


















 Биология
БиологияПохожие презентации:










Основные закономерности наследования признаков
1. Основные закономерности наследования признаков
2.
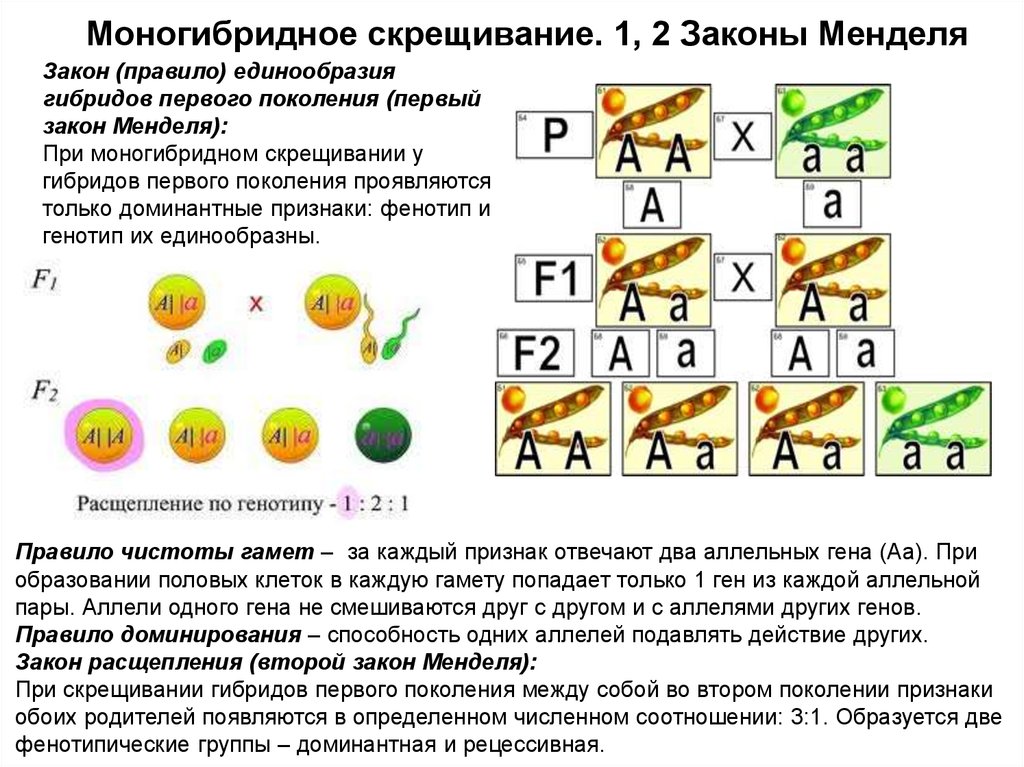
Моногибридное скрещивание. 1, 2 Законы МенделяЗакон (правило) единообразия
гибридов первого поколения (первый
закон Менделя):
При моногибридном скрещивании у
гибридов первого поколения проявляются
только доминантные признаки: фенотип и
генотип их единообразны.
Правило чистоты гамет – за каждый признак отвечают два аллельных гена (Аа). При
образовании половых клеток в каждую гамету попадает только 1 ген из каждой аллельной
пары. Аллели одного гена не смешиваются друг с другом и с аллелями других генов.
Правило доминирования – способность одних аллелей подавлять действие других.
Закон расщепления (второй закон Менделя):
При скрещивании гибридов первого поколения между собой во втором поколении признаки
обоих родителей появляются в определенном численном соотношении: 3:1. Образуется две
фенотипические группы – доминантная и рецессивная.
3. Неполное доминирование. Промежуточное наследование
4.
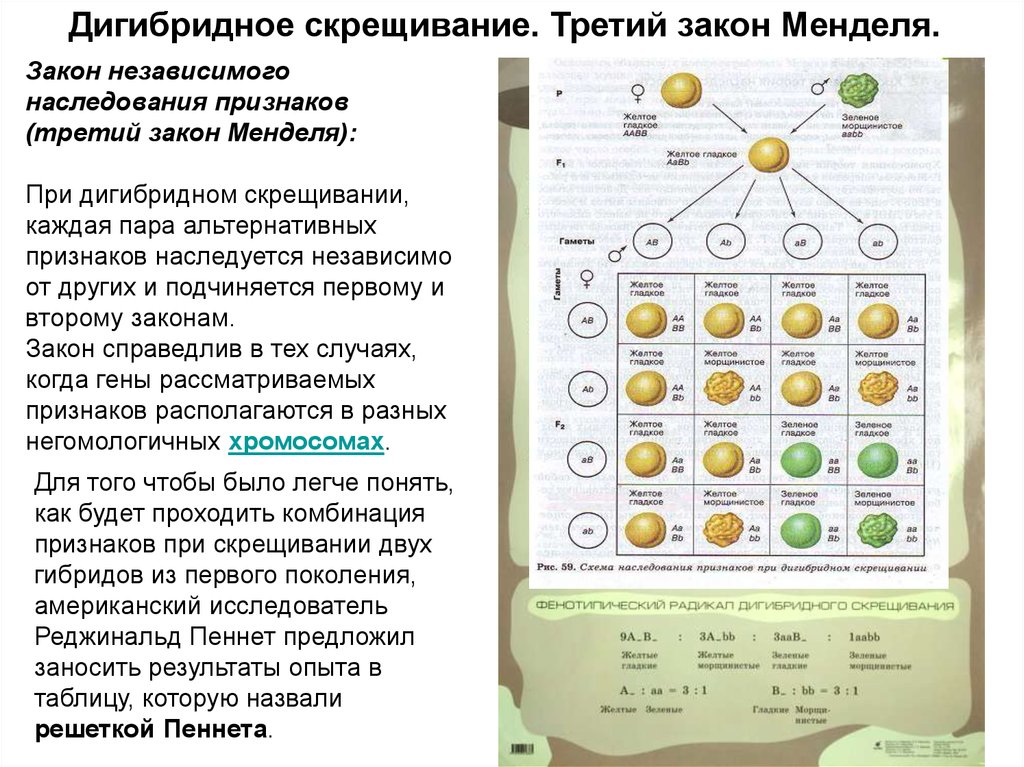
Дигибридное скрещивание. Третий закон Менделя.Закон независимого
наследования признаков
(третий закон Менделя):
При дигибридном скрещивании,
каждая пара альтернативных
признаков наследуется независимо
от других и подчиняется первому и
второму законам.
