
















































 Биология
Биология Экология
ЭкологияПохожие презентации:










Еволюція екосистем
1.
Еволюція екосистем2.
Структура БіогеоценозівСтруктура
ставкового
біоценозу
Луки. Структура
біоценозу
Схема заростання озера
3.
Структура БіогеоценозівБіоценоз
Консорції
1
Консорція 1
Консорція 1
Консорція 1
2
Консорція 1
3
Сінузії
Структура
біоценозу
тропічного лісу
на острові
Калімантан
4
4.
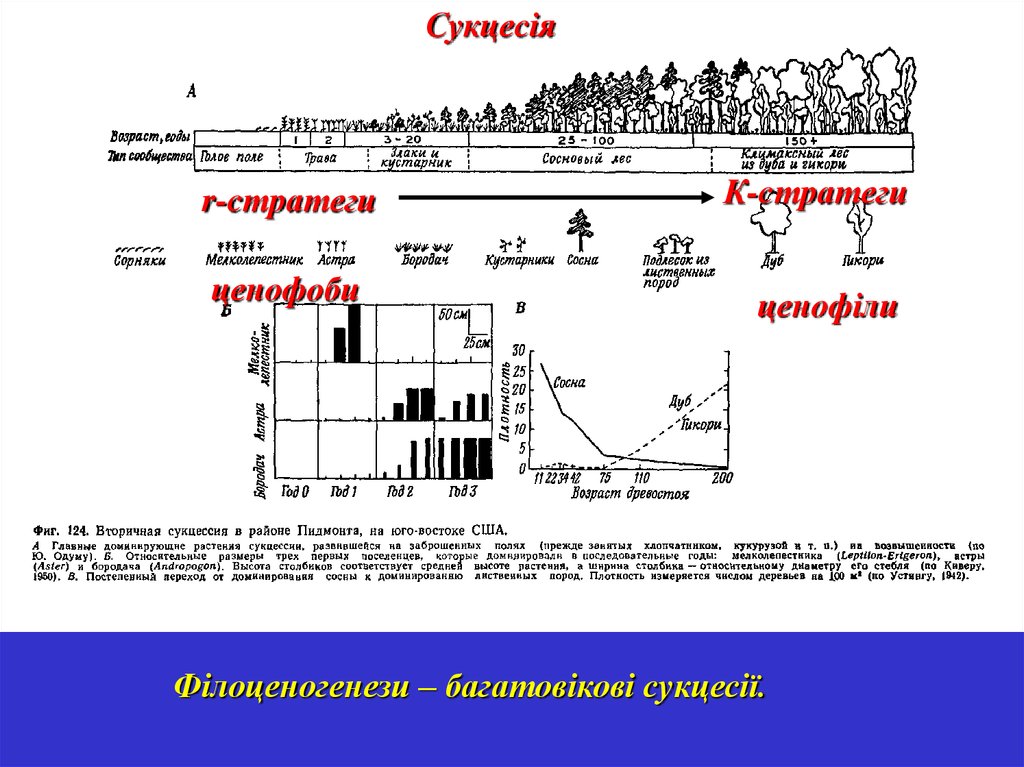
Сукцесіяr-стратеги
ценофоби
К-стратеги
ценофіли
Філоценогенези – багатовікові сукцесії.
5.
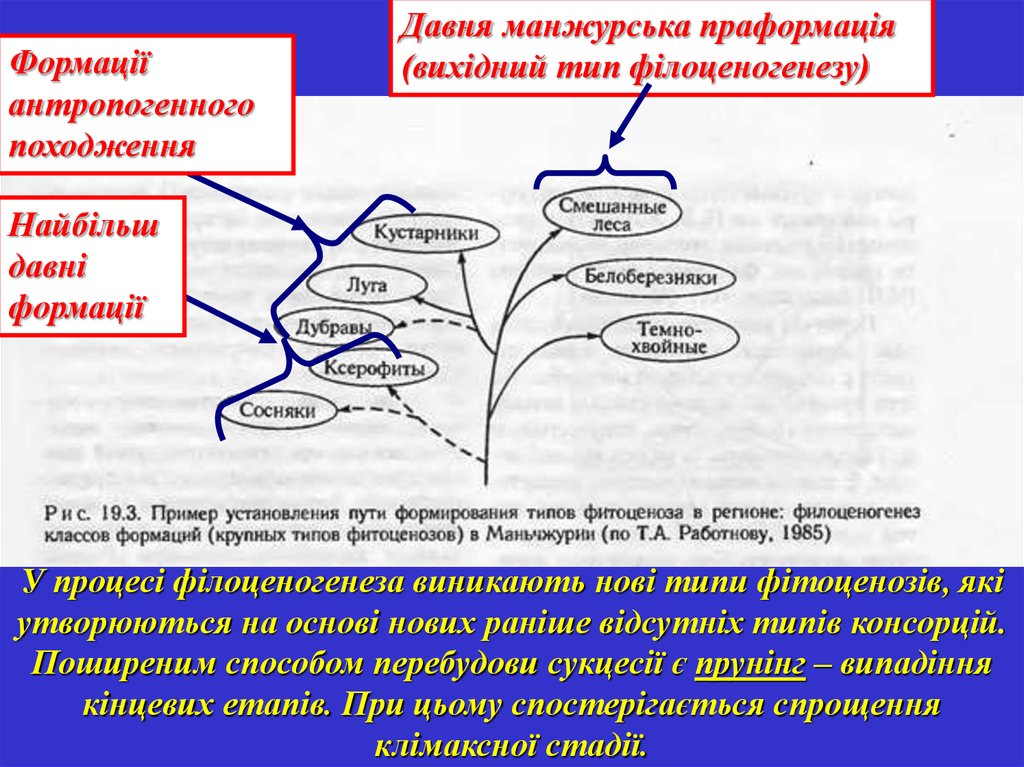
Формаціїантропогенного
походження
Давня манжурська праформація
(вихідний тип філоценогенезу)
Найбільш
давні
формації
У процесі філоценогенеза виникають нові типи фітоценозів, які
утворюються на основі нових раніше відсутніх типів консорцій.
Поширеним способом перебудови сукцесії є прунінг – випадіння
кінцевих етапів. При цьому спостерігається спрощення
клімаксної стадії.
6.
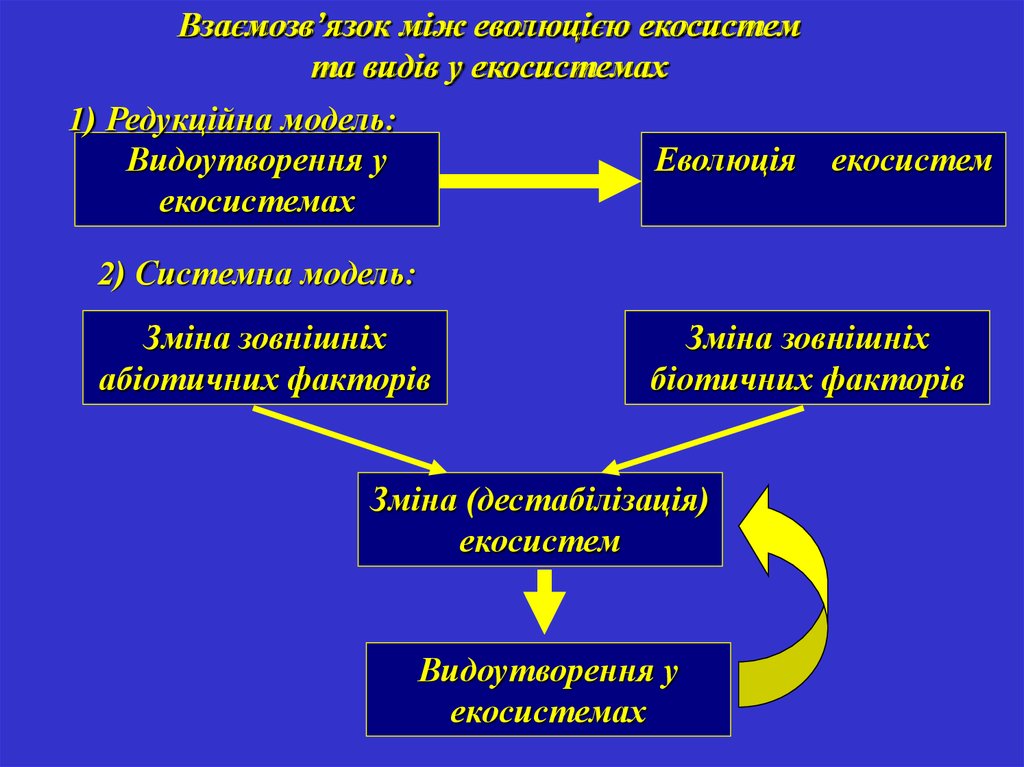
Взаємозв’язок між еволюцією екосистемта видів у екосистемах
1) Редукційна модель:
Видоутворення у
екосистемах
Еволюція
екосистем
2) Системна модель:
Зміна зовнішніх
абіотичних факторів
Зміна зовнішніх
біотичних факторів
Зміна (дестабілізація)
екосистем
Видоутворення у
екосистемах
7.
Ми не можемо чекати милостей від природи!Бобри (?)
Плотина з гілок каміння і намулу - результат діяльності
кількох поколінь бобрів. Деякі боброві плотини сягають кількох
кілометрів у довжину.
8.
Некогерентна еволюція – швидкі і частонезбалансовані зміни у нестабільних екосистемах
Погіршення умов,
посилення боротьби за
існування
дестабілізація
екосистем
Послаблення боротьби за
існування
Порушення
зв’язку
середовищеорганізм в
онтогенезі
Посилення
неспецифічної
елімінації
Зняття тиску
стабілізуючого
добору
Радіація форм
Швидке видоутворення
інадаптивних форм
Вимирання
видів (елізія)
Утворення
екологічних
вакансій
Вселення видів –
імігрантів (інвазія)
9.
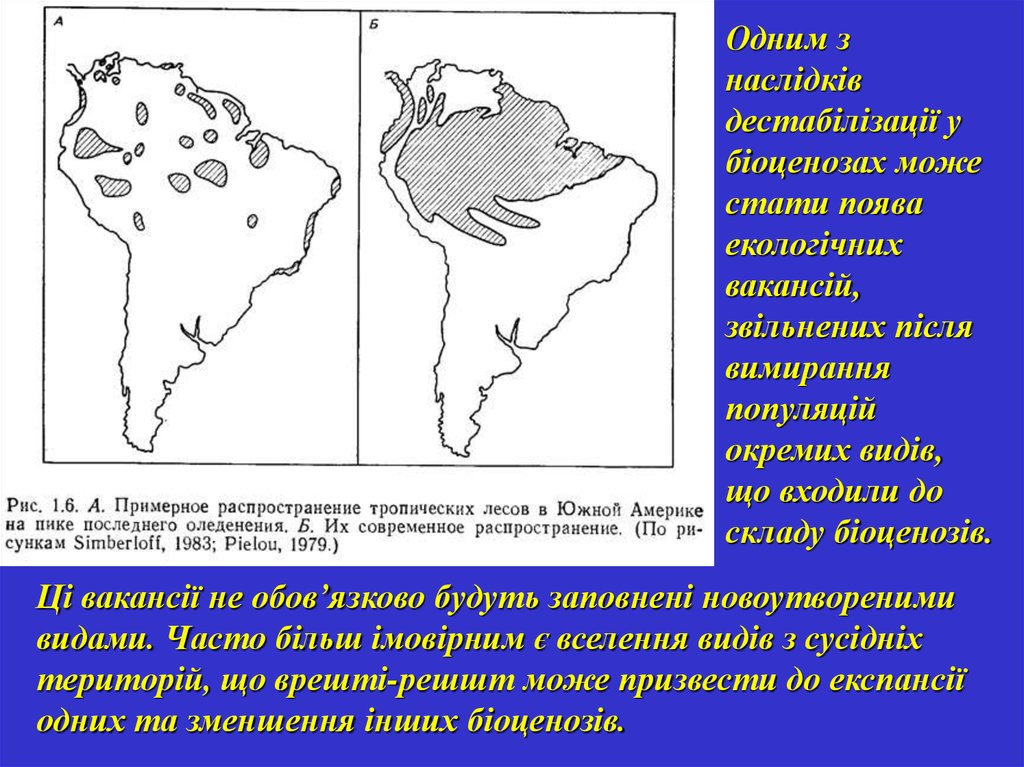
Одним знаслідків
дестабілізації у
біоценозах може
стати поява
екологічних
вакансій,
звільнених після
вимирання
популяцій
окремих видів,
що входили до
складу біоценозів.
