обратите
внимание, что цитоплазма клеток тесно прижата к
клеточным стенкам. Это свидетельствует о том, что
клетки находятся в состоянии полного насыщения
водой, т.е. в состоянии тургора.
1
2
3
Клетки листа элодеи –
E. сanadensis:
1 - клеточная стенка;
2 - цитоплазма;
3 - хлоропласты.
44. Плазмолиз в клетках листа элодеи канадской – E. сanadensis: 1 – клетка в тургесцентном состоянии; 2 – плазмолизированные
•Задание 2. Изучение плазмолиза и деплазмолиза в клетках листа элодеиканадской - E. сanadensis:
• Плазмолиз характерен для растительных клеток, у которых имеется жесткая
клеточная стенка, благодаря чему их плазмалемма в обычных условиях прижата
действием тургорного давления к клеточной стенке. При помещении растительной
клетки в гипертонический раствор скорость диффузии воды из клетки будет
превышать скорость диффузии воды в клетку из окружающего раствора. Объем
клеточного сока при этом уменьшится, вследствие чего произойдет сокращение
объема центральной вакуоли и отделение протопласта от клеточной стенки. Это
явление получило название плазмолиза. Если плазмолизированные клетки пребывают
в этом состоянии короткое время, то они остаются живыми. При помещении такой
клетки в гипотонический раствор хлорида натрия вода начнет поступать в клетку, в
результате чего произойдет деплазмолиз – возвращение клетки в тургесцентное
состояние.
•Для наблюдения плазмолиза в приготовленном временном препарате листа элодеи
замените воду гипертоническим раствором хлорида натрия. С этой целью на
предметное стекло с одного края покровного стекла нанесите каплю гипертонического
раствора, а с противоположного края стекла с помощью фильтровальной бумаги
удалите воду, находящуюся под покровным стеклом. В результате на место воды под
покровное стекло начнет поступать гипертонический раствор хлорида натрия. Для
полной замены воды гипертоническим раствором указанную процедуру повторите 2-3
раза. По прошествии 5 - 10 минут, при микроскопии препарата найдите
плазмолизированные клетки.
•Для наблюдения деплазмолиза на поверхность временного микропрепарата листа
элодеи с плазмолизированными клетками нанесите несколько капель
дистиллированной воды непосредственно у края покровного стекла. После этого с
помощью фильтровальной бумаги удалите гипертонический раствор хлорида натрия,
находящийся под стеклом. По окончании указанной процедуры изучите препарат под
малым и большим увеличением микроскопа. Обратите внимание на восстановление
положения клеточной мембраны, характерного для клетки в состоянии тургора.
Объясните, какое свойство плазмалеммы лежит в основе явлений плазмолиза и
деплазмолиза.
2
2
1
2
Плазмолиз в клетках
листа элодеи канадской –
E. сanadensis:
1 – клетка в тургесцентном
состоянии;
2 – плазмолизированные
клетки.
45.
Что изображено на электронограмме?Какая структура соответствует указанной цифре?
1
1
46.
Что изображено на электронограмме?Чему соответствуют темные и светлые полосы?
2
47.
Что изображено на рисунке?Какие структуры соответствует указанным цифрам?
48.
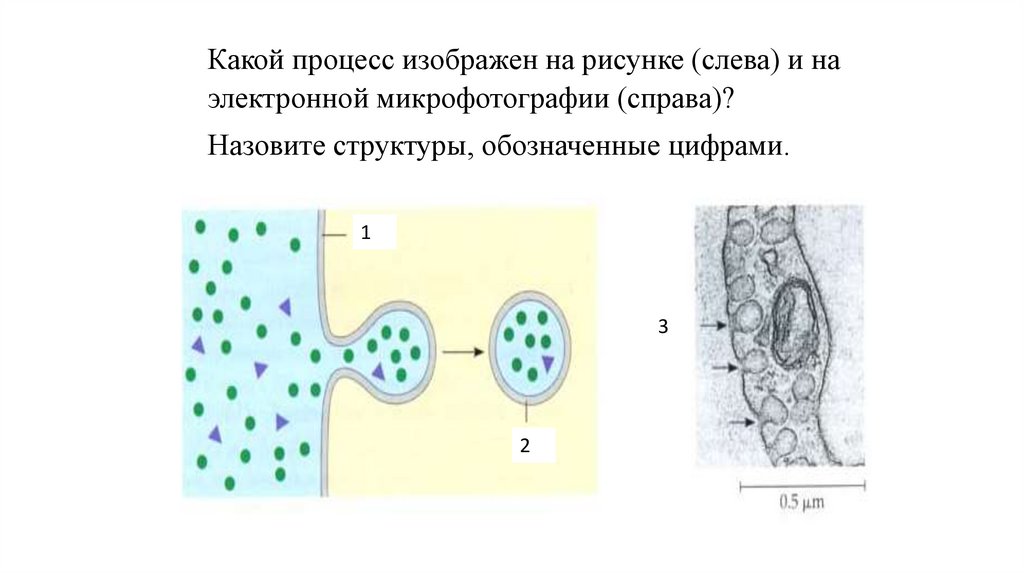
Какой процесс изображен на рисунке (слева) и наэлектронной микрофотографии (справа)?
