")






")




. А - плоская нейральная пластинка. B- нейральная пластинка (с подлежащей хордой -")



 у куриного зародыша. Образование медуллярного тяжа из которого")
")



 и антериорной висцеральной энтодермы (AVE) в формирование головы и головного")
. Стадии трёх и")

 гены контролируют спецификацию в передне-заднем направлении заднего отдела головного (ромбомеры)")




")



 у Drosophila вдоль дорсо-вентральной и")
 у Drosophila вдоль дорсо-вентральной и")


 Гены кодируют ТФ со структурой bHLH, но не проявляют большой")


")




. Отметить эпендимный, маргинальный, мантийный")
 и крыши (BMP) нервной трубки под воздействием одноимённых")

. Становление градиентов паракринных факторов в нервной трубке в")
. Экспрессия Shh (в нижней")
, инициированная участком хорды")
 и сигнальных")
")



 нейронов (спецификация LIM-содержащих нейронов спинного мозга.")
")

 – эволюционно появляется у позвоночных. В эмбриогенезе формируется дорсолатерально в передне-заднем")
")



")










 - мембранные или секреторные белки ряда (эпителиальных) клеток ограничивают рост аксонов в")




 Биология
БиологияПохожие презентации:










Нейрогенез (у высших животных: ЦНС - нервная трубка периферическая, НС - нервный гребень и плакоды)
1. Лекция 12 НЕЙРОГЕНЕЗ (у высших животных: ЦНС- нервная трубка периферическая НС - нервный гребень и плакоды)
Лекция 12НЕЙРОГЕНЕЗ
(у высших животных: ЦНСнервная трубка
периферическая НС - нервный
гребень и плакоды)
2. Основные производные эктодермального зародышевого листка позвоночных
3. Основные составлящие нервной системы
4. Нейроны – биполярные клетки, аксон растет от тела клетки
5. Глиальные клетки позвоночных изолируют аксоны, содержат миелиновые мебраны, обеспечивая передачу электрических импульсов от
тела нейрона к аксону6.
• В течение развития нейральныестволовые клетки дают начало
нейронам ЦНС
• Они также – источник всех типов
макроглии ЦНС (астроцитов,
олигодендроцитов)
7. НЕЙРУЛЯЦИЯ 1. Начальная стадия нейрогенеза у позвоночных - нейруляция. У позвоночных участок эктодермы специфицируется как
НЕЙРУЛЯЦИЯ1. Начальная стадия нейрогенеза у позвоночных нейруляция.
У позвоночных участок эктодермы
специфицируется как нейроэктодерма и клетки
этого участка приобретают вид столбчатого
эпителия.
Этот район называется нейральной пластинкой.
Затем инвагинацией и замыканием нейральной
пластинки формируется нервная трубка.
Различают первичную и вторичную нейруляцию.
2. Дифференцировка нервной трубки в переднезаднем и дорсо-вентральном направлениях
3. Образование нервного гребня
8. Нейруляция у амфибий (первичная)
9. Нейроглия дрозофилы Edenfeld et al. 2005, Curr.Opin. Neuribiol. 15, 34-39
10. Виды нейроглии у насекомых Freeman, Doherty, 2006, Trends in Neurosci. 29, 82-90
11. Хотя миелиновая оболочка глии у многих беспозвоночных отсутствует, у некоторых беспозвоночных она встречается и отличается от