Ці вакансії не обов’язково будуть заповнені новоутвореними
видами. Часто більш імовірним є вселення видів з сусідніх
територій, що врешті-решшт може призвести до експансії
одних та зменшення інших біоценозів.
10.
При доместикації імовірність прояву відхилень онтогенезузбільшується. Так у різних видів свійських тварин з’являється
рябе забарвлення, невластиве для їхніх диких сородичів
Рябий кіт
Рябий кінь
Рябий поні
Рябий пес
11.
При несприятливих умовах репродуктивні бар’єри міжвидами можуть порушуватись.
В деяких місцях
на Далекому
Сході основну
масу популяцій
складають
гібриди
благородного і
плямистого
оленей
Кряква та американська чорна качка
активно гібридизують з 1960-х рр.
12.
Поява морфозу може бути частковоскомпенсована завдяки зміненій
поведінці.
Так особина трупіала Sturnella neglecta,
яка мала аномально довгий дзьоб, була
знайдена у незвичному для трупіалів місці
з пухким грунтом, де птах здобував їжу в
незвичний для трупіалів спосіб: він
зондував грунт у пошуках безхребетних.
13.
ЕзогенезПапуга кеа (Nestor notabilis ) харчувався фруктами та нектаром
квітів, доки до Нової Зеландії не було завезено вівців. Після цього
кеа став хижаком.
14.
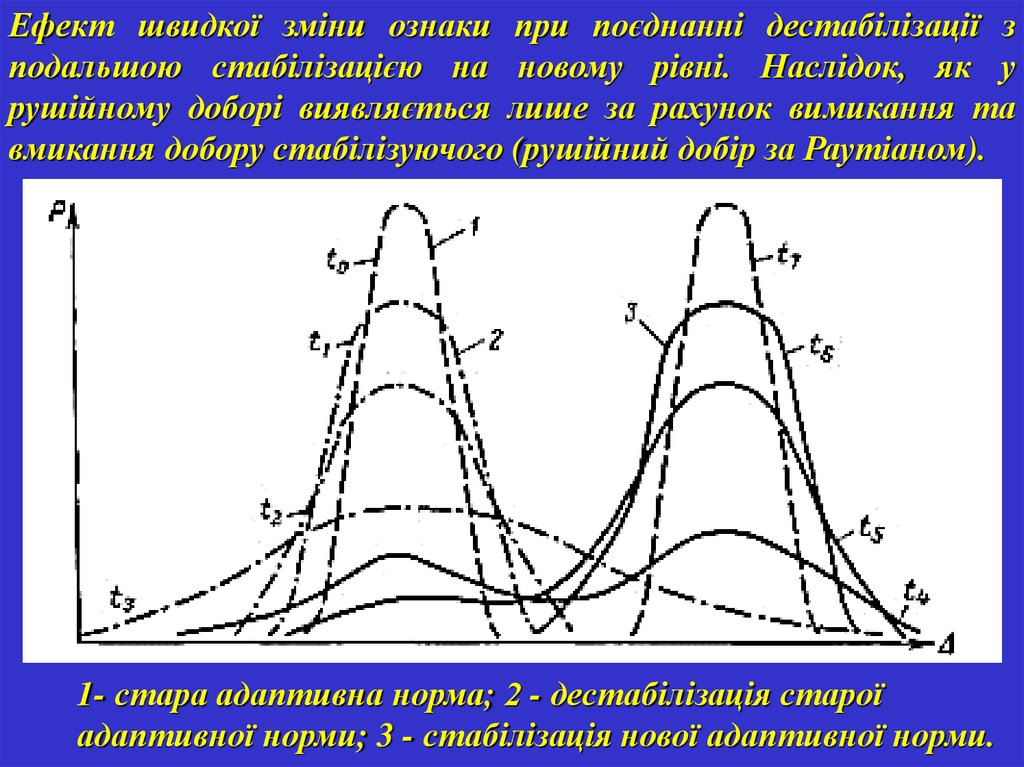
Ефект швидкої зміни ознаки при поєднанні дестабілізації зподальшою стабілізацією на новому рівні. Наслідок, як у
рушійному доборі виявляється лише за рахунок вимикання та
вмикання добору стабілізуючого (рушійний добір за Раутіаном).
1- стара адаптивна норма; 2 - дестабілізація старої
адаптивної норми; 3 - стабілізація нової адаптивної норми.
15.
Симпатричне видоутворення у природі:молюски Аральського моря
Донні
фільтратори,
занурені у
грунт
Позагрунтові
донні
фільтратори
Вихідне
положення
(початок 80-х
рр.)
Cerastoderma
isthmicum
Сучасний
стан (2000-і
рр.)
Dreissena (3
спеціалізовані
види)
Cerastoderma
sp.
Cerastoderma
isthmicum
16.
Спеціогенез(видоутворення)
Cerastoderma isthmicum - намулоїд (вихідна форма - 1)
та Cerastoderma sp. - фільтруюча форма (2)
17.
Видоутворення в умовах островних екосистемАдаптивна радіація гавайських квіткарок - приклад
видоутворення, викликаного екологічними причинами (способом
харчування).
Бруньки
Соковиті плоди
Насіння і ягоди
Нектар
Комахи у
корі дерев
18.
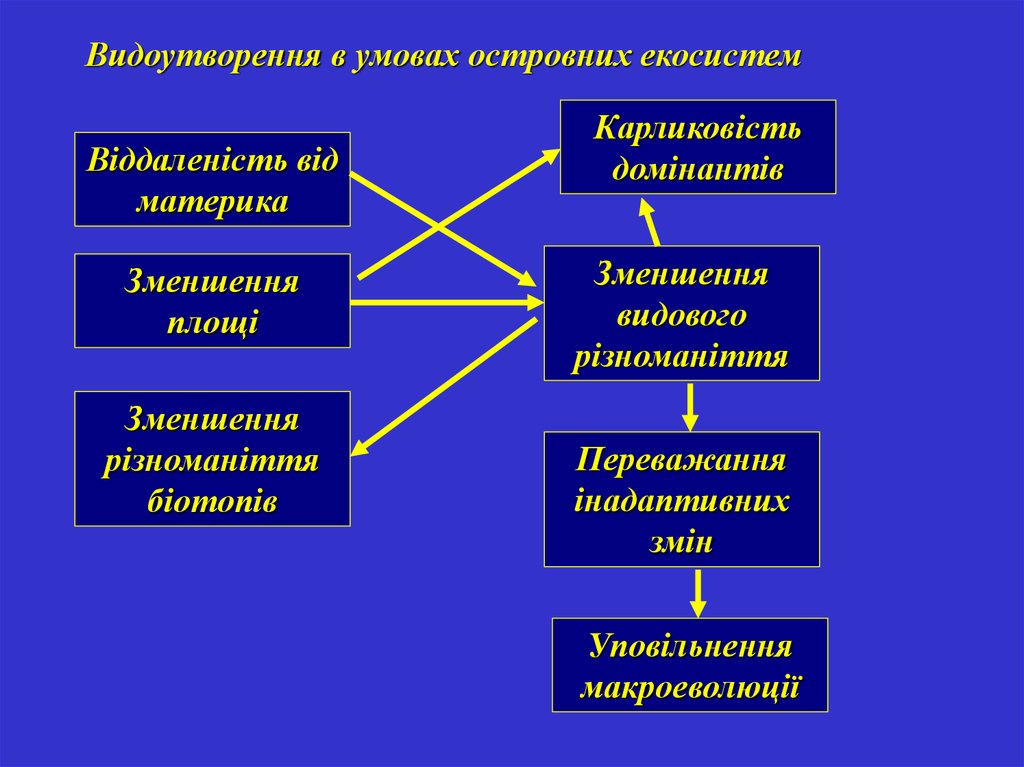
Видоутворення в умовах островних екосистемВіддаленість від
материка
Зменшення
площі
Зменшення
різноманіття
біотопів
Карликовість
домінантів
Зменшення
видового
різноманіття
Переважання
інадаптивних
змін
Уповільнення
макроеволюції
19.
Карликовийстегодон
20.
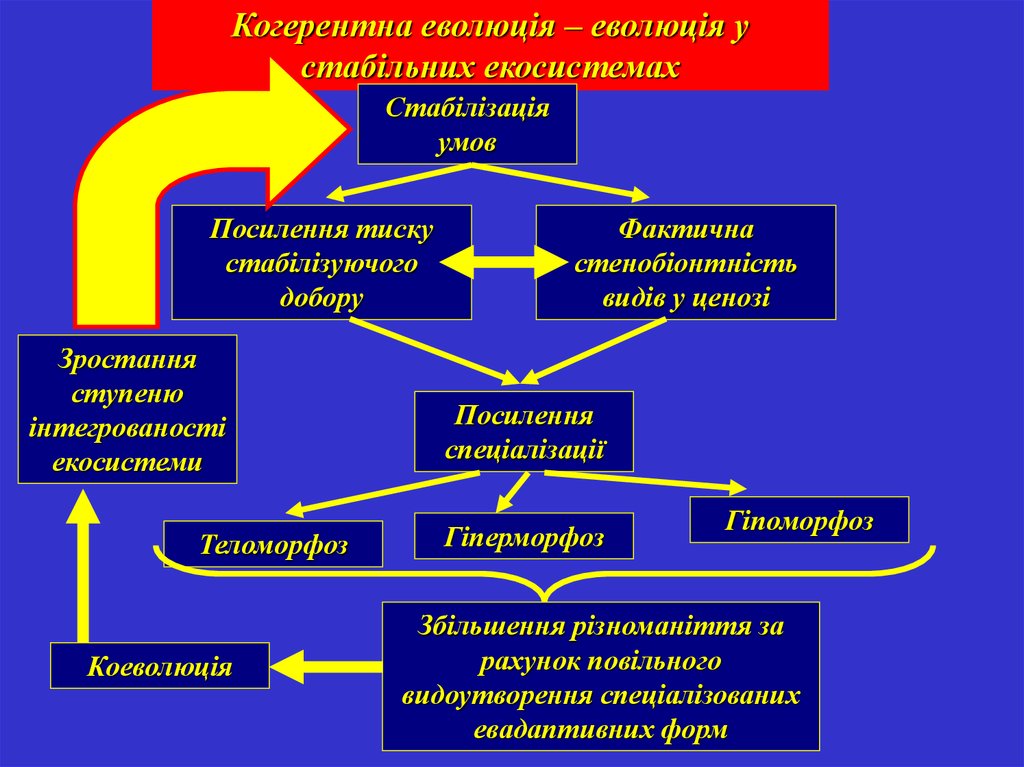
Когерентна еволюція – еволюція устабільних екосистемах
Стабілізація
умов
Посилення тиску
стабілізуючого
добору
Зростання
ступеню
інтегрованості
екосистеми
Теломорфоз
Коеволюція
Фактична
стенобіонтність
видів у ценозі
Посилення
спеціалізації
Гіперморфоз
Гіпоморфоз
Збільшення різноманіття за
рахунок повільного
видоутворення спеціалізованих
евадаптивних форм
21.