Закон справедлив в тех случаях,
когда гены рассматриваемых
признаков располагаются в разных
негомологичных хромосомах.
Для того чтобы было легче понять,
как будет проходить комбинация
признаков при скрещивании двух
гибридов из первого поколения,
американский исследователь
Реджинальд Пеннет предложил
заносить результаты опыта в
таблицу, которую назвали
решеткой Пеннета.
5.
6. Пример дигибридного наследования признаков
7. Анализирующее скрещивание
Анализирующее скрещивание - приполной доминантности среди особей с
доминантными признаками невозможно
отличить гомозиготы от гетерозигот, а в
этом часто возникает необходимость
(например, чтобы определить, чистородна
или гибридна данная особь). С этой целью
проводят анализирующее скрещивание,
при котором исследуемая особь
доминантными признаками скрещивается
с рецессивной гомозиготой. Если
потомство от такого скрещивания
окажется однородным, значит, особь
гомозиготна (ее генотип АА). Если же в
потомстве будет 50% особей с
доминантными признаками, а 50% с
рецессивными, значит, особь
гетерозиготна. Смотрите схему.
8. Пример анализирующего скрещивания
9. Анализирующее скрещивание при дигибридном наследовании признаков
10. Основные типы взаимодействия генов
* Полное доминирование:
Явление, при котором доминантный ген полностью подавляет
рецессивный. Гетерозиготные организмы по фенотипу всегда
точно соответствуют родителю, гомозиготному по доминантному
гену.
* Неполное доминирование:
Явление, при котором доминантный ген не полностью подавляет работу рецессивного, в результате
развивается промежуточный признак.
Примером может служить окраска цветка у растения ночная красавица. У которого при скрещивании
красных и белых цветков среди гибридов первого поколения все цветки розовые.
* Множественный аллелизм:
Явление, при котором ген представлен не двумя аллелями (как в случаях полного или неполного
доминирования), а гораздо большим их числом. Пример – трёхчленная серия аллелей, определяющая
окраску меха у кошек. Чёрная окраска меха B доминирует над шоколадной b, которая, в свою очередь,
доминирует над светло-коричневой b1. (B>b>b1).
* Кодоминирование:
Явление независимого друг от друга проявления обоих аллелей в фенотипе гетерозиготы, иными
словами – отсуствие доминантно-рецессивных отношений между аллелями. Пример – взаимодействие
аллелей (IA и IB), определяющих четвёртую группу крови человека.
* Сверхдоминирование:
более сильное проявление признака в гетерозиготе, а не в гомозиготе. Так, у дрозофилы имеется
рецессивный летальный ген, гетерозиготы по которому обладают большей жизнеспособностью, чем
доминантные гомозиготы.
11. Взаимодействие неаллельных генов:
• * Комплементарность (лат. – «комплиментум» - дополнение) – один извидов взаимодействия неаллельных генов, при котором эти гены
дополняют действие друг друга, и признак появляется только при
сочетании двух доминантных аллелей разных генов (например, А и В)
При этом каждый из взаимодействующих неаллельных генов в
отсутствии другого не обеспечивает формирования признака.
Классическим примером комплементарного
взаимодействия генов является наследование
формы гребня у кур. При скрещивании кур,
имеющих розовидный и гороховидный
гребень, все первое поколение имеет
ореховидный гребень.
При скрещивании гибридов первого поколения
у потомков наблюдается расщепление по
форме гребня: 9 ореховидных: 3 розовидных:
3 гороховидных: 1 листовидный. Генетический
анализ показал, что куры с розовидным
гребнем имеют генотип А_bb, с гороховидным
— ааВ_, с ореховидным — А_В_ и с
листовидным — ааbb, то есть развитие
розовидного гребня происходит в том случае,
если в генотипе имеется только один
доминантный ген — А, гороховидного —
наличие только гена В, сочетание генов А В
обусловливает появление ореховидного
гребня, а сочетание рецессивных аллелей
этих генов — листовидного.
12. При комплементарном взаимодействии генов в дигибридном скрещивании получаются расщепления потомков отличные от менделевского.
Однако все они являются видоизменениями общейменделевской формулы 9:3:3:1.
Расщепление в F2 признаков при
комплементарном взаимодействии
генов
9 : 7 Появление у душистого
горошка потомства, имеющего
цветки пурпурной окраски, при
скрещивании родительских форм с
белыми цветками.
9 : 6 : 1 Появление у тыквы
потомства с дисковидной формой
плода при скрещивании родителей,
имеющих сферическую форму
плода.
9 : 3 : 4 Появление у мышей
окраски шерсти типа агути при
скрещивании родительских форм
черного и белого цвета.
9 : 3 : 3 : 1 Появление у дрозофилы
потомства с темно-красными
глазами (дикого типа) при
скрещивании родительских форм с
ярко-красными (scarlet) и
коричневыми (brown) глазами.
П р и м е ч а н и е . Жирным
шрифтом выделен класс, признак
которого обусловлен
комплементарным
взаимодействием генов.
13.
