








 от размеров самок (L, мм) у мизиды Paramysis lacustris из Каунасского вдхр. (а);")
, их плодовитость снижается, несмотря на")

")

")











































 линейно снижается с увеличением ее численности (N).")












 Социология
СоциологияПохожие презентации:










Факторы динамики численности популяций
1. Лекции 11 – 12 Факторы динамики численности популяций
2.
Численность популяции никогда не остаетсяпостоянной, она постоянно
претерпевает
сезонные или многолетние изменения, которые
обусловлены многочисленными факторами.
В общем виде изменения численности
популяции за единицу времени
определяются
следующим образом:
3.
= (В + im) – (D + em),где:
В - рождаемость, или количество новорожденной
молоди, произведенной времени всеми имеющимися в
популяции половозрелыми особями;
D – смертность, или количество погибших особей в
популяции, независимо от причины их смерти – старость,
выедание хищниками, болезни, промысел, гибель в
природных катастрофах и т.д.;
im – иммиграция, или число особей, поступивших в
данную популяцию из других популяций;
em – эмиграция, или число особей, убывших из данной
популяции в другие популяции.
Все эти параметры определяются для определенных
промежутков времени (сутки, месяц, год и т.д.) и имеют
размерность [особь·время-1].
4.
Если величинамиim и em можно
пренебречь, то изменения численности
популяции будут определяться разностью
между
величинами
рождаемости
и
смертности:
= В – D,
При В = D численность популяции
остается постоянной, поскольку
.
При В > D она возрастает,
а при В < D – снижается.
5.
Рождаемость в популяциях определяется,преимущественно свойствами особей и популяций,
из которых важнейшими являются:
Абсолютная, или индивидуальная плодовитость –
число потомков (яйца, личинки, молодь, семена,
споры и т.п.), производимых материнской особью за
один акт размножения;
Длительность
генеративного
цикла,
или
промежутка
времени
между
двумя
последовательными актами размножения у отдельных
особей;
Время генерации, или продолжительность жизни
одного поколения.
Общая численность (плотность) популяции;
Доля особей,
производящих потомство, в
популяции.
6.
Тем не менее, на все эти параметрырождаемости
существенное
воздействие
оказывают факторы внешней среды.
Смертность в популяциях, напротив,
определяют,
прежде всего, внешние
факторы, которые кратко были перечислены
выше.
Поэтому, хотя каждый вид имеет
генетически закрепленную максимальную
длительность жизни, в природных условиях
до нее доживает лишь незначительная часть
популяции.
7.
Абсолютная плодовитость (F)У
одноклеточных
организмов,
размножающихся
делением надвое (бактерии, протисты), F = 1.
Плодовитость гомойотермных животных очень низка –
от 1 до 5 – 7, очень редко до 12 – 15 детенышей (серая
полевка, крыса) и 20 – 22 яиц (куриные птицы).
У пойкилотермных организмов Е, как правило,
значительно выше – от нескольких десятков и сотен – у
травянистых
растений,
коловраток,
ракообразных,
брюхоногих моллюсков, амфибий, рептилий – до сотен
тысяч и миллионов – у древесных растений, грибов,
крупных двустворчатых моллюсков и рыб.
Если число потомков, произведенных за один акт
размножения, больше единицы, их совокупность, особенно
у животных, часто называют кладкой или пометом.
8.
Ниже своего максимальногоуровня,
плодовитость, особенно у видов с достаточно
высокими F, может изменяться в широких
пределах под воздействием факторов внешней
среды (температура, световой режим, количество
корма и его состав, плотность популяции и т.д.),
размеров
и
физиологического
состояния
организма.
У птиц и млекопитающих F практически не
зависит от размеров самок.
У большинства пойкилотермных животных, а
также многих древесных растений в условиях,
близких к оптимальным, значения F возрастают с
увеличением размеров материнских особей (L).
9.
Это связь обычно выражается степенным уравнением:F = mLn
где m и n - константы.
В логарифмических координатах данное уравнение
трансформируется в уравнение прямолинейной регрессии
lgF = lgm + nlgL
У моллюсков, ракообразных и насекомых значения
показателя степени n чаще всего изменяются в пределах от
2 до 3, у рыб обычно n > 3.
10. Зависимость плодовитости (F, яйца·особь-1) от размеров самок (L, мм) у мизиды Paramysis lacustris из Каунасского вдхр. (а);
Кайраккумского вдхр. (б) и авандельты Дона (в).F
а
б
в
L, мм
Зависимость плодовитости (F , яйца·особь-1) от размеров L (мм)
у P.lacustris из авандельты Дона следует уравнению: F = 0,159L2,11,
а из Каунасского вдхр. - уравнению F = 0,008L3,10.
