































 Биология
БиологияПохожие презентации:
")
")






")

Трансляция. Переход от нуклеиновых кислот к белкам
1.
ТрансляцияПереход от нуклеиновых кислот к
белкам
Генетическая информация, находящаяся в клетке в виде
ДНК и воспроизводящаяся в клеточных поколениях путем
редупликации ДНК, реализуется через биосинтез белка.
2.
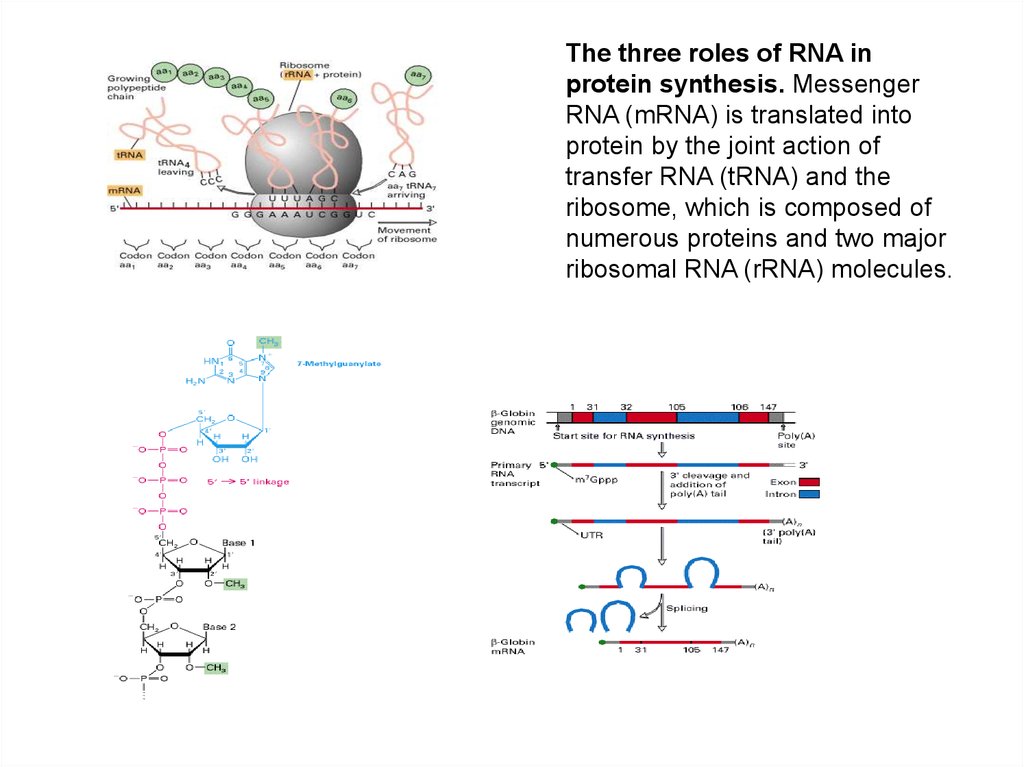
The three roles of RNA inprotein synthesis. Messenger
RNA (mRNA) is translated into
protein by the joint action of
transfer RNA (tRNA) and the
ribosome, which is composed of
numerous proteins and two major
ribosomal RNA (rRNA) molecules.
3.
4.
Предтрансляционный этапГЕНЕТИЧЕСКИЙ КОД КАК
СИСТЕМА
5.
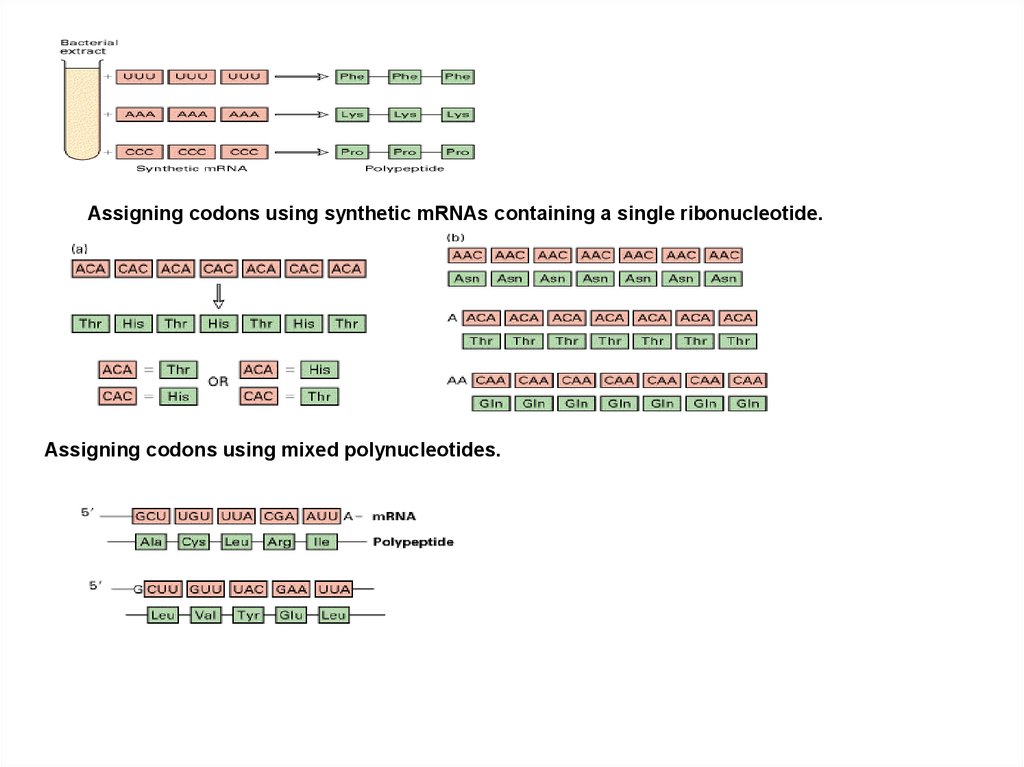
Assigning codons using synthetic mRNAs containing a single ribonucleotide.Assigning codons using mixed polynucleotides.
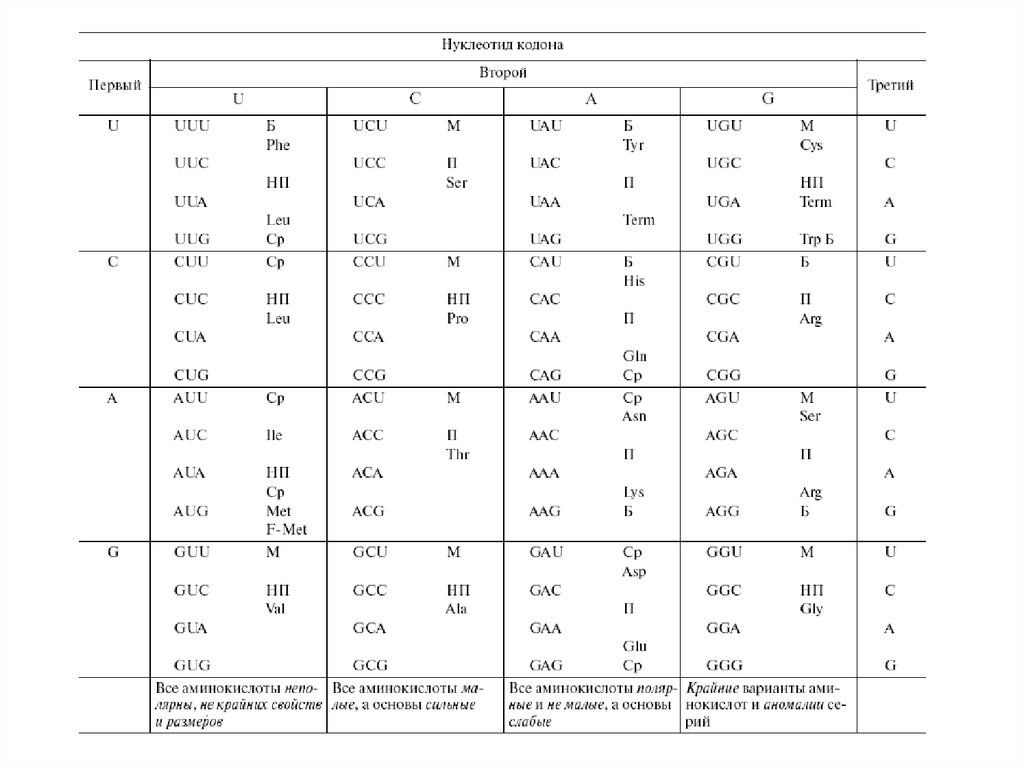
6. Свойства генетического кода:
1. Все кодоны однозначны, то есть каждый кодирует единственнуюаминокислоту. Иначе говоря, в направлении кодон => аминокислота
генетический код однозначен.
2. Обратное соответствие в направлении аминокислота => кодон
неоднозначно, и это свойство называется вырожденностью. Отдельные
аминокислоты кодируются группами (сериями) кодонов-синонимов. 18
серий из 20 содержат от двух до шести кодонов, две серии (Меt и Тrр) не
вырождены, содержат по одному кодону. Средняя вырожденность
генетического кода приблизительно три кодона на серию.
7. Свойства генетического кода:
Генетический код содержит также знакипунктуации (начала и конца) трансляции.
3.
–
–
–
–
Кодоны AUG, UUG и GUG у прокариот помимо
кодирования аминокислот кодируют также инициацию
трансляции.
Однако однозначность кодирования при этом не
нарушается, так как инициирующие знаки
локализованы в определенном окружении (контексте),
способном образовывать самокомплементарные
"шпильки".
Три "вакантных" триплета— UAA (ochre), UAG (amber)
и UGA (ораl) — не кодируют аминокислот, а
выполняют роль терминальных знаков трансляции
(стоп-кодонов, нонсенс-кодонов или терминальных
нонсенсов).
В норме ими заканчиваются все цистроны, то есть
транслируемые гены, единицы трансляции.