таковой позвоночных, причём биохимическаяструктура этого миелина варьирует. Вероятно, первичная функция
миелина глиальных клеток животных, окружающая аксоны изоляция последних от возможных контактов с клетками других
структур.
12. Нейруляция первичная и вторичная Роль вертикальных и планарных сигналов в индукции ЦНС Градиенты факторов, ответственных за
спецификацию структурвдоль передне-задней и дорсо-вентральной осей у зародыша
амфибии ( По Gilbert, 2003; модифицированная схема)
13. Нейруляция у куриного зародыша (вид сверху). А - плоская нейральная пластинка. B- нейральная пластинка (с подлежащей хордой -
зачатокголовы), С- нервная бороздка, D- начало формирования нервной трубки в
головном отделе, Е- головные и спинные отделы нервной трубки
14. Первичная нейруляция у куриного зародыша. Изгибание нейральной пластинки в районах контакта с хордой и эктодермой. Замыкание
нервных валиков и образование трубки, образование нервного гребня15. Окончание нейруляции в головном отделе куриного зародыша 24-х часов развития. В каудальном отделе ещё происходит гаструляция
16. Особенности гаструляции у млекопитающих. Формирующаяся хорда у зародыша мыши контактирует с энтодермальными клетками первичной
кишки (и, возможно, c пластинкой дна нервной трубки). Реснички клетокузелка (слева) важны для направления миграции клеток
17. Вторичная нейруляция в хвостовом районе (после 25-ого сомита) у куриного зародыша. Образование медуллярного тяжа из которого
путёмкавитации образуется нервная трубка
18. Нейруляция у человека (формирование нервной трубки)
19. Нейруляция у человека. Важность своевременного закрытия переднего и заднего нейропоров для нормального развития. Дефекты,
связанные с нарушением закрытия нейропоров в разныхрайонах -(anencephaly, spina bifida) и замыкания всей нервной трубки
(craniorachischisis)
20. Смена типов кадгерина в клетках эктодермы в ходе нейруляции обусловливает образование нервной трубки и нервного гребня
21. Основные производные эктодермального зародышевого листка позвоночных
22. Участие двух сигнальных центров: узелка (node) и антериорной висцеральной энтодермы (AVE) в формирование головы и головного
мозга млекопитающих. AVE экспрессирует гены – ТФ: Lim1, Otx2,Hex и секретируемых факторов: Cerberus, Dickkopf. Узелок – гены
секретируемых факторов: хордина, Noggin и др.
23. Региональная специфичность нервной трубки и её производных вдоль передне-задней оси (развитие мозговых структур). Стадии трёх и
пяти мозговых пузырей и т.д.24. Головной мозг позвоночных
EmbryonicForebrain
Midbrain
Hindbrain
Shark
Goose
Human
25. Гомеотические (гомеозисные) гены контролируют спецификацию в передне-заднем направлении заднего отдела головного (ромбомеры)
мозга и всего спинного мозга мыши26. Факторы, участвующие в закладке различных отделов головного и спинного мозга млекопитающих