Назовите структуры, обозначенные цифрами.
1
3
2
49.
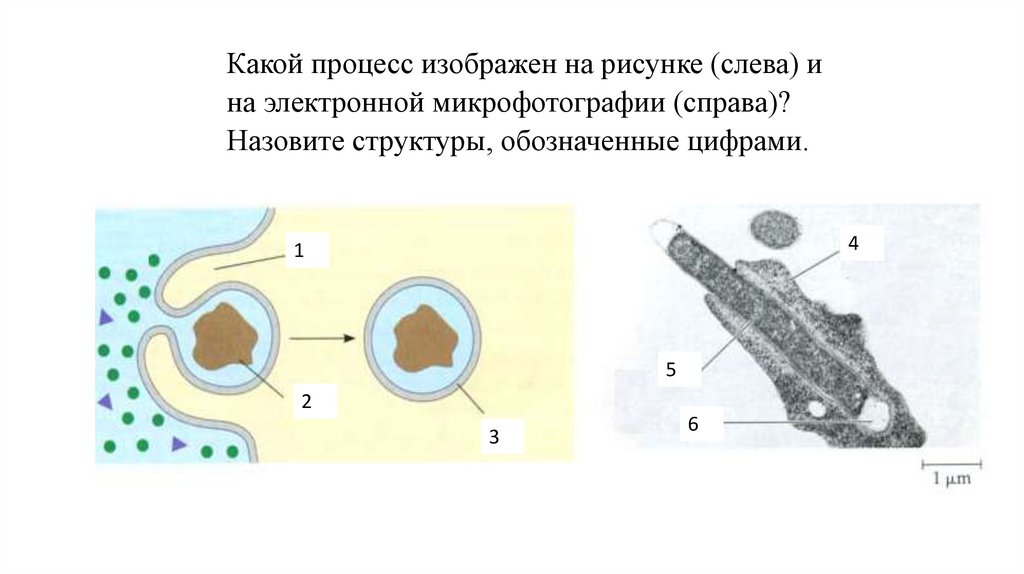
Какой процесс изображен на рисунке (слева) ина электронной микрофотографии (справа)?
Назовите структуры, обозначенные цифрами.
4
1
5
2
3
6
50.
ТЕСТЫ ДЛЯОБСУЖДЕНИЯ
51.
Вопрос 1.Текучесть плазмалеммы и ее способность к
самосборке обусловлены свойствами входящих в
ее состав молекул:
а) липидов;
б) белков;
в) полисахаридов;
г) всех указанных соединений.
52.
Вопрос 2.Белок клатрин принимает участие в процессах:
а) диффузии;
б) осмоса;
в) эндоцитоза;
г) транспорта ионов Na+ и K+;
д) транспорта глюкозы и Na+.
53.
Вопрос 3. Транспорт, при котором белокпереносчик функционирует только в отношениимолекул или ионов одного вида называется
а) унипортом;
б ) копортом;
в) симпортом;
г) антипортом.
54.
Вопрос 4. Одновременный транспорт глюкозы иNa+ осуществляется путем
а) антипорта;
б) унипорта;
в) симпорта;
г) экзоцитоза.
55.
Вопрос 5. Укажите, какая функция поверхностногоаппарата клетки осуществляется с участием белка
гликофорина,
являющегося
структурным
компонентом агглютининогенов эритроцитов крови
человека
а) транспортная;
б) метаболическая;
в) маркерная;
г) локомоторная.
56.
Домашнее задание кзанятию №4:
Гиалоплазма и система синтеза, сегрегации и
внутриклеточного транспорта биополимеров
(вакуолярная система):
- гиалоплазма и органеллы;
- рибосомы и их функции;
- эндоплазматический ретикулум;
- аппарат Гольджи;
- лизосомы, циклы лизосом.