









 Биология
БиологияПохожие презентации:





")




Механізми утворення різноманітності антитіл i вклітинних рецепторів
1.
ЛЕКЦІЯ 12.ТЕМА: МЕХАНІЗМИ УТВОРЕННЯ
РІЗНОМАНІТНОСТІ АНТИТІЛ I ВКЛІТИННИХ РЕЦЕПТОРІВ.
Молекулярні механізми
перебудови генів антитіл та ВкР.
Соматичні гіпермутації в генах
імуноглобулінів.
"Помилки" під час рекомбінації.
1
2.
В нормі клітина має експресувати лишеодин тип функціонально перебудованих генів
важкого й легкого ланцюгів (явище алельного
виключення генів імуноглобулінів).
Біологічне значення: кожна лімфоїдна
клітина має нести рецептори лише однієї
специфічності.
2
3.
Алельневиключення
генів
імуноглобулінів
можна
пояснити
механічними перешкодами для одночасної
деконденсації хроматину двох гомологічних
хромосом, яке є необхідною умовою
перебігу реаранжування.
3
4.
Молекулярні механізмиперебудови генів антитіл та ВкР.
Молекулярні механізми з’єднання
V- і J-сегментів генів легких ланцюгів
або V-, D-, J-генних сегментів важких
ланцюгів
зумовлені
внутрішньохромосомними делеціями.
4
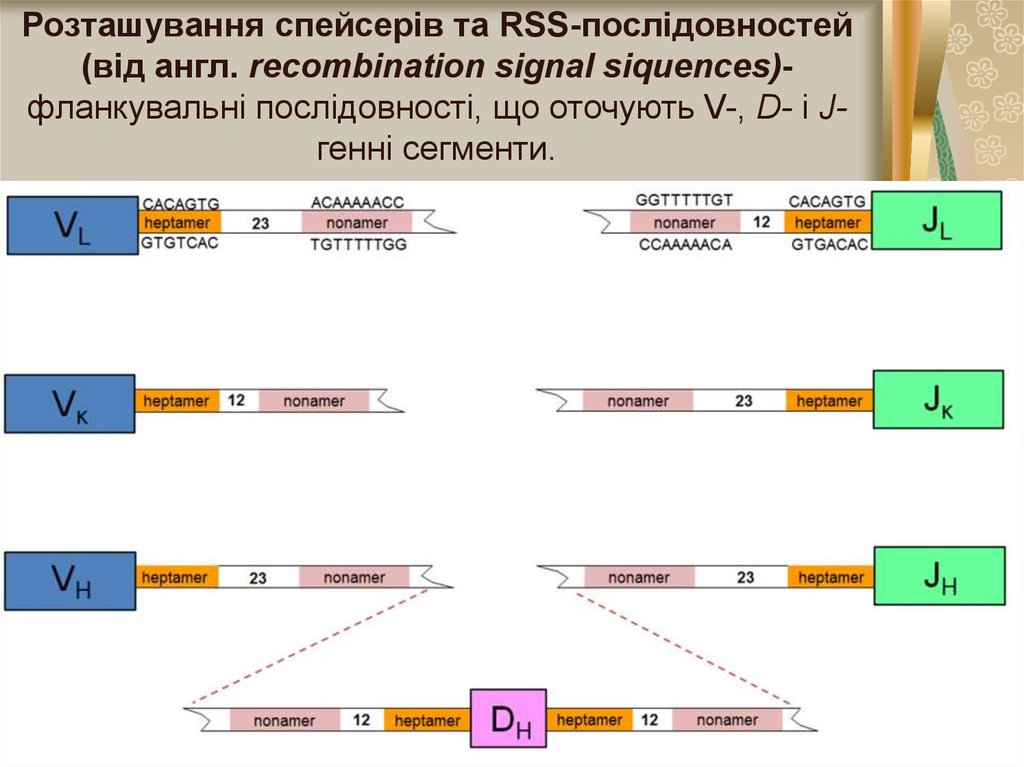
5.
Розташування спейсерів та RSS-послідовностей(від англ. rесотbіnаtion signal siquences)фланкувальні послідовності, що оточують V-, D- і Jгенні сегменти.