11. Часто достижения самками определенного возраста (обычно близкого к предельному), их плодовитость снижается, несмотря на
FЧасто достижения самками
определенного
возраста
(обычно
близкого
к
предельному), их плодовитость
Возраст
снижается,
несмотря на
продолжающийся
рост
их
Возрастные изменения
плодовитости (F) у не- размеров. Это
обусловлено
которых ракообразных
старческими изменениями в
организме.
12.
Характеризменения
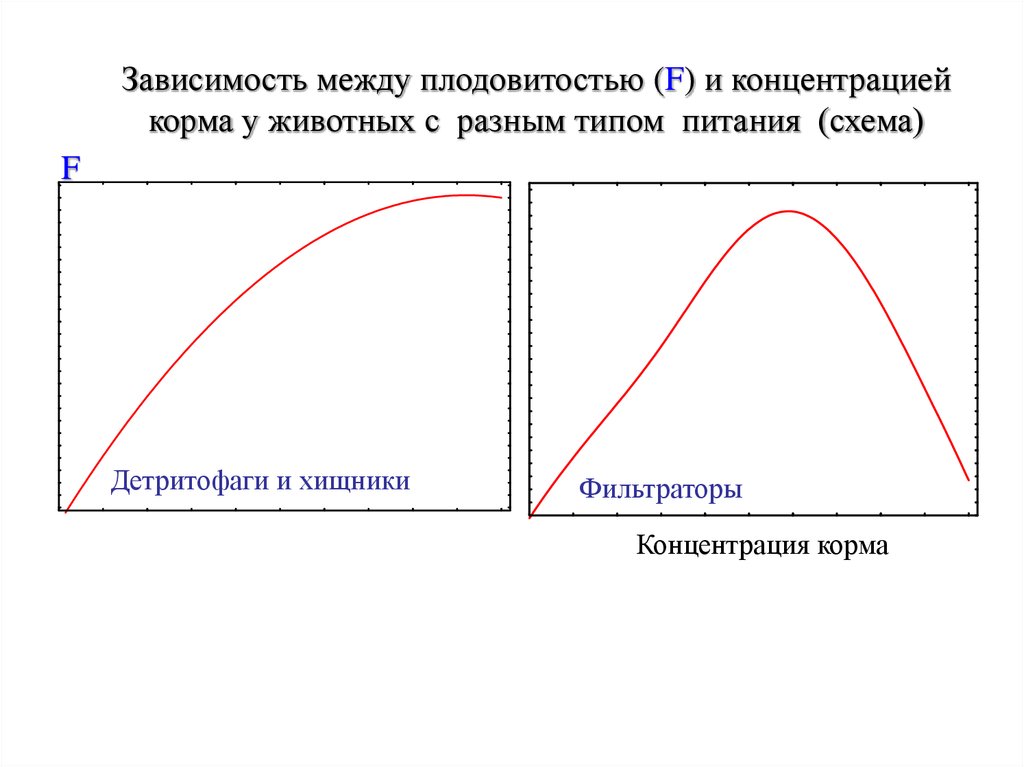
плодовитости
пойкилотермных животных с концентрацией
корма в среде (C) зависит от типа их питания.
Животных в этом отношении можно
разделить на:
хищников, включая и растительноядных
животных,
детритофагов,
или
«мусорщиков»
(потребителей крупных органических остатков);
фильтраторов.
13.
Зависимость между плодовитостью (F) и концентрациейкорма у животных с разным типом питания (схема)
F
Детритофаги и хищники
Фильтраторы
Концентрация корма
14. Влияние плотности популяции на плодовитость животных (схема)
аб
в
N
Плотность
а – кривая «дрозофильного типа»
б – кривая «типа Олли»
в – кривая промежуточного типа
популяции
15.
Однако во всех случаях выявлена однаобщая закономерность – при достижении
некоторой критической плотности популяции
плодовитость особей в ней начинает снижаться
вплоть до нулевого значения.
Это является одним из важнейших
механизмов регуляции численности популяций
по принципу отрицательной обратной связи.
16. Длительность генеративного цикла (G)
У одноклеточныхорганизмов (бактерии,
протисты) G соответствует промежутку времени
между двумя их последовательными делениями.
У многоклеточных организмов, особенно у
мелких зоопланктонных видов (ракообразные,
коловратки), дающих за сезон размножения
несколько кладок, G часто принимается равным
длительности эмбрионального развития яиц.
17.
У гомойотермных животных длительностьэмбриогенеза (беременность у млекопитающих,
насиживание яиц у птиц) отличается высокой
видовой специфичностью.
Она не зависит от температуры, но может
изменяться
в
некоторых
пределах
под
воздействием других факторов – условий питания,
плотности популяции и т.д.
18.
Длительностьбеременности
у
млекопитающих
в
целом
возрастает
с
увеличением их размеров: от 20 - 21 суток у
мышей и крыс до 660 суток у слона.
Однако из этого правила имеются
многочисленные
исключения. Например, у
барсука (масса тела до 16 кг) беременность
длится дольше (357 суток), чем у гораздо более
крупной лошади (400 кг и 330 суток).