Мутационное возникновение нонсенсов внутри гена
приводит к преждевременной терминации трансляции
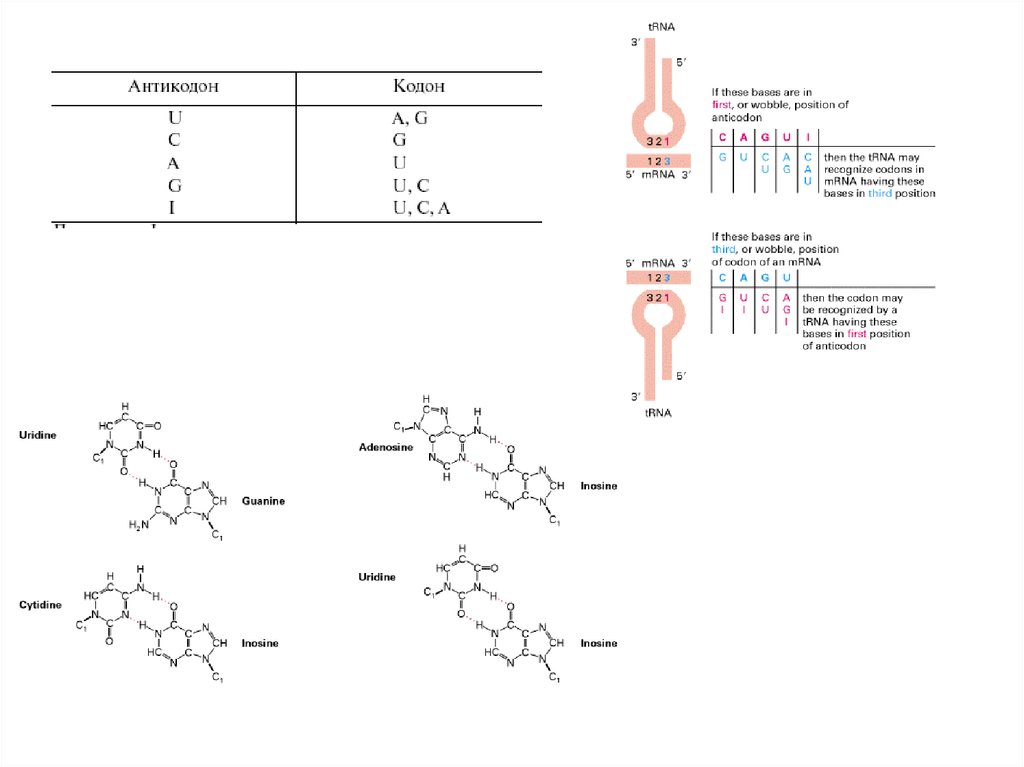
8. Неоднозначное спаривание:
Однозначность кода в направлении кодон => аминокислота
обеспечивается строгой специфичностью АА-тРНК-синтетаз.
Каждая АА-тРНК-синтетаз узнает единственную аминокислоту,
поэтому неоднозначность исключена или маловероятна.
В основе систематической вырожденности лежат правила
неоднозначности спаривания кодон— антикодон, установленные
Ф. Криком. Один антикодон может узнавать один, два или три
кодона, различающиеся по третьей позиции.
Согласно правилам неоднозначного спаривания, систематическая
вырожденность в парах кодонов обеспечивается отдельными
фракциями т-РНК, имеющими U, G или I (инозин) в трех позициях
антикодонов.
Всего генетический код требует не менее 32 фракций т-РНК.
9.
10.
11. Система генетического кода:
• Анализ таблицы генетического кода как целого позволяетвыявить удивительные свойства его регулярности и симметрии.
• Обозначим позиции нуклеотидов в кодонах (5'—1—2—3—3')
через х -- у -- z, где х - приставка, у - корень, z - окончание, а ху
— основа кодона.
• Эти термины отражают сходные понятия лингвистики.
– Корни слов определяют их смысл. Все мутации, затрагивающие
корень (у) кодона, также изменяют кодовую серию, то есть
нарушают смысл кодона.
– Приставки слов тоже участвуют в определении смысла, хотя и не
так жестко, а многие их изменения меняют смысл слов. Замены в
приставках (х) кодонов чаще всего изменяют их смысл, но иногда
являются синонимическими.
– Окончания слов обычно участвуют в словоизменении, то есть в
синонимических преобразованиях. Аналогично 70% замен в
окончаниях (z) кодонов синонимические.
– Наконец, приставка и корень слова образуют его основу, несущую
полную или доминирующую смысловую нагрузку. Основа кодона
(ху) тоже играет ключевую роль в особенностях генетического кода.
12. Групповые свойства кода:
Легко заметить, что столбцы генетического кода отвечающие
определенным корням кодонов, имект некоторые групповые свойства.
Третий столбец (корень А) содержит полярные аминокислоты немалые
по размеру, а все основы кодонов слабые.
Второй столбец (корень С) содержит аминокислоты, малые по
размеру, а все основы кодонов сильные.
Первый столбец (корень U) включает неполярные аминокислоты
разных (но не крайних) размеров, а свойства кодонов неоднозначны.
Четвертый столбец (корень G) содержит все крайние и аномальные
варианты аминокислот и кодовых серий: самую реактивную
аминокислоту (Суs), самую большую и плоскую (Тrр), самую большую и
корявую (Агg), самую маленькую (Glу), часть единственной несвязной
серии (Sег) и неоднозначный терминальный нонсенс (UAG), который в
ряде случаев кодирует 21-ю аминокислоту - селеноцистеин (Sес).
Таким образом, если групповые свойства корней (и столбцов)
отвечают каким-то общим правилам их возникновения, то четвертый
столбец скорее напоминает свалку всего, что не попало в первые три
столбца по групповым правилам их формирования.
13.
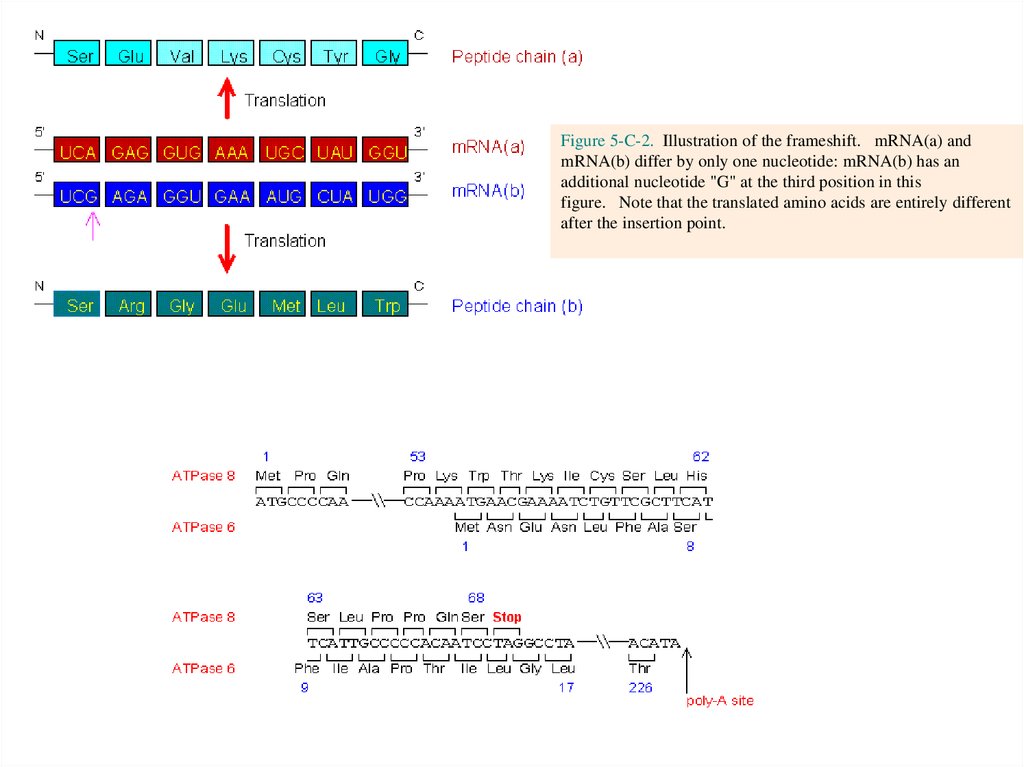
Figure 5-C-2. Illustration of the frameshift. mRNA(a) andmRNA(b) differ by only one nucleotide: mRNA(b) has an
additional nucleotide "G" at the third position in this
figure. Note that the translated amino acids are entirely different
after the insertion point.
14. Предтрансляционный этап
Строение и функциитРНК
15.
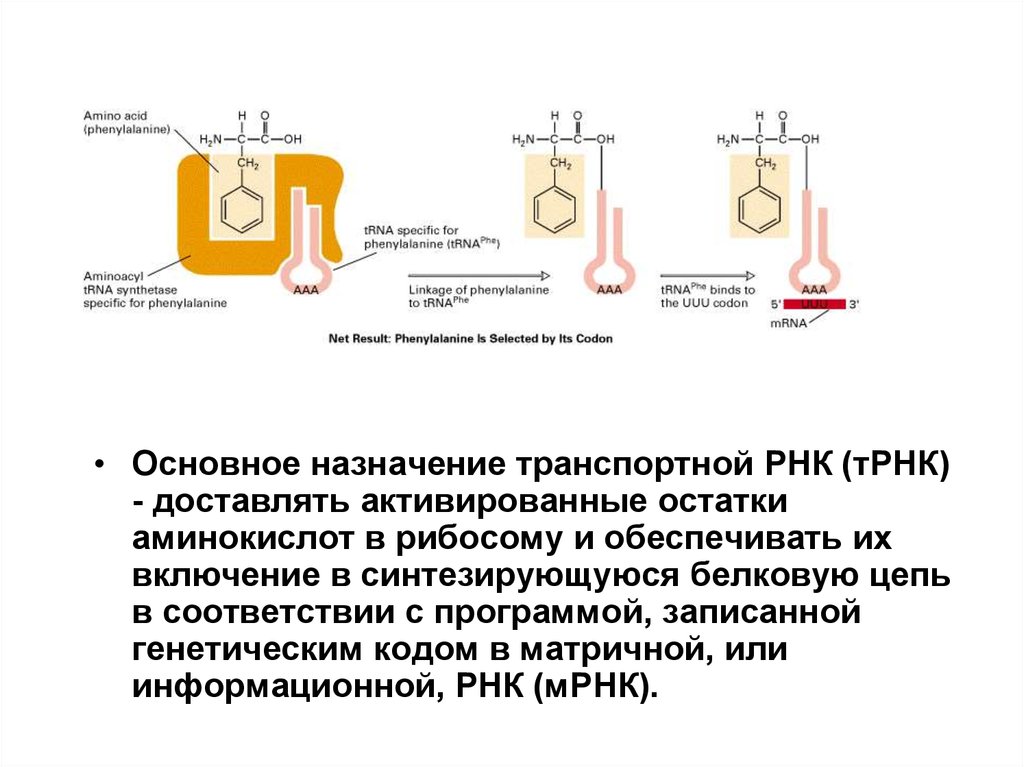
• Основное назначение транспортной РНК (тРНК)- доставлять активированные остатки
аминокислот в рибосому и обеспечивать их
включение в синтезирующуюся белковую цепь
в соответствии с программой, записанной
генетическим кодом в матричной, или
информационной, РНК (мРНК).
16. Общие сведения:
• Впервые нуклеотидная последовательность молекулытРНК — дрожжевой аланиновой тРНК, была расшифрована
в 1965 году в лаборатории Р. Холли.
