
. Мембрана клеток в покое обладает высокой проницаемостью для")


")





 были известны очень")
")


")


")

.")












 Биология
БиологияПохожие презентации:










Мембранный потенциал
1.
Мембранный потенциалВ состоянии покоя между наружной и внутренней мембраной клетки
имеется разность потенциалов, которая называется мембранным
потенциалом (МП) или потенциалом покоя. Его величина
колеблется от -30 до – 100 мВ (у разных клеток).
Первая теория объясняющая возникновение МП была разработана
Ю.Бернштейном (1902). МП создается за счет неравномерного
распределения ионов по обе стороны от мембраны, и различной
проницаемости мембраны для этих ионов. В покое мембрана хорошо
проницаема для калия, которого много внутри клетки, поэтому он
выходит из нее. Органических анионов также много внутри клетки,
но они выйти не могут, т.к. мембрана для них непроницаема.
2.
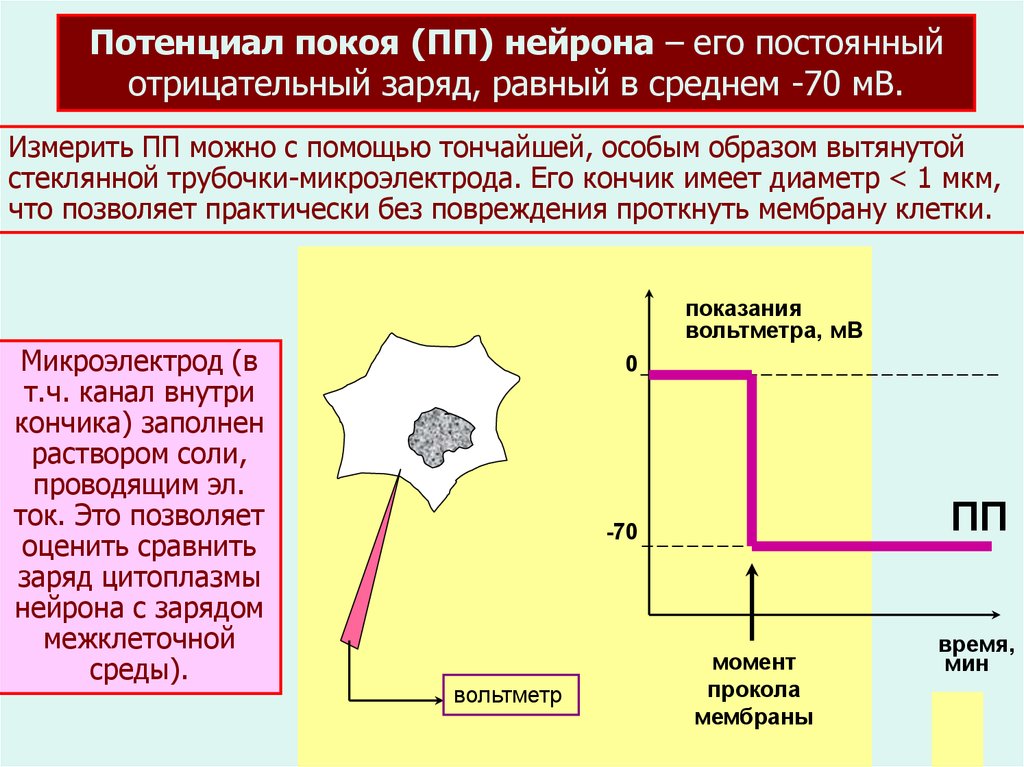
Потенциал покоя (ПП) нейрона – его постоянныйотрицательный заряд, равный в среднем -70 мВ.
Измерить ПП можно с помощью тончайшей, особым образом вытянутой
стеклянной трубочки-микроэлектрода. Его кончик имеет диаметр < 1 мкм,
что позволяет практически без повреждения проткнуть мембрану клетки.
Микроэлектрод (в
т.ч. канал внутри
кончика) заполнен
раствором соли,
проводящим эл.
ток. Это позволяет
оценить сравнить
заряд цитоплазмы
нейрона с зарядом
межклеточной
среды).
показания
вольтметра, мВ
0
ПП
-70
вольтметр
момент
прокола
мембраны
время,
мин
2
3. Механизмы возникновения мембранного потенциала (Ю.Бернштейн, 1902). Мембрана клеток в покое обладает высокой проницаемостью для
ионов калия, низкой для ионов натрия и не проницаема дляорганических анионов
4.
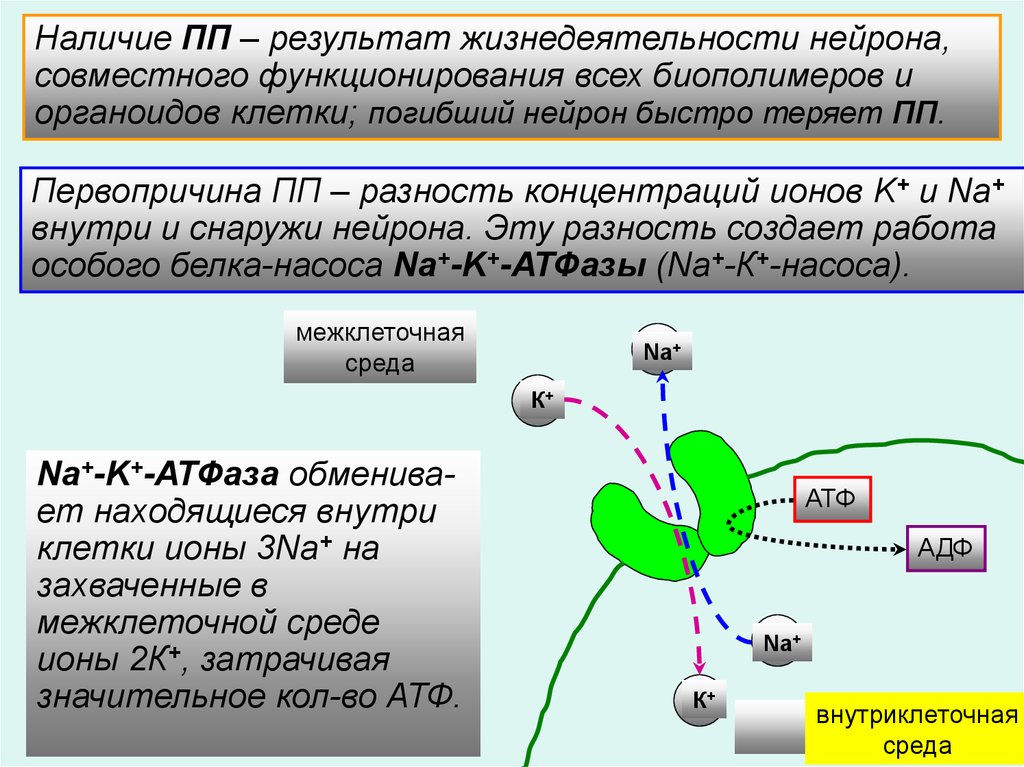
Наличие ПП – результат жизнедеятельности нейрона,совместного функционирования всех биополимеров и
органоидов клетки; погибший нейрон быстро теряет ПП.
Первопричина ПП – разность концентраций ионов K+ и Na+
внутри и снаружи нейрона. Эту разность создает работа
особого белка-насоса Na+-K+-АТФазы (Na+-К+-насоса).
межклеточная
среда
Na+
К+
Na+-K+-АТФаза обменивает находящиеся внутри
клетки ионы 3Na+ на
захваченные в
межклеточной среде
ионы 2К+, затрачивая
значительное кол-во АТФ.
АТФ
АДФ
Na+
К+
внутриклеточная
4
среда
5.
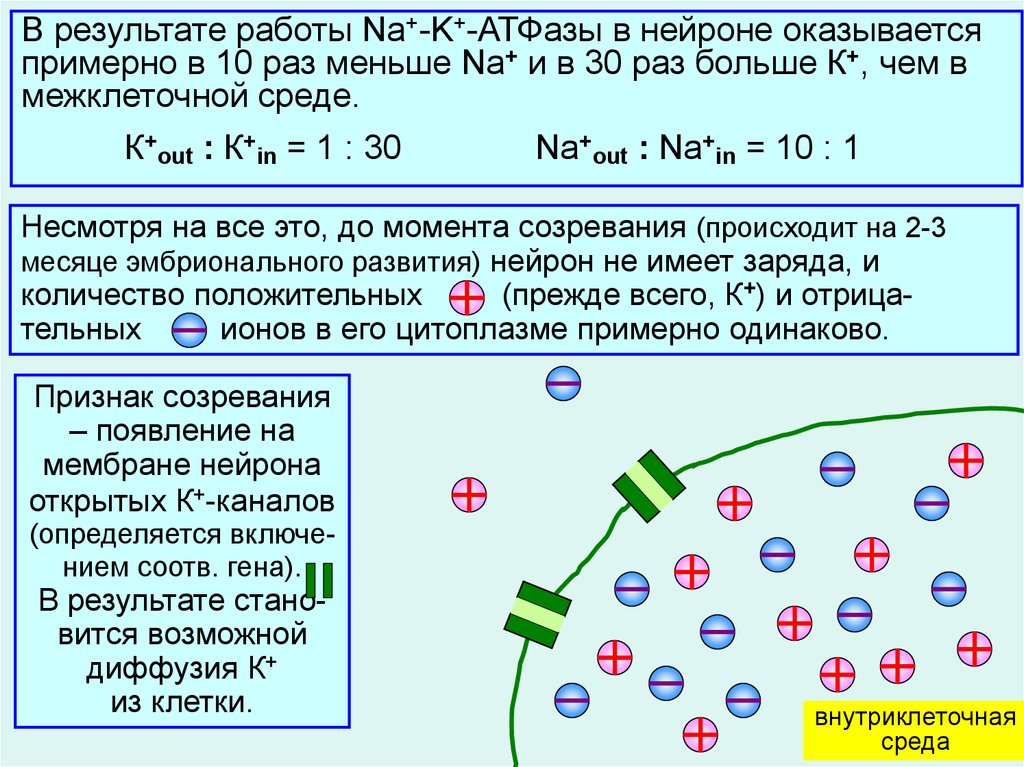
В результате работы Na+-K+-АТФазы в нейроне оказываетсяпримерно в 10 раз меньше Na+ и в 30 раз больше К+, чем в
межклеточной среде.