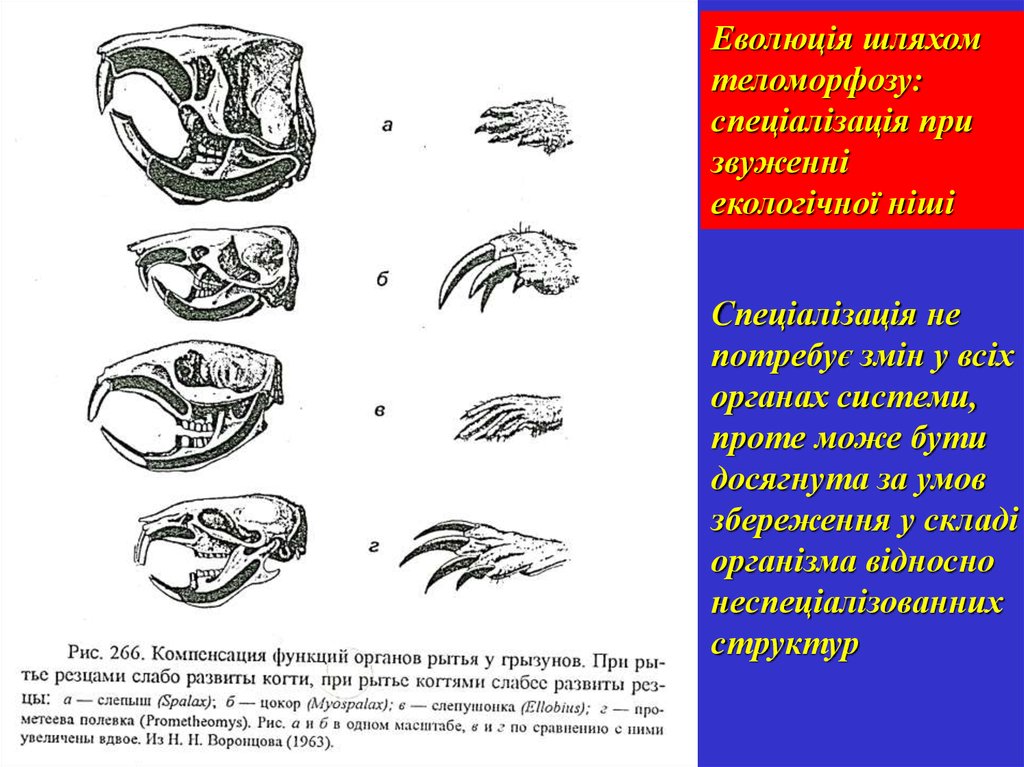
Еволюція шляхомтеломорфозу:
спеціалізація при
звуженні
екологічної ніші
Спеціалізація не
потребує змін у всіх
органах системи,
проте може бути
досягнута за умов
збереження у складі
організма відносно
неспеціалізованних
структур
22.
Еволюція шляхом гіперморфозу: Закон Копа-ДепереПредком могутнього плейстоценового південного слона був
невеличкий ссавець мерітерій.
23.
Еволюція шляхомгіпоморфозу
Амбістома
Аксолотль
24.
Коеволюція - еволюція екологічнопов’язаних таксонів
Симбіонт - хазяїн
Хижак - жертва
25.
Мурахи-листорізихарчуються грибом, який вони
вирощують на субстраті з
подрібненого листя.
А з аскоміцетом-паразитом
плантацій мурахи борються за
допомогою антибіотиків, які
виділяють аскоміцети, яких
мурахи переносять у криптах
на поверхні свого тіла.
26.
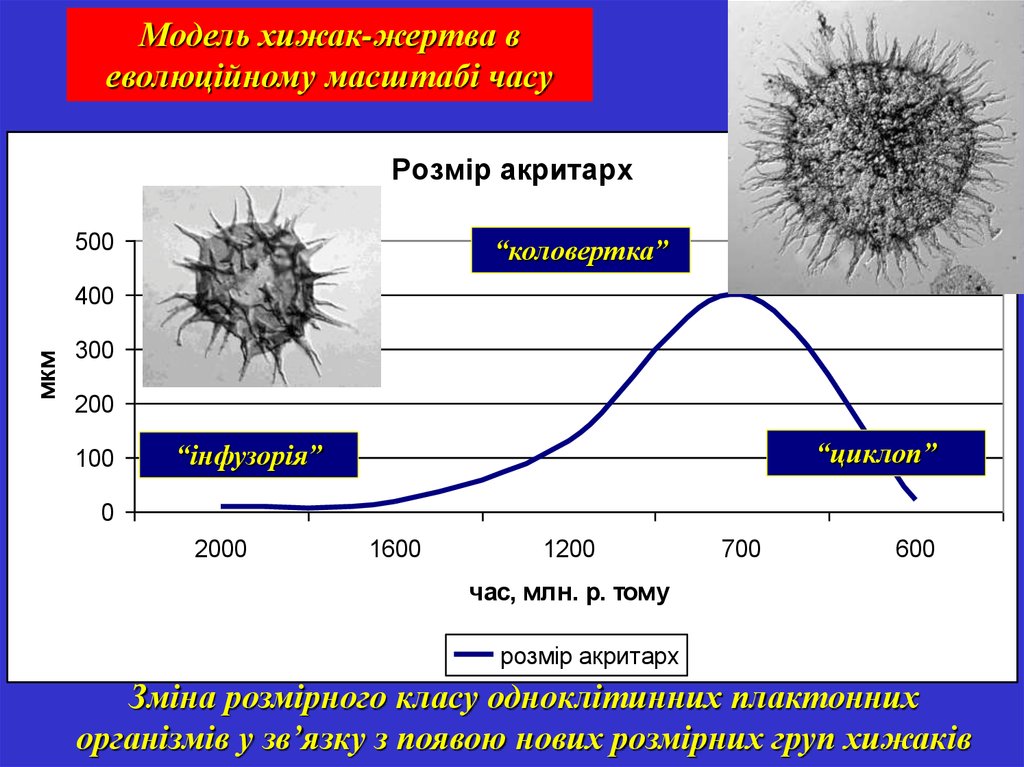
Модель хижак-жертва веволюційному масштабі часу
Розмір акритарх
500
“коловертка”
мкм
400
300
200
100
“циклоп”
“інфузорія”
0
2000
1600
1200
700
600
час, млн. р. тому
розмір акритарх
Зміна розмірного класу одноклітинних плактонних
організмів у зв’язку з появою нових розмірних груп хижаків
27.
Взаємодія хижака та його жертви веволюційному масштабі часу
1. Поява нового типу фітофага (“ жертва 1”)
2. Поява нового типу хижака . “ Жертва 1” випереджає хижака за
ступенем спеціалізації, проте риси спецчіалізації жертви є
інадаптивними
3. Прогресивна спеціалізація хижака йде швидше ніж жертви 1,
має комплексний характер, інадаптивний груз при цьому є меншим
4. Повільне зростання пресу хижака на “ жертву 1 ” . Початок
вимирання “жертви 1”. Вивільнення екологічної ліцензії, яку
починає опановувати фітофаг нового покоління: “жертва 2”, яка
стає додатковим (не головним) ресурсом для хижака
5. Хижак та “ жертва 2” витісняють “ жертву 1” шляхом прямої
та непрямої конкуренції відповідно. “ Жертва 2” стає основним
ресурсом для хижака по мірі вимирання “жертви 1”.
28.
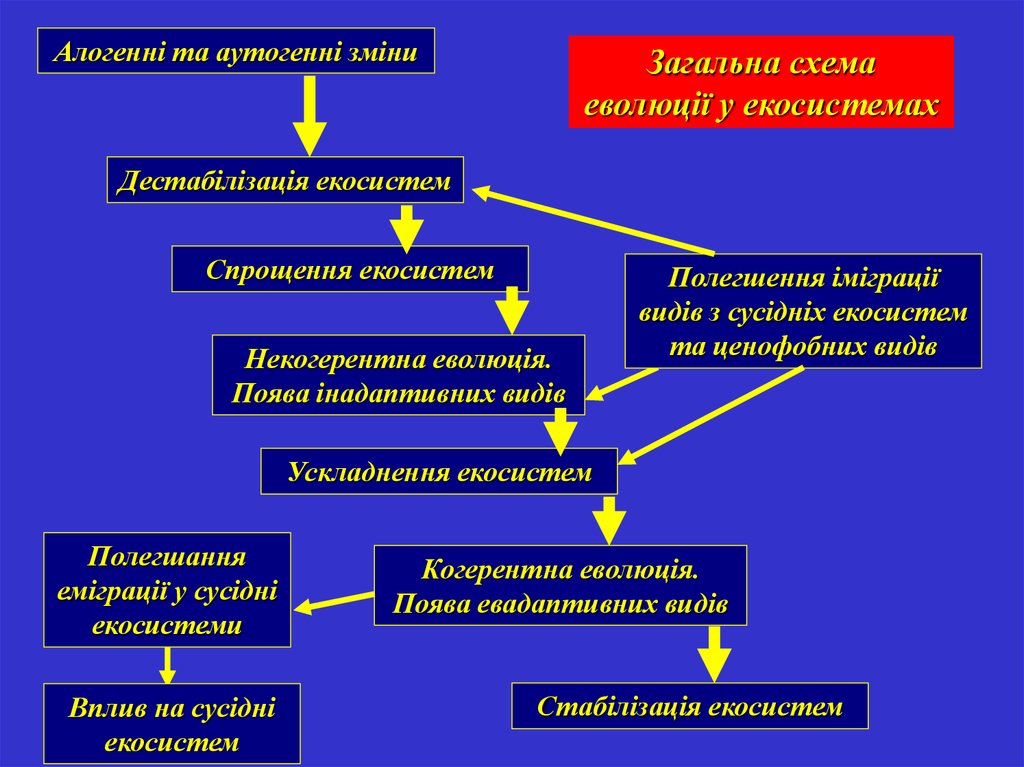
Алогенні та аутогенні зміниЗагальна схема
еволюції у екосистемах
Дестабілізація екосистем
Спрощення екосистем
Некогерентна еволюція.
Поява інадаптивних видів
Полегшення іміграції
видів з сусідніх екосистем
та ценофобних видів
Ускладнення екосистем
Полегшання
еміграції у сусідні
екосистеми
Вплив на сусідні
екосистем
Когерентна еволюція.
Поява евадаптивних видів
Стабілізація екосистем
29.
Елементарні акти філоценогенезу:1) езогенез – перегрупування зв’язків у екосистемі –
перебудова екологічної ніші
2) спеціогенез – видоутворення – поява нововї екологічної
ніші
3) елізія – вимирання виду, зникнення екологічної ніші
4) інвазія – іміграція виду до екрсистеми – вбудовування ніші
5) субституція – заміщення видів вікаріатів (заміна ніші) за
рахунок конкурентного витіснення
Прунінг – спрощення сукцесії за рахунок випадіння кінцевих
стадій.
30.
Обмін видамиВ результаті непрямої конкуренції американська
норка повністю знищила норку європейську.
31.
Вікаріати(вікаріантні
види) – види, що
займають
подібні
екологічні ніші і
здатні
заміщувати
один - одного
32.