• С тех пор были опубликованы данные о нуклеотидной
последовательности (называемой первичной структурой)
более чем 1700 видов различных тРНК из
прокариотических и эукариотических организмов.
• Все тРНК имеют общие черты как в их первичной
структуре, так и в способе складывания полинуклеотидной
цепи во вторичную структуру за счет взаимодействий
между основаниями нуклеотидных остатков.
• Помимо цитоплазматических тРНК, кодируемых ядерной
ДНК, в эукариотических клетках имеются особые виды
тРНК, кодируемые автономными геномами митохондрий и
хлоропластов.
17. Первичная структура тРНК:
• тРНК - относительно небольшие молекулы, длина их цепейварьирует от 74 до 95 нуклеотидных остатков.
• Все тРНК имеют одинаковый З'-конец, построенный из двух
остатков цитозина и одного — аденозина (ССА-конец).
• Именно З'-концевой аденозин связывается с
аминокислотным остатком при образовании аминоацилтРНК.
• ССА-конец присоединяется ко многим тРНК с помощью
специального фермента. В других случаях он считывается с
кодирующего данную тРНК гена.
• Нуклеотидный триплет, комплементарный кодону для
аминокислоты (антикодон), находится приблизительно в
середине цепи тРНК.
• В отдельных положениях последовательности практически у
всех видов тРНК встречаются одни и те же (консервативные)
нуклеотидные остатки. В некоторых положениях могут
находиться или только пуриновые, или только
пиримидиновые основания (их называют
полуконсервативными остатками).
18.
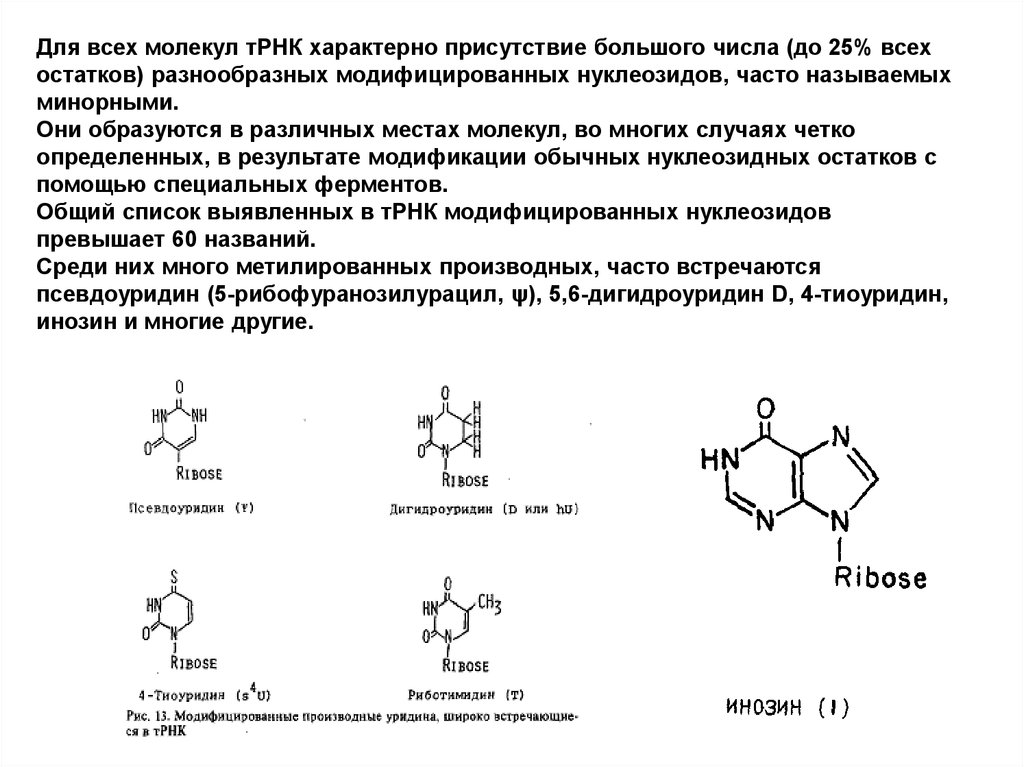
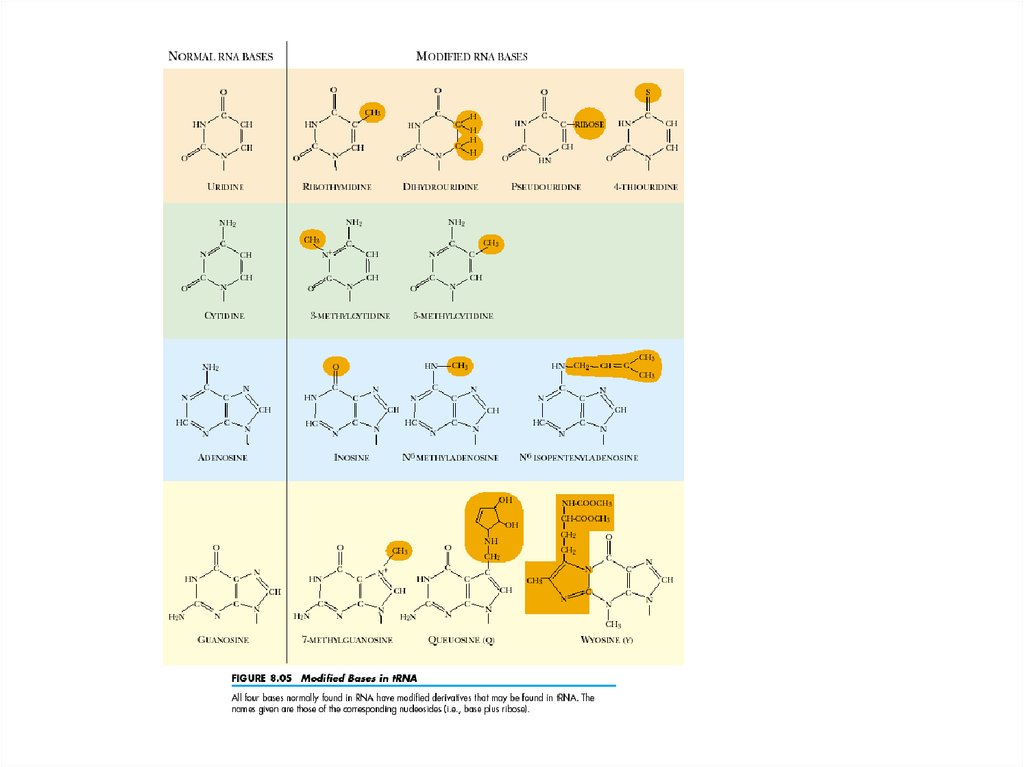
Для всех молекул тРНК характерно присутствие большого числа (до 25% всехостатков) разнообразных модифицированных нуклеозидов, часто называемых
минорными.
Они образуются в различных местах молекул, во многих случаях четко
определенных, в результате модификации обычных нуклеозидных остатков с
помощью специальных ферментов.
Общий список выявленных в тРНК модифицированных нуклеозидов
превышает 60 названий.
Среди них много метилированных производных, часто встречаются
псевдоуридин (5-рибофуранозилурацил, ψ), 5,6-дигидроуридин D, 4-тиоуридин,
инозин и многие другие.
19.
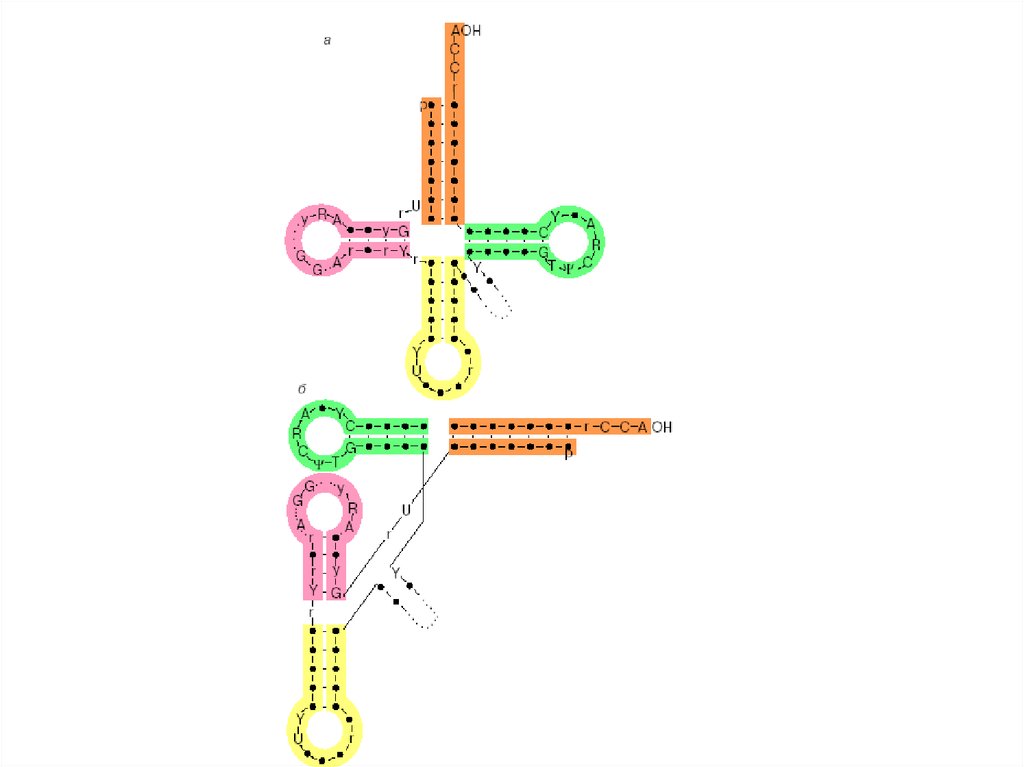
20. Вторичная структура тРНК
Анализ нуклеотидной последовательности уже
первой расшифрованной дрожжевой аланиновой
тРНК выявил возможность складывания цепи во
вторичную структуру за счет
взаимокомплементарности участков цепи.
Три фрагмента цепи оказываются комплементарными
при складывании их на себя, образуя
шпилькообразные структуры. Кроме того, 5'-конец
комплементарен участку, близкому к З'-концу цепи,
при их антипараллельном расположении; они
формируют так называемый акцепторный стебель.
В результате образуется структура,
характеризующаяся наличием четырех стеблей и
трех петель, которая получила название "клеверного
листа".
Стебель с петлей формируют ветвь.
Слева и справа от антикодоновой ветви расположены
D- и Т-ветви, соответственно названные так из-за
присутствия в их петлях необычных консервативных
нуклеозидов дигидроуридина (D) и тимидина (Т).
Нуклеотидные последовательности всех изученных
тРНК могут быть сложены в аналогичные структуры.