5
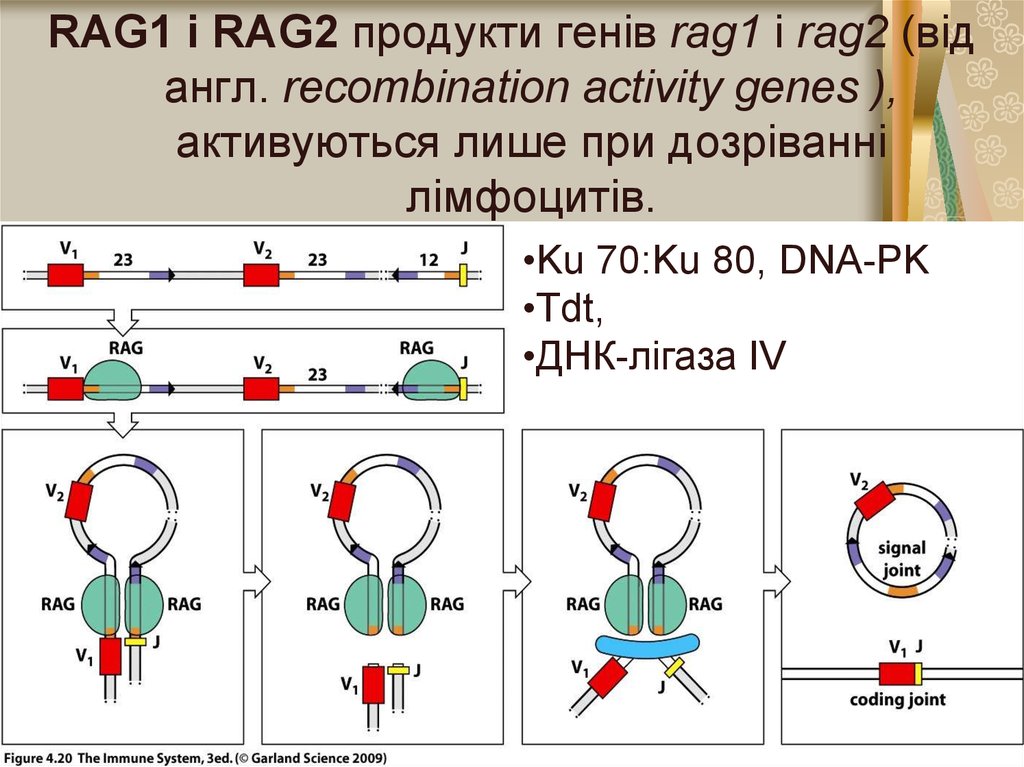
6.
RAG1 і RAG2 продукти генів rag1 і rag2 (відангл. rесотbіnаtіоn activity genes ),
активуються лише при дозріванні
лімфоцитів.
•Ku 70:Ku 80, DNA-PK
•Tdt,
•ДНК-лігаза ІV
6
7.
Антигеннезалежна перебудовагенів імуноглобулінів призводить до
формування функціональних ІgМта ІgD-рецепторів наївних В-клітин.
Антигензалежна – до переключення
синтезу з мембранної на
секреторну форму ІgМ або
переключення синтезу на інші
класи антитіл.
7
8.
Соматичні гіпермутації в генахімуноглобулінів.
Створення різноманітності антитіл може
бути результатом точкових соматичних
мутацій та "помилок" під час V-D-J
рекомбінацій.
8
9.
Соматичні мутації практично невідбуваються в ділянках, що кодують
сталі
ділянки
імуноглобулінових
ланцюгів.
Мутації виникають як в екзонах, так
і
в
інтронах,
однак
частіше
відбуваються в першій і другій
гіперваріабельних ділянках (CDR1 і
CDR2).
9
10.
Процесисоматичного
гіпермутагенезу
залежать
від
ферменту - індукованої активацією
дезамінази цитидину (AICD від англ.
activation induced citidine deaminase),
який вносить зміни в певні ділянки
ДНК, які потім репаруються з
помилками.
10
11.
"Помилки" під час рекомбінації.місця з'єднання V-D , D-J і V-Jсегментів можуть зміщуватися в частину
5'- і З' ділянок "сусідніх" генетичних
сегментів;
під час об’єднання V-сегментів з
відповідними D- і J-сегментами можуть
виникати вставки додаткових нуклеотидів
між сегментами, що об’єднуються, а також
втрати нуклеотидів через активність
певних екзонуклеаз.
11
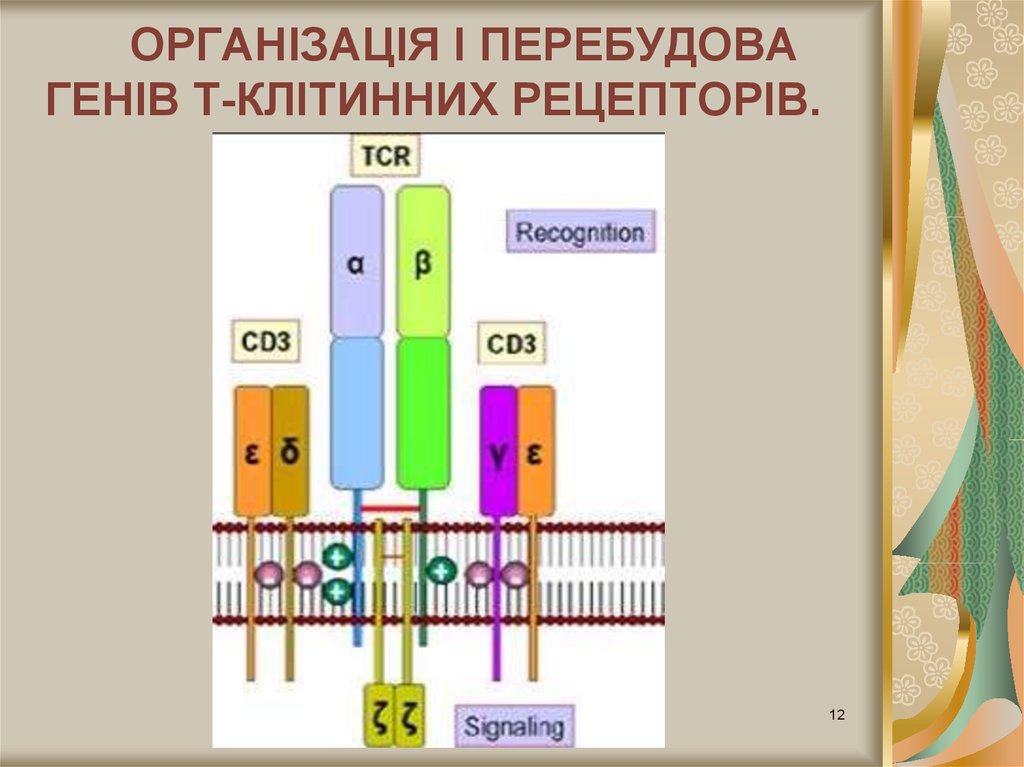
12.