У
сумчатых,
хищных
и
других
млекопитающих,
рождающих
слепых
и
беспомощных детенышей, беременность обычно
короче, чем у копытных с сопоставимой массой
тела, детеныши которых способны следовать за
матерью уже через несколько минут после
рождения.
19.
Период насиживания у птиц в целомвозрастает с увеличением массы яиц – от 12 суток
(колибри, масса яйца 20 мг) до 70-100 суток
(императорский пингвин, масса яйца – до 450 г).
Однако самые крупные среди современных
птиц яйца африканского страуса (до 2 кг)
насиживаются всего 35 - 42 суток, примерно
столько же, как и небольшие яйца лебедя (35 г).
20.
Время генерации (Тo), или продолжительностьжизни одного поколения.
Равно сумме длительностей эмбрионального и
ювенильного периодов жизненного цикла особей.
У моноциклических видов (лососевые рыбы и
др.), погибающих сразу же после вымета своей
первой и единственной кладки, Tо совпадает со
средней длительностью жизни половозрелых особей.
У полициклических видов, производящих две и
более кладок за жизненный цикл, Tо определяется
по специальным демографическим таблицам.
21.
Доля размножающихся самок в общейчисленности популяции (α ) равна
α = N♀/N,
где
N♀ - численность размножающихся самок,
N – общая численность популяции.
22.
Скорость размножения особи равна среднемуколичеству потомков (яйца, личинки, молодь и т.п.),
производимому одной особью (самкой) за сутки (Br):
Br =
где
F – плодовитость особи;
G - длительности генеративного цикла, который в
ряде случаев можно принять равным длительности
эмбриогенеза (De)
23.
Скорость размножения популяции равна общемуколичеству потомков, произведенному популяцией
за сутки (Bp):
Bp =
где F – средняя плодовитость самок
G – длительность генеративного цикла
N - численность популяции
α – доля размножающихся самок в ней.
24.
Bp =25. Пример для дафнии Daphnia magna
F = 30 яицG при 20оС = 3 суток
N = 500 экз м-3
α = 0,1
Отсюда,
Br = 30/3 = 10 яиц• сут-1,
Bp = (30 500• 0,1)/3 = 500 яиц • м-3•сут-1
26.
Удельная рождаемостьПри отсутствии в популяции смертности и
достаточном
количестве
пищевых
и
пространственных ресурсов, рост ее численности
может быть очень быстрым.
Например,
если
инфузория
Paramecium
caudautum делится надвое всего раз в сутки, то
численность ее популяции будет удваиваться
каждые сутки.
При отсутствии смертности число потомков
одной инфузории уже через год достигла бы 2364
(или 10110) особей.
27.
Втаком
случае
увеличение
численности
популяции в геометрической прогрессии описывается
экспоненциальным уравнением:
Nt = Noebt
где N0 – начальная численность особей
при t = 0,
Nt - численность особей ко времени t,
b - константа, называемая удельной рождаемостью.
Значение b означает количество потомков, которое
производится в популяции за единицу времени в расчете
на одну особь. Оно имеет размерность [время-1].
28.
Значение b может быть рассчитано,если
известны
два
значения
численности популяции N1 и N2 в
моменты времени t1 и t2:
29.
Рассчитаем значение b для популяцииинфузории за период времени t2 – t1 = 1 суткам.
Поскольку численность популяции каждые сутки
удваивается, то N2 = 2N1.
Отсюда:
b = (ln2N1 - lnN1)/1 =
= ln2 + lnN1 - lnN1 = ln2.
Отсюда b = 0,69 сут-1,
Уравнение роста для популяции инфузории в
численной форме имеет вид:
Nt = N0e0,69t
30.
Однако большинство видов размножается путемпродуцирования молоди или яиц, число которых в
одной кладке обычно больше единицы.
Значения
b для
популяций таких видов
рассчитываются иными способами.
Рассмотрим
вначале
виды
с
партеногенетическим размножением, у которых
потомство производят все половозрелые особи.
Их
примером
являются
ветвистоусые
ракообразные – дафнии и др.
31.
Допустим, популяция состоит только изполовозрелых особей.
Если ее начальная численность (N1) равна 1 экз,
то через 1 сутки она достигнет:
N2 = 1 + F/D.
Отсюда:
b = [ln(1 + F/D) - ln1]/1
Поскольку F/D представляет собой скорость
размножения особей ( Br) и ln1 = 0, получаем:
b = ln(1 + Br)
32. Пример для партеногенетической популяции Daphnia magna.
E = 30 яиц; D = 3 сут.Отсюда:
b = ln(1 + 30/3) = 2,40 сут-1
33.
Если в популяцииимеются особи, не
производящие
потомство
(молодь,
неяйценосные самки, самцы), в полученное
уравнение вносится поправочный коэффициент
α, характеризующий отношение численности
яйценосных самок (N♀) к общей численности
популяции (N), т.е. α = N♀/N .
Тогда:
b = ln(1 + αBr)
34. Пример для двуполой популяции Asellus aquaticus
F = 30 яицD при 20оС – 14 сут.