В дополнение к трем петлям клеверного листа в
структуре тРН К выделяют также дополнительную,
или вариабельную, петлю (V-петлю).
Ее размеры резко различаются у разных тРНК,
варьируя от 4 до 21 нуклеотида.
21. Третичная структура тРНК:
Общие принципы складывания цепей различных
тРНК в компактную третичную структуру оказались
универсальными.
За счет взаимодействия элементов вторичной
структуры формируется третичная структура,
которая получила название L-формы из-за сходства
с латинской буквой L.
За счет стэкинга оснований акцепторный стебель и
Т-стебель клеверного листа образуют одну
непрерывную двойную спираль, а два других стебля
— антикодоновый и D — другую непрерывную
двойную спираль.
При этом D- и Т-петли оказываются сближенными и
скрепляются между собой путем образования
дополнительных, часто необычных пар оснований.
В образовании этих пар, как правило, принимают
участие консервативные или полуконсервативные
остатки.
ССА-конец тРНК и ее антикодоновый триплет
находятся на максимальном удалении один от
другого (расстояние около 8 нм), причем основания
антикодона обращены внутрь угла L-образной
молекулы.
22.
23. Аминоацил-тРНК-синтетазы:
• Как уже указывалось, для каждой из 20аминокислот в клетках есть специальный
фермент, осуществляющий синтез
соответствующей аминоацил-тРНК (общее
название — аминоацил-тРНК-синтетазы, для
краткости будем называть их синтетазами).
• Все ферменты этой группы катализируют
реакции аминоацилирования тРНК по
принципиально сходному механизму.
• Реакция сопряжена с расщеплением аденозин5'-трифосфата (АТР) и протекает в две стадии.
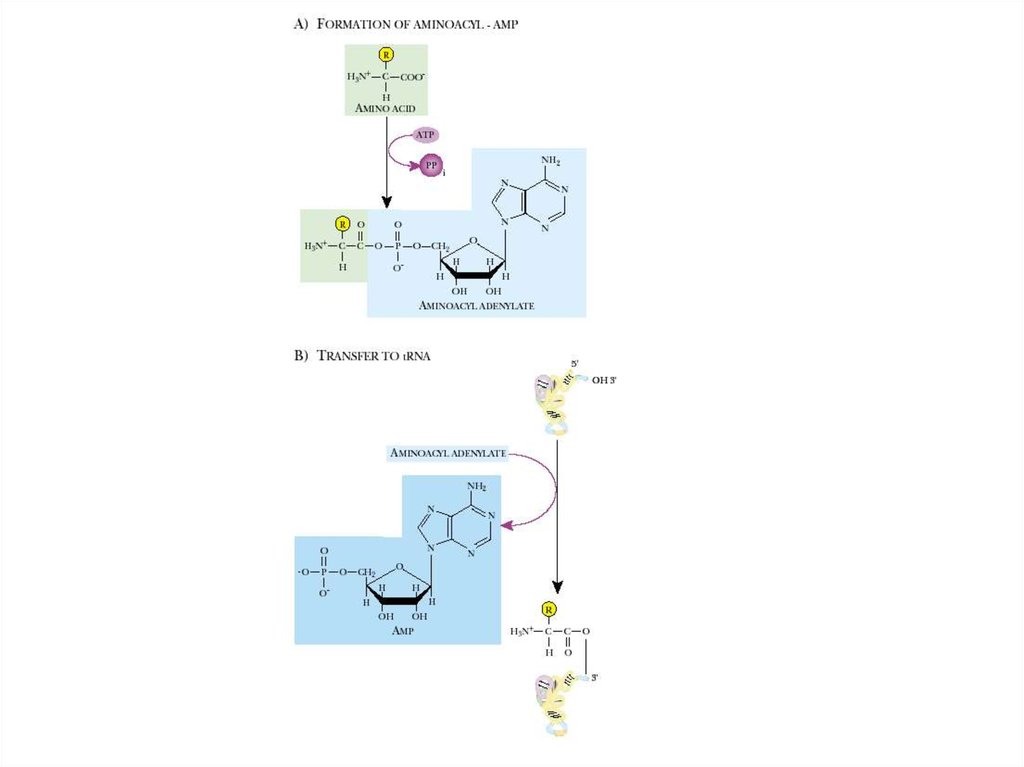
24. Механизм аминоацилирования:
На первой стадии происходит
активация аминокислоты с
использованием энергии,
запасенной в АТР.
На второй стадии активированный
аминоацильный остаток
переносится на одну из
гидроксильных групп рибозного
кольца концевого аденозина на ССАконце тРНК.
Например, валил-тРНК-синтетаза
катализирует суммарную реакцию
синтеза валил-тРНК:
Val + ATP + тРНК ----Val-тРНК +
АМР+РР
Идущую через следующие две
стадии:
– Е + Val + АТР-—— Е*(Val-АМР) +
РР,
– Е*(Val-АМР)----------АМР + Val-тРНК.
25.
26. Точность работы:
Исключительно низкая частота ошибок при аминоацилировании
тРНК (<10-4) является непременным условием реализации
генетического кода.
При биосинтезе белков в рибосомах выбор аминокислоты,
включающейся в растущую белковую цепь, зависит
исключительно от адапторной молекулы тРНК, к которой она
прикреплена.
Если на предрибосомном этапе произошла ошибка и к тРНК
присоединилась аминокислота, не соответствующая
специфичности антикодона, то эта ошибка уже не может быть
исправлена на последующих этапах белкового синтеза.
27. Основы узнавания тРНК аминоацил-тРНК-синтетазами:
Набор узнавания, естественно разный для разных пар тРНК синтетаза, включает нуклеотиды, занимающие одни и те же
положения в структуре большинства тРНК.
Это следующие участки тРНК:
– 1. Антикодон (остатки 34—36). Участие антикодона,
определяющего на рибосоме включение аминокислоты в
растущую цепь белка, еще и в отборе этой аминокислоты на
стадии реакции аминоацилирования тРНК, предполагалось еще
в 60-е годы исходя из уникальности этого элемента в структуре
молекулы.
– 2. Нуклеотид 73, предшествующий ССА-концу. Присутствие в
этом положении того или другого пуринового нуклеотида (А
или G) коррелирует с типом аминокислот, присоединяемых к
тРНК. Если в этом положении находится А, то тРНК
акцептирует гидрофобные аминокислоты, а если G — то
полярные.
– 3. Первые три пары нуклеотидов акцепторного стебля (1-72, 2—
71, 3-70). В разных случаях в узнавании синтетазой может
вовлекаться от одной до трех пар нуклеотидов акцепторного
стебля.
28. Отбор аминокислот:
• Аминокислоты построены из специфического радикала,аминогруппы и карбоксильной группы.
• Вклад различных фрагментов молекулы аминокислоты
исследовали с помощью неприродных синтетических
аналогов аминокислот, в которых были изменены
специфический радикал, аминогруппа или карбоксильная
группа.
• Было показано для различных аминоацил-тРНК-синтетаз,
что необходимым элементом узнавания всех аминокислот
является аминогруппа. Модификация этой группы
приводит к резкому падению сродства аминокислоты к
ферменту.
• Как принято считать, карбоксильная группа играет
меньшую роль на стадии первичного связывания
аминокислоты, однако безусловно важна на последующих
каталитических этапах реакции аминоацилирования.
• Специфическое распознавание аминокислот аминоацилтРНК-синтетазами осуществляется за счет взаимодействий
со специфическими радикалами аминокислот.
29. Аминоацил-тРНК-синтетазы:
• Несмотря на то что эти белки катализируют однуреакцию, они оказались чрезвычайно
разнообразными по размеру, субъединичному
строению и последовательности аминокислот.
• Например, молекулярные массы АРСаз кишечной
палочки варьируют от 51 000 до 384 000, при этом
часть ферментов функционирует в виде мономеров,
часть — в виде димеров, встречаются также и
тетрамеры.
• Оказалось, что между всеми АРСазами
действительно нет сходства, но их можно разделить
на два класса и внутри каждого из классов
обнаруживаются характерные последовательности
аминокислот (так называемые мотивы) и сходные
элементы третичной структуры.
30. Классификация АСРаз:
31. ГИПОТЕЗЫ О ПРОИСХОЖДЕНИИ АРСаз:
• Оказалось, что N-концевой участок аланиновойАРСазы длиной 368 аминокислот способен
синтезировать аминоациладенилат; фрагмент
длиной 385 аминокислот связывает, но не
аминоацилирует тРНК, а фрагмент длиной в 461
аминокислоту успешно аминоацилирует не только
тРНК, но и мини- и микроспирали.
• При этом урезанный фермент, представляющий
собой собственно центральный каталитический
домен АРСазы аминоацилирует тРНК хуже, чем
полный фермент, а аминоацилирование миниспиралей осуществляется с одинаковой эффективностью как целой АРСазой, так и ее N-концевым
доменом.
32. Гипотеза Шиммеля:
П. Шиммель выдвинул гипотезу,
согласно которой и антикодоновый
домен тРНК, и ответственный за
взаимодействие с ним домен АРСаз
являются более поздним
эволюционным приобретением.
Исходная, предковая тРНК
представляла собой короткую
шпильку типа микроспирали, к
которой присоединялись
аминокислоты при помощи предков
современных АРСаз — коротких
полипептидов, содержащих
нуклеотидсвязывающие мотивы.
В "дорибосомную" эпоху синтез
коротких полипептидов
осуществлялся непосредственно на
молекуле предковой тРНК путем
переноса аминокислоты,
присоединенной к З'-гидроксильной
группе концевого рибозного остатка,
на аминогруппу аминокислоты,
присоединенной к 2'-гидроксилу того
же остатка рибозы.
Аналогичным образом происходит
современный биосинтез белка на
рибосоме, за исключением того, что
растущий пептид переносится с одной
молекулы тРНК на другую.
33. Гипотеза Шиммеля:
• Для обеспечения такого древнего синтеза пептидовнеобходимы были по крайней мере два фермента,
способные присоединять аминокислоты к 2'- и к З'-ОН
рибозы, поскольку для этих реакций требуется совершенно
различная пространственная организация субстратов в
активном центре.
• Эти ферменты и стали родоначальниками двух классов
современных АРСаз.
• В дальнейшем белоксинтезирующий аппарат стал более
сложным, для хранения информации о большом
количестве разнообразных белков возник генетический
код, и для обеспечения запрограммированного синтеза
белка на РНК-матрице к предковым тРНК присоединился
антикодоновый домен, а к АРСазам - дополнительные
модули, позволяющие узнавать антикодон тРНК и таким
образом повысить специфичность и точность
аминоацилирования.