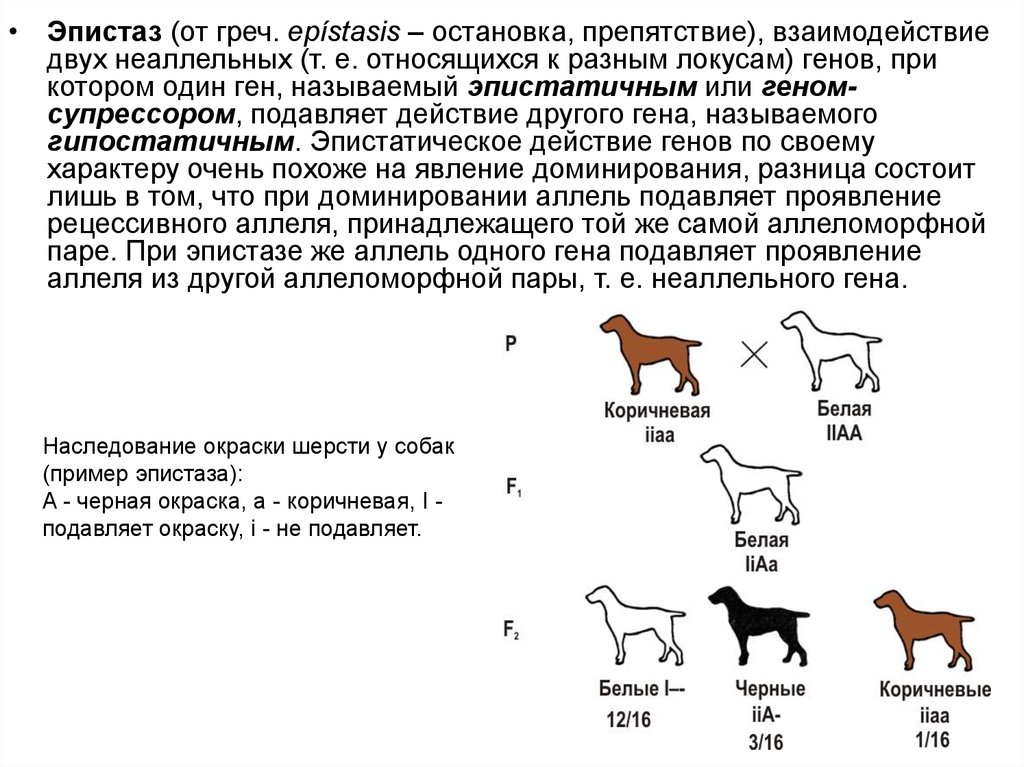
• Эпистаз (от греч. epístasis – остановка, препятствие), взаимодействиедвух неаллельных (т. е. относящихся к разным локусам) генов, при
котором один ген, называемый эпистатичным или геномсупрессором, подавляет действие другого гена, называемого
гипостатичным. Эпистатическое действие генов по своему
характеру очень похоже на явление доминирования, разница состоит
лишь в том, что при доминировании аллель подавляет проявление
рецессивного аллеля, принадлежащего той же самой аллеломорфной
паре. При эпистазе же аллель одного гена подавляет проявление
аллеля из другой аллеломорфной пары, т. е. неаллельного гена.
Наследование окраски шерсти у собак
(пример эпистаза):
A - черная окраска, а - коричневая, I подавляет окраску, i - не подавляет.
14.
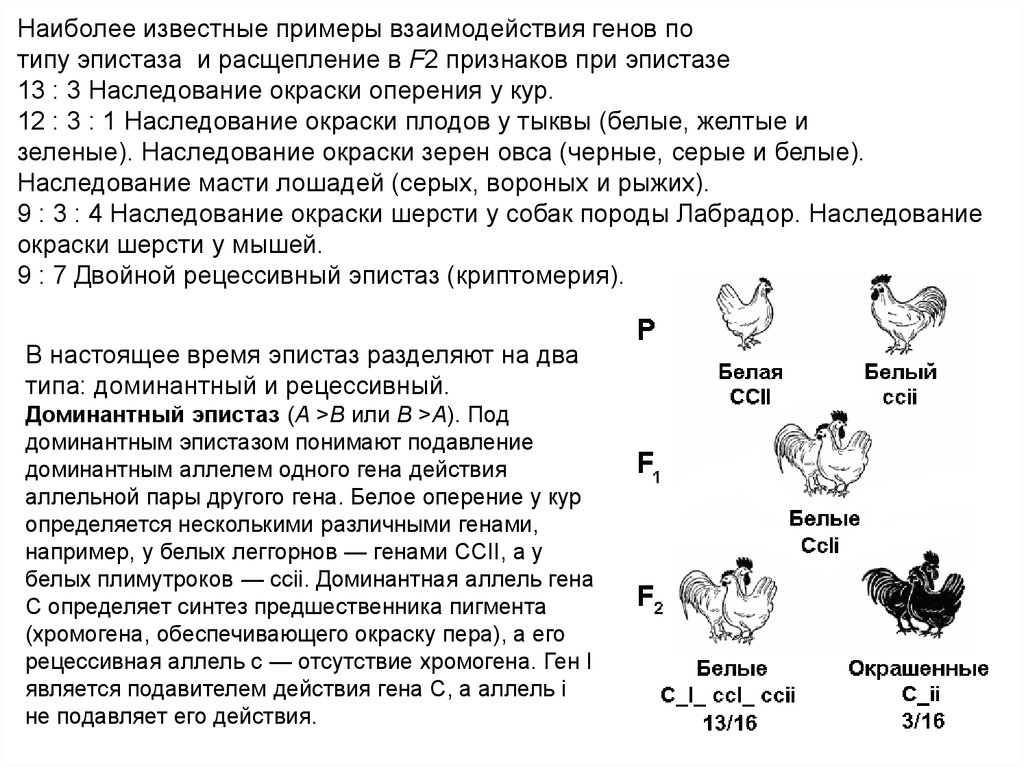
Наиболее известные примеры взаимодействия генов потипу эпистаза и расщепление в F2 признаков при эпистазе
13 : 3 Наследование окраски оперения у кур.
12 : 3 : 1 Наследование окраски плодов у тыквы (белые, желтые и
зеленые). Наследование окраски зерен овса (черные, серые и белые).
Наследование масти лошадей (серых, вороных и рыжих).
9 : 3 : 4 Наследование окраски шерсти у собак породы Лабрадор. Наследование
окраски шерсти у мышей.