27. Участие гомеотических генов в спецификации нейральных структур вдоль передне-задней оси позвоночных и дрозофилы (ортологичные
гены отмечены одинаковым цветом)28. Экспериментальное увеличение концентрации ретиноевой кислоты в заднем отделе головного мозга вызывает превращение ромбомеров
одного вида в другой29. Образование различных слоев нервной трубки и образование различных отделов мозга в ходе нейрогенеза
30. Пролиферация и миграция дифференцирующихся нейробластов в ЦНС млекопитающих (будущие головной и спинной отделы)
31. Градиент «изнутри наружу» миграции нейронов при образовании слоев коры переднего отдела мозга макаки-резус
32. Миграция нейрона из вентрикулярного слоя в один из верхних слоев по отростку глиальной клетки. А-схема, B- адгезия нейрона к
отростку глиальной клетки. Внизу (С) миграция нейрона в мозжечкемлекопитающего
33.
• Гены, ответственные за нейрогенез убеспозвоночных и позвоночных – весьма
консервативны (ортологичные гены)
• Пронейральные гены
• Нейрогенные гены
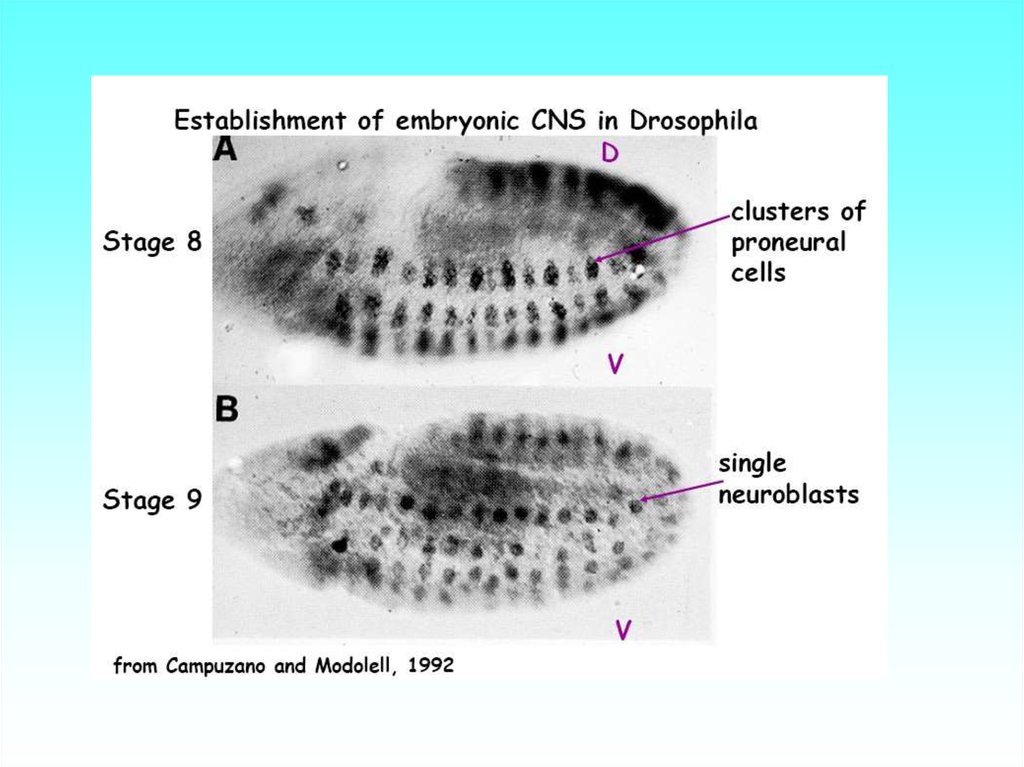
34. Последовательная спецификация линии нейробластов (в виде кластеров клеток) у Drosophila вдоль дорсо-вентральной и
передне-задней осей. За нейральнуюспецификацию - (А) у дрозофилы в ходе гаструляции отвечают факторы Dpp и Sog
(decapentaplegic и short gastrulation). Они гомологичны факторам позвоночных
соответственно BMP и chordin. Результатом их взаимодействия является активация
пронейральных basicHLH- транскрипционных факторов achaete, scute (у
млекопитающих гомолог -MASH-1,2 )
35. Последовательная спецификация линии нейробластов (в виде кластеров клеток) у Drosophila вдоль дорсо-вентральной и
передне-задней осей(продолжение).
Delta/Notch-сигналинг определяет дифференцировку пронейральных клеток
на нейральные и и ненейральные (эктодермальные).
36.
37.
38.
• Пронейральные гена дрозофилыкодируют белки-ТФ с характерным
доменом: basic- Helix-Loop-Helix
• 4 гена A-S –Complex
• Нейрогенные гены: (Atonal-семейства)то же ТФ имеют bHLH-структуру, но
лишь на 50% гомологичны первым
39. Гены нейрональной дифференцировки дрозофилы (biparous/tap) Гены кодируют ТФ со структурой bHLH, но не проявляют большой
гомологии спервыми двумя семействами ПГ и
действуют на более поздних этапах
нейрогенеза (в ходе формирования
нейронов).
Сходны с геном нейрогенина позвоночных
40. Пронейральные гены дрозофилы семейства achaete/scute
41. Часть нейрогенных генов дрозофилы могут экспрессироваться в ненейронных клетках и тогда их белковые продукты действуют как
антогонисты ТФ –продуктампронейральных генов
• Эти гены кодируют белки, аналогичные белкам пронейральных
генов. Они являются ТФ, со структурой двух типов:
• HLH (extamacrochaeta –emc), не имеющего основного домена
связывания ДНК
• basicHLH “Hairy -семейства” ( hairy, E(spl)-C, deadpan).
Вместо E-бокса эти гены содержат N-бокс, кодирующий
основный (basic) домен другого типа
Оба эти обстоятельства не дают возможности их гетеродимерам с