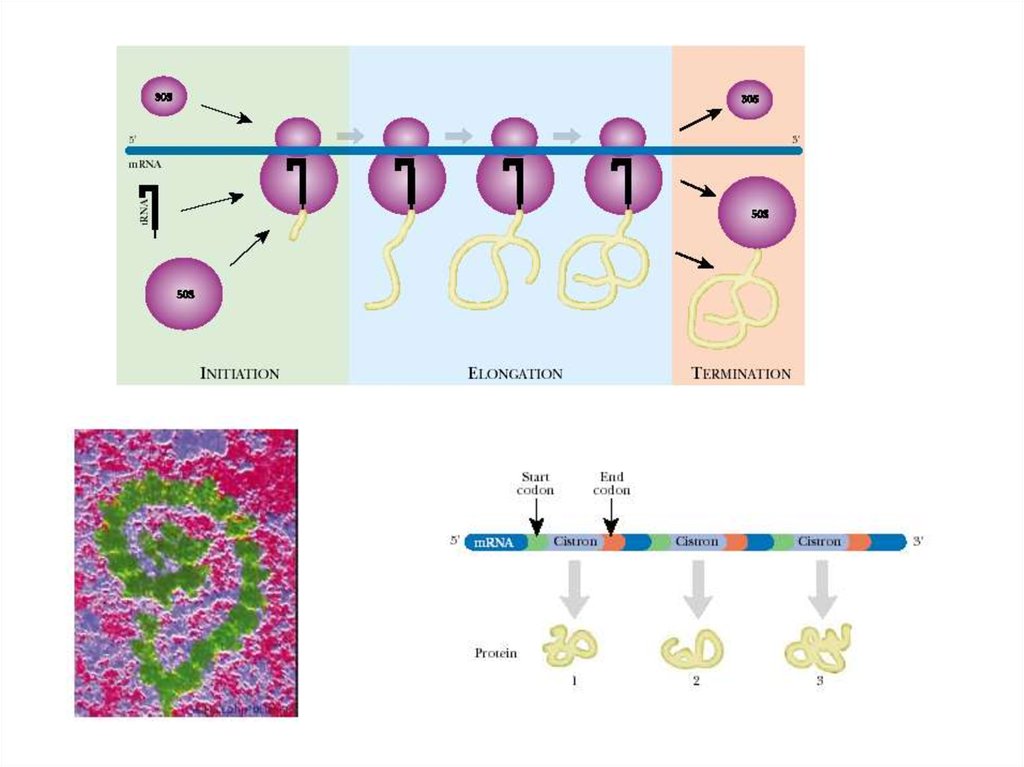
34. Синтез белка: общие сведения:
• Процесс создания химической структуры белка (синтезполипептидной цепи) и в значительной мере ее физическое
сворачивание в функционально активную белковую глобулу
осуществляются рибосомой.
• Количество рибосом в клетке сильно варьирует — от тысяч до
десятков тысяч на клетку — в зависимости от интенсивности
белкового синтеза в данном типе клеток.
• Каждая рибосома полностью прочитывает одну молекулу мРНК
и в соответствии с ее программой синтезирует одну молекулу
белка, после чего может быть запрограммирована другой
молекулой мРНК и произвести другую молекулу белка и т.д.
• Обычно одна молекула мРНК читается сразу несколькими
рибосомами, двигающимися вдоль мРНК друг за другом и, таким
образом, независимо синтезирующими идентичные молекулы
белка, но с соответствующим отставанием. Такой динамический
комплекс одной мРНК с несколькими рибосомами называется
полирибосомой.
35. Рибосома:
Химически рибосома есть рибонуклеопротеид: она
состоит из специальной рибосомной РНК и
специальных рибосомных белков, находящихся в
комплексе друг с другом.
Физически рибосома представляет собой компактную
частицу специфической формы, лишенную внутренней
и внешней симметрии, грубо аппроксимируемую
сферой с диаметром около 30 нм.
Функционально это молекулярная машина,
протягивающая вдоль себя цепь мРНК, считывающая
закодированную в мРНК генетическую информацию и
параллельно, в соответствии с кодом, синтезирующая
полипептидную цепь белка из поступающих в нее
аминокислотных остатков.
В процессе работы рибосома потребляет энергию
гидролиза гуанозинтрифосфата (ГТФ).
Очевидно, что детальное знание структуры рибосомы
является необходимой базой для понимания
механизмов работы этой молекулярной машины.
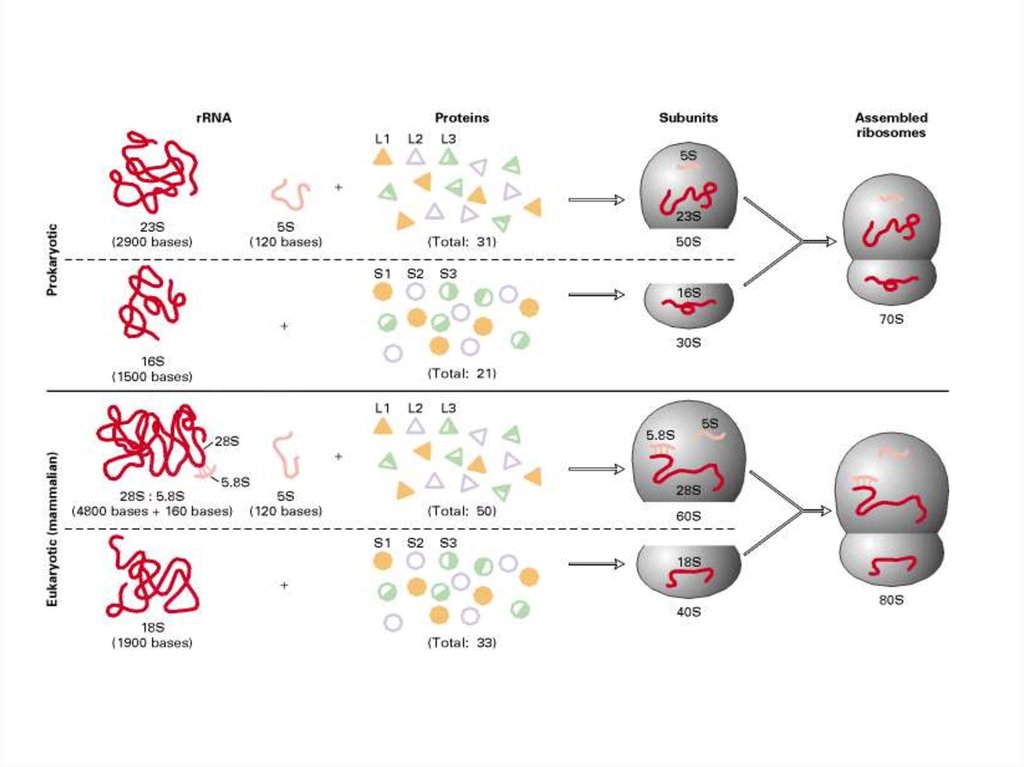
36. ПРИНЦИП № 1: ДВЕ НЕРАВНЫЕ СУБЧАСТИЦЫ
• Электронно-микроскопические изображения рибосом яснопоказывают, что эти округлые частицы подразделяются на
две неравные части.
• Действительно, если в среде, окружающей рибосомы,
понизить концентрацию ионов магния или каким-либо еще
образом увеличить электростатическое отталкивание
фосфатных групп рибосомной РНК, то рибосомная частица
диссоциирует на две неравные субчастицы с
соотношением их масс около 2:1.
• Полные рибосомные частицы и их субчастицы принято
обозначать в соответствии с их коэффициентами
седиментации (скоростями осаждения) в
ультрацентрифуге, выражаемыми в единицах Сведберга
(S).
• Бактериальная рибосома с молекулярной массой около
3•106 имеет коэффициент седиментации 70S и
обозначается как 70S-частица, а несколько более крупная
рибосома эукариотических организмов предстает как 80Sчастица.
37.
38. ПРИНЦИП № 2: САМОСВОРАЧИВАНИЕ РИБОСОМНОЙ РНК В КОМПАКТНОЕ ЯДРО
Каждая рибосомная
субчастица содержит одну
молекулу высокополимерной
рибосомной РНК,
составляющую от половины
до двух третей всей массы
субчастицы.
Изолированные цепи
высокополимерной
рибосомной РНК в условиях,
гасящих электростатическое
отталкивание их фосфатных
групп (высокие концентрации
солей, особенно ионов
магния), способны
сворачиваться в компактные
частицы характерной формы,
причем компактно свернутая
23S РНК напоминает по форме
полусферическую большую
субчастицу рибосомы, а 16S
РНК —удлиненную малую
субчастицу.
39. ПРИНЦИП № 3: СБОРКА РАЗНООБРАЗНЫХ БЕЛКОВ НА РНК
Каждая рибосомная субчастица содержит много молекул
рибосомных белков, и все они разные.
Многочисленные рибосомные белки могут играть двоякую роль в
современной рибосоме.
С одной стороны, они могут непосредственно участвовать в
функциях связывания субстратов и каталитических функциях
рибосомы, локализуясь в соответствующих функциональных
центрах и обеспечивая их своими активными группами.
С другой — рибосомные белки могут служить стабилизаторами
или модификаторами определенных локальных структур
рибосомной РНК и таким образом поддерживать их в
функционально активном состоянии или способствовать их
переключениям из одного состояния в другое.
В частности, в отношении главной каталитической функции
рибосомы — ее пептидил-трансферазной активности,
ответственной за образование пептидных связей, имеются все
основания полагать, что эта активность обеспечивается
локальной структурой рибосомной РНК большой субчастицы, но
некоторые рибосомные белки оказываются необходимыми для
поддержания (стабилизации) этой структуры.
40.
41.