9 : 7 Двойной рецессивный эпистаз (криптомерия).
В настоящее время эпистаз разделяют на два
типа: доминантный и рецессивный.
Доминантный эпистаз (А >В или В >А). Под
доминантным эпистазом понимают подавление
доминантным аллелем одного гена действия
аллельной пары другого гена. Белое оперение у кур
определяется несколькими различными генами,
например, у белых леггорнов — генами ССII, а у
белых плимутроков — ccii. Доминантная аллель гена
С определяет синтез предшественника пигмента
(хромогена, обеспечивающего окраску пера), а его
рецессивная аллель с — отсутствие хромогена. Ген I
является подавителем действия гена С, а аллель i
не подавляет его действия.
15.
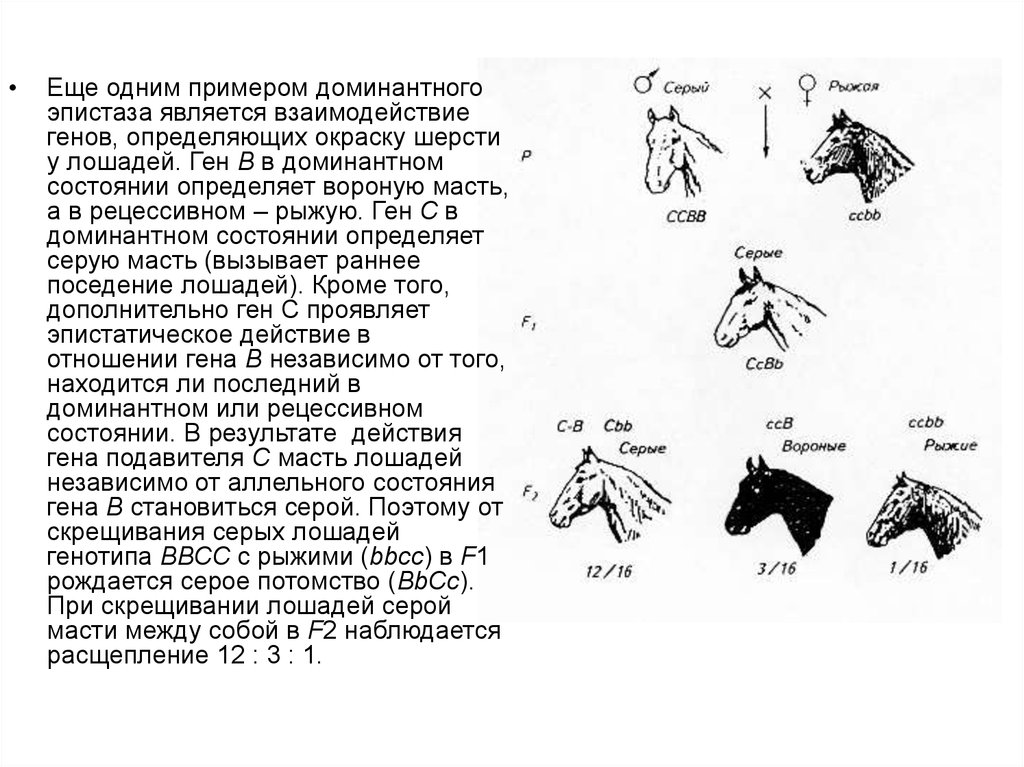
Еще одним примером доминантного
эпистаза является взаимодействие
генов, определяющих окраску шерсти
у лошадей. Ген В в доминантном
состоянии определяет вороную масть,
а в рецессивном – рыжую. Ген С в
доминантном состоянии определяет
серую масть (вызывает раннее
поседение лошадей). Кроме того,
дополнительно ген С проявляет
эпистатическое действие в
отношении гена В независимо от того,
находится ли последний в
доминантном или рецессивном
состоянии. В результате действия
гена подавителя С масть лошадей
независимо от аллельного состояния
гена В становиться серой. Поэтому от
скрещивания серых лошадей
генотипа ВВСС с рыжими (bbсс) в F1
рождается серое потомство (ВbСс).
При скрещивании лошадей серой
масти между собой в F2 наблюдается
расщепление 12 : 3 : 1.
16.
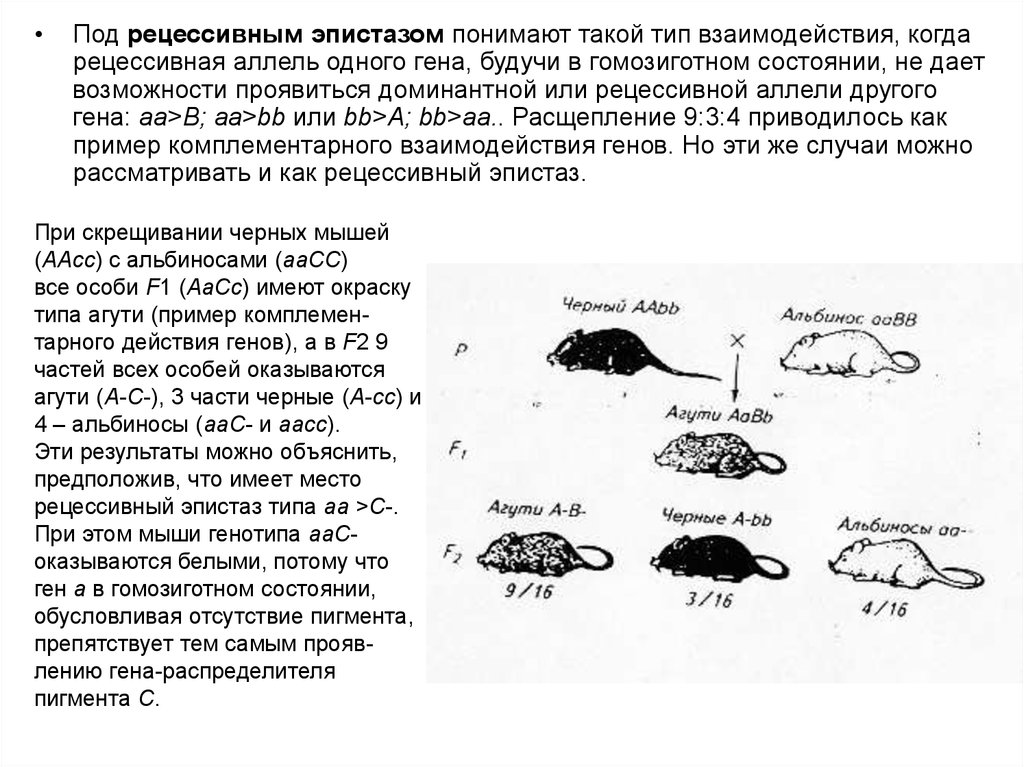
Под рецессивным эпистазом понимают такой тип взаимодействия, когда
рецессивная аллель одного гена, будучи в гомозиготном состоянии, не дает
возможности проявиться доминантной или рецессивной аллели другого
гена: аа>В; аа>bb или bb>А; bb>аа.. Расщепление 9:3:4 приводилось как
пример комплементарного взаимодействия генов. Но эти же случаи можно
рассматривать и как рецессивный эпистаз.