К+out : К+in = 1 : 30
Na+out : Na+in = 10 : 1
Несмотря на все это, до момента созревания (происходит на 2-3
месяце эмбрионального развития) нейрон не имеет заряда, и
количество положительных
(прежде всего, К+) и отрицательных
ионов в его цитоплазме примерно одинаково.
Признак созревания
– появление на
мембране нейрона
открытых К+-каналов
(определяется включением соотв. гена).
В результате становится возможной
диффузия К+
из клетки.
5
внутриклеточная
среда
6.
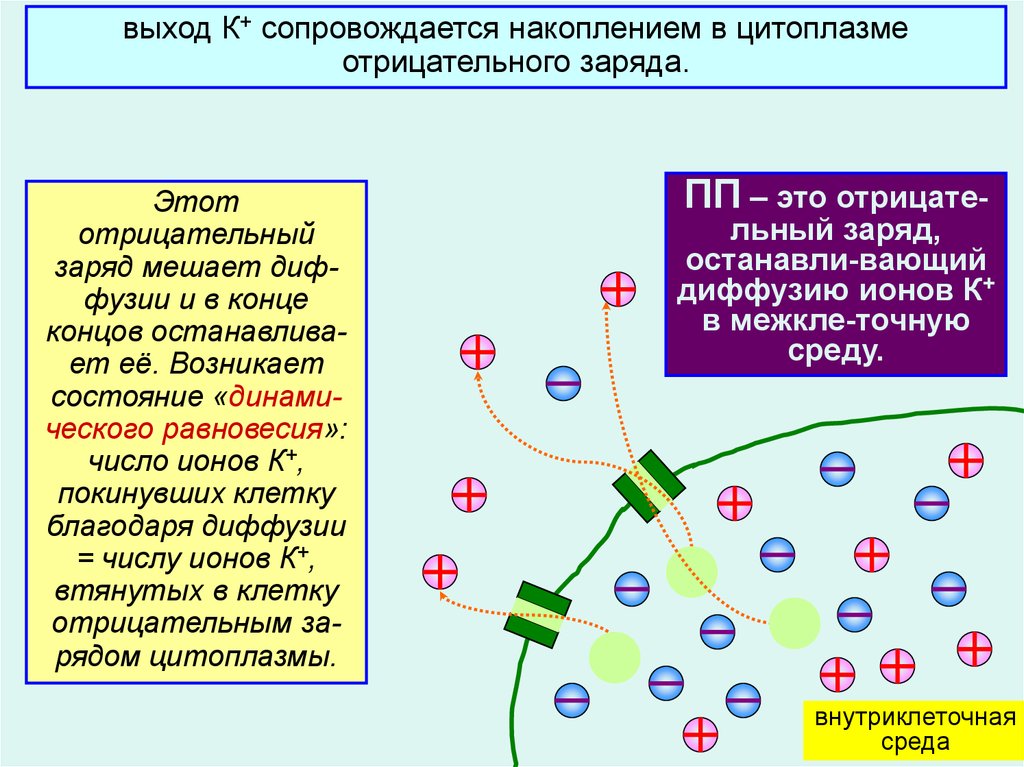
выход К+ сопровождается накоплением в цитоплазмеотрицательного заряда.
Этот
отрицательный
заряд мешает диффузии и в конце
концов останавливает её. Возникает
состояние «динамического равновесия»:
число ионов К+,
покинувших клетку
благодаря диффузии
= числу ионов К+,
втянутых в клетку
отрицательным зарядом цитоплазмы.
ПП – это отрицате-
льный заряд,
останавли-вающий
диффузию ионов К+
в межкле-точную
среду.
6
внутриклеточная
среда
7.
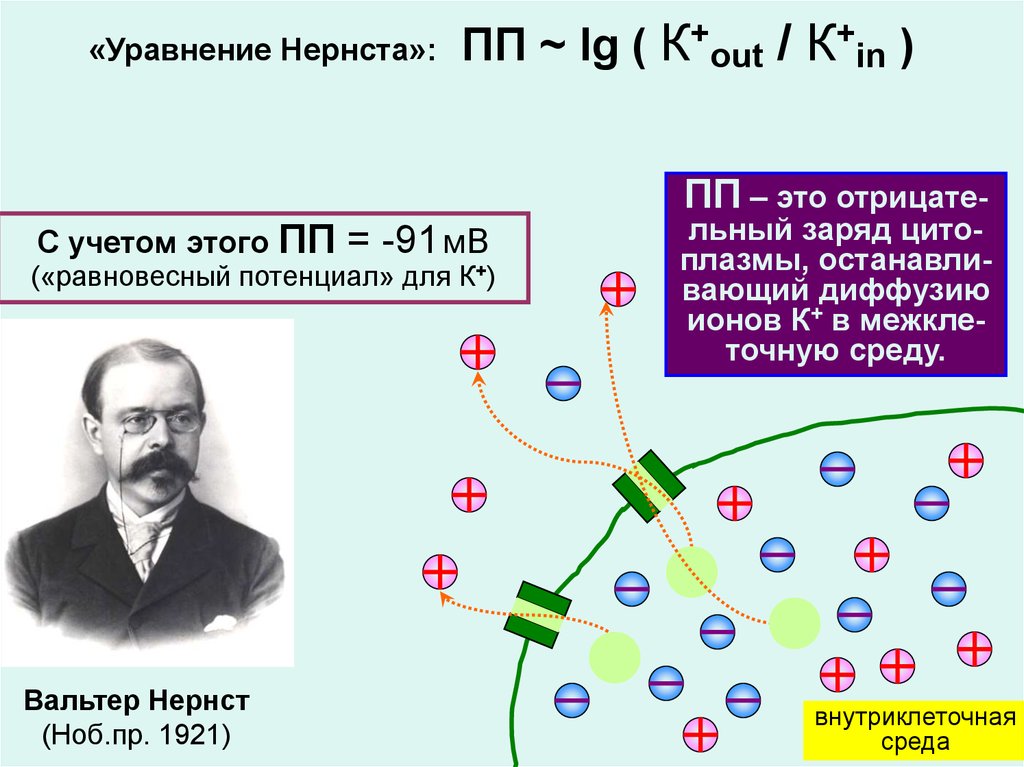
«Уравнение Нернста»:ПП ~ lg ( К+out / К+in )
ПП – это отрицате-
С учетом этого ПП
= -91 мВ
(«равновесный потенциал» для К+)
Вальтер Нернст
(Ноб.пр. 1921)
льный заряд цитоплазмы, останавливающий диффузию
ионов К+ в межклеточную среду.
7
внутриклеточная
среда
8.
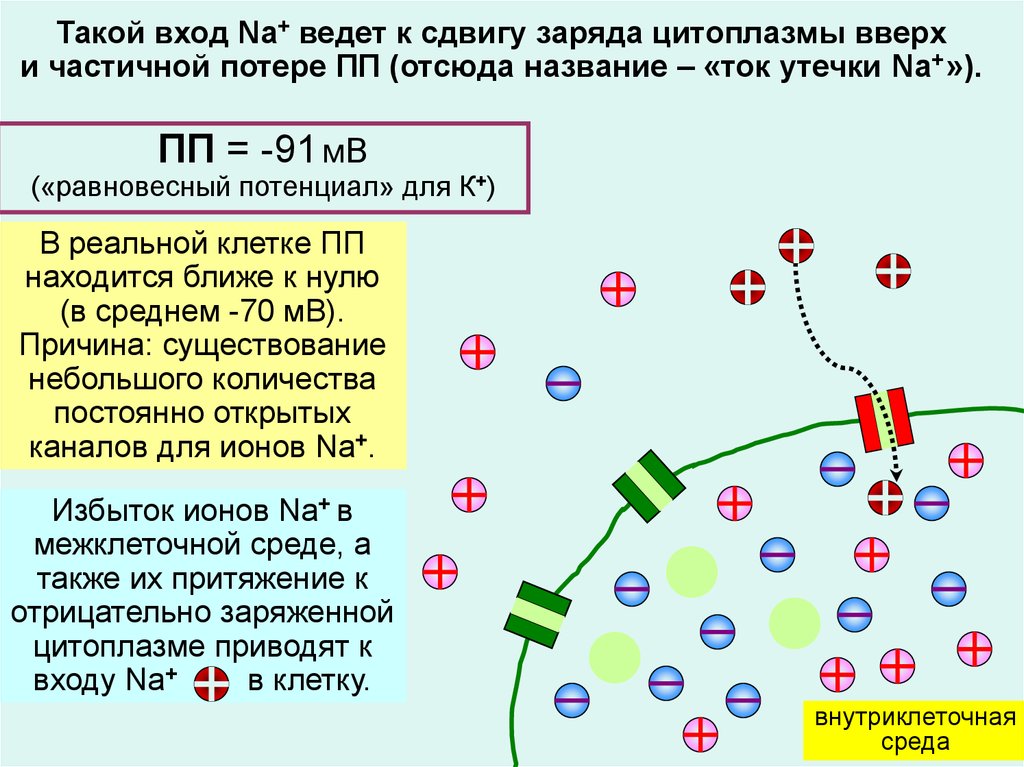
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверхи частичной потере ПП (отсюда название – «ток утечки Na+ »).
ПП = -91 мВ
(«равновесный потенциал» для К+)
В реальной клетке ПП
находится ближе к нулю
(в среднем -70 мВ).
Причина: существование
небольшого количества
постоянно открытых
каналов для ионов Na+.
Избыток ионов Na+ в
межклеточной среде, а
также их притяжение к
отрицательно заряженной
цитоплазме приводят к
входу Na+
в клетку.
8
внутриклеточная
среда
9.