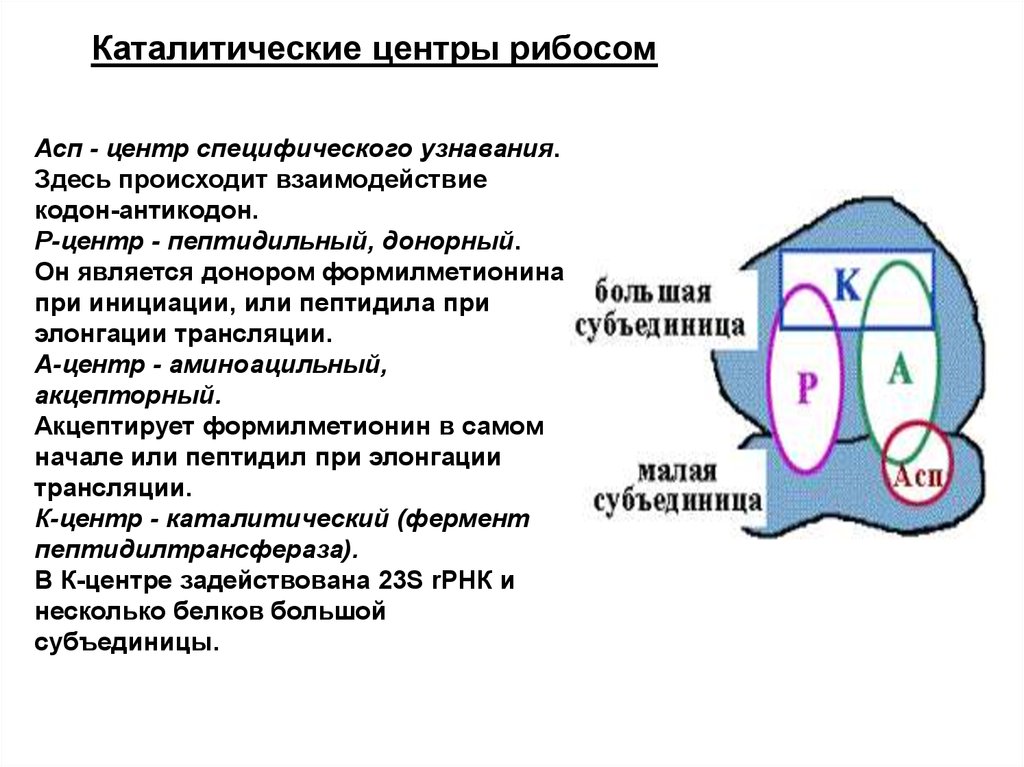
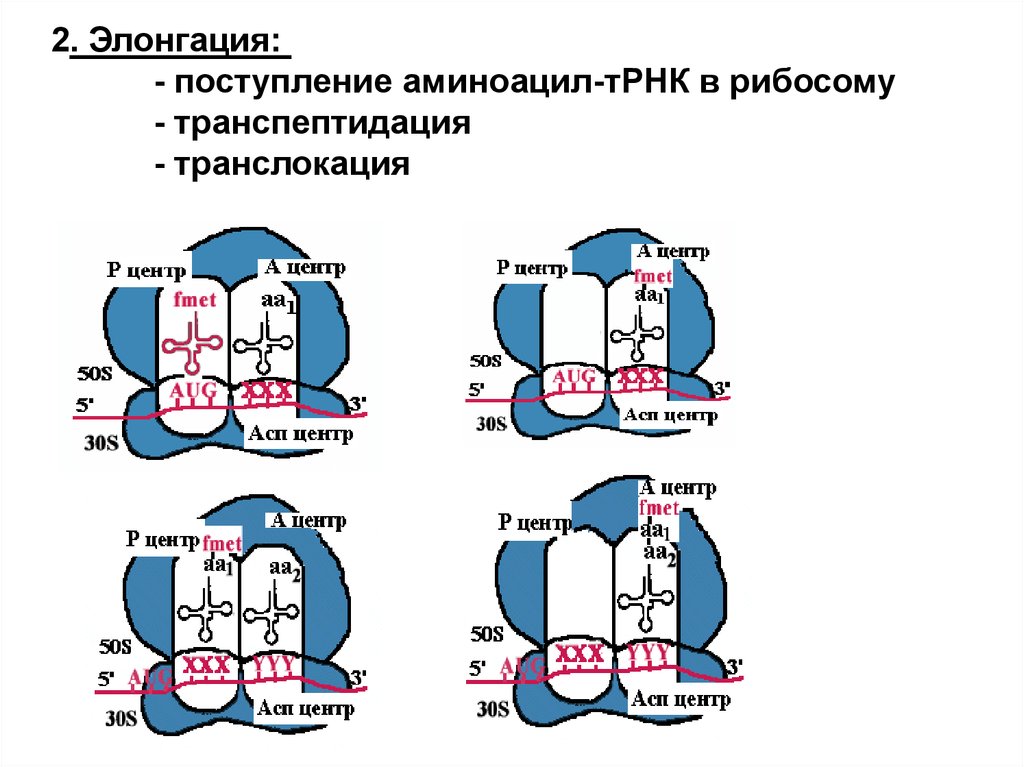
Каталитические центры рибосомАсп - центр специфического узнавания.
Здесь происходит взаимодействие
кодон-антикодон.
Р-центр - пептидильный, донорный.
Он является донором формилметионина
при инициации, или пептидила при
элонгации трансляции.
А-центр - аминоацильный,
акцепторный.
Акцептирует формилметионин в самом
начале или пептидил при элонгации
трансляции.
К-центр - каталитический (фермент
пептидилтрансфераза).
В К-центре задействована 23S rРНК и
несколько белков большой
субъединицы.
42.
43.
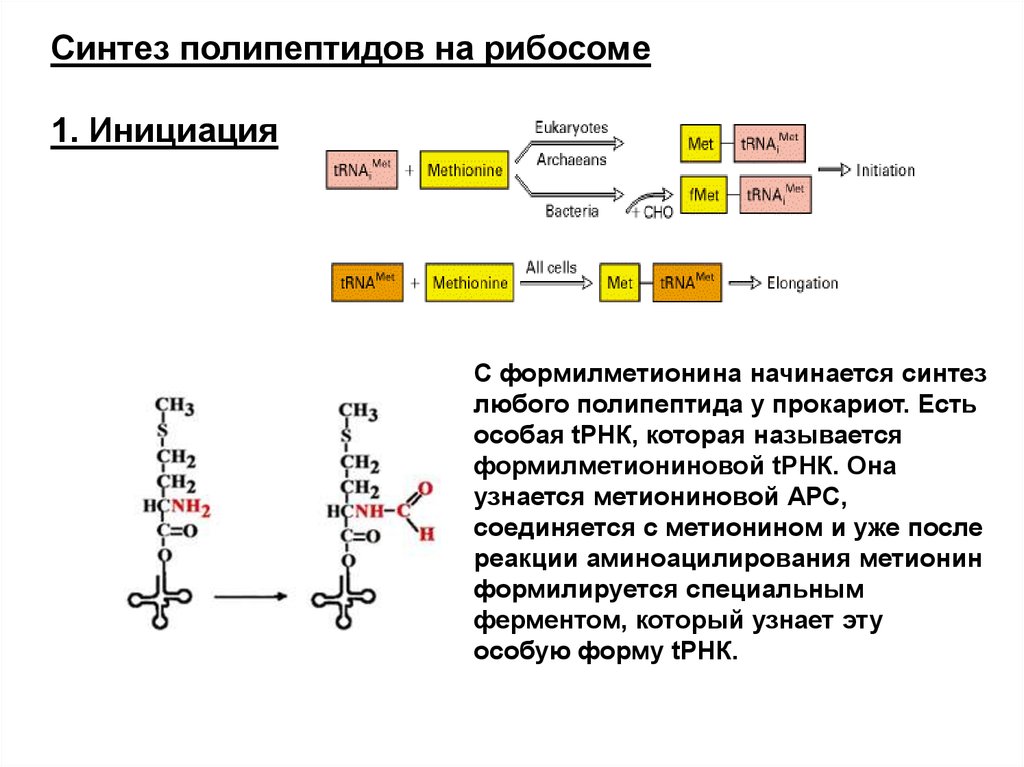
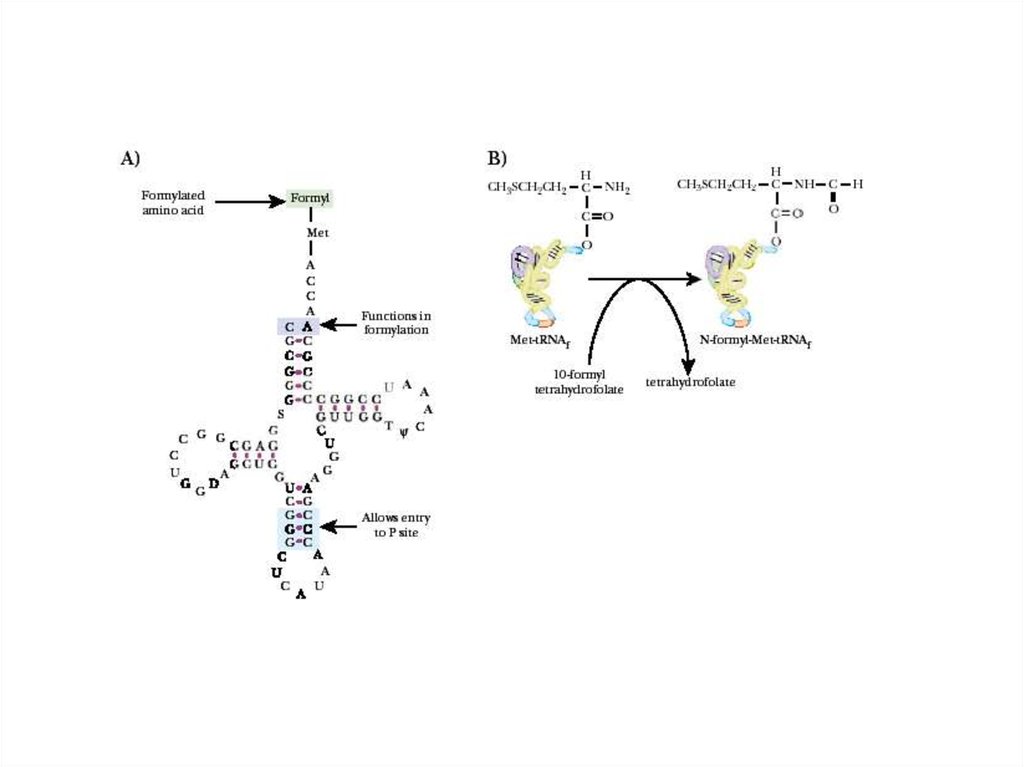
Синтез полипептидов на рибосоме1. Инициация
С формилметионина начинается синтез
любого полипептида у прокариот. Есть
особая tРНК, которая называется
формилметиониновой tРНК. Она
узнается метиониновой АРС,
соединяется с метионином и уже после
реакции аминоацилирования метионин
формилируется специальным
ферментом, который узнает эту
особую форму tРНК.
44.
45.