белками пронейральных генов нормально функционировать в
качестве положительных ТФ.
42. Функции транскрипционных факторов и сигнальных молекул беспозвоночных, ортологичных генам позвоночных (см. ранее)
43.
• Спецификация идифференцировка клеток
спинного мозга
44. Нейруляция первичная и вторичная Роль вертикальных и планарных сигналов в индукции ЦНС Градиенты факторов, ответственных за
спецификацию структурвдоль передне-задней и дорсо-вентральной осей у зародыша
амфибии (S. Gilbert, 2003, модифицированная схема)
45. Осевое паттернирование нервной трубки. Указаны паракринные факторы, принимающие участие в нейруляции вдоль передне-задней оси
зародыша.46. Участие паракринных факторов в дорсо-вентральном паттернировании нервной трубки- последовательность индуцирующих воздействий
хорды и эпидермиса на нейральнуюпластинку (Sonic hedgehog- Shh, факторы семейства BMP)
47. Развитие спинного мозга человека. Дифференцировка слоев мозга (поперечный срез). Отметить эпендимный, маргинальный, мантийный
слои,silcus limitans, расположение различных типов нейронов
48. Образование новых сигнальных центров в пластинке дна (Shh) и крыши (BMP) нервной трубки под воздействием одноимённых
индуцирующих сигналов из хордыи эктодермы. Гипотетически, соответствующие участки нервной трубки высших
позвночных могли произойти из примыкающих структур хордомезодермы и
эктодермы
49. Поляризация нервной трубки доль дорсо-вентральной оси
50. Дорсо-вентральная спецификация нервной трубки (продолжение). Становление градиентов паракринных факторов в нервной трубке в
дорсовентральном направлении, определяющие дифференцировку нейронов(мотонейронов, вставочных) и глии
51. Дорсо-вентральная спецификация нервной трубки (поперечный срез нервной трубки куриного зародыша). Экспрессия Shh (в нижней
части) -маркированазелёным цветом, экспрессия дорcалина (в верхней части) -маркирована
голубым цветом. Моторные нейроны, дифференцирующиеся при умеренных
концентрациях Shh, окрашены в желто-оранжевый цвет.
52. Каскадная индукция в вентральной части нервной трубки (вентральных V3 и моторных нейронов), инициированная участком хорды
прилежащим кнервной трубке. Эктопическая индукция моторных нейронов в срединной
части нервной трубки участками хорды, пластинки дна нервной трубки и
т.п.районов, секретирующих Shh.
53. Региональная активация в нервной трубке различных факторов транскрипции (Pax3,6,7; Mash1,2, Lim1,2,3; Isl1,2) и сигнальных
молекул (Delta, Serrate, Notch) определяющих спецификациюнейральных клеток в дорсо-вентральном направлении
54. Поляризация нервной трубки вдоль передне-задней оси (продолжение)
55. Поляризация нервной трубки вдоль проксимо-дистальной оси
56. Семейство факторов транскрипции LIM, включающих подсемейство Islet Lim transcription factors are responsive for axon target