α = 0,1.
Тогда
b = ln (1 + 0,1 . 30/14) = 0,194 сут-1
.
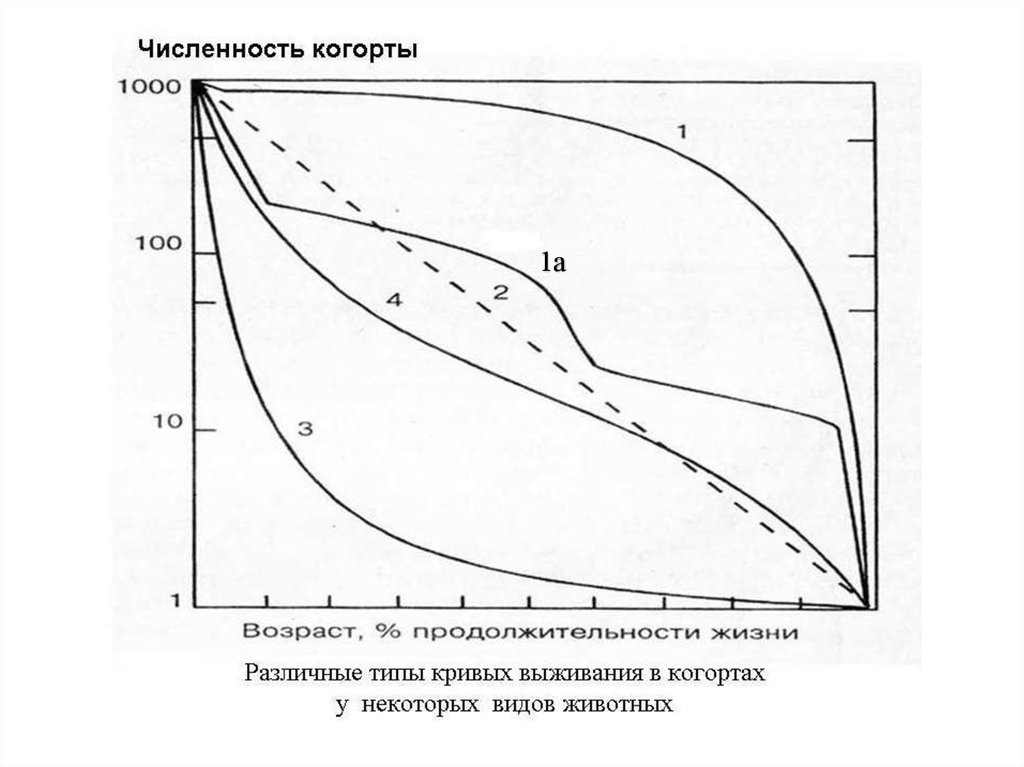
35. Типы смертности в популяциях
Мы будет анализировать смертность нево всей популяции, а в отдельных ее
когортах, или группах одновременно
отрожденных особей.
Все особи в популяциях
I-го типа
составляют одну когорту.
Выделяют четыре основных
типа
смертности в когортах:
36.
1а37. I-й тип смертности
Отражаетситуацию,
при
которой
смертность до определенного возраста низкая,
а затем быстро возрастает.
Характерен для многих насекомых с
неполным превращением, пресмыкающихся,
большинства млекопитающих, в том числе
человека.
38. Ia-тип смертности
Ступенчатая кривая характернадля
видов с метаморфозом, у которых
смертность резко возрастает при переходе
от одной стадии жизненного цикла к
другой.
Их примером являются насекомые с
полным превращением – бабочки и др.
Смертность их яиц и имаго выше, чем
личинок и куколок.
39. II-ой тип смертности
Характеризует относительное постоянствосмертности во всех возрастах.
Свойственен
преимущественно
выводковым
птицам,
мелким
млекопитающим (мыши, кролики)
и
некоторым другим видам, напр., рептилиям.
40. III-й тип смертности
Характеризуется очень высокой смертностью враннем возрасте и относительно низкой в старших
возрастах. Встречается у организмов с очень высокой
плодовитостью и отсутствием форм заботы о потомстве
– древесных растений, многих рыб, морских
двустворчатых
моллюсков
(устрицы,
мидии),
иглокожих.
Эти виды производят огромное количество семян,
мелких икринок, личинок и т.п. До половой зрелости
доживает лишь их ничтожная часть, однако смертность
взрослых особей, после достижения ими определенных
размеров достаточно низка.
Поэтому III тип смертности иногда называют
устричным типом смертности.
41.
42.
Кривые выживанияв когортах состоятельных
людей в Римской империи в I – II ст. н.э. на примере
столицы
империи
г.
Рима
и
одной
из
североафриканских провинций.
43.
Удельную скорость смертности (d) в когорте(при
любых ее типах)
за определенный
промежуток времени (t2–t1) можно рассчитать,
если известны значения численности когорты в
начале и конце этого промежутка (N2 и N1).
Поскольку N2 < N1, значения d являются
отрицательным и имеют размерность [время-1].