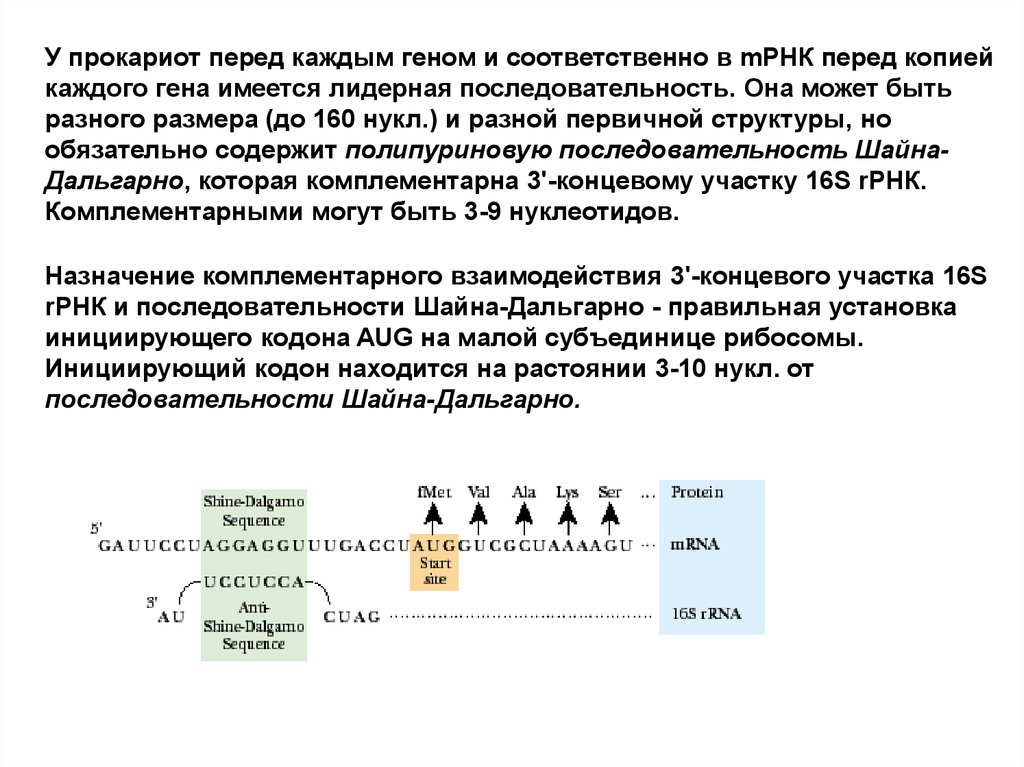
У прокариот перед каждым геном и соответственно в mРНК перед копиейкаждого гена имеется лидерная последовательность. Она может быть
разного размера (до 160 нукл.) и разной первичной структуры, но
обязательно содержит полипуриновую последовательность ШайнаДальгарно, которая комплементарна 3'-концевому участку 16S rРНК.
Комплементарными могут быть 3-9 нуклеотидов.
Назначение комплементарного взаимодействия 3'-концевого участка 16S
rРНК и последовательности Шайна-Дальгарно - правильная установка
инициирующего кодона AUG на малой субъединице рибосомы.
Инициирующий кодон находится на растоянии 3-10 нукл. от
последовательности Шайна-Дальгарно.
46.
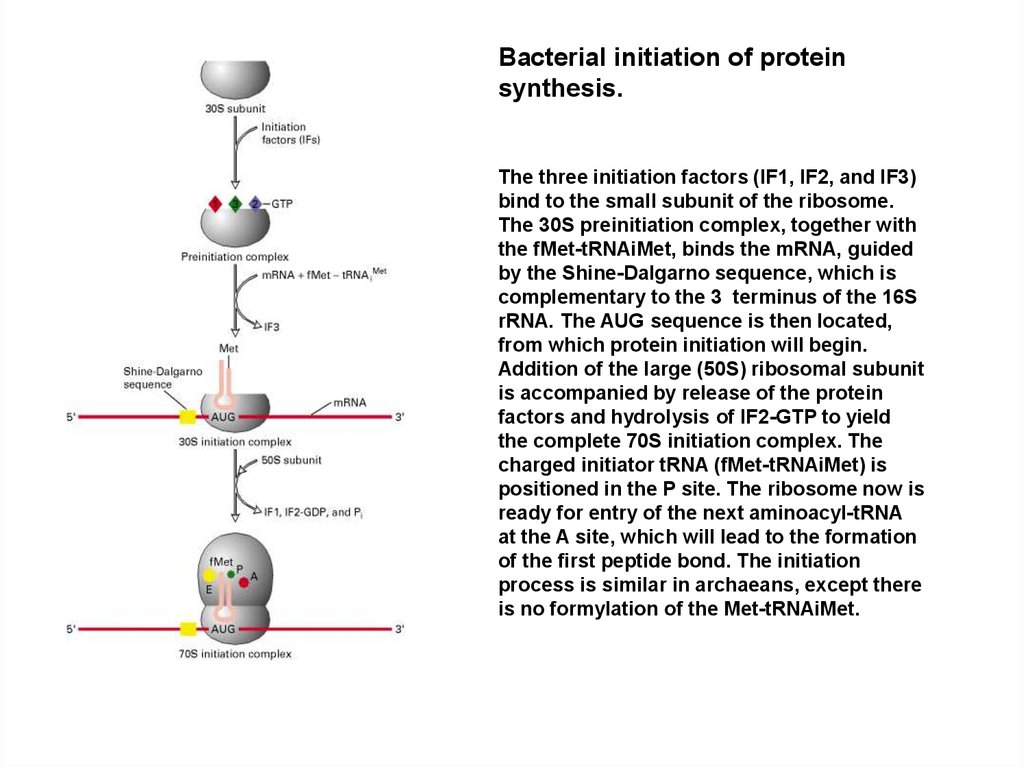
Bacterial initiation of proteinsynthesis.
The three initiation factors (IF1, IF2, and IF3)
bind to the small subunit of the ribosome.
The 30S preinitiation complex, together with
the fMet-tRNAiMet, binds the mRNA, guided
by the Shine-Dalgarno sequence, which is
complementary to the 3 terminus of the 16S
rRNA. The AUG sequence is then located,
from which protein initiation will begin.
Addition of the large (50S) ribosomal subunit
is accompanied by release of the protein
factors and hydrolysis of IF2-GTP to yield
the complete 70S initiation complex. The
charged initiator tRNA (fMet-tRNAiMet) is
positioned in the P site. The ribosome now is
ready for entry of the next aminoacyl-tRNA
at the A site, which will lead to the formation
of the first peptide bond. The initiation
process is similar in archaeans, except there
is no formylation of the Met-tRNAiMet.
47.
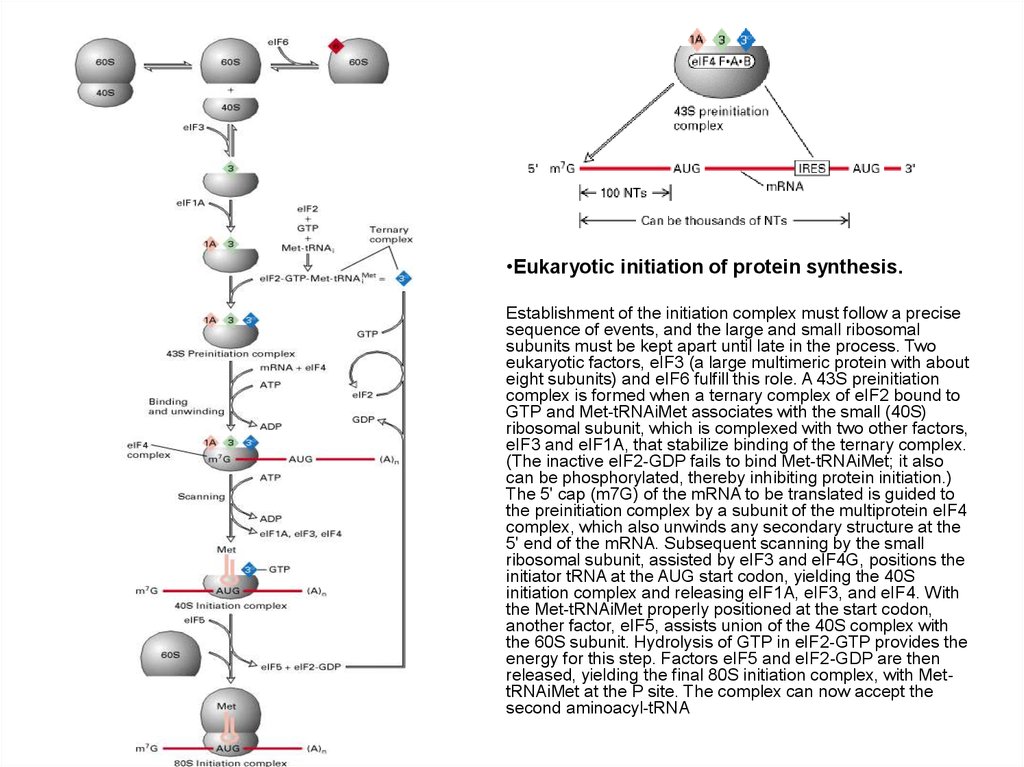
•Eukaryotic initiation of protein synthesis.Establishment of the initiation complex must follow a precise
sequence of events, and the large and small ribosomal
subunits must be kept apart until late in the process. Two
eukaryotic factors, eIF3 (a large multimeric protein with about
eight subunits) and eIF6 fulfill this role. A 43S preinitiation
complex is formed when a ternary complex of eIF2 bound to
GTP and Met-tRNAiMet associates with the small (40S)
ribosomal subunit, which is complexed with two other factors,
eIF3 and eIF1A, that stabilize binding of the ternary complex.
(The inactive eIF2-GDP fails to bind Met-tRNAiMet; it also
can be phosphorylated, thereby inhibiting protein initiation.)
The 5' cap (m7G) of the mRNA to be translated is guided to
the preinitiation complex by a subunit of the multiprotein eIF4
complex, which also unwinds any secondary structure at the
5' end of the mRNA. Subsequent scanning by the small
ribosomal subunit, assisted by eIF3 and eIF4G, positions the
initiator tRNA at the AUG start codon, yielding the 40S
initiation complex and releasing eIF1A, eIF3, and eIF4. With
the Met-tRNAiMet properly positioned at the start codon,
another factor, eIF5, assists union of the 40S complex with
the 60S subunit. Hydrolysis of GTP in eIF2-GTP provides the
energy for this step. Factors eIF5 and eIF2-GDP are then
released, yielding the final 80S initiation complex, with MettRNAiMet at the P site. The complex can now accept the
second aminoacyl-tRNA
48.