При скрещивании черных мышей
(AAсс) с альбиносами (ааСС)
все особи F1 (AaСс) имеют окраску
типа агути (пример комплементарного действия генов), а в F2 9
частей всех особей оказываются
агути (А-С-), 3 части черные (А-сс) и
4 – альбиносы (ааС- и aaсс).
Эти результаты можно объяснить,
предположив, что имеет место
рецессивный эпистаз типа аа >С-.
При этом мыши генотипа ааСоказываются белыми, потому что
ген а в гомозиготном состоянии,
обусловливая отсутствие пигмента,
препятствует тем самым проявлению гена-распределителя
пигмента С.
17.
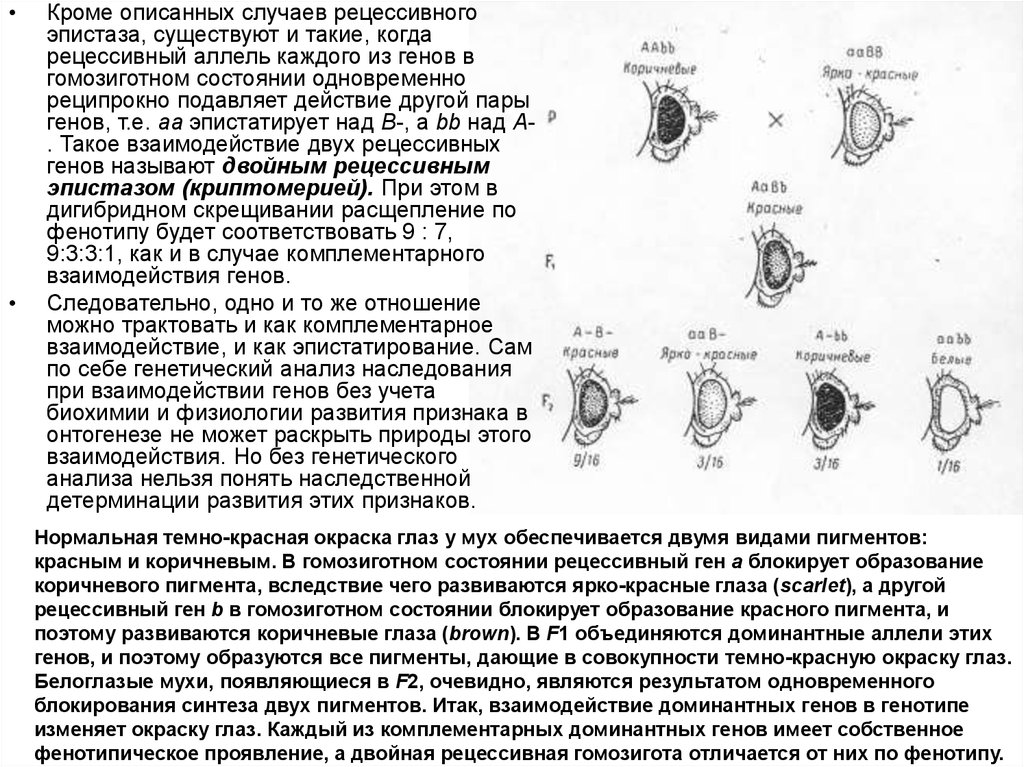
Кроме описанных случаев рецессивного
эпистаза, существуют и такие, когда
рецессивный аллель каждого из генов в
гомозиготном состоянии одновременно
реципрокно подавляет действие другой пары
генов, т.е. аа эпистатирует над В-, a bb над А. Такое взаимодействие двух рецессивных
генов называют двойным рецессивным
эпистазом (криптомерией). При этом в
дигибридном скрещивании расщепление по
фенотипу будет соответствовать 9 : 7,
9:3:3:1, как и в случае комплементарного
взаимодействия генов.
Следовательно, одно и то же отношение
можно трактовать и как комплементарное
взаимодействие, и как эпистатирование. Сам
по себе генетический анализ наследования
при взаимодействии генов без учета
биохимии и физиологии развития признака в
онтогенезе не может раскрыть природы этого
взаимодействия. Но без генетического
анализа нельзя понять наследственной
детерминации развития этих признаков.
Нормальная темно-красная окраска глаз у мух обеспечивается двумя видами пигментов:
красным и коричневым. В гомозиготном состоянии рецессивный ген а блокирует образование
коричневого пигмента, вследствие чего развиваются ярко-красные глаза (scarlet), а другой
рецессивный ген b в гомозиготном состоянии блокирует образование красного пигмента, и
поэтому развиваются коричневые глаза (brown). В F1 объединяются доминантные аллели этих
генов, и поэтому образуются все пигменты, дающие в совокупности темно-красную окраску глаз.