Они показывают число погибших особей в
популяции за единицу времени в расчете на одну
особь.
44.
Если за достаточно длительный периодвремени значения d остаются постоянным, то
численность когорты за этот период снижается
экспоненциально.
В случае постоянства d за весь жизненный
цикл получаем экспоненциальное снижение
численности когорты:
Nt = Noe-dt,
где No начальная численность особей в
когорте, Nt – ее численность к возрасту t, d –
удельная скорость смертности.
Легко видеть, что постоянство d в когорте
соответствует II-му типу смертности.
45.
Отсюда возможно определить среднее значениемаксимальной длительности жизни особей (tmax).
Таковой можно принять возраст, до которого
доживает 1% от начальной численности когорты.
Если No = 100 особей, то Nt = 1 особь. Тогда
1 = 100e– dt
Отсюда
tmax = ln100/d = 4,6/d.
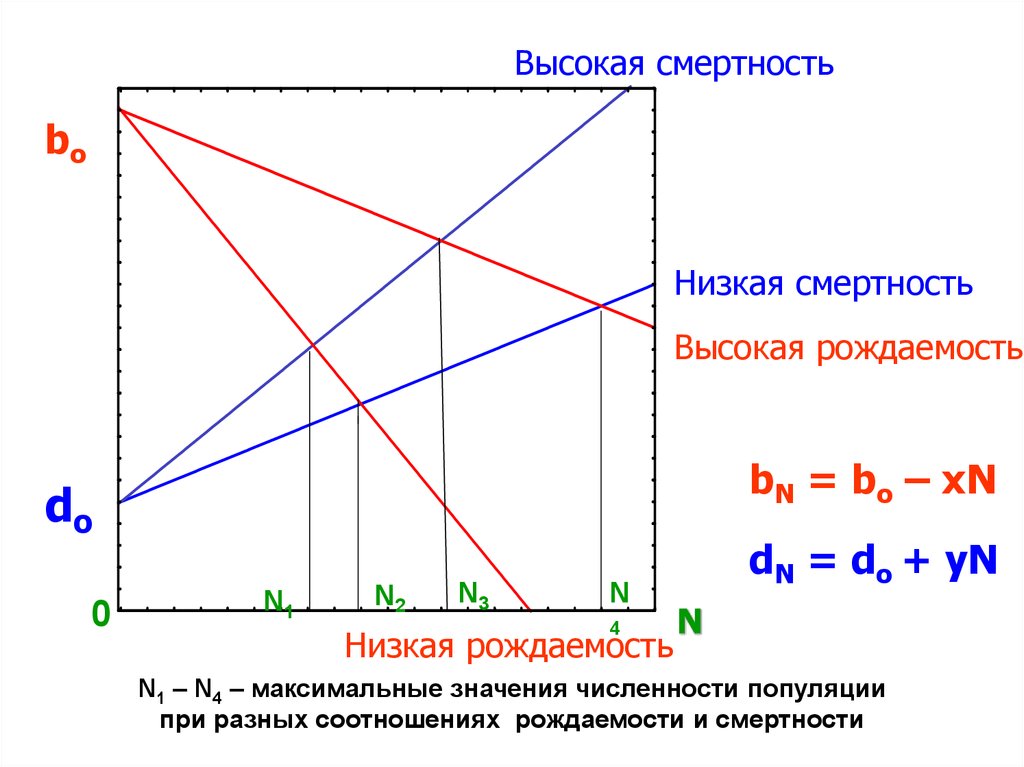
46. Сопряженные изменения рождаемости и смертности
Рассмотрим простейший случай, когдаудельная скорость смертности (d) линейно
возрастает,
а
удельная
скорость
рождаемости (b) линейно снижается с
увеличением плотности популяции (N).
47.
Высокая смертностьbo
Низкая смертность
Высокая рождаемость
bN = bo – xN
do
0
N1
N2
N3
N
4
Низкая рождаемость
dN = do + yN
N
N1 – N4 – максимальные значения численности популяции
при разных соотношениях рождаемости и смертности
48.
ТогдаbN = bo – xN;
dN = do + yN
bN и dN –удельные скорости рождаемости и
смертности при плотности N,
bo и do их значения при N, близкой к нулю,
х и у – эмпирические коэффициенты.
49.
Когда bo = do, что соответствует точкепересечения прямых) на графике, скорость роста
численности популяции
становится равной
нулю.
Отсюда популяция достигает максимально
возможной при данных условиях численности
(Nmax), или т.наз. «стационарного состояния».
Чем сильнее рождаемость снижается, а
смертность возрастает с увеличением плотности,
тем более низкими становятся значения Nmax
50.
При стационарном состоянии популяции,т.е. при bN=dN
соблюдается следующее
соотношение:
bo – xN = do + yN
Отсюда
Nmax = (bo – do)/(x + y)
Тогда плотность популяции может
изменяться в следующих пределах:
0 ≤ N ≤ Nmax
51.