ОРГАНІЗАЦІЯ I ПЕРЕБУДОВАГЕНІВ Т-КЛІТИННИХ РЕЦЕПТОРІВ.
12
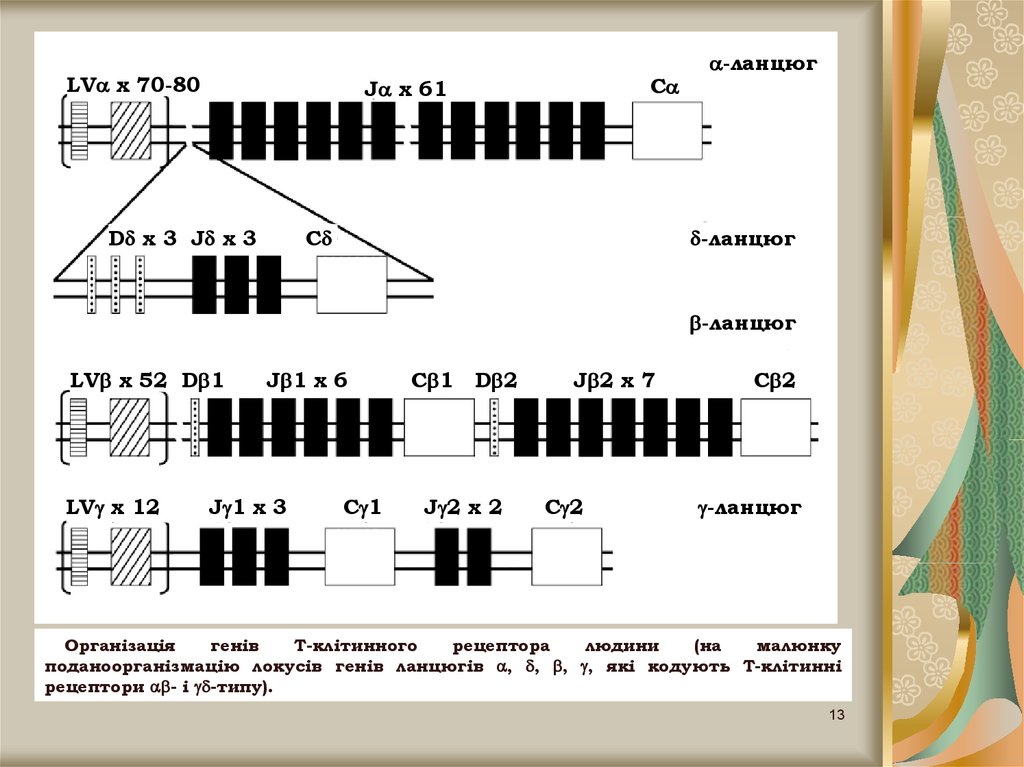
13.
LV x 70-80C
J x 61
D x 3 J x 3
-ланцюг
-ланцюг
C
-ланцюг
LV x 52 D 1
LV x 12
J 1 x 6
J 1 x 3
C 1
C 1
D 2
J 2 x 2
J 2 x 7
C 2
C 2
-ланцюг
Організація
генів
Т-клітинного
рецептора
людини
(на
малюнку
поданоорганізмацію локусів генів ланцюгів , , , , які кодують Т-клітинні
рецептори - і -типу).
13
14.
Схема рекомбінації зародкової ДНК ланцюгів (а) і (б)Т-клітинного рецептора людини.
а
LV x 70-80
J x 61
C
-ланцюг
VJ-рекомбінація
LV 1J
C
-ланцюг
б
LV x 52 D 1
C
J 1 x 6
C 1
D 2
J 2 x 7
C 2
VDJ-рекомбінація
14
LV 1D 1J C 1