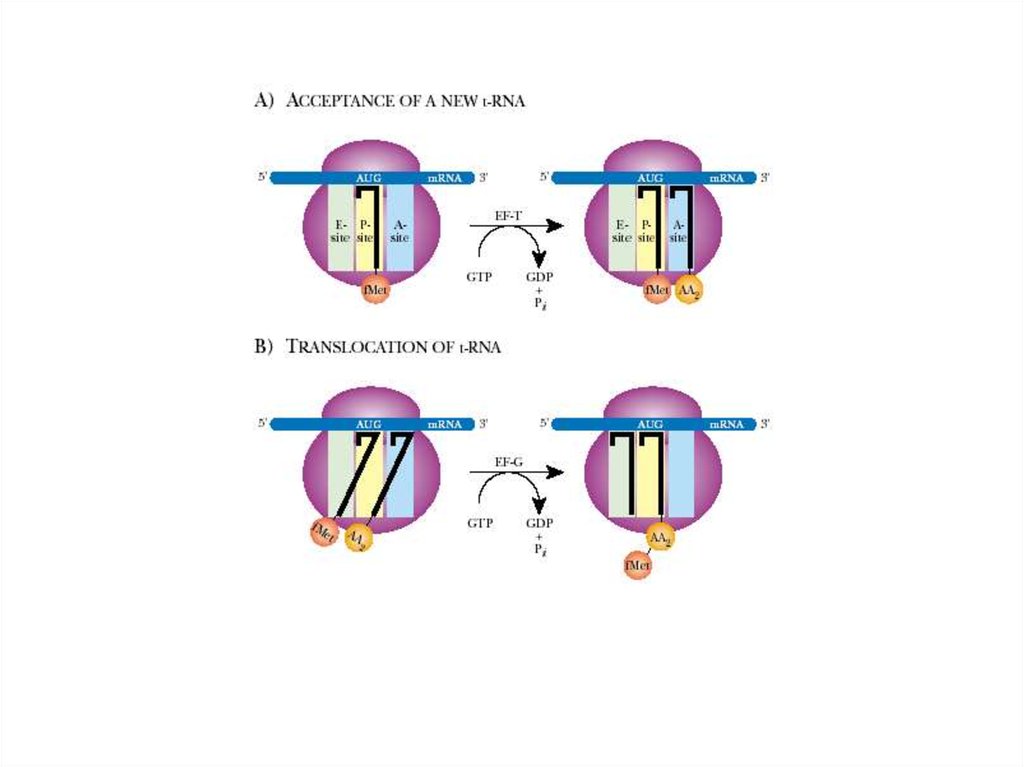
2. Элонгация:- поступление аминоацил-тРНК в рибосому
- транспептидация
- транслокация
49.
50.
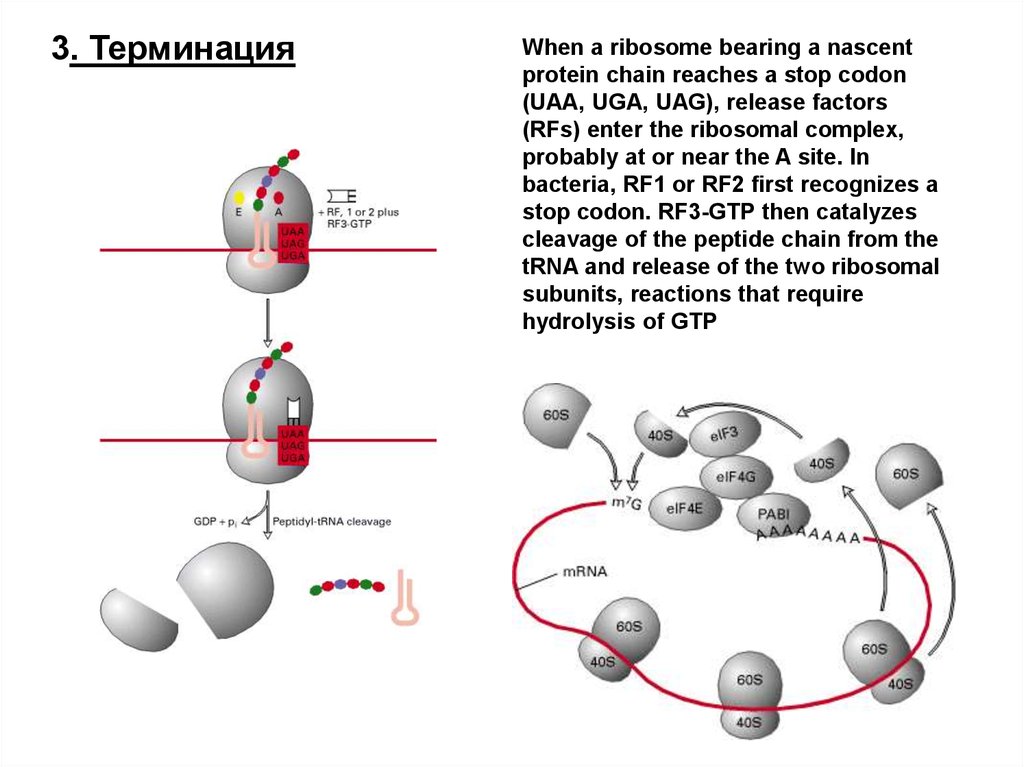
3. ТерминацияWhen a ribosome bearing a nascent
protein chain reaches a stop codon
(UAA, UGA, UAG), release factors
(RFs) enter the ribosomal complex,
probably at or near the A site. In
bacteria, RF1 or RF2 first recognizes a
stop codon. RF3-GTP then catalyzes
cleavage of the peptide chain from the
tRNA and release of the two ribosomal
subunits, reactions that require
hydrolysis of GTP
51.
52.
53.
54.
55.
56.
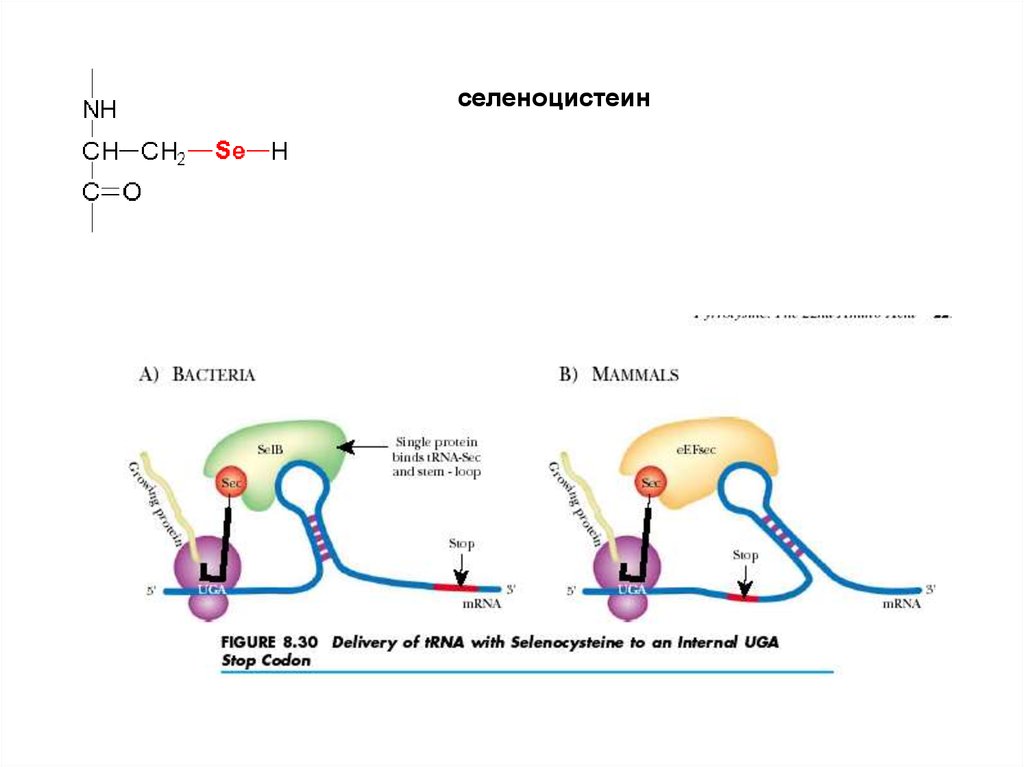
селеноцистеин57.
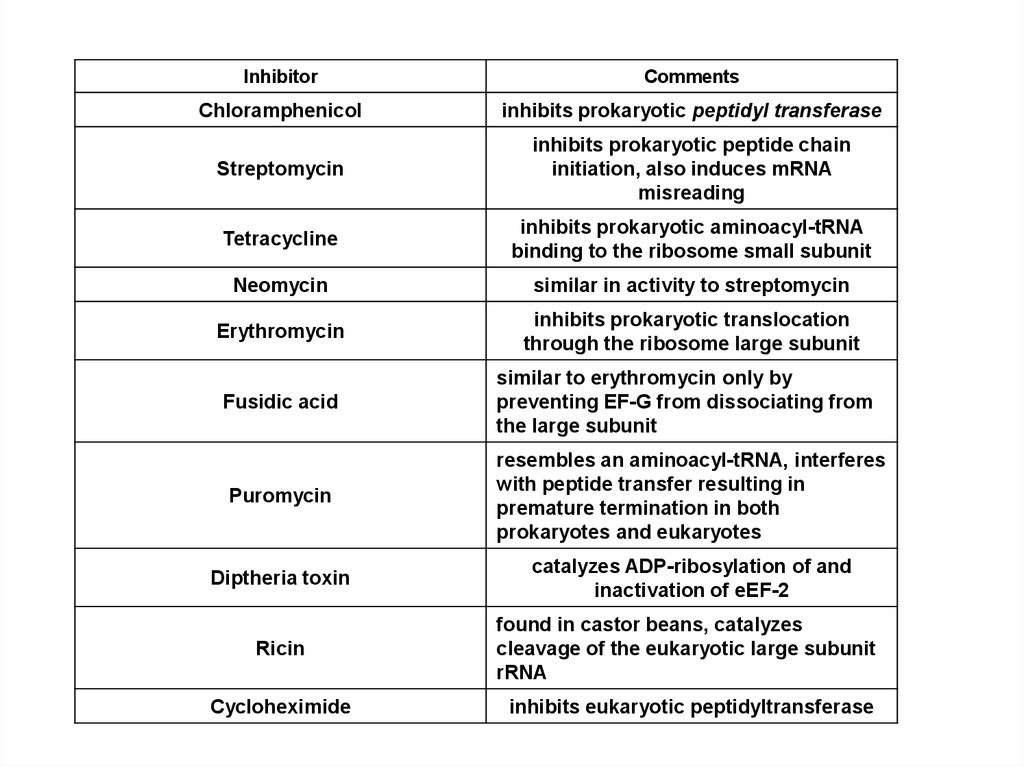
InhibitorComments
Chloramphenicol
inhibits prokaryotic peptidyl transferase
Streptomycin
inhibits prokaryotic peptide chain
initiation, also induces mRNA
misreading
Tetracycline
inhibits prokaryotic aminoacyl-tRNA
binding to the ribosome small subunit
Neomycin
similar in activity to streptomycin
Erythromycin
inhibits prokaryotic translocation
through the ribosome large subunit
Fusidic acid
similar to erythromycin only by
preventing EF-G from dissociating from
the large subunit
Puromycin
resembles an aminoacyl-tRNA, interferes
with peptide transfer resulting in
premature termination in both
prokaryotes and eukaryotes
Diptheria toxin
catalyzes ADP-ribosylation of and
inactivation of eEF-2
Ricin
found in castor beans, catalyzes
cleavage of the eukaryotic large subunit
rRNA
Cycloheximide
inhibits eukaryotic peptidyltransferase