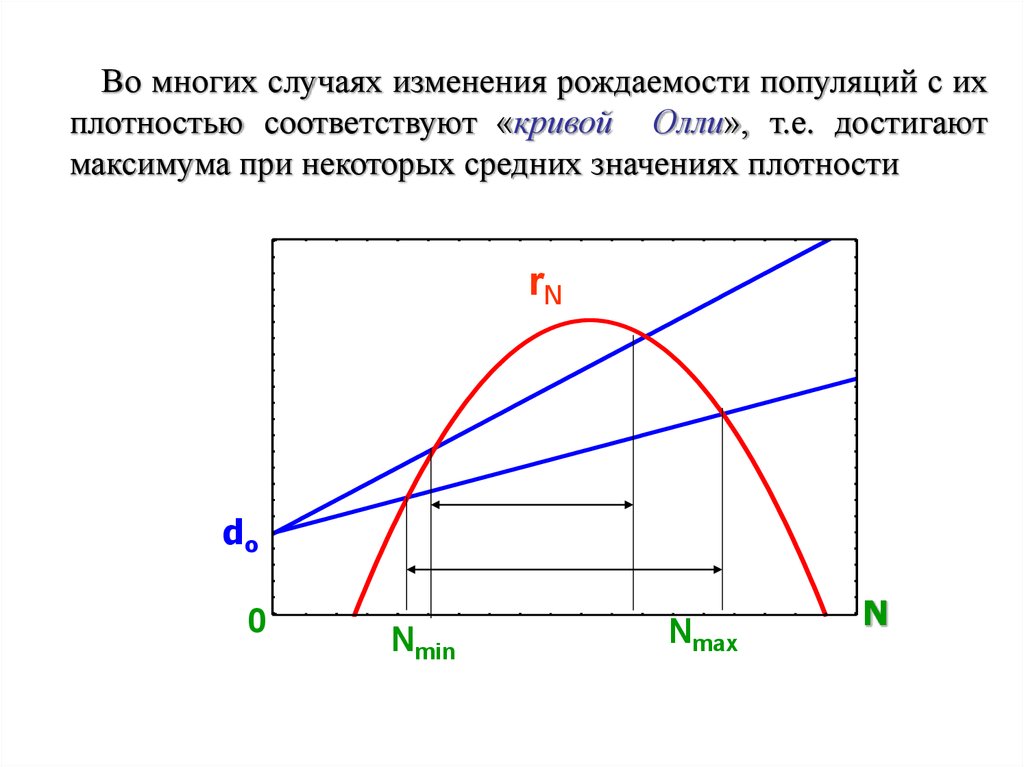
Во многих случаях изменения рождаемости популяций с ихплотностью соответствуют «кривой Олли», т.е. достигают
максимума при некоторых средних значениях плотности
rN
do
0
Nmin
Nmax
N
52.
В таком случае существует не толькомаксимальный (Nmax), но и минимальный
(Nmin) предел численности популяций.
Отсюда ее плотность может изменяться в
пределах:
Nmin ≤ N ≤ Nmax.
Чем выше смертность в популяции, тем в
более узких пределах будет изменяться ее
численность.
53.
Рост численности популяцийРостом
популяции
называется
увеличение
численности составляющих ее особей во времени в
результате процессов размножения или иммиграции.
При этом размеры, масса или возраст особей не имеют
значения.
Перед
рассмотрением
количественных
закономерностей роста популяций необходимо выделить
следующие основные понятия.
Абсолютная скорость роста популяции (С), или
прирост ее численности за (ΔN) за единицу времени (Δt),
т.е.:
С=
54.
Значениеимеет размерность
[особи·время-1].
Когда значения Δt достаточно малы,
абсолютный
прирост
характеризует
мгновенную скорость роста популяции, т. е.
или первую производную численности
по времени.
55.
Относительная скорость роста популяции (С’)определяется как
прирост численности за единицу
времени в расчете на одну особь:
_
_
где N – средняя численность популяции за период Δt.
Значения С’ имеют размерность [время-1].
Когда Δt значения достаточно малы, получаем
удельную скорость роста численности популяции (r):
56.
Удельную скорость роста популяции можнорассчитать согласно:
где N1 и N2 – численность популяции в моменты
времени t1 и t2.
Значения r имеют размерность [время-1].
Если значения r в период времени t1 – t2 остаются
постоянными, то рост численности популяции в
этом интервале является экспоненциальным.
57.
В таком случае абсолютная скорость ростапопуляции возрастает прямо пропорционально ее
достигнутой численности:
Исходя из
,
получаем:
Интегрированием последнего уравнения получаем
уравнение экспоненциального роста:
Nt = Noert,
где No – начальная численность особей,
Nt – численность особей ко времени t.
58.
В полулогарифмических координатахграфик этого уравнения
представляет
прямую линию:
lnNt = lnNo + rt
59. Примеры экспоненциального роста популяции от No = 10 экз при трех разных значениях r. Слева – обычные координаты, справа –
N, экзr = 0,6
4000
4000
r = 0,4
800
3000
400
2000
r = 0,2
80
40
1000
8
4
0
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
Время, условные единицы
Примеры экспоненциального роста популяции от No = 10 экз
при трех разных значениях r.