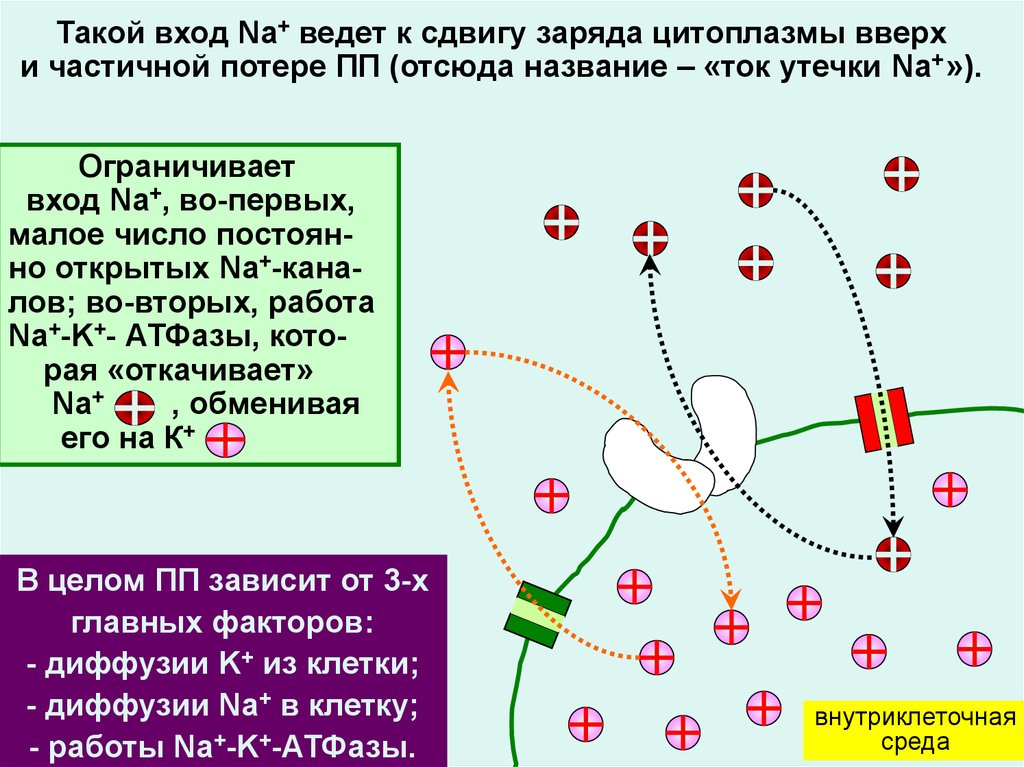
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверхи частичной потере ПП (отсюда название – «ток утечки Na+ »).
Ограничивает
вход Na+, во-первых,
малое число постоянно открытых Na+-каналов; во-вторых, работа
Na+-K+- АТФазы, которая «откачивает»
Na+
, обменивая
его на К+
В целом ПП зависит от 3-х
главных факторов:
- диффузии K+ из клетки;
- диффузии Na+ в клетку;
- работы Na+-K+-АТФазы.
9
внутриклеточная
среда
10.
+ ведет к сдвигу заряда цитоплазмы вверхТакой вход
Naклетки
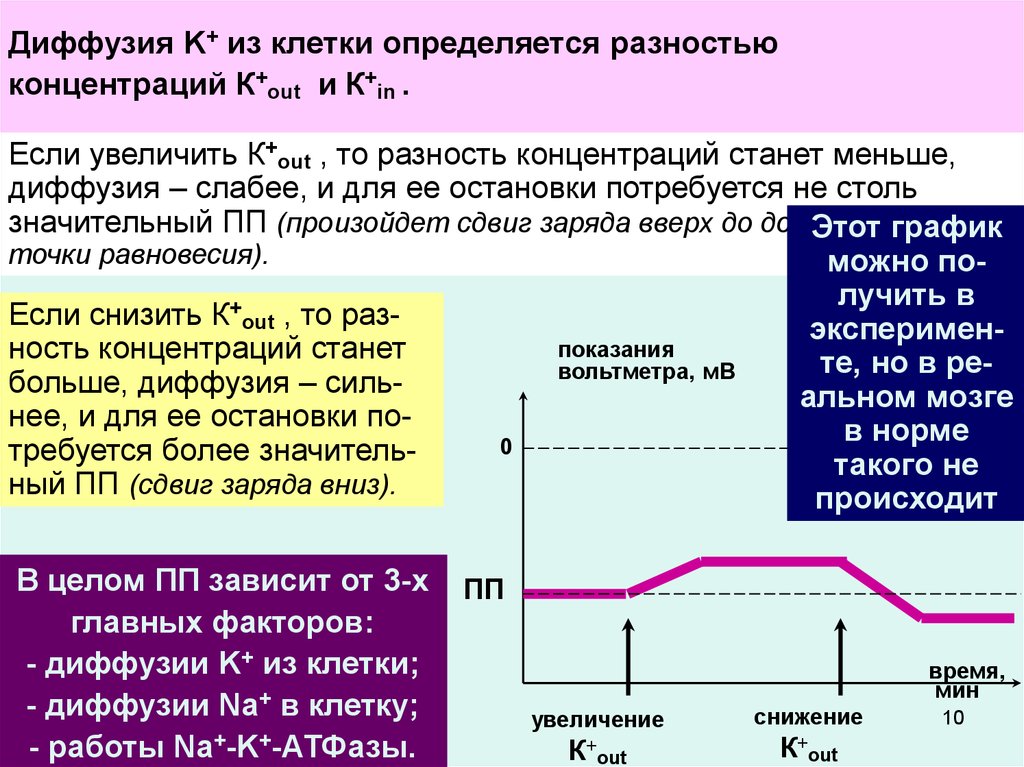
Диффузия
K+ из
определяется разностью
и частичной потере
ПП
(отсюда название – «ток утечки» Na+).
+
+
концентраций К out и К in .
Если увеличить К+out , то разность концентраций станет меньше,
диффузия – слабее, и для ее остановки потребуется не столь
значительный ПП (произойдет сдвиг заряда вверх до достижения
новой
Этот график
точки равновесия).
можно получить в
+
Если снизить К out , то разэксперименпоказания
ность концентраций станет
те, но в револьтметра, мВ
больше, диффузия – сильальном мозге
нее, и для ее остановки пов норме
0
требуется более значительтакого не
ный ПП (сдвиг заряда вниз).
происходит
В целом ПП зависит от 3-х
главных факторов:
- диффузии K+ из клетки;
- диффузии Na+ в клетку;
- работы Na+-K+-АТФазы.
ПП
время,
мин
увеличение
К+out
снижение
К+out
10
11.
+ ведет к сдвигу заряда цитоплазмы вверхТакой вход
Диффузия
Na+ Na
в клетку
зависит, прежде всего, от концентра-+
и частичной потере ПП (отсюда название – «ток утечки» Na ).
ции постоянно открытых Na+-каналов на мембране.
Эта величина, в свою очередь, является стабильным свойством
конкретного нейрона. Чем больше таких каналов, тем ПП ближе к
нулю, чем меньше – тем ПП ближе к уровню -91 мВ.
Чем ближе ПП к нулю, тем
возбудимее нейрон (такие
нужны, например, в центрах
бодрствования);
чем ближе ПП к уровню -91 мВ,
тем ниже возбудимость (минимальна в центрах, запускающих движения).
В целом ПП зависит от 3-х
главных факторов:
- диффузии K+ из клетки;
- диффузии Na+ в клетку;
- работы Na+-K+-АТФазы.
показания
вольтметра, мВ
0
-60
-70
-80
А: нейрон с большим числом
Na+-каналов
Б: нейрон со средним
числом Na+-каналов
(ПП= -70 мВ)
В: нейрон с малым числом
Na+-каналов
ПП нейрона А
ПП нейрона Б
ПП нейрона В
время, мин 11
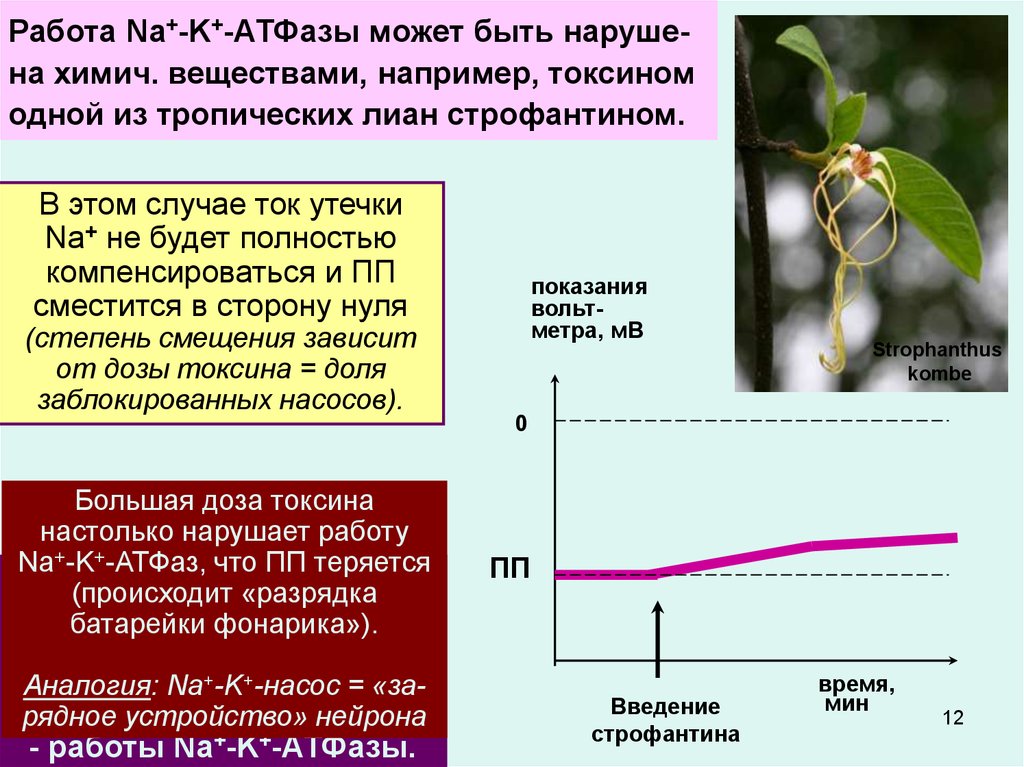
12.
Работа Na+-K+-АТФазы может быть нарушена химич. веществами, например, токсиномодной из тропических лиан строфантином.
В этом случае ток утечки
Na+ не будет полностью
компенсироваться и ПП
сместится в сторону нуля
(степень смещения зависит
от дозы токсина = доля
заблокированных насосов).
Большая доза токсина
настолько нарушает работу
Na+-K+-АТФаз, что ПП теряется
В целом
ПП зависит
от 3-х
(происходит
«разрядка
главных факторов:
батарейки
фонарика»).
- диффузии+ K++ из клетки;
Аналогия: Na -K -насос
= «за+ в клетку;
-рядное
диффузии
Na
устройство» нейрона
- работы Na+-K+-АТФазы.
показания
вольтметра, мВ
Strophanthus
kombe
0
ПП
Введение
строфантина
время,
мин
12
13. Na,K –ATФаза ингибируется уабаином
Na,K –ATФаза ингибируется
уабаином
В 1785 году Визеринг сообщил об использовании листьев
наперстянки для лечения сердечной недостаточности.