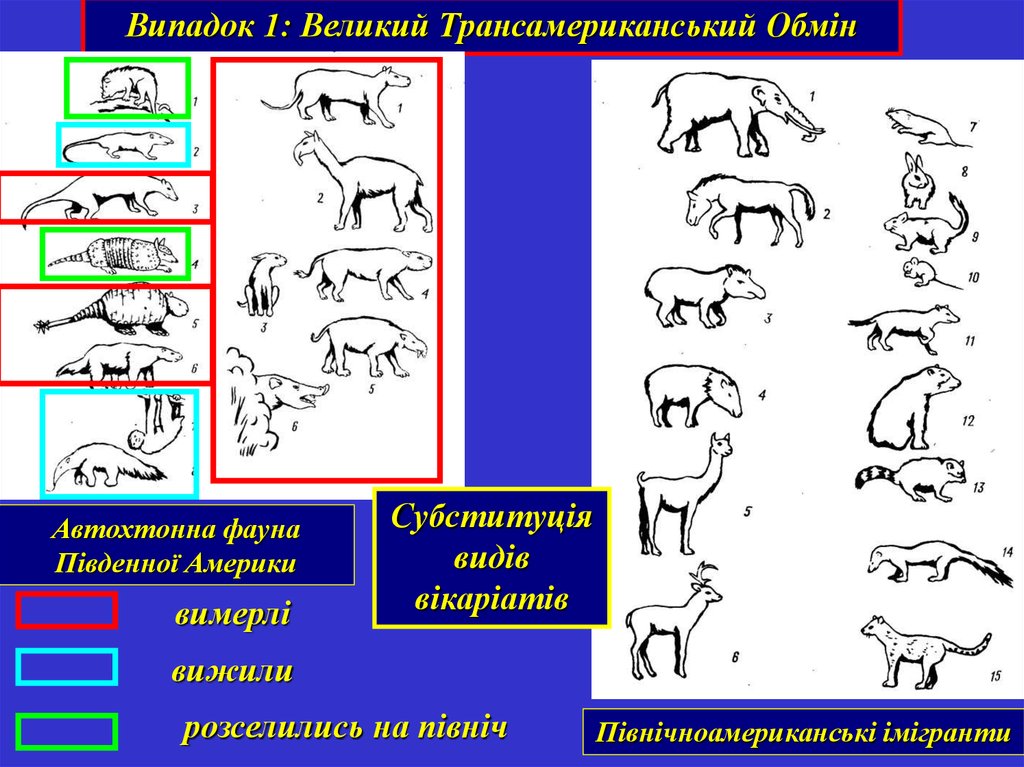
Випадок 1: Великий Трансамериканський ОбмінАвтохтонна фауна
Південної Америки
вимерлі
Субституція
видів
вікаріатів
вижили
розселились на північ
Північноамериканські імігранти
33.
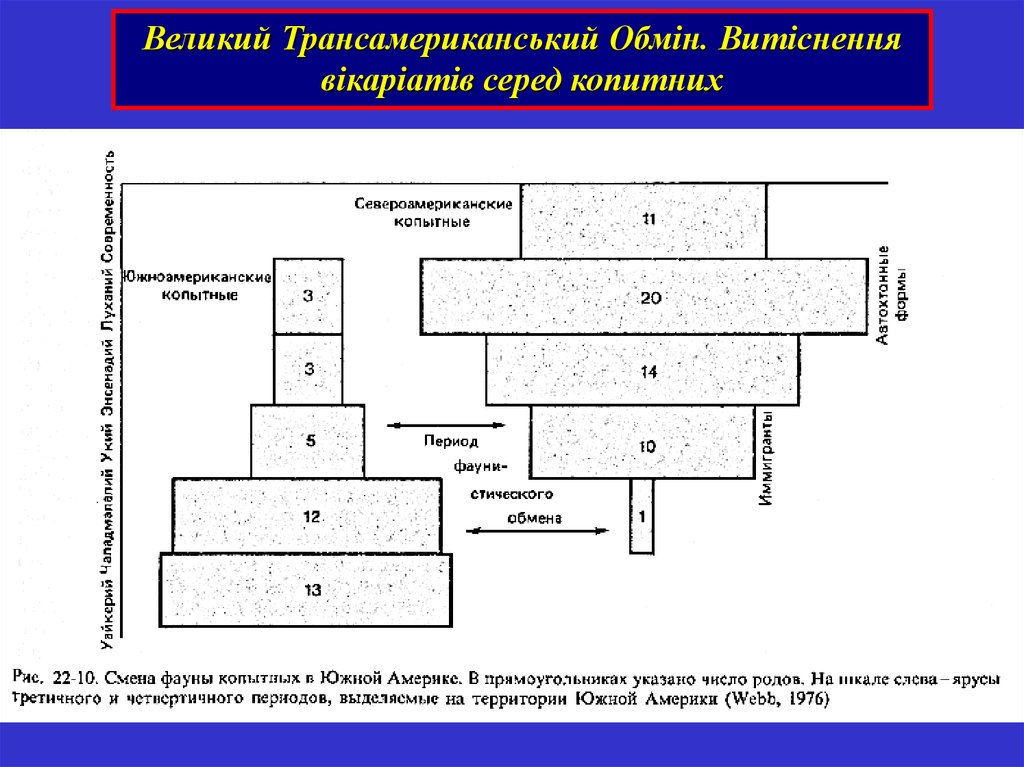
Великий Трансамериканський Обмін. Витісненнявікаріатів серед копитних
34.
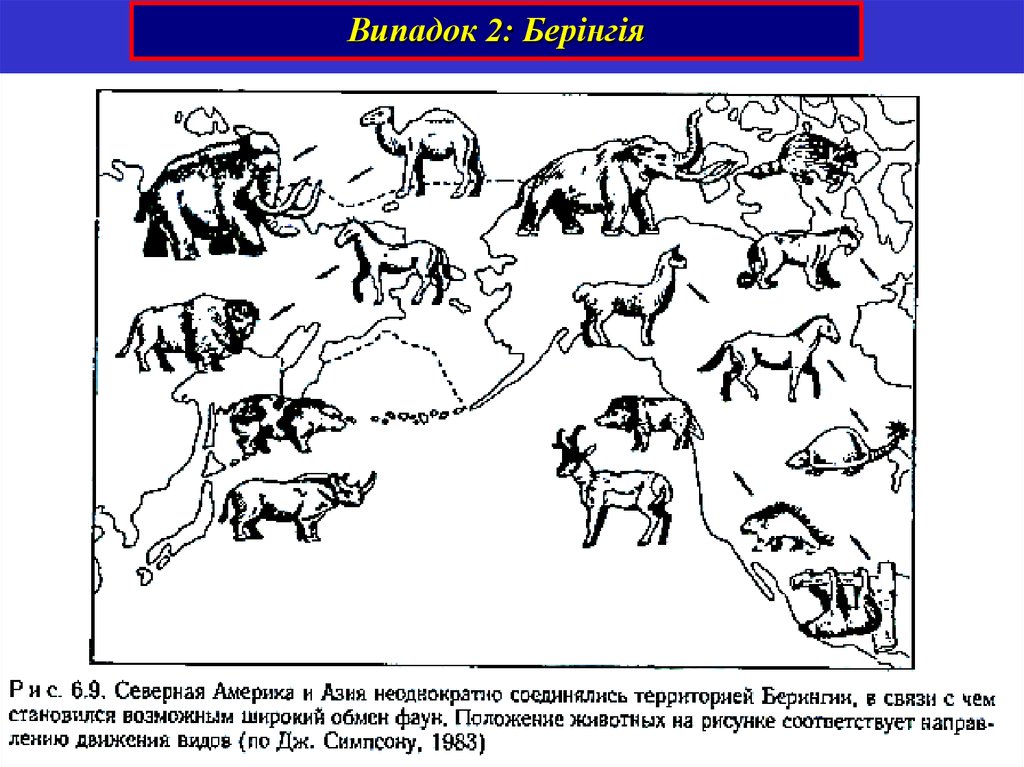
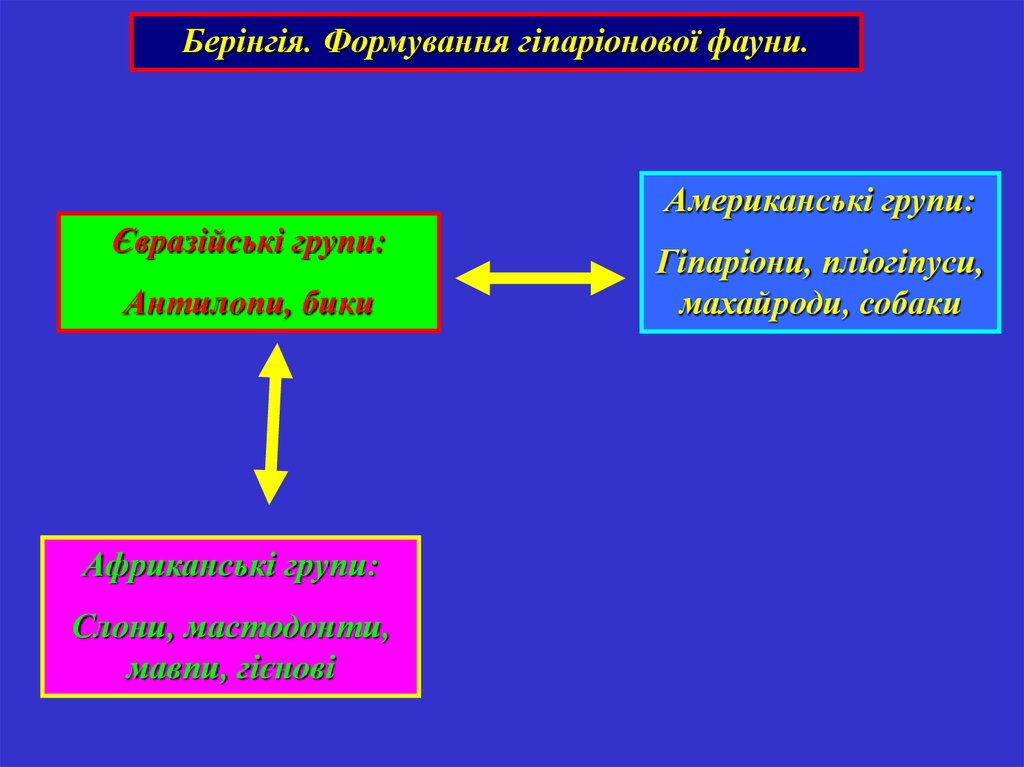
Випадок 2: Берінгія35.
Берінгія. Формування гіпаріонової фауни.Американські групи:
Євразійські групи:
Антилопи, бики
Африканські групи:
Слони, мастодонти,
мавпи, гієнові
Гіпаріони, пліогіпуси,
махайроди, собаки
36.