Белоглазые мухи, появляющиеся в F2, очевидно, являются результатом одновременного
блокирования синтеза двух пигментов. Итак, взаимодействие доминантных генов в генотипе
изменяет окраску глаз. Каждый из комплементарных доминантных генов имеет собственное
фенотипическое проявление, а двойная рецессивная гомозигота отличается от них по фенотипу.
18. Соотношение фенотипических классов дигибридного расщепления в F2 при различных типах взаимодействия генов
Взаимодействие междуаллелями
Генотипы
генами
Aa
Bb
Aи B
-
-
Отсутствует
-
+
Отсутствует
+
+
Отсутствует
9
3
+
+
аa маскирует Ви bb
9
3
+
+
А- маскирует Ви bb
12
3
1
+
{А- маскирует
В- и bb
bb маскирует Аи aa}
12(+1)?
3
1
+
{aa- маскирует
В- и bb
bb маскирует Аи aa}
+
{А- маскирует
В- и bb
B- маскирует Аи aa}
+
+
+
ААВВ
ААВb
1
AaBB
2
AaBb
2
3
AAbb
4
6
9
Aabb
aaBB
aaBb
aabb
1
2
1
2
1
1
2
3
1
3
1
4
7
15
1
Знак «–» – неполное доминирование, знак «+» – полное доминирование; знак вопроса около
фенотипических классов 12(+1) означает возможность расщепления как 12:3:1, так и 13:3.
19.
Полимерия
Полимерией называют однозначное влияние двух, трех и более неаллельных генов на
развитие одного и того же признака. Такие гены называют полимерными, или
множественными, и обозначают одной буквой с соответствующим индексом, например,
А1, А2, а1, а2. Полимерные гены контролируют большинство количественных признаков
организмов: высоту растения, массу семян, масличность семян, содержание сахара в
корнеплодах сахарной свеклы, удойность коров, яйценоскость, вес тела и т.д.
Кумулятивная полимерия.
В этом случае степень проявления признака зависит от
числа доминантных аллелей в генотипе особи. Так,
красная окраска зерен пшеницы определяется двумя и
более парами генов. Каждый из доминантных генов этих
аллелей определяет красную окраску, рецессивные гены
определяют белый цвет зерен. Один доминантный аллель
дает не очень сильно окрашенные зерна. Если в генотипе
присутствуют два доминантных аллеля, интенсивность
окраски повышается. Лишь в том случае, когда организм
оказывается гомозиготным по всем парам рецессивных
генов, зерна не окрашены.
20.
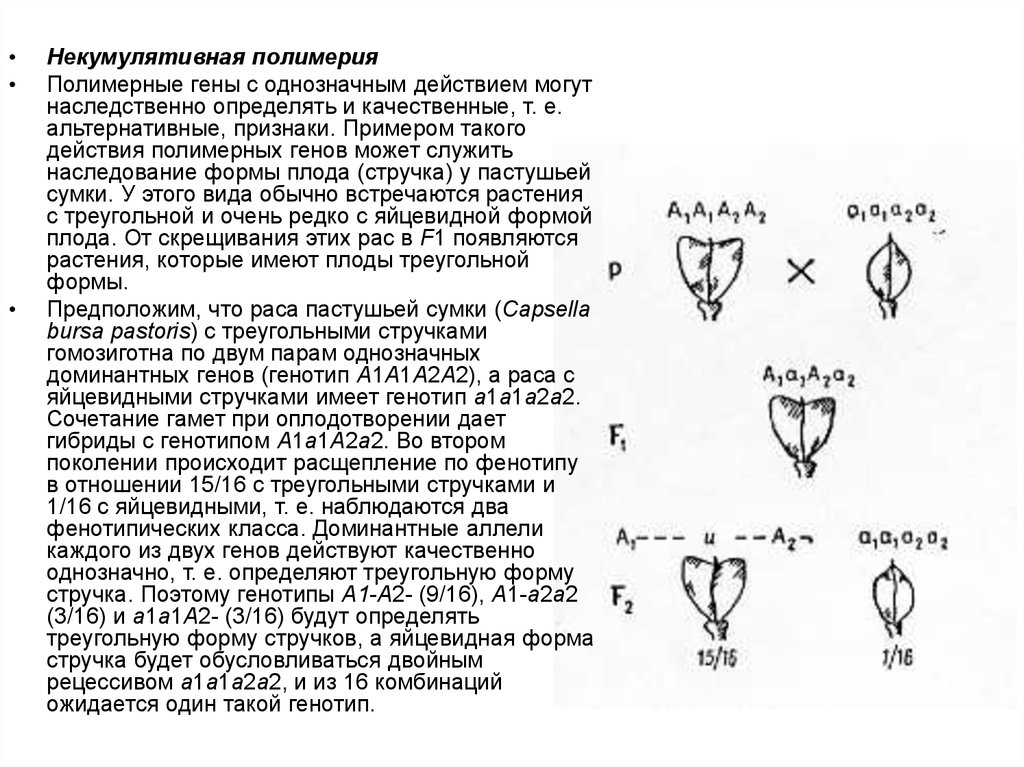
Некумулятивная полимерия
Полимерные гены с однозначным действием могут
наследственно определять и качественные, т. е.
альтернативные, признаки. Примером такого
действия полимерных генов может служить
наследование формы плода (стручка) у пастушьей
сумки. У этого вида обычно встречаются растения
с треугольной и очень редко с яйцевидной формой
плода. От скрещивания этих рас в F1 появляются
растения, которые имеют плоды треугольной
формы.
Предположим, что раса пастушьей сумки (Capsella
bursa pastoris) с треугольными стручками
гомозиготна по двум парам однозначных
доминантных генов (генотип А1А1А2А2), а раса с
яйцевидными стручками имеет генотип а1а1а2а2.
Сочетание гамет при оплодотворении дает
гибриды с генотипом А1а1А2а2. Во втором
поколении происходит расщепление по фенотипу
в отношении 15/16 с треугольными стручками и
1/16 с яйцевидными, т. е. наблюдаются два
фенотипических класса. Доминантные аллели
каждого из двух генов действуют качественно
однозначно, т. е. определяют треугольную форму
стручка. Поэтому генотипы А1-А2- (9/16), A1-a2a2
(3/16) и a1a1A2- (3/16) будут определять
треугольную форму стручков, а яйцевидная форма
стручка будет обусловливаться двойным
рецессивом а1а1а2а2, и из 16 комбинаций
ожидается один такой генотип.