selection during motor neuron axon guidance57. Спецификация различных групп двигательных нейронов в передне-заднем и дорсо-вентральном направлениях в зависимости от активации
в нихразличных транскрипционных факторов LIM-семейства (Lim1,3; Isl1,2).
Иннервация моторными нейронами различных мышц определяется
позициями моторных нейронов в спинном мозге (на разных уровнях вдоль
передне-задней оси и в колонках на одном уровне)
58. Организация двигательных (моторных) нейронов (спецификация LIM-содержащих нейронов спинного мозга.
59. Нервный гребень (только у позвоночных)
Клетки нервного гребня позвоночных –предшественники периферической
(парасимпатической и симпатической)
нервной системы и её глии (шванновских
клеток), но не только их.
60.
• Нервный гребень (НГ) – производное эмбриональнойэктодермы (существует временно), закладывается по
краям нейральной пластинки и окончательно
формируется при отделении нервной трубки от
эктодермы в ходе нейруляции. Клетки НГ мигрируют в
процессе или по завершении нейруляции и образуют
популяции клеток в различных тканях. Иногда клетки
НГ называют в виду их важности «четвёртым
зародышевым листком».
• Из клеток НГ формируются нейроны и глия ПНС,
некоторые элементы скелета челюстно-лицевого
скелета (кости, хрящи, связки, мышцы),
гладкомышечные клеткт ствола аорты и некоторых
артерий,хондроциты, меланоциты кожи,
разновидности гормон-продуцирующих клеток
некоторых желез внутренней секреции (надпочечников
и щитовидной железы).
61. Нервный гребень (НГ) – эволюционно появляется у позвоночных. В эмбриогенезе формируется дорсолатерально в передне-заднем
направлении и продуцирует клетки несколькихтипов в зависимости от различной локализации его отделов.
Головной отдел - производные НГ: хрящи, кости и соединительная
ткань головы, нейроны черепно-мозговых нервов и клетки глии.
Клетки НГ входят также в состав висцерального скелета
(одонтобласты, структуры челюсти, среднего уха), в состав тимуса.
Туловищный отдел НГ -меланоциты (синтезируют пигмент
меланин), мигрируют дорсолатерально в эктодерму или через
передний отдел склеротомов вентролатерально. Те клетки, которые
остаются в склеротоме, формируют ганглии дорсальных корешков
(содержат сенсорные нейроны). Мигрирующие вентрально образуют
мозговое вещество надпочечников, симпатические ганглии и
кластеры нервных клеток, окружающие аорту (вблизи сердца)
Клетки НГ крестцового отдела образуют парасимпатические
ганглии.
62. Производные клеток нервного гребня (НГ)
63. Миграция клеток нервного гребня в туловищном отделе куриного зародыша (два пути миграции у птиц и млекопитающих, но не у рыб и
лягушек). Дорсолатерально -меланоциты, заселяют кожу, волосяныелуковицы; вентрально- через передний отдел склеротомов, становятся
сенсорными и симпатическими нейронами, швановскими клетками,
хромафинными клетками.
64. Специфические маркеры НГ свидетельствуют о миграции клеток НГ куриного зародыша вентрально через антериорные отделы склеротомов