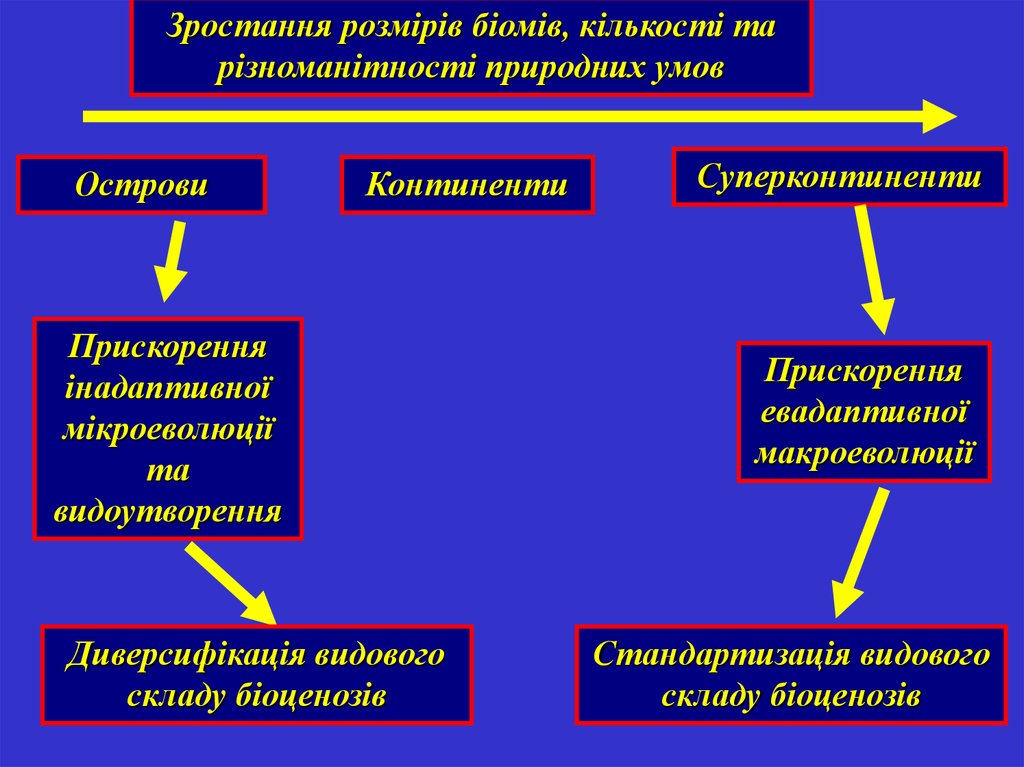
Зростання розмірів біомів, кількості тарізноманітності природних умов
Острови
Континенти
Прискорення
інадаптивної
мікроеволюції
та
видоутворення
Диверсифікація видового
складу біоценозів
Суперконтиненти
Прискорення
евадаптивної
макроеволюції
Стандартизація видового
складу біоценозів
37.
Еволюціяфітохорій у
піздньому полеозої
та мезозої (за
С.В.Мейеном та
В.А.Вахрамєєвим)
38.
39.
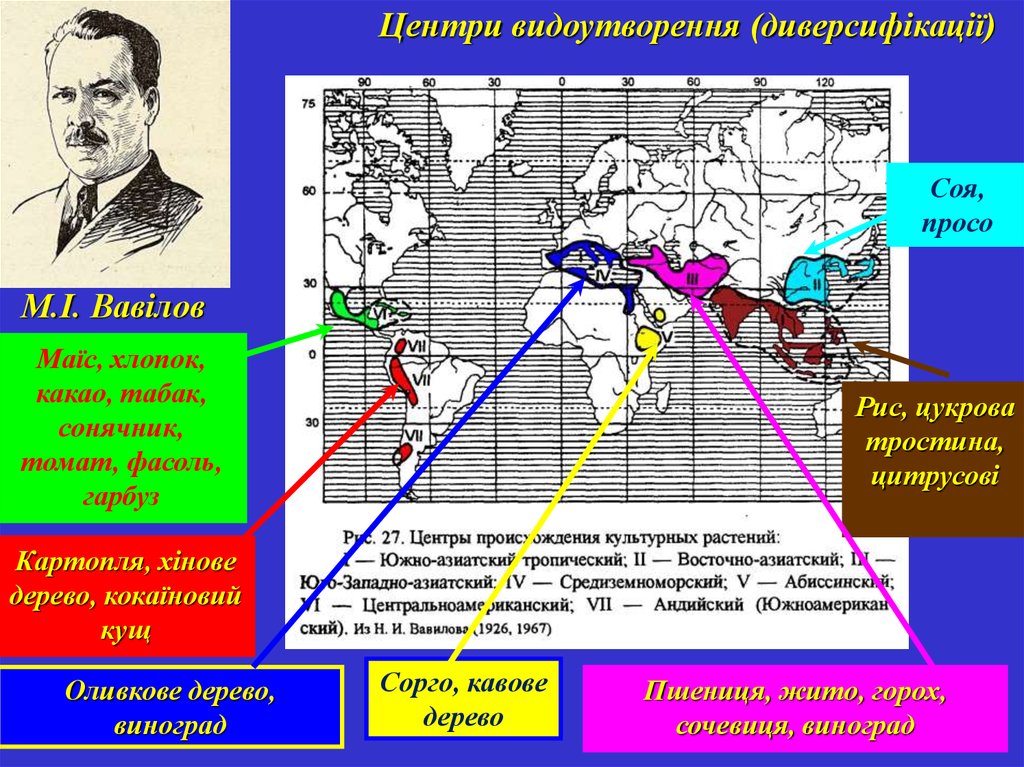
Центри видоутворення (диверсифікації)Соя,
просо
М.І. Вавілов
Маїс, хлопок,
какао, табак,
сонячник,
томат, фасоль,
гарбуз
Рис, цукрова
тростина,
цитрусові
Картопля, хінове
дерево, кокаїновий
кущ
Оливкове дерево,
виноград
Сорго, кавове
дерево
Пшениця, жито, горох,
сочевиця, виноград
40.
Особливості центрів диверсифікіціїЦентр диверсифікації
Периферія
Максимальне різноманіття
умов, сприятливі умови
Багато ендеміків
Більше генералістів
Найбільш молоді
представники таксонів
Більш давні та примітивні
форми
Більше домінантних
алелей
Більше рецесивних алелей
Повний набір
гомологічних рядів
мінливості
41.
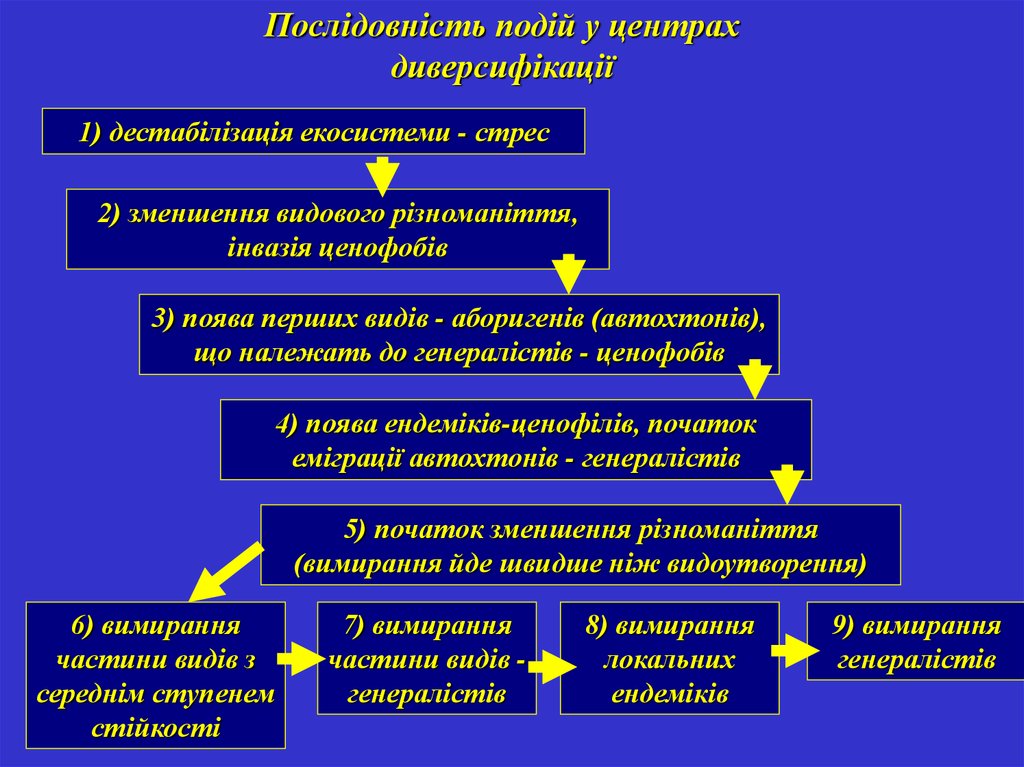
Послідовність подій у центрахдиверсифікації
1) дестабілізація екосистеми - стрес
2) зменшення видового різноманіття,
інвазія ценофобів
3) поява перших видів - аборигенів (автохтонів),
що належать до генералістів - ценофобів
4) поява ендеміків-ценофілів, початок
еміграції автохтонів - генералістів
5) початок зменшення різноманіття
(вимирання йде швидше ніж видоутворення)
6) вимирання
частини видів з
середнім ступенем
стійкості
7) вимирання
частини видів генералістів
8) вимирання
локальних
ендеміків
9) вимирання
генералістів
42.
Моделі фітоспредінга: 1) модель екваторіальної помпиНеобхідна умова: кріооера, наявність суттєвих кліматичних відмінностей
полярна зона
помірна зона
Позатропічне поширення
і персистування реліктів
субтропіки
тропіки
Центри диверсифікації
43.
Моделі фітоспредінга: 2)модель поліградієнтного стануНеобхідна умова: термоера, відсутність суттєвих
кліматичних відмінностей
субтропіки
Приполярні області
44.
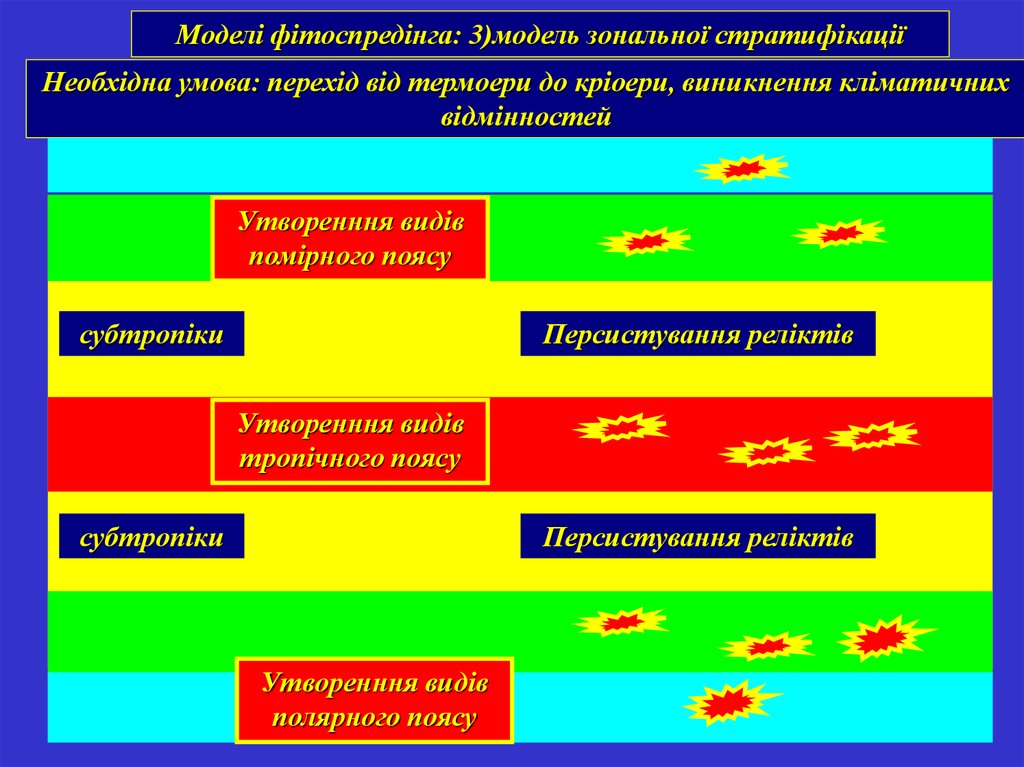
Моделі фітоспредінга: 3)модель зональної стратифікаціїНеобхідна умова: перехід від термоери до кріоери, виникнення кліматичних
відмінностей
Утворенння видів
помірного поясу
субтропіки
Персистування реліктів
Утворенння видів
тропічного поясу
субтропіки
Утворенння видів
Приполярні полярного
області поясу
Персистування реліктів
45.