21.
ДЕЙСТВИЕ ГЕНОВ-МОДИФИКАТОРОВ• Гены-модификаторы – это гены, влияющие на проявление
признаков (количественных или качественных), контролируемых
другими неаллельными генами.
Примером действия геновмодификаторов является
контроль за расположением
окрашенных участков шерсти
(пятен) на теле
некоторых животных. При
наличии доминантного гена S
(генотип SS или Ss) окраска
шерсти у мышей, морских
свинок, собак, лошадей и
других животных
равномерная и пятнистость
отсутствует. Если ген
находится в рецессивном
состоянии (генотип ss) –
пятна интенсивно проЯвляются.
22.
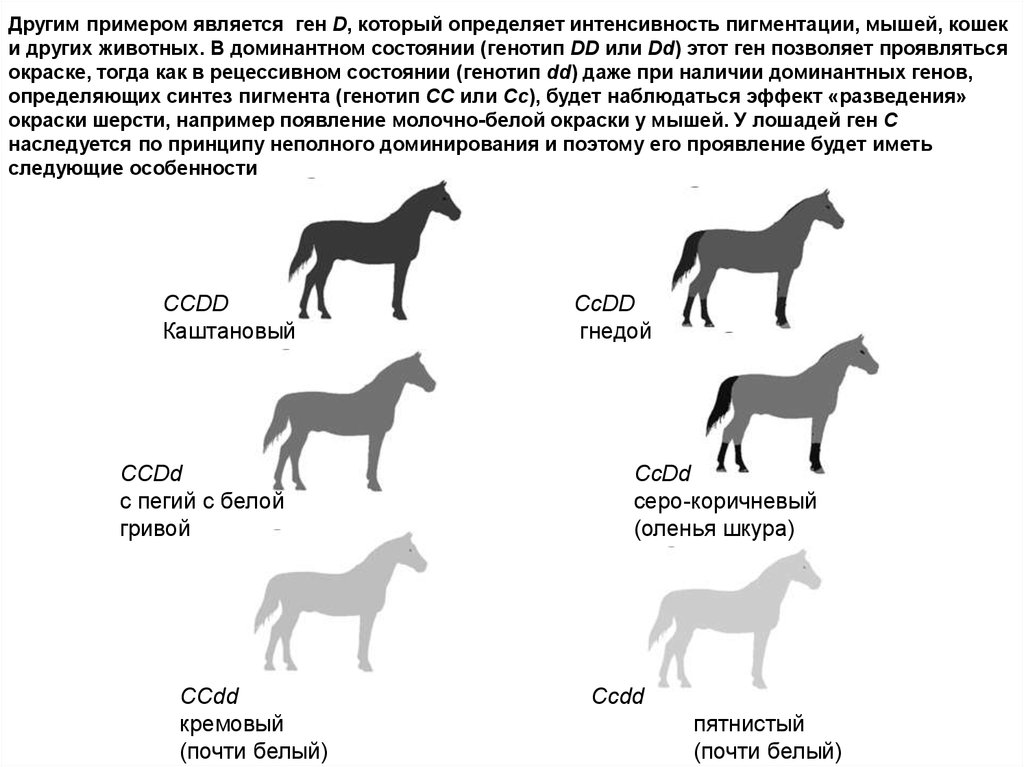
Другим примером является ген D, который определяет интенсивность пигментации, мышей, кошеки других животных. В доминантном состоянии (генотип DD или Dd) этот ген позволяет проявляться
окраске, тогда как в рецессивном состоянии (генотип dd) даже при наличии доминантных генов,
определяющих синтез пигмента (генотип СС или Сс), будет наблюдаться эффект «разведения»
окраски шерсти, например появление молочно-белой окраски у мышей. У лошадей ген С
наследуется по принципу неполного доминирования и поэтому его проявление будет иметь
следующие особенности
ССDD
Каштановый
CCDd
с пегий c белой
гривой
ССdd
кремовый
(почти белый)
CcDD
гнедой
CcDd
серо-коричневый
(оленья шкура)
Ccdd
пятнистый
(почти белый)
23.
Плейотропи́я (от греч. πλείων — «больше» и греч. τρέπειν — «поворачивать,
превращать») — явление множественного действия гена. Выражается в
способности одного гена влиять на несколько фенотипических признаков. Таким
образом, новая мутация в гене может оказать влияние на некоторые или все
связанные с этим геном признаки.
Летальное действие гена – одно из
разновидностей плейотропиии.
Один ген, определяющий какой-либо
признак, влияет так же на
жизнеспособность в целом.
При скрещивании лис с платиновой
окраской резко падала плодовитость.
Гены платиновой окраски в
гомозиготном состоянии вызывали
смертность плода на внутриутробной
стадии развития. Для избегания таких
ситуаций скрещивают платиновых лис с
чернобурыми.
Каракульские ягнята с серой окраской
(ширази) возникли в результате мутации
от овец с черной окраской. Гены серой
окраски в гомозиготном состоянии
вызывают гибель ягнят в раннем
возрасте. Для получения
жизнеспособных серых ягнят
скрещивают серых овец с черными.
Доминантные гомозиготы погибают в
период эмбрионального развития.
24. ВЛИЯНИЕ ФАКТОРОВ ВНЕШНЕЙ СРЕДЫ НА ПРОЯВЛЕНИЕ ПРИЗНАКОВ.