Слева – обычные координаты,
справа – полулогарифмические координаты
60.
Уравнениеэкспоненциально
роста
можно получить и другим способом.
Если значения удельных скоростей
рождаемости (b) и смертности (d) в
популяции остаются постоянными, то:
Nt = Noebt - Noe-dt = Noe(b – d)t
Поскольку b и d - постоянные величины,
их разность b - d = r представляет собой
удельную
скорость роста численности
популяции.
61.
Вприродных
условиях
экспоненциальный рост встречается очень
редко. Его примерами являются молодые
пионерные популяции, которые попадают в
условия избытка пространства и ресурсов,
отсутствия хищников, паразитов
и
конкурентов.
Поэтому модель экспоненциального роста
рассматривают
в
качестве
оценки
потенциальных возможностей увеличения
численности популяции.
62.
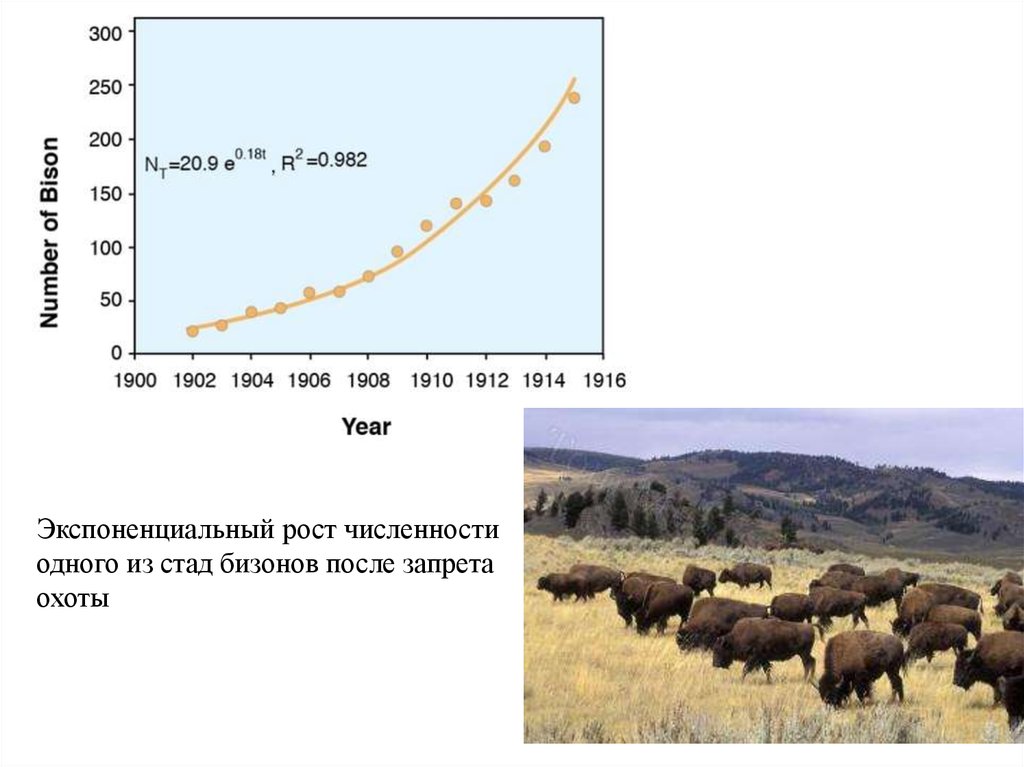
Экспоненциальный рост численностиодного из стад бизонов после запрета
охоты
63.