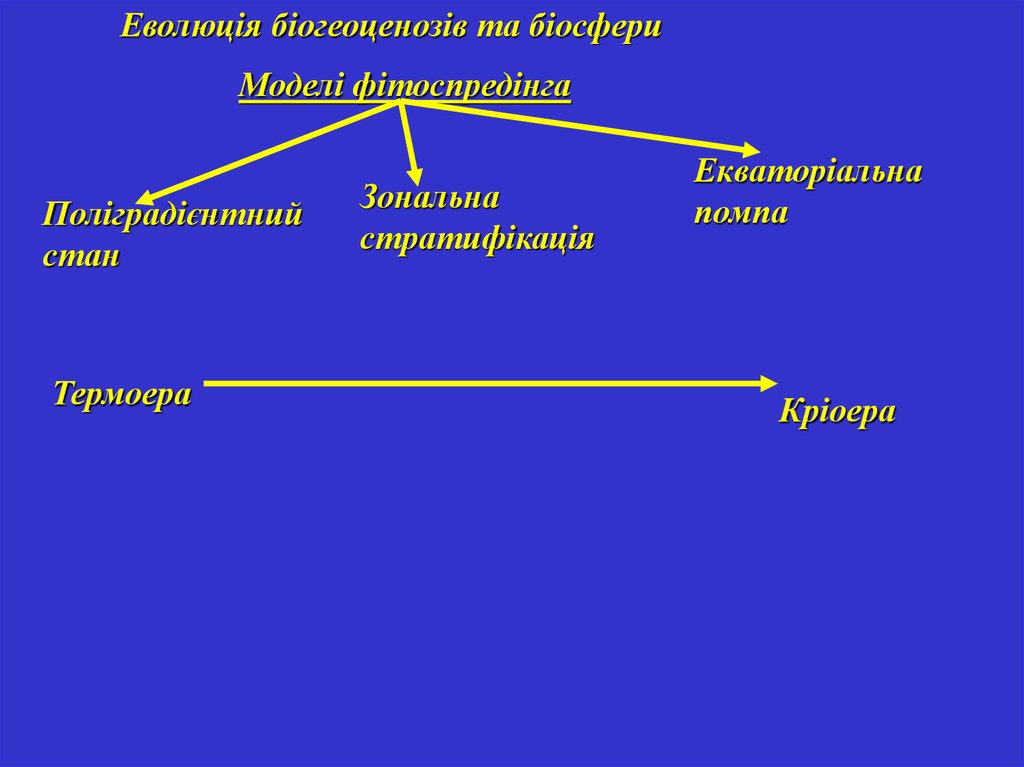
Еволюція біогеоценозів та біосфериМоделі фітоспредінга
Поліградієнтний
стан
Термоера
Зональна
стратифікація
Екваторіальна
помпа
Кріоера
46.
Вимирання у еволюціїФонове
вимирання
Антропогенове
вимирання
Вимирання
видів вікаріатів
Великі
вимирання
(глобальні
біосферні
перебудови)
47.
Роди-одноденки48.
Великі вимиранняКількість видів морських тварин в різні періоди фанерозоя
49.
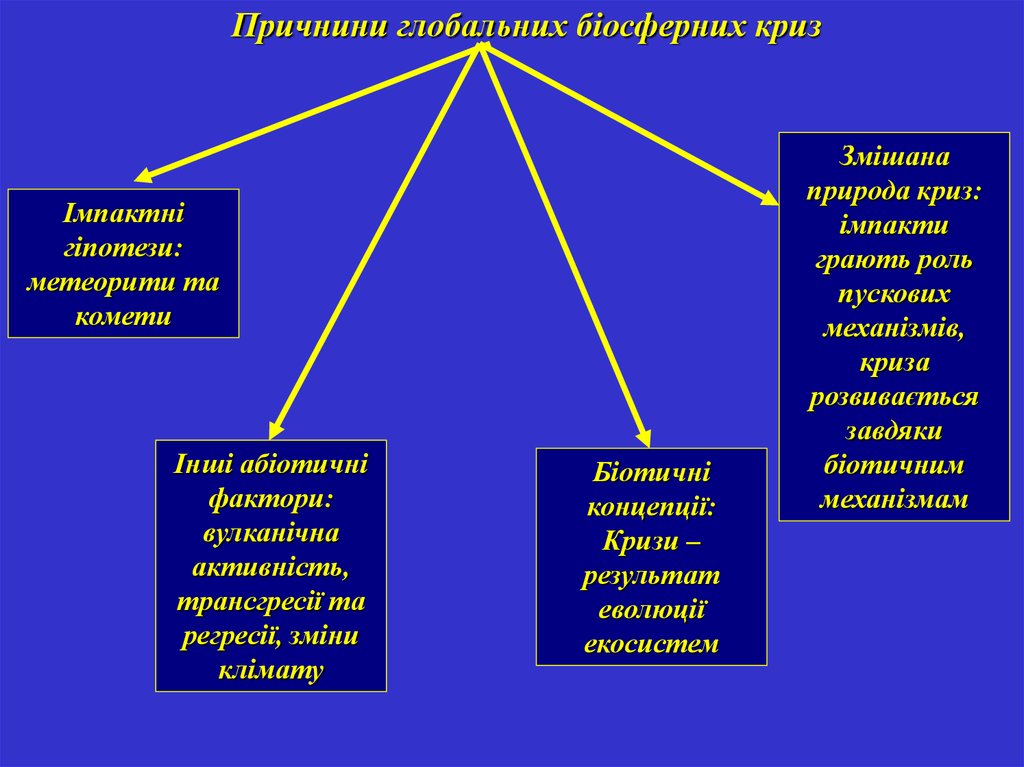
Причнини глобальних біосферних кризІмпактні
гіпотези:
метеорити та
комети
Інші абіотичні
фактори:
вулканічна
активність,
трансгресії та
регресії, зміни
клімату
Біотичні
концепції:
Кризи –
результат
еволюції
екосистем
Змішана
природа криз:
імпакти
грають роль
пускових
механізмів,
криза
розвивається
завдяки
біотичним
механізмам
50.
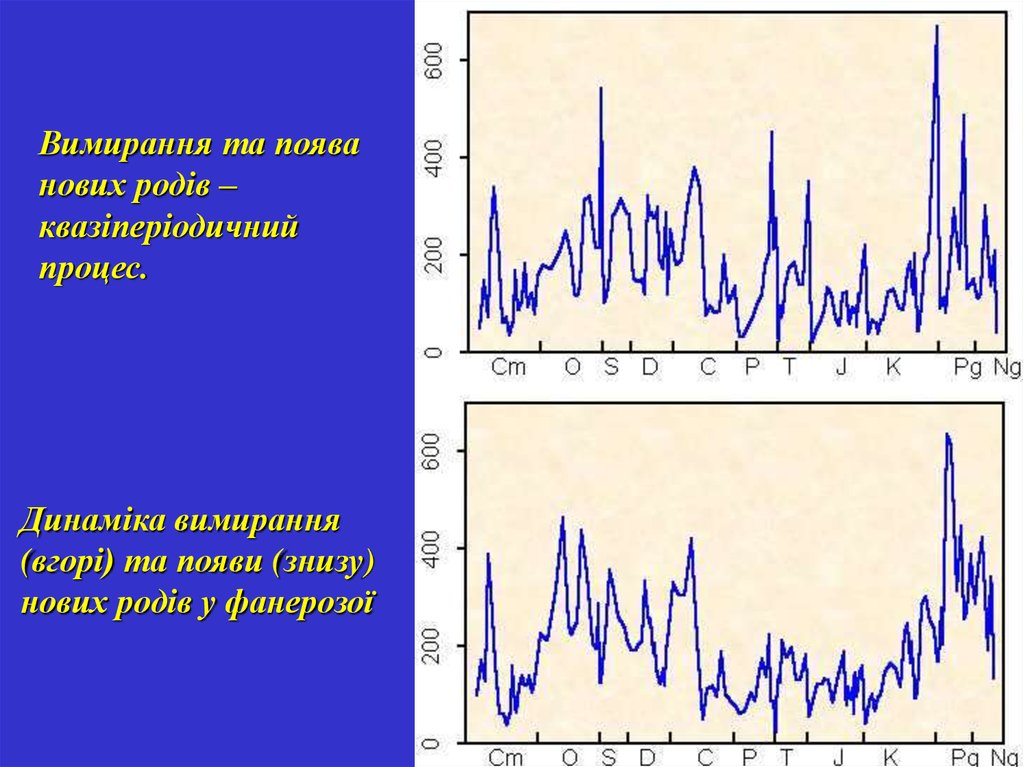
Вимирання та появанових родів –
квазіперіодичний
процес.
Динаміка вимирання
(вгорі) та появи (знизу)
нових родів у фанерозої
51.
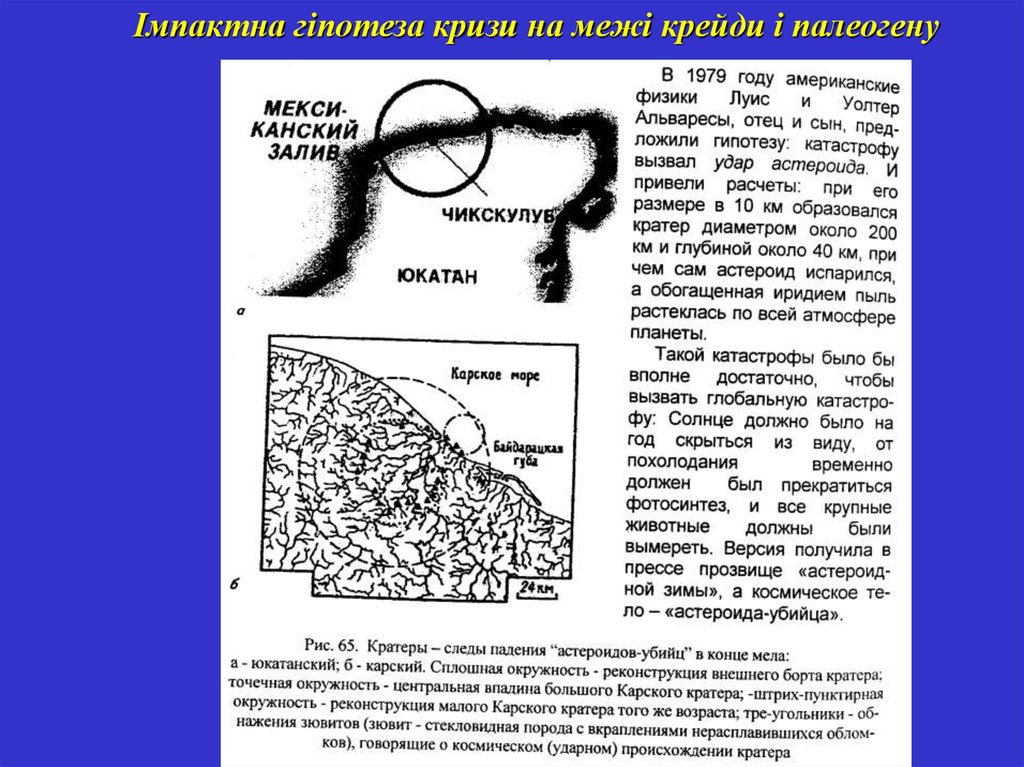
Імпактна гіпотеза кризи на межі крейди і палеогену52.