• Пенетрантность.Рассматривая действие гена, его аллелей,
необходимо учитывать не только генные
взаимодействия и действие генов-модификаторов, но
и модифицирующее действие среды, в которой
развивается организм. Известно, что у примулы
окраска цветка розовая (Р-) - белая (рр) наследуется
по моногибридной схеме, если растения развиваются
в интервале температур 15-25°С. Если же растения F2
вырастить при температуре 30-35°С, то все цветки у
них оказываются белыми. Наконец, при выращивании
растений F2 в условиях температуры, колеблющейся
около 30°С, можно получить разнообразные
соотношения от 3Р:1рр до 100% растений с белыми
цветками. Такое варьирующее соотношение классов
при расщеплении в зависимости от условий внешней
среды или от условий генотипической среды носит
название варьирующей пенетрантности: Это понятие
подразумевает возможность проявления или
непроявления признака у организмов, одинаковых по
исследуемым генотипическим факторам.
Триада рас китайских примул.
25.
Уже упоминался пример плейотропного действия гена - доминантная платиновая окраска
лисиц с рецессивным летальным действием. Как показал Д.К. Беляев с сотрудниками ,
можно добиться рождения живых щенков, гомозиготных по доминантной аллели
платиновой окраски, если варьировать длину дня для беременных самок. Таким образом,
пенетрантность проявления летального эффекта может быть снижена (уже не будет
100%-ной).
Пенетрантность характеризует процент особей, проявляющих в генотипе
данный ген, по отношению к общему числу особей, у которых он мог бы
проявиться.
26. Экспрессивность - степень выраженности признака при реализации генотипа в различных условиях среды. Эта выраженность
фенотипическогопроявления гена связана с изменчивостью признака в пределах нормы
реакции. Экспрессивность выражают количественно.
Пенетрантность и экспрессивность гена Lobe у D. melanogaster. Для этого
доминантного гена характерна варьирующая
экспрессивность: размер глава изменяется от нуля (А) до нормального (Е). Данный ген пенетрант
ен только у 75% носителей (А-Д)
27.
Кролики так называемой гималайской породы имеют своеобразную пеструю окраску меха:основной фон шерстного покрова - белый; мордочки, кончики ушей и лап, а также хвосты черные. Размер черных пятен зависит от температуры содержания гималайских кроликов. При
обычных «комнатных» температурах зверьки имеют такую окраску, как сказано выше, при
повышенных температурах размер черных пятен уменьшается, а при температурах выше 30 °С на
всем теле формируется белая шерсть; при пониженных температурах размер черных пятен
увеличивается, а при температурах близких к 0 °С — гималайские кролики приобретают чисто
черную окраску.
28.
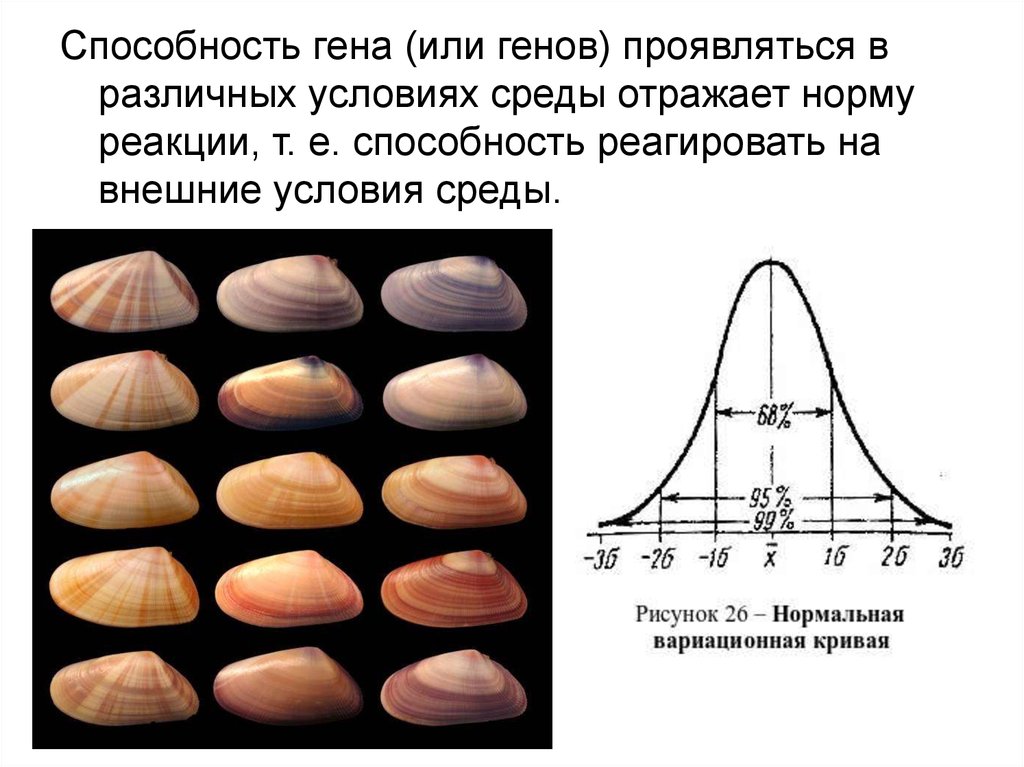
Способность гена (или генов) проявляться вразличных условиях среды отражает норму
реакции, т. е. способность реагировать на
внешние условия среды.
29.
Но́рма реа́кции — способность генотипа формировать в онтогенезе, в
зависимости от условий среды, разные фенотипы. Она характеризует долю
участия среды в реализации признака и определяет модификационную
изменчивость вида. Чем шире норма реакции, тем больше влияние среды и
тем меньше влияние генотипа в онтогенезе. Один и тот же ген в разных
условиях среды может реализоваться в несколько проявлений признака
(фенов). В каждом конкретном онтогенезе из спектра проявлений признака
реализуется только один. Аналогично один и тот же генотип в разных условиях
среды может реализоваться в целый спектр потенциально
возможных фенотипов, но в каждом конкретном онтогенезе реализуется
только один фенотип. Под наследственной нормой реакции понимают
максимально возможную ширину этого спектра: чем он шире, тем шире норма
реакции. Фенотипическое значение любого количественного признака
определяется, с одной стороны, его генотипическим значением, с другой
стороны — влиянием среды.