Действующим началом являлся дигитоксин, соединение,
относящееся к сердечным гликозидам – группе стероидных Огликозидов. Наиболее известным соединением этого ряда
является уабаин.
в 1950-60 годы
синтезирован уабаин и др. гликозиды
Показана их способность избирательно
связываться и блокировать активность АТФазы
нервной ткани. Уабаин использован для
выделения и идентификации АТФазы нервной
ткани
14. Наперстянка (дигиталис)
Симптомыотравления:
1) Сердечный приступ
2) Тошнота, рвота
3) Диарея
4) Головная боль
5) Медленный пульс
6) Делирий
Первая помощь при отравлении: очистить желудок и
кишечник, принять активированный уголь, обратиться к
врачу. Можно выпить крепкого кофе.
15. Интересные факты о наперстянке
• Один из побочныхэффектов лекарств с
наперстянкой –
изменённое
восприятие цветов
• Воздействие
наперстянки на
сердце было открыто
только 1775 году
Уильямом
Уизерингомю
16. Уравнение Гольдмана-Ходжкина-Катца
Уравнение ГольдманаХоджкина-КатцаPK [K ]вн PNa [Na ]вн PCl [Cl ]нар
RT
Vm ln
F P [K ] P [Na ] P [Cl ]
K
нар Na
нар Cl
вн
17. Биоэлектрические явления в возбудимых тканях
Физиологию возбудимых тканей изучаетэлектрофизиология.
Электрофизиология –исследует
электрические проявления жизнедеятельности
клеток, тканей и органов для выяснения
природы биоэлектрических феноменов и их
физиологического значения, а также для их
использования в качестве показателей
функционирования организма.
18. Основные задачи электрофизиологии:
• 1. Изучение физических и химических механизмовэлектрических процессов в живых тканях, т.е. природы
генерации биопотенциалов.
• 2. Исследование свойств живых тканей как проводников
электричества.
• 3. Исследование действия электрического тока на
процессы жизнедеятельности.
• Важность изучения электрофизиологии определяется
тем, что:
• 1. Многие из электрофизиологических методов широко
используются в клинике для диагностики (ЭКГ – сердце,
ЭЭГ – мозг, миография – мышцы и т.д.
• 2. Нарушения биоэлектрических процессов приводит к
возникновению различных заболеваний. Поэтому знание
электрофизиологии необходимо для понимания
механизмов патогенеза многих заболеваний.
• 3. Методы электростимуляции используются в
кардиологии, неврологии, спортивной медицине,
косметологии.
19. В основе реакций организма лежит раздражимость - способность реагировать на действие различных факторов изменением функции.
В основе реакций организма лежит раздражимость способность реагировать на действие различных факторовизменением функции.
• Возбудимость - способность отвечать на действие
раздражителя возбуждением. В электрофизиологии
термином «возбуждение» обозначают 2 процесса:
• 1.Процесс временной деполяризации мембраны клетки, т.е.
генерацию потенциала действия (ПД).
• 2.Специфический ответ ткани на возникший в ней ПД
(проведение нервного импульса, сокращение мышцы,
выделение секрета железой и т. д.).
• Возбудимостью обладают нервная, мышечная и секреторная
ткани, которые называют «возбудимыми тканями».
• Проводимость – способность передавать возбуждение на
расстояние.
• Раздражители (стимулы) – различные факторы внешней
среды, способные вызывать возбуждение.
• Возбудимость различных тканей неодинакова. Ее величину
оценивают по порогу раздражения - минимальной силе
раздражителя, которая способна вызвать возбуждение.
20. Явления связанные с природным электричеством (молнии, статическое электричество, электрические рыбы и др.) были известны очень
давноОднако их научное изучение началось лишь в 17 веке.
21. Бенджамин Франклин (1706—1790)
Разработал (1750) теорию
электрических явлений, согласно
которой электричество представляет
особую “электрическую жидкость»,
пронизывающую все тела.
Если появляется ее излишек, то тело
заряжается положительно, когда ее
недостает — отрицательно. Им было
введено понятие положительного и
отрицательного заряда и их
обозначение: «+» и «-».
В 1753 доказал электрическую природу
молнии (опыт со змеем) и
тождественность земного и
атмосферного электричества. Изобрел
молниеотвод.
Его опыты стимулировали
исследования врачей и физиологов,
которые начали использовать действие
электрических зарядов для лечения
болезней
22.
23. Впервые искусственную электрическую искру получили от электрической машины, изобретённой Герике - шар из серы, насаженный на
ось,укреплённую в деревянном штативе. При вращении шар электризовался
ладонями рук.
• Зная, что стекло не проводит
электричество, Мушенбрук в
1745 г. взял стеклянную колбу,
наполненную водой, опустил в
нее медную проволоку,
соединенную с кондуктором
электрической машины и
попросил своего помощника
вращать шар.
• Он предположил, что заряды,
поступавшие с кондуктора,
будут накапливаться в
стеклянной банке. Когда он
стал отсоединять медную
проволоку то получил сильный
удар тока.
Так была изобретена
лейденская банка
24. Первые электрофизиологические опыты. Луиджи Гальвани (1737-1798)
Он обратил внимание на
сокращение препарата задних
лапок лягушки при
прикосновении к ним скальпеля,
если рядом вращалась
электрофорная машина.
Решил проверить будет ли
аналогично действовать
атмосферное электричество.
Для этого он подвешивал
препараты лапок с помощью
медных крючках на перилах
балкона. Обнаружил, что когда
ветер раскачивал лапки и они
соприкасались с железными
перилами, происходит
сокращение мышц
25.
Итальянский физиолог Луиджи
Гальвани из Болоньи сделал
важное наблюдение. Он
заметил, что тело мертвой
лягушки вздрагивало вблизи
электрической машины, как
только из машины
извлекались искры. Такое же
явление он заметил, когда
повесил препарированные
ножки лягушки на медную
проволоку на балконной
решетке и когда от ветра
ножки лягушки прикасались к
железу решетки.
Гальвани верно приписал
вздрагивание ножек действию
электричества,
26. Лаборатория Гальвани
27. Алессандро Вольта (1745-1827)
Вольта повторил опыт Гальвани,
но объяснил, его результаты тем,
что в цепи из двух разнородных
металлов (цинк и медь) и
электролита (физиологический
раствор) возникает ток, который
вызывает сокращение мышцы.
То есть источником тока является
не живая ткань, а металлы, а
живая ткань является лишь
проводником электрического тока.
Спор между А.Вольта и Л.Гальвани
оказал огромное влияние на
развитие физиологии.
А.Вольта создал генератор
электрического тока –
гальванический элемент (вольтов
столб).
Ввел понятие об электродвижущей
силе, предложил ее единицу –
Вольт.
28. Второй опыт Л. Гальвани
• Для того, чтобы доказать, что ткани животных тоже способныгенерировать электрический ток Л.Гальвани исключил из опыта
металлы
• Он препарировал седалищный нерв вдоль бедра лапки
лягушки, а затем набрасывал его с помощью стеклянного
крючка на мышцы голени, что вызывало сокращение этих мышц
(второй опыт Гальвани).
29. Первый «балконный» опыт Л.Гальвани (1786 г).
• Гальвани повторилэтот опыт в условиях
лаборатории,
прикасаясь к
препаратам пинцетами,
сделанными из
различных металлов.
• Максимальный эффект
возникал если
использовался пинцет
сделанный из меди и
цинка.
30.
Джованни Альдини (1762 - 1834),
хотя именно он открыл новую
страницу сопротивления
смертоносному началу природы.
Альдини, судебный медик из
Болоньи, включил в свои
исследования и человеческие тела, а
именно, трупы заключенных,
казненных по приговору суда. В
сферу экспериментов ученого попали
и обезглавленные тела, и
отрубленные головы, но так же и
нерасчлененные трупы, которые, по
всей видимости, приходилось
выкупать, на что ученый тратил свои
личные гонорары: всего лишь один
из примеров беззаветного служения
науке. Впрочем, гальванизируемые
человеческие трупы в целом
показывали ту же картину, что и
тушки лягушек, свежеубиенные туши
коров - но Альдини работал, не
покладая рук, добиваясь тонкой
дифференциации движений.
Результаты своих опытов ученый
тщательно фиксировал.
Триумф исследовательской программы Дж. Альдини произошел в Лондоне 17 января 1803 года. В этот день,
имевший тогда уже европейскую известность ученый провел демонстрационный эксперимент с телом
повешенного Джорджа Фостера. Меняя параметры тока - напряжение, силу, частоту, а также точки крепления
электродов, Альдини продемонстрировал широкий спектр возможностей, открываемых гальванизацией трупа.
При активировании лицевых нервов публика видела гримасы ужаса и боли, при гальванизации верхних
конечностей Фостер вскидывал руки, вращал ими, сжимал и разжимал кисти; ученый, манипулируя
электродами, извлекал из тела повешенного различные звуки, а палитра моторных реакций повешенного в
ряде случаев превосходила возможности обычного живого тела: так, у Фостера шевелились волосы и
двигались уши. Но особое впечатление на публику произвел своеобразный танец, который Джованни Альдини
удавалось воспроизводить и в прежних экспериментах (правда, с обезглавленными трупами): Фостер кружился,
поднимал ноги, совершая ряд узнаваемых танцевальных движений... Так описывали лондонские газеты этот
впечатляющий Death Dance на следующий день. Несколько человек из публики упали в обморок.
31. Дальнейшие исследования
• В 1838 г. Матеуччи обнаружил, что можнозарегистрировать электрический ток (ток
покоя), который течет от ее
неповрежденной поверхности к поперечному
разрезу.
• В 1841 г. Дюбуа Реймон установил, что
поврежденный участок имеет
отрицательный заряд, а неповрежденный –
положительный. Ввел понятия
«возбуждение», «возбудимые ткани»
32. Что такое возбудимые ткани
• Это ткани, клетки которых способны в ответ нараздражение генерировать специфическую реакцию –
возбуждение.
• Возбуждение – это процесс перехода живой клетки из
состояния покоя в состояние активности. Это
сопровождается появлением в клетке высокоамплитудного
электрического сигнала – потенциала действия (нервного
импульса).
• Раздражение – процесс действия раздражителя, который
действуя на возбудимую клетку, вызывает возбуждение.
• Раздражитель ( фактор внешней или внутренней среды, в
нервной системе это ионный электрический ток) может
вызвать раздражение если его сила равна или превышает
определенную величину – порог раздражения.
33. Все электрические процессы разворачиваются на цитоплазматической мембране. Клеточные мембраны состоят из жидкой фазы липидов и
встроенных в липидыбелковых молекул. Молекулы липидов организованы в бислойную мембрану (6
нм).Полярные гидрофильные головки обращены к поверхностям мембраны,
гидрофобные к хвосты вытянуты к середине бислоя. Мембрана плохо пропускает
воду, практически непроницаемы для ионов т.к. липиды плотно упакованы в
мембране и между ними нет практически никаких расстояний.
34. Белковые молекулы частично погружены в слой липидов, либо с внеклеточной стороны, либо с внешней стороны. Некоторые белки
полностью пронизывают мембрану. Именно они (трансмембранные)образуют структуры, образующие движение ионов через мембрану ( ионные
переносчики и ионные каналы)
35.
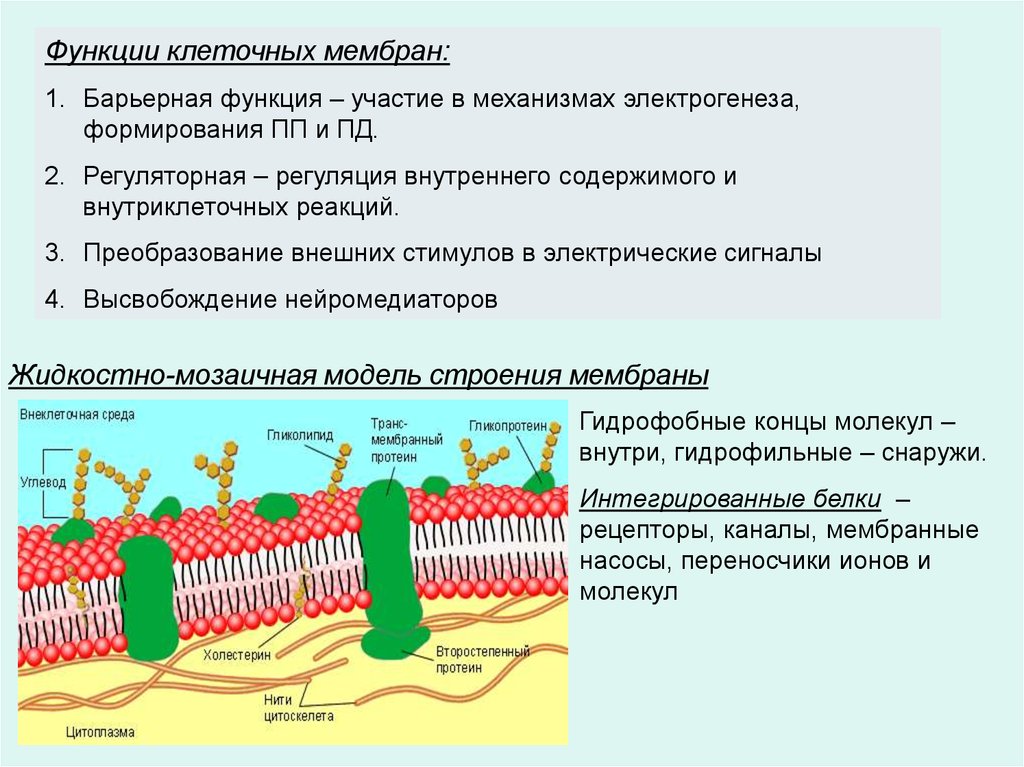
Функции клеточных мембран:1. Барьерная функция – участие в механизмах электрогенеза,
формирования ПП и ПД.
2. Регуляторная – регуляция внутреннего содержимого и
внутриклеточных реакций.
3. Преобразование внешних стимулов в электрические сигналы
4. Высвобождение нейромедиаторов
Жидкостно-мозаичная модель строения мембраны
Гидрофобные концы молекул –
внутри, гидрофильные – снаружи.
Интегрированные белки –
рецепторы, каналы, мембранные
насосы, переносчики ионов и
молекул
36.
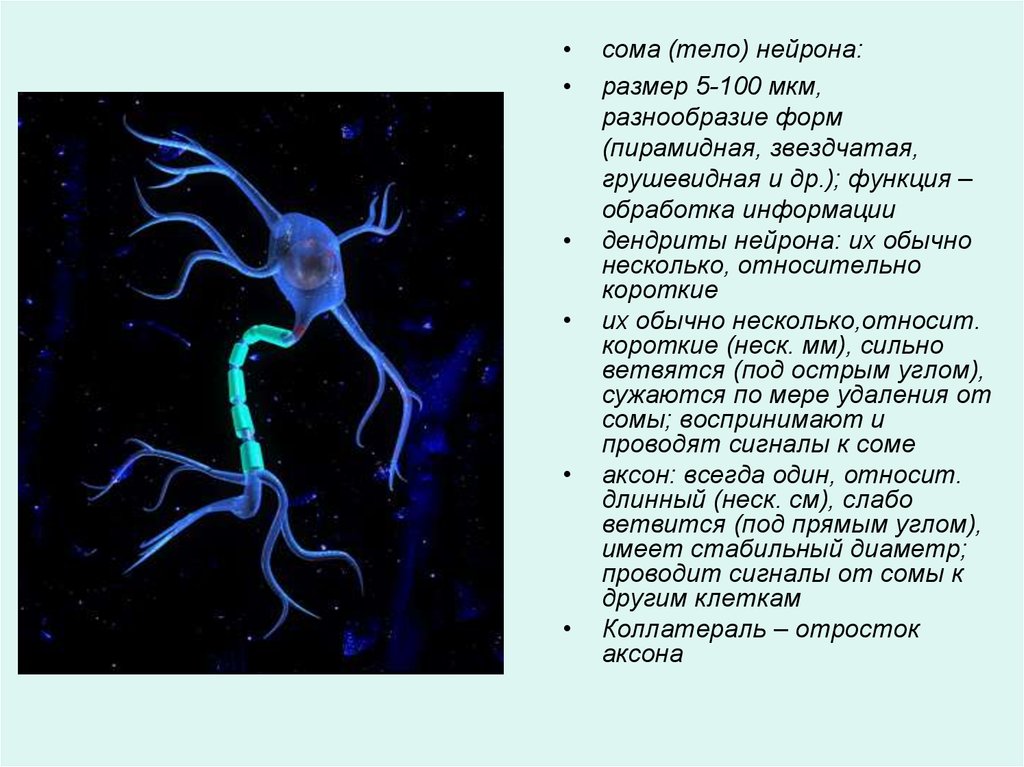
сома (тело) нейрона:
размер 5-100 мкм,
разнообразие форм
(пирамидная, звездчатая,
грушевидная и др.); функция –
обработка информации
дендриты нейрона: их обычно
несколько, относительно
короткие
их обычно несколько,относит.
короткие (неск. мм), сильно
ветвятся (под острым углом),
сужаются по мере удаления от
сомы; воспринимают и
проводят сигналы к соме
аксон: всегда один, относит.
длинный (неск. см), слабо
ветвится (под прямым углом),
имеет стабильный диаметр;
проводит сигналы от сомы к
другим клеткам
Коллатераль – отросток
аксона
37.
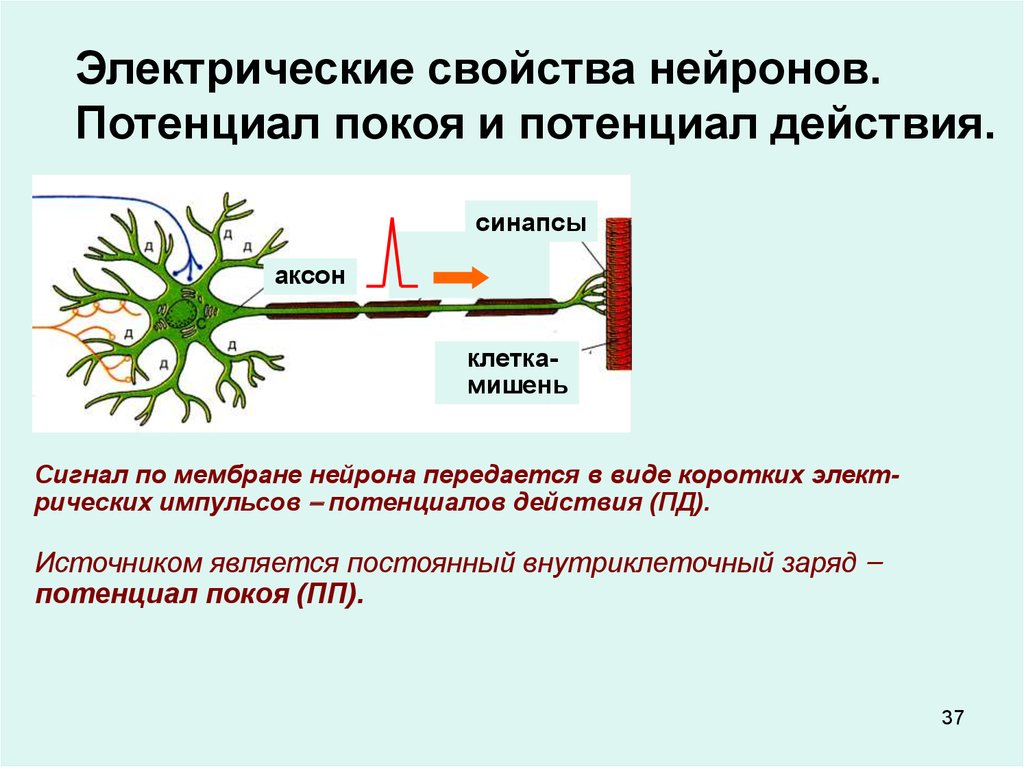
Электрические свойства нейронов.Потенциал покоя и потенциал действия.
синапсы
аксон
клеткамишень
Сигнал по мембране нейрона передается в виде коротких электрических импульсов – потенциалов действия (ПД).
Источником является постоянный внутриклеточный заряд –
потенциал покоя (ПП).
37
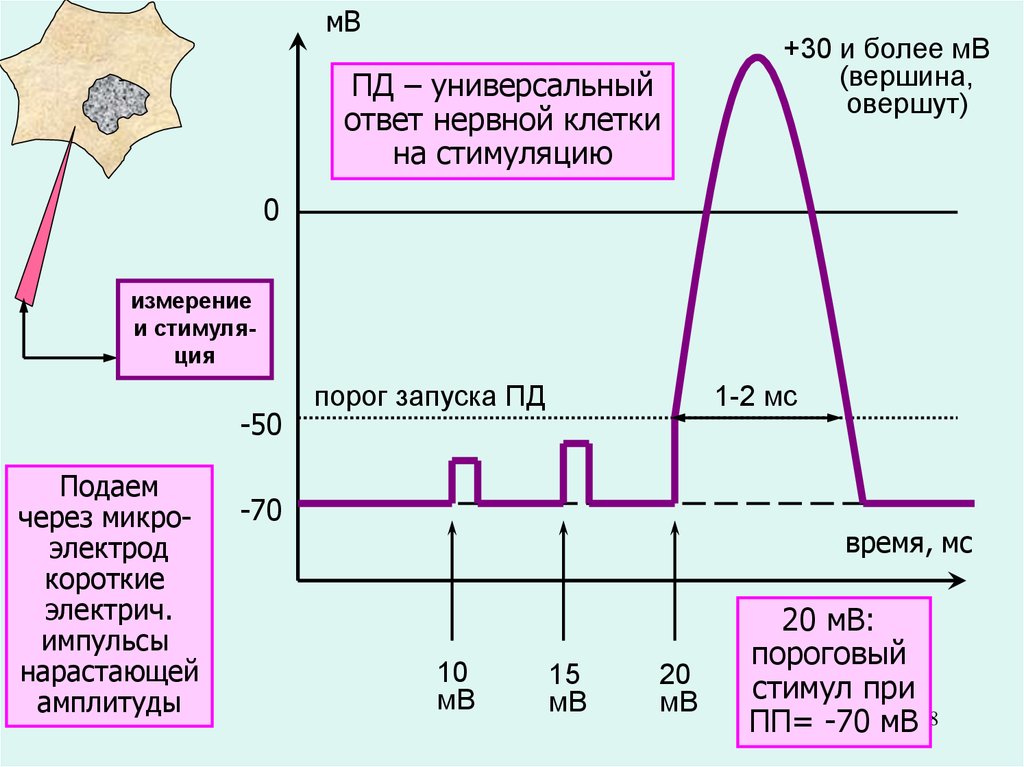
38.
мВПД – универсальный
ответ нервной клетки
на стимуляцию
+30 и более мВ
(вершина,
овершут)
0
измерение
и стимуляция
-50
Подаем
через микроэлектрод
короткие
электрич.
импульсы
нарастающей
амплитуды
порог запуска ПД
1-2 мс
-70
время, мс
10
мВ
15
мВ
20
мВ
20 мВ:
пороговый
стимул при
ПП= -70 мВ38
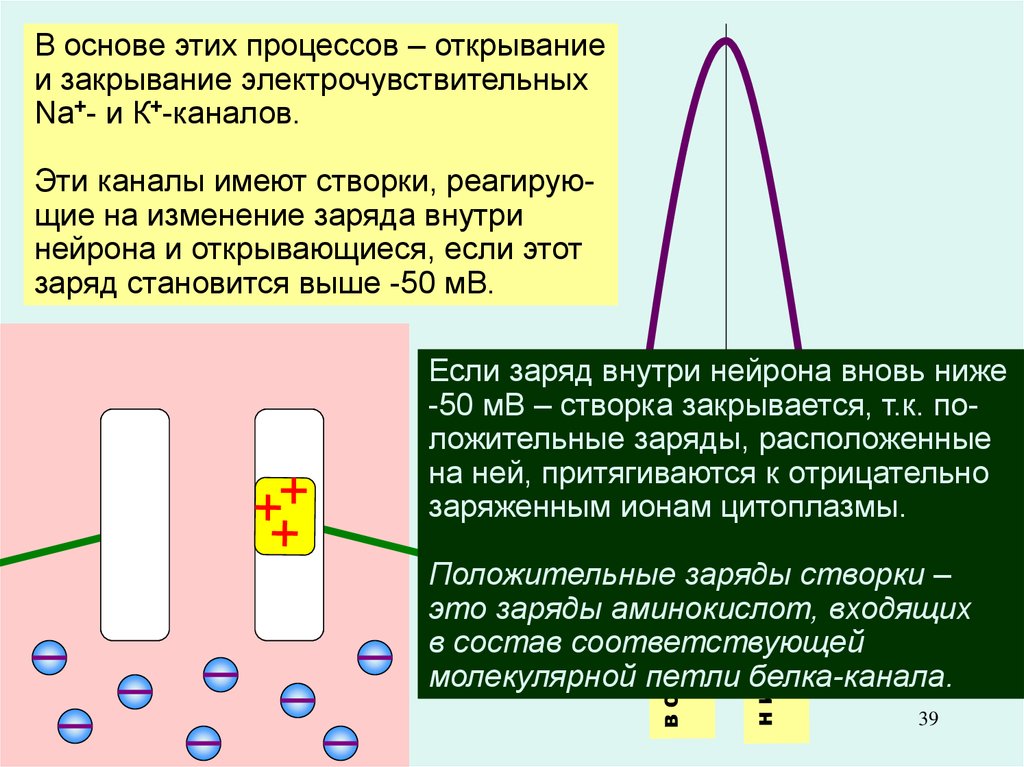
39.
В основе этих процессов – открываниеи закрывание электрочувствительных
Na+- и К+-каналов.
Эти каналы имеют створки, реагирующие на изменение заряда внутри
нейрона и открывающиеся, если этот
заряд становится выше -50 мВ.
Восходящая фаза
(деполяризация):
вход в клетку
«порции» Na+.
Нисходящая фаза
(реполяризация):
выход из клетки
примерно такой
же «порции» К+.
Если заряд внутри нейрона вновь ниже
-50 мВ – створка закрывается, т.к. повход Na+
выход К+
ложительные
заряды, расположенные
на ней, притягиваются к отрицательно
заряженным ионам цитоплазмы.
Положительные заряды створки –
это заряды аминокислот, входящих
в состав
ПП соответствующей
молекулярной петли белка-канала.
39
40.
Открытие электрочувствительного Na+-канала «разрешает»+ в клетку.
В вход
основе
процессов
– открывание
Naэтих
Открытие
электрочувствительного
и закрывание
электрочувствительных
К+-канала «разрешает»
выход К+ из клетки.
Na+- +и К+-каналов.
Na -каналы открываются очень быстро после стимула и
самопроизвольно
закрываются
примерно через 0.5 мс.
Эти
каналы имеют створки,
реагирующие
на изменение
заряда медленно
внутри
К+-каналы
открываются
– в течение примерно
нейрона
и открывающиеся,
если этот они в большинстве
0.5 мс после
стимула; закрываются
заряд
становится
-50 мВ.
своем
к моментувыше
снижения
заряда нейрона до уровня ПП.
Восходящая
фаза: вход в
Для
закрытия Na+-канаклетку
лов
на пике Na
ПД+.служит
«порции»
дополнительная
Нисходящая
(внутриклеточная,
фаза: выход из И-)
инактивационная,
клетки –пристворка
h-ворота.
мерностворка
такой же
Вторая
«порции» К+. А-) –
(активационная,
m-ворота.
Именно разная скорость открытия
Если+ заряд внутри+ нейрона вновь ниже
Na -каналов и К -каналов позволяет
-50 мВ – створка закрывается, т.к. повозникнуть сначала восходящей, а
ложительные заряды, расположенные
затем – нисходящей фазе ПД.
на ней, притягиваются к отрицательно
(сначала ионы
Na+цитоплазмы.
вносят в нейрон
заряженным
ионам
положительный заряд, а затем ионы К+ выносят его,
возвращая
Положительные
заряды
створки –
клетку
в исходное
состояние.
это
заряды
аминокислот,
входящих
в состав соответствующей
молекулярной петли белка-канала.
40
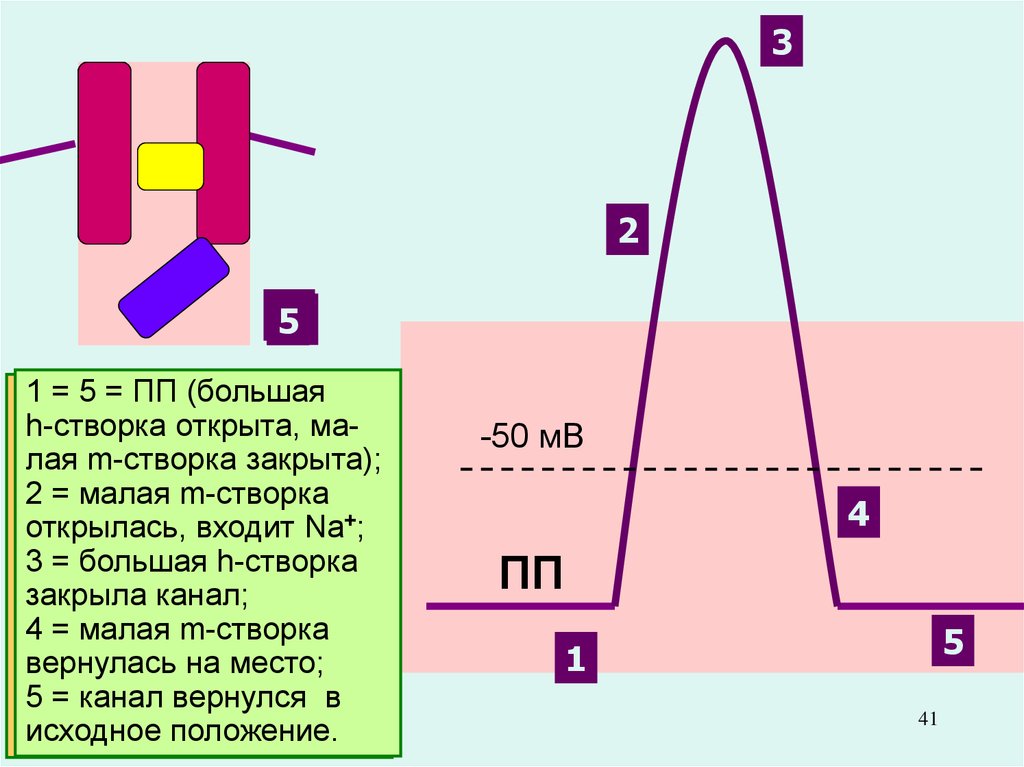
41.
32
1
3
4
5
2
1 = 5 = ПП (большая
h-створка
открыта,
маДля
закрытия
Na+-каналаяна
m-створка
лов
пике ПД закрыта);
служит
2 = малая m-створка
дополнительная
открылась, входит Na+;
(внутриклеточная,
3 = большая h-створка
инактивационная,
И-)
закрыла–канал;
створка
h-ворота
4 = малая
m-створка
Вторая
створка
вернулась на место;
(активационная,
А-) –
5 = канал вернулся в
m-ворота.
исходное положение.
Именно разная скорость открытия
Na+-каналов и К+-каналов позволяет
возникнуть сначала восходящей, а
-50 мВ
затем – нисходящей фазе ПД.
(сначала ионы Na+ вносят 4в нейрон
положительный заряд, а затем ионы ПП
К+ выносят его, возвращая
клетку в исходное состояние.
5
1
41
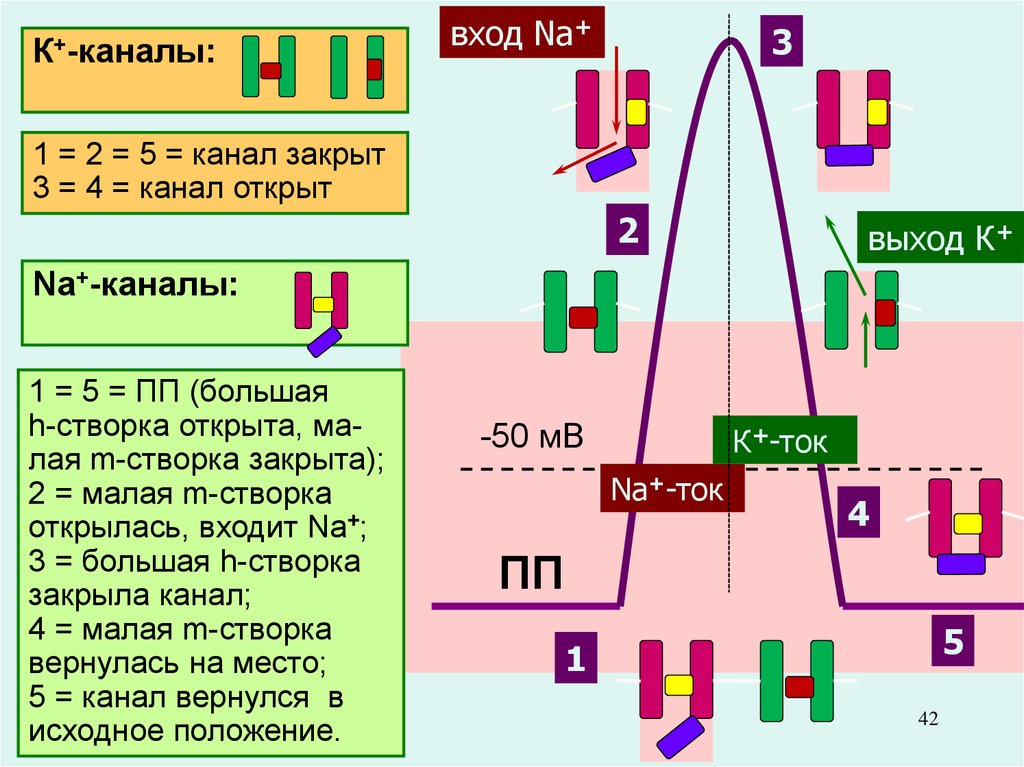
42.
К+-каналы:вход Na+
3
1 = 2 = 5 = канал закрыт
3 = 4 = канал открыт
2
выход К+
Na+-каналы:
1 = 5 = ПП (большая
h-створка открыта, малая m-створка закрыта);
2 = малая m-створка
открылась, входит Na+;
3 = большая h-створка
закрыла канал;
4 = малая m-створка
вернулась на место;
5 = канал вернулся в
исходное положение.
Именно разная скорость открытия
Na+-каналов и К+-каналов позволяет
возникнуть сначала восходящей,
а
+
-50 мВ
К фазе
-ток ПД.
затем – нисходящей
+-ток
Na
(сначала ионы Na+ вносят 4в нейрон
положительный заряд, а затем ионы ПП
К+ выносят его, возвращая
клетку в исходное состояние.
5
1
42
43.
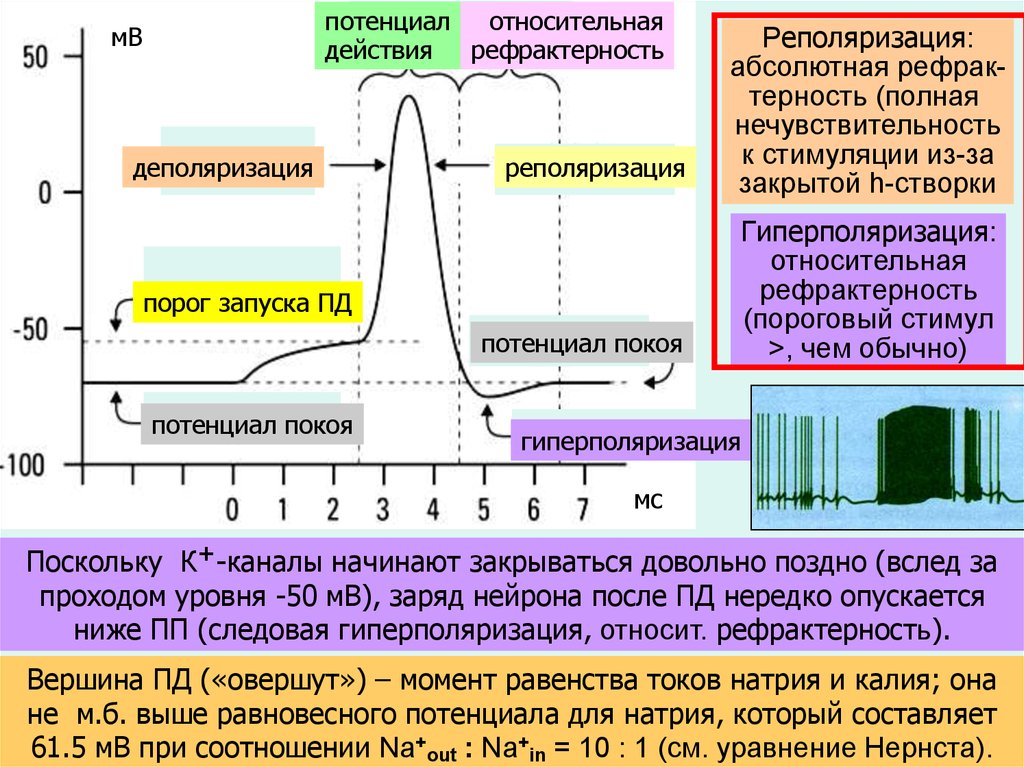
потенциалотносительная
действия рефрактерность
мВ
деполяризация
реполяризация
порог запуска ПД
потенциал покоя
потенциал покоя
Реполяризация:
абсолютная рефрактерность (полная
нечувствительность
к стимуляции из-за
закрытой h-створки
Гиперполяризация:
относительная
рефрактерность
(пороговый стимул
>, чем обычно)
гиперполяризация
мс
Поскольку К+-каналы начинают закрываться довольно поздно (вслед за
проходом уровня -50 мВ), заряд нейрона после ПД нередко опускается
ниже ПП (следовая гиперполяризация, относит. рефрактерность).
Вершина ПД («овершут») – момент равенства токов натрия и калия; она
не м.б. выше равновесного потенциала для натрия, который составляет
43
61.5 мВ при соотношении Na+out : Na+in = 10 : 1 (см. уравнение Нернста).
44.
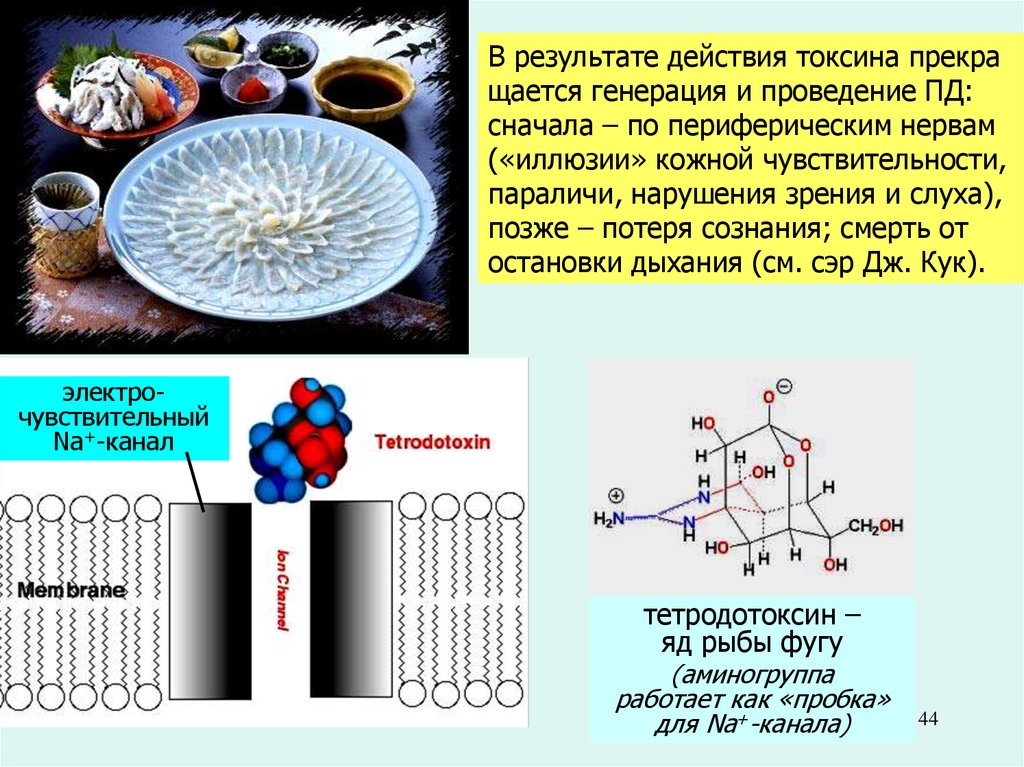
В результате действия токсина прекращается генерация и проведение ПД:
сначала – по периферическим нервам
(«иллюзии» кожной чувствительности,
параличи, нарушения зрения и слуха),
позже – потеря сознания; смерть от
остановки дыхания (см. сэр Дж. Кук).
электрочувствительный
Na+-канал
тетродотоксин –
яд рыбы фугу
(аминогруппа
работает как «пробка»
для Na+-канала)
44
45.
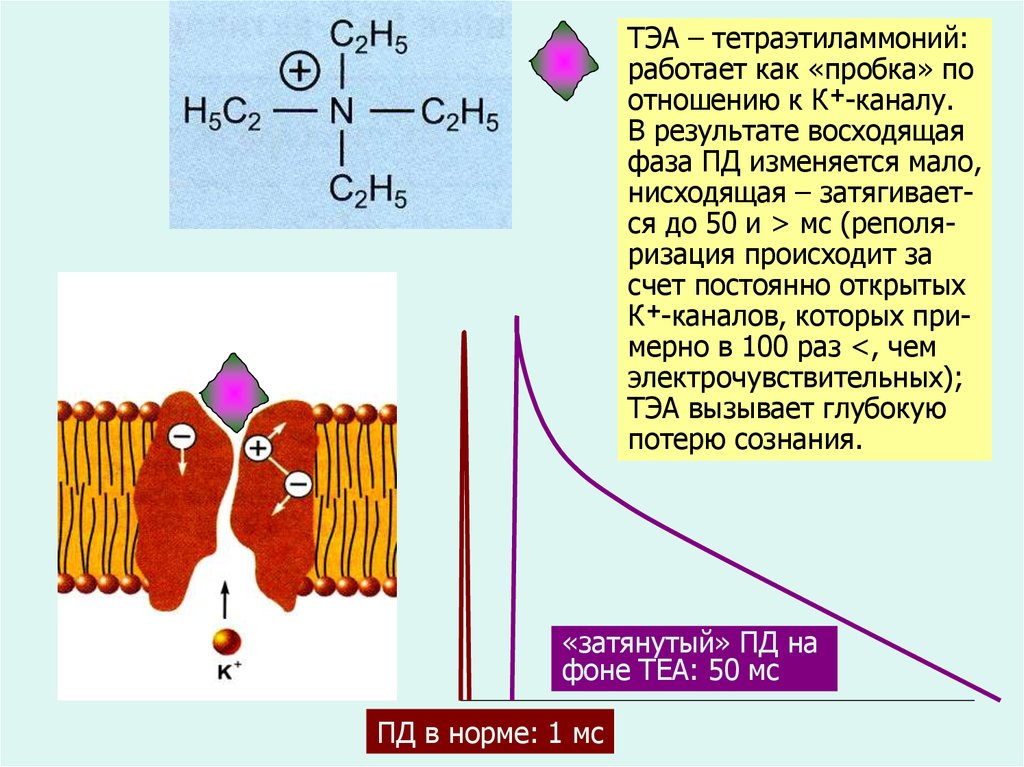
ТЭА – тетраэтиламмоний:работает как «пробка» по
отношению к К+-каналу.
В результате восходящая
фаза ПД изменяется мало,
нисходящая – затягивается до 50 и > мс (реполяризация происходит за
счет постоянно открытых
К+-каналов, которых примерно в 100 раз <, чем
электрочувствительных);
ТЭА вызывает глубокую
потерю сознания.
«затянутый» ПД на
фоне ТЕА: 50 мс
ПД в норме: 1 мс
46. Проведение возбуждения в нервных волокнах
• При распространении возбуждения по безмиелиновому нервномуволокну местные токи, возникающие между возбужденным участком
(заряженным отрицательно), и невозбужденными (заряженными
положительно) деполяризуют мембрану до критического уровня, что
приводит к генерации ПД в соседних участках (I - непрерывное
проведение).
• Наличие у миелиновых волокон оболочки (с высоким электрическим
сопротивлением), приводит к тому, что местные токи могут возникать
только между перехватами Ранвье (лишенных миелина), где и
происходит генерация ПД. Возбуждение «перепрыгивает» через
участки нервного волокна, покрытые миелином (II – сальтаторное,
скачкообразное проведение).
47.
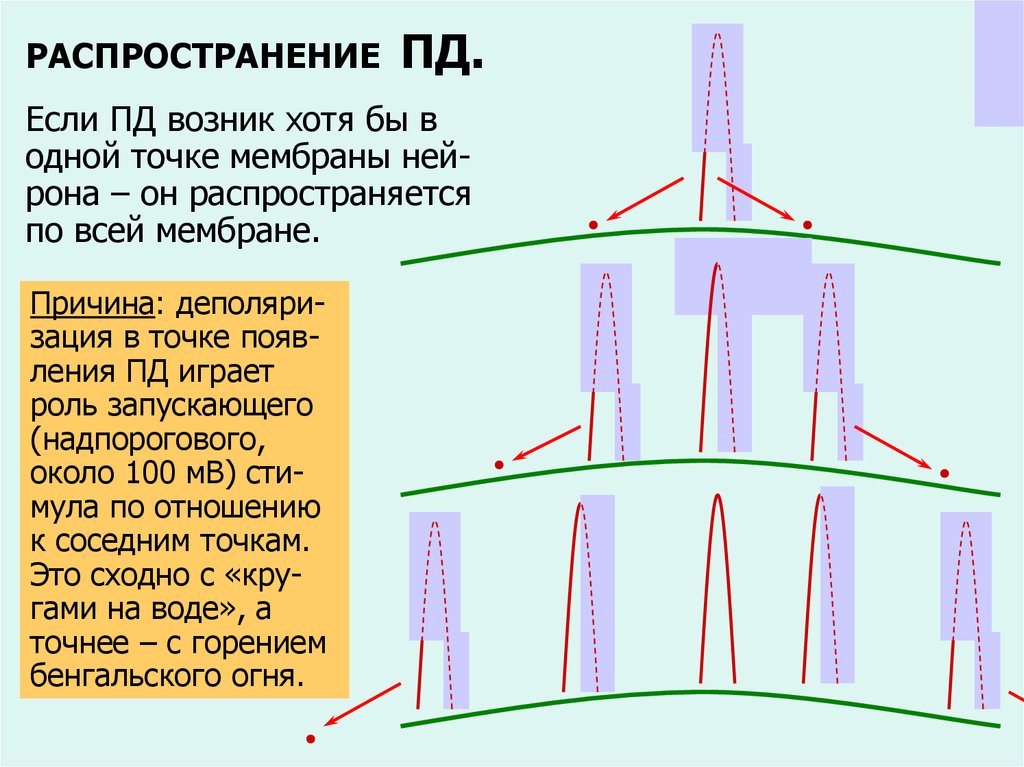
РАСПРОСТРАНЕНИЕПД.
Если ПД возник хотя бы в
одной точке мембраны нейрона – он распространяется
по всей мембране.
Причина: деполяризация в точке появления ПД играет
роль запускающего
(надпорогового,
около 100 мВ) стимула по отношению
к соседним точкам.
Это сходно с «кругами на воде», а
точнее – с горением
бенгальского огня.
стимул
48.
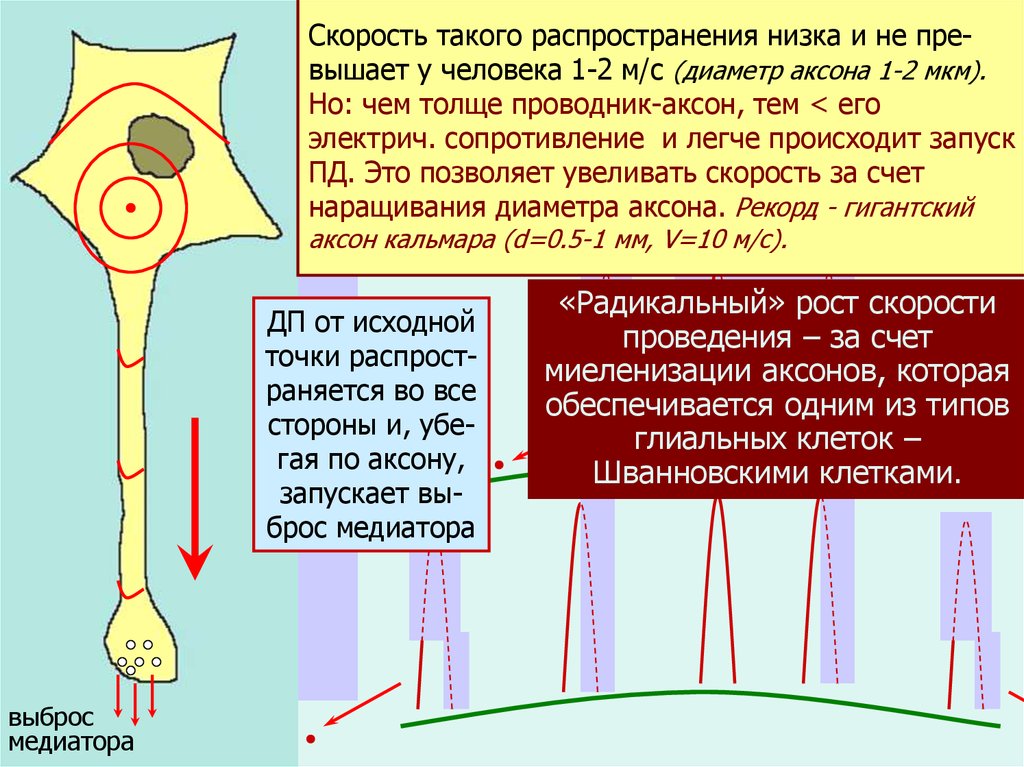
Скорость такого распространения низка и не преРАСПРОСТРАНЕНИЕвышаетПД.
у человека 1-2 м/с (диаметр аксона 1-2 мкм).
Но: чем толще проводник-аксон, тем < его
Если ПД возник хотя
бы в
электрич. сопротивление и легче происходит запуск
одной точке мембраны
нейПД. Это позволяет увеливать скорость за счет
рона – он распространяется
наращивания диаметра аксона. Рекорд - гигантский
по всей мембране.аксон кальмара (d=0.5-1 мм, V=10 м/с).
Причина: деполяриДП от исходной
зация в точке появточки распростления ПД играет
раняется во все
роль запускающего
(надпорогового, стороны и, убегая по аксону,
около 100 мВ) стизапускает вымула по отношению
брос медиатора
к соседним точкам.
Это сходно с «кругами на воде», а
точнее – с горением
бенгальского огня.
выброс
медиатора
стимул
«Радикальный»
рост скорости
проведения – за счет
миеленизации аксонов, которая
обеспечивается одним из типов
глиальных клеток –
Шванновскими клетками.
49.