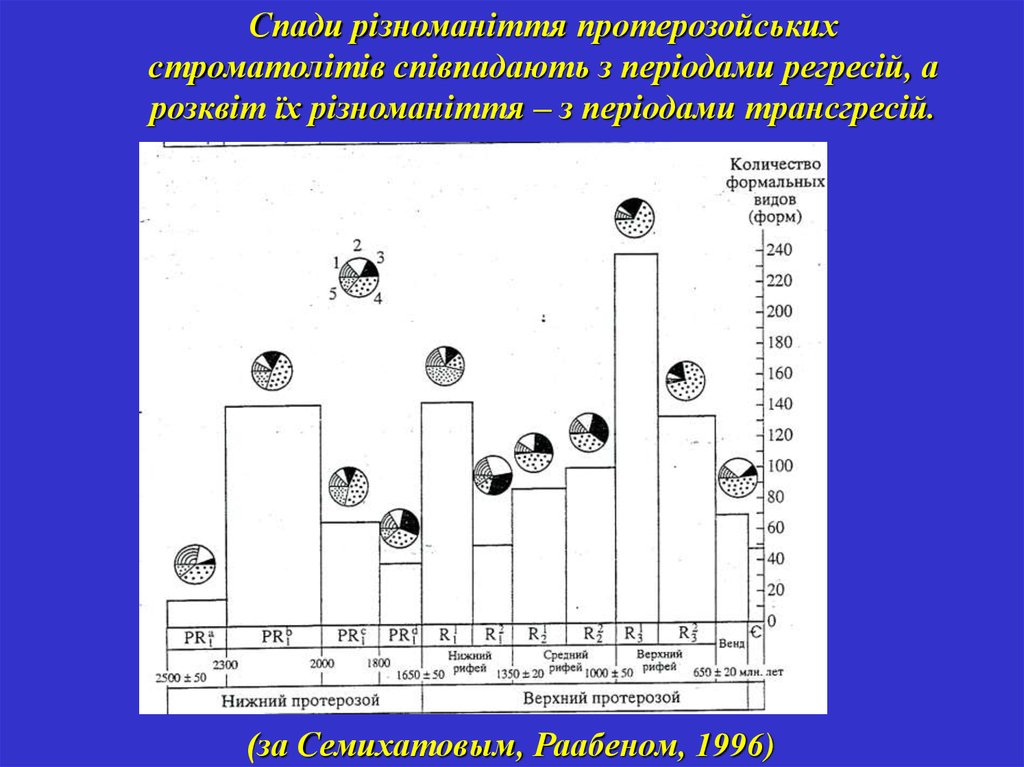
Спади різноманіття протерозойськихстроматолітів співпадають з періодами регресій, а
розквіт їх різноманіття – з періодами трансгресій.
(за Семихатовым, Раабеном, 1996)
53.
Вимирання комах в результаті ангіоспермізації:приклад кризи біотичної природи
54.
Характер вимиранняпланктонних
фораменіфер на
границі крейди та
палеогену
55.
Дякую за увагу!56.
Деякі глобальні закономірності еволюціїбіорізноманіття
57.
Мінливість на рівні біогеоценозуУсереднене таксономічне навантаження у різних екомерах (групах
екологічних ніш), які займають Metazoa (за Киселев, 2004 ).
58.
Закон росту середньої тривалості існування родів у процесіеволюції
Середня тривалість існування родів у млн. років
59.
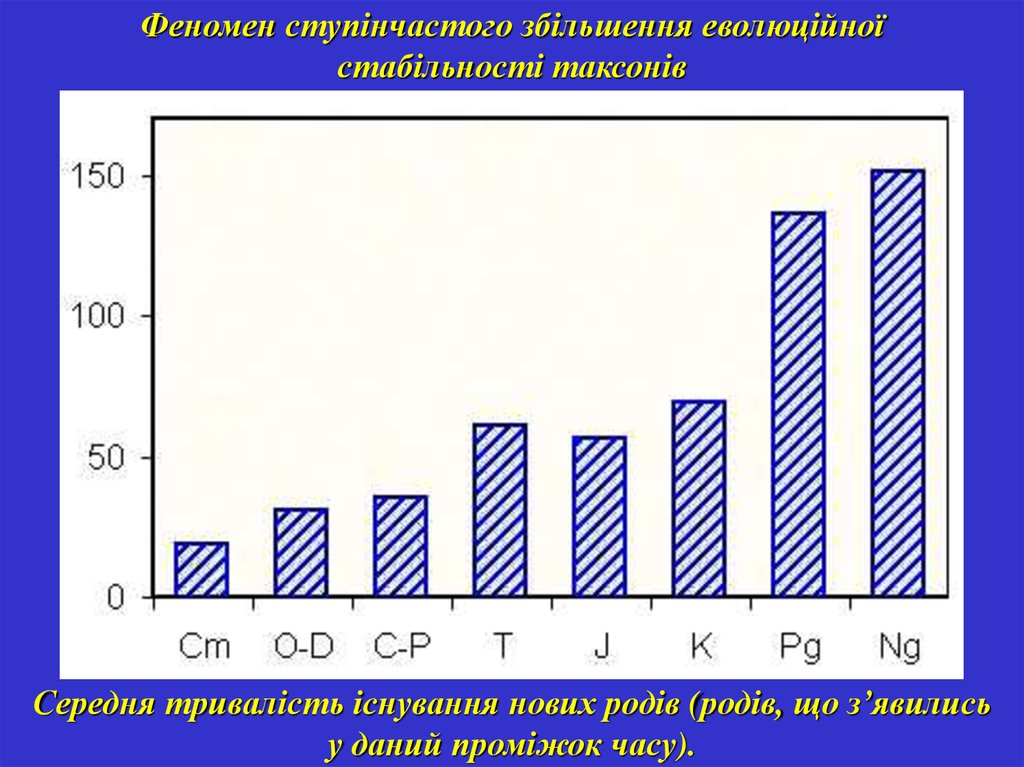
Феномен ступінчастого збільшення еволюційноїстабільності таксонів
Середня тривалість існування нових родів (родів, що з’явились
у даний проміжок часу).
60.
Розподіл родів з різноюкількістю видів на прикладі
родини Curculionidae (за
Е.Майром).
А) Співвідношення числа таксонів різних рангів фанерозойської
морської біоти (g-роди, f-родини, o- ряди, c-класи, p-типи);
Б) Зміна кількості середнього числа субтаксонів у таксоні (за
Марков, 2003).
61.
Крива сумарої тривалості існування62.
Найбільш звичайна форма алометричної залежності морфофункціонального параметра від маси тіла для старшого таксоната субтаксонів. Характерно, що в межах заданого інтервалу
значень маси морфо-функцірнальний параметр має більшу
мінливість у видів старшого таксону (dPt > dPg ), а в межах
одного діапазону значень параметру маса більш мінлива в межах
субтаксону (dMt < dMg ) (за Гродницкий, 2002)
63.
Дякую за увагуDinohyus - хижий предок сучасної свині
64.
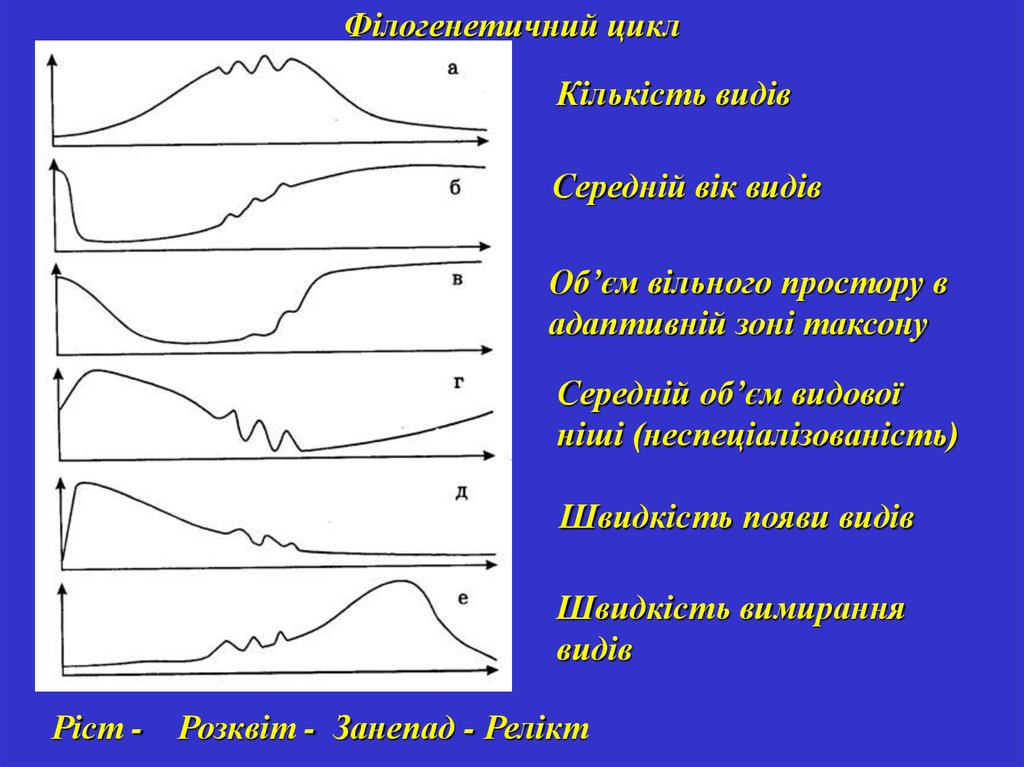
Філогенетичний циклКількість видів
Середній вік видів
Об’єм вільного простору в
адаптивній зоні таксону
Середній об’єм видової
ніші (неспеціалізованість)
Швидкість появи видів
Швидкість вимирання
видів
Ріст - Розквіт - Занепад - Релікт
65.
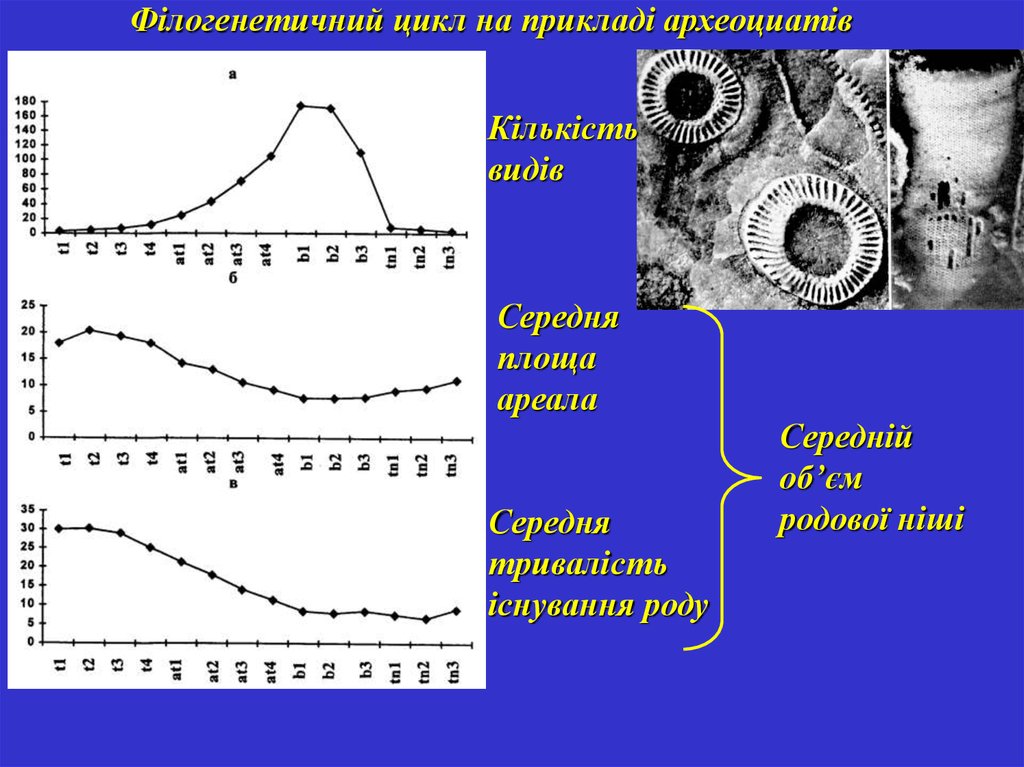
Філогенетичний цикл на прикладі археоциатівКількість
видів
Середня
площа
ареала
Середня
тривалість
існування роду
Середній
об’єм
родової ніші
66.