(роль эфрина- Ephrin и эфринового рецептора-Eph в постериорной частисклеротома, запрещающих миграцию в этом отделе путём отталкивания
мигрирующих клеток)
65. Роль белков RhoB и Slug в клетках НГ, способствующих миграции клеток. (Rho участвует в упрочении цитоскелета для облегчения
миграции, а Slug участвует в ослаблении прочных контактов междуклетками НГ)
66. Последовательное ограничение путей дифференцировки клеток НГ (гипотетическая модель)
67. Дифференцировка клеток НГ определяется последовательными индуцирующими воздействиями паракринных факторов FGF и NGF
(NGF-обусловливает миграцию клеток, действуя как хемоаттрактант икак ингибитор апоптоза)
68. Конечные стадии дифференцировки клеток нервного гребня туловищного отдела. Альтернативные варианты дифференцировки в
хромоафинные клеткимозгового вещества надпочечников и в симпатические нейроны.
69.
• Черепные плакоды –третий источник
нервных клеток (и не
только их)
70. Черепные плакоды - временные локальные утолщения эктодермы между эпидермальной и нейральной эктодермой головы. Формирование
плакод индуцируется соседними тканями.Преплакодный эпибранхиальный (подковообразный) зачаток образуется дорсально, там,
где жаберные карманы контактируют с эпидермисом. Эта структура затем расщепляется,
образуя парные зачатки обонятельных, хрусталиковых плакод, плакод ряда черепномозговых нервов, слуховых структур, непарный зачаток аденогипофиза. Эти структуры
впоследствии расщепляются, формируя плакоды коленчатую, лицевую и узловатую
плакоды, которые служат материалом для сенсорных нейронов лицевого, языкоглоточного в
блуждающего черепных нервов.
71. Черепные плакоды
72.
•Как растут аксоны?73. Схематическое изображение моторного нейрона. Электрические импульсы приходят от дендритов и передаются по аксону. Длина аксона
может быть больше метра. Конус роста аксона имеетдвигательный и сенсорный аппараты. По окончании движения он
образует синапс с клеткой ткани-мишени
74. Внутриклеточные причины роста аксонов: взаимодействия актиновых и миозиновых структур
75.
• Внешние факторы управлениядвижением конуса роста аксона
(проецирование его на тканимишени) в принципе те же, что и
при миграции клеток нервного
гребня (молекулы аттракции и
репульсии).
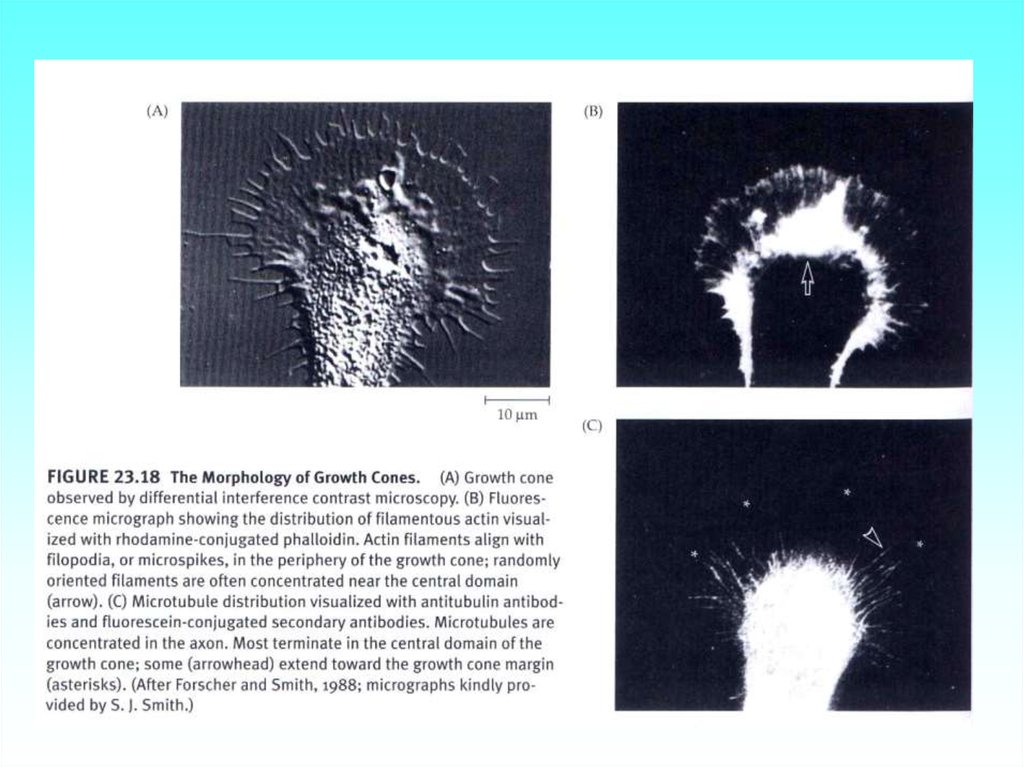
76. Конус роста аксона нейрональной клетки бражника. Видны микрошипики (филоподии, содержащие микрофиламенты, окрашены в зелёный
цвет, иориентированные радиально к аксону) и микротрубочки (окрашены красным
цветом) ориентированы параллельно к аксону).
77.
78. Семафорины (наряду с эфринами) - мембранные или секреторные белки ряда (эпителиальных) клеток ограничивают рост аксонов в
нежелательном направлении(репелленты). Внизу трансгенный (по семафорину) фибробласт куриного зародыша
избирательно угнетает выросты механорецепторного аксона. Наоборот , нетрины
являются хемоаттрактантами и привлекают к себе растущие аксоны.