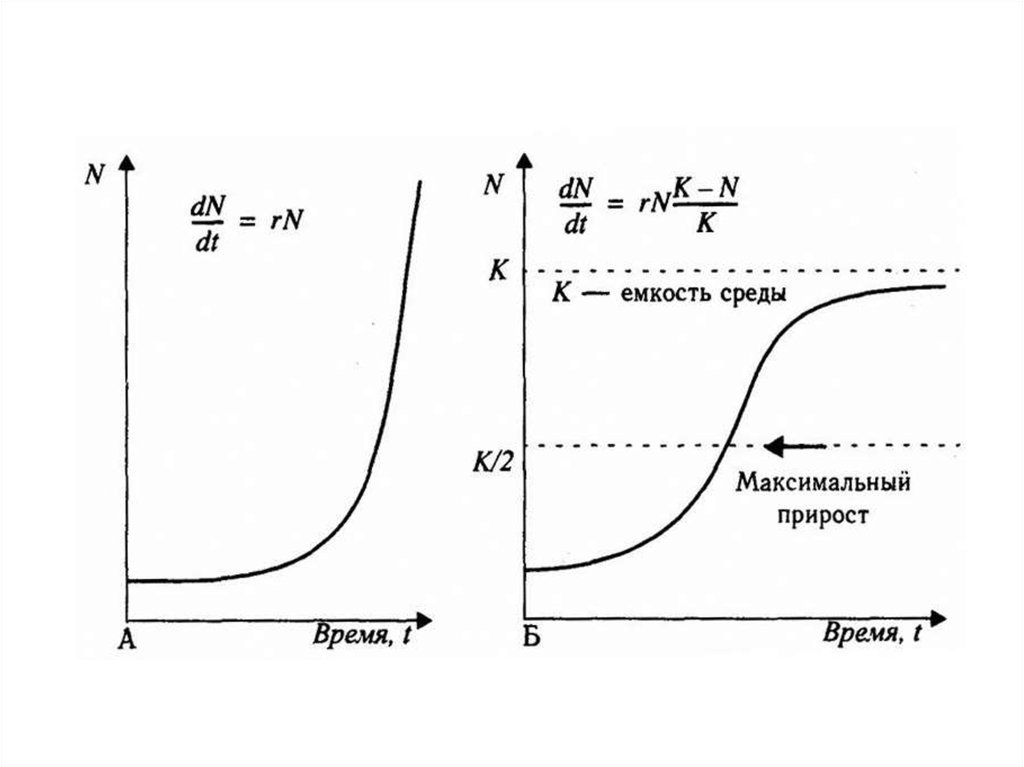
Логистический рост численности популяцииВ природе, как правило, с ростом численности
популяции скорость ее роста снижаться и в
конечном итоге падает до нуля. Это приводит к
стабилизации
численности
популяции
на
определенном уровне.
64. Рассмотрим простейший случай, когда удельная скорость роста популяции (r) линейно снижается с увеличением ее численности (N).
65.
Тогда связь между r и N выражается уравнениемпрямолинейной регрессии:
rN = r - aN
где rN – удельная скорость роста популяции при
плотности N,
r – ее максимальное значение при N, близкой к
нулю,
а – эмпирический коэффициент.
При некоторой численности популяции (N = К)
значение rN становится равным нулю. Тогда:
r – aK = 0, или
a = r/K
66.
Подставляя a = r/K в уравнениеrN = r - aN
получаем:
rN = r –
или
отсюда:
2
67.
68.
Преобразуем уравнение:Получаем:
Значение
= Z является константой. Тогда:
- ZN2
Параметр ZN2 характеризует сопротивление среды
росту численности популяции.
69.
При низких N («экологический вакуум»)значения zN2
близки к нулю, отсюда рост
численности популяции практически не будет
отличаться от экспоненциального.
С повышением численности значения zN2
будут возрастать, поэтому рост численности будет
замедляться и все более отличаться от
экспоненциального.
При некотором значении N = K скорость
роста популяции станет равной нулю, т.е.
популяция
достигнет
своей
максимальной
численности (Nmax).
70.
Интегрирование уравненияпозволяет получить уравнение логистического роста:
где Nt – численность популяции к моменту времени t; К – ее
максимальное значение (Nmax); r – удельная скорость роста
популяции при минимальной плотности; а – «константа
интегрирования»,
определяющая
положение
кривой
относительно начала координат.
, где No – численность особей при t = 0.
71.
Графиклогистического
уравнения
представляет
S-образную
кривую,
асимптотически
приближающуюся
к
предельному значению плотности популяции
К = Nmax.
Такой тип роста численности популяции
называется логистическим ростом.
72.
Пример расчета уравнения логистического ростадля лабораторной популяции дрозофилы
Возраст
популяции, сутки
Число особей
Возраст
популяции, сутки
Число особей
0
12
20
1556
2
20
22
1804
4
33
24
1957
6
56
26
2056
8
93
28
2104
10
155
30
2115
12
258
32
2123
14
431
34
2131
16
720
36
2133
18
1203
38
2133
73.
t = 0 cуток, N = 12 особей;t = 2 cуток, N = 20 особей.
Тогда
r = (ln20–ln12)/(2-0) = 0,2554 сут-1
Поскольку K = Nmax = 2133 особи, то
а = ln[(2133 – 12)/12] = 5,175
Тогда
74. Рост численности популяции дрозофилы
2400Численность особей
2000
1600
1200
800
400
0
0
5
10
15
20
25
Время, сутки
30
35
40
1
2
Видно, что экспоненциальная фаза роста
популяции
дрозофилы (1) закончилась на 18-е сутки роста, после чего
началось быстрое снижение скорости роста, приведшее к
стабилизации ее численности.
75.
Емкость средыКогда
численность
популяции
стабилизируется на определенном уровне, говорят,
что для нее достигнут предел емкости среды.
Емкость среды – способность природного
окружения
обеспечивать
нормальную
жизнедеятельность в единице пространства
определенного числа особей определенной массы
без заметного нарушения самого окружения.
76.
Емкость среды проявляет свое действиечерез лимитирующие факторы – запасов
пищи, минеральных солей, пространства,
влияние особей друг на друга в результате
повышения плотности.
Поддержание
численности
на
определенном
уровне
происходит
посредством механизмов отрицательной
обратной связи.