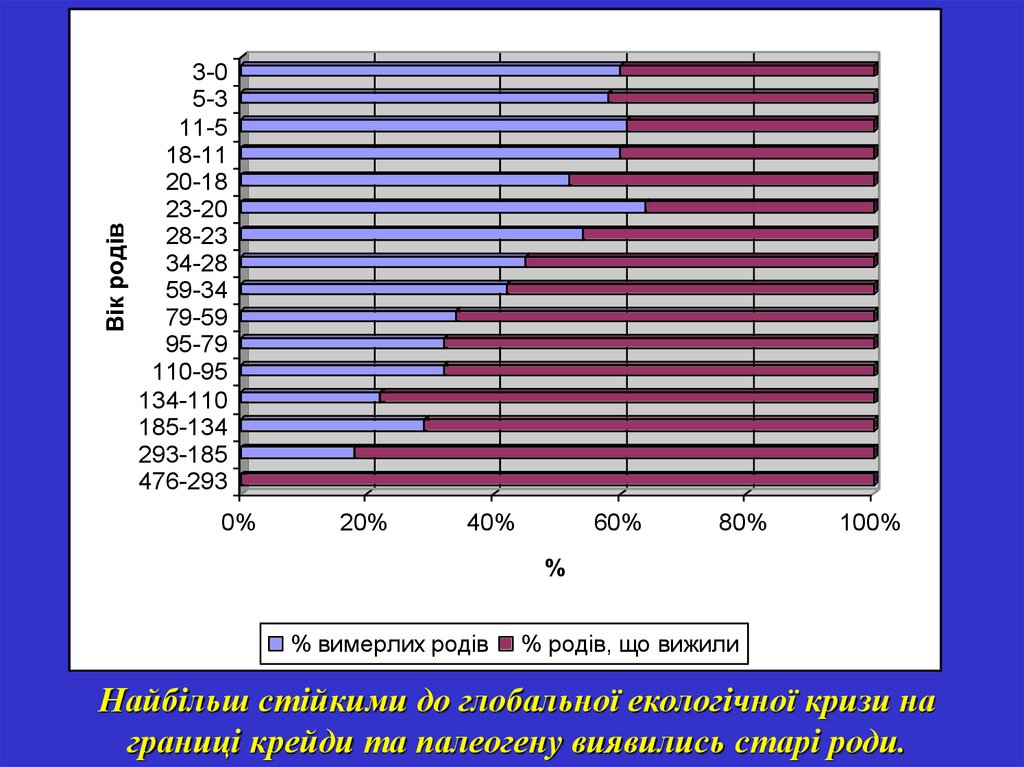
Вік родів3-0
5-3
11-5
18-11
20-18
23-20
28-23
34-28
59-34
79-59
95-79
110-95
134-110
185-134
293-185
476-293
0%
20%
40%
60%
80%
100%
%
% вимерлих родів
% родів, що вижили
Найбільш стійкими до глобальної екологічної кризи на
границі крейди та палеогену виявились старі роди.
67.
Темпи еволюційних перетворень1 Дарвін : зміна розмірів органа на 0,001 його величини
за 1000 років
1 Сімпсон : зміна розмірів органа на 1 стандартне
(квадратичне) відхилення за 1 млн. років
1 Холдейн : зміна розмірів органа на 1 стандартне
(квадратичне) відхилення за 1 покоління
Час напіввимирання таксона : час за який вимирає 1/2
частина видів таксона
Вік напівсучасної фауни (фауни, в якій 1/2 видів
сучасна, а друга половина вимерла).
68.
Вік напівсучасної фауни родів деяких таксонівСсавці - 4 млн р.
Птахі - 10 млн. р.
Амфібії - 10 млн. р.
Рептилії - 20 млн. р.
Риби - 30-50 млн. р.
Комахи - 40 млн. р.
Фораменіфери - 230 млн. р.
69.
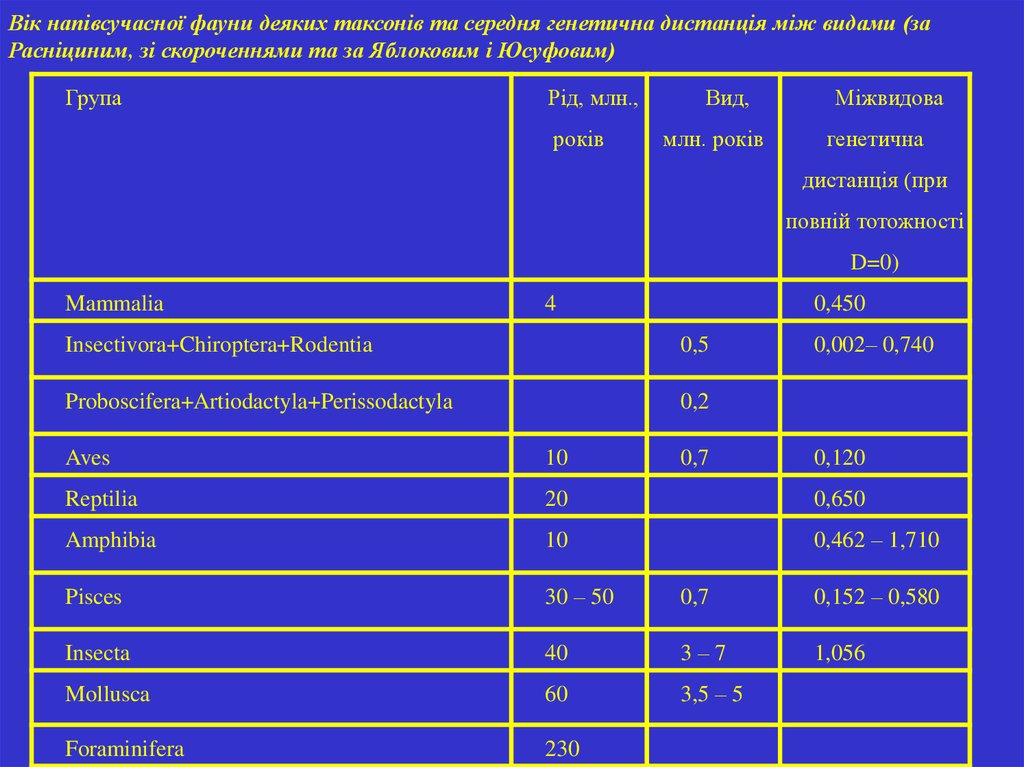
Вік напівсучасної фауни деяких таксонів та середня генетична дистанція між видами (заРасніциним, зі скороченнями та за Яблоковим і Юсуфовим)
Група
Рід, млн.,
років
Вид,
млн. років
Міжвидова
генетична
дистанція (при
повній тотожності
D=0)
Mammalia
4
0,450
Insectivora+Chiroptera+Rodentia
0,5
Proboscifera+Artiodactyla+Perissodactyla
0,2
0,7
0,002– 0,740
Aves
10
0,120
Reptilia
20
0,650
Amphibia
10
0,462 – 1,710
Pisces
30 – 50
0,7
0,152 – 0,580
Insecta
40
3–7
1,056
Mollusca
60
3,5 – 5
Foraminifera
230
70.
Живі викопні роди, які існуютьна Землі десяткисотні міліонів років
71.
72.
Типи живих викопнихВиди – ізоляти (мешканці
рефугіумів)
Види – мешканці
реліктових біогеоценозів
Види, що займають
підпорядковане
положення у
біогеоценозах
Гаттерія
Деревовидна
папороть
Fannia
canicularis
73.
Популяції видуDelamarentulus tristani
Silv. з групи Protura
мешкають у КостаРіці та у Західній
Африці. Ізоляція
популяцій цього виду
відбулася близько
80млн. років тому.
Еволюційному стазису сприяє стабільність умов,
стабільність екологічної ніші, яка більша у видів, що
займають підпорядковане положення у біоценозах.
Види, що займають домінуючі позиції у біоценозах (віоленти)
навпаки виявляються найбільш чутливими до перебудови
екосистем завдяки прунінгу – випадіння останнях стадій
сукцесії.
74.
Фактори, що сприяють швидкій еволюції груп:Статеве розмноження
Можливість дії статевого добору
Складність будови
Великі розміри
Довгий онтогенез
Мала кількість нащадків
Турбота про нащадків
Великий діапазон внутрішньовидової мінливості ознак
Мінімальний діапазон внутрішньовидової генетичної мінливості
Складність міжгенних взаємодій
Панівні позиції в біогеоценозі
Різноманіття умов середовища
Максимально комфортні абіотичні умови
Кондиціонування зовнішнього середовища
Великий ареал
Складність екосистем
75.
Інадаптації та евадаптаціїІнадаптація - неглибоке, однобічне пристосування, яке
здійснюється за рахунок порушення координаційних зв’язків у
організмі.
76.
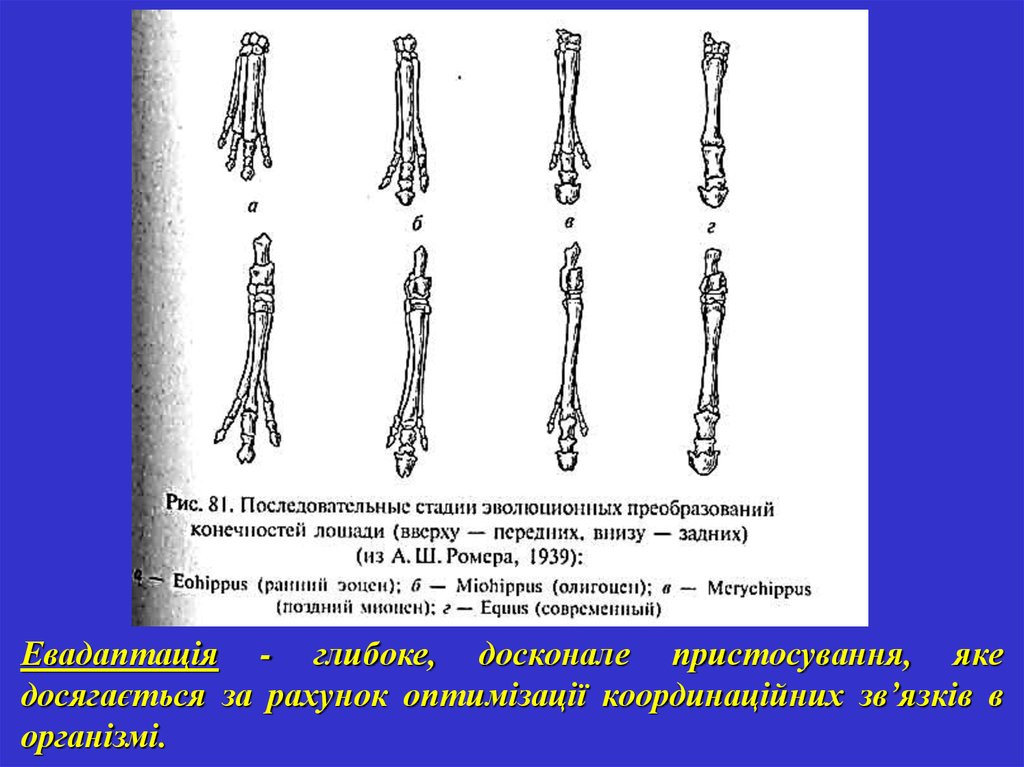
Евадаптація - глибоке, досконале пристосування, якедосягається за рахунок оптимізації координаційних зв’язків в
організмі.
77.
Група креодонтівдосягла швидкого
успіху, проте згодом
поступилася
справжнім хижим.
Оксієна
78.
Адаптивний компроміс...а у пінгвіна спеціалізована
для плавання.
Форма тіла тіла
альбатроса
оптимальна для
польоту...
На відміну від
спеціалізованих
видів, генералісти,
такі як кайри,
мають проміжний
фенотип, компромісний для пересування у двох різних
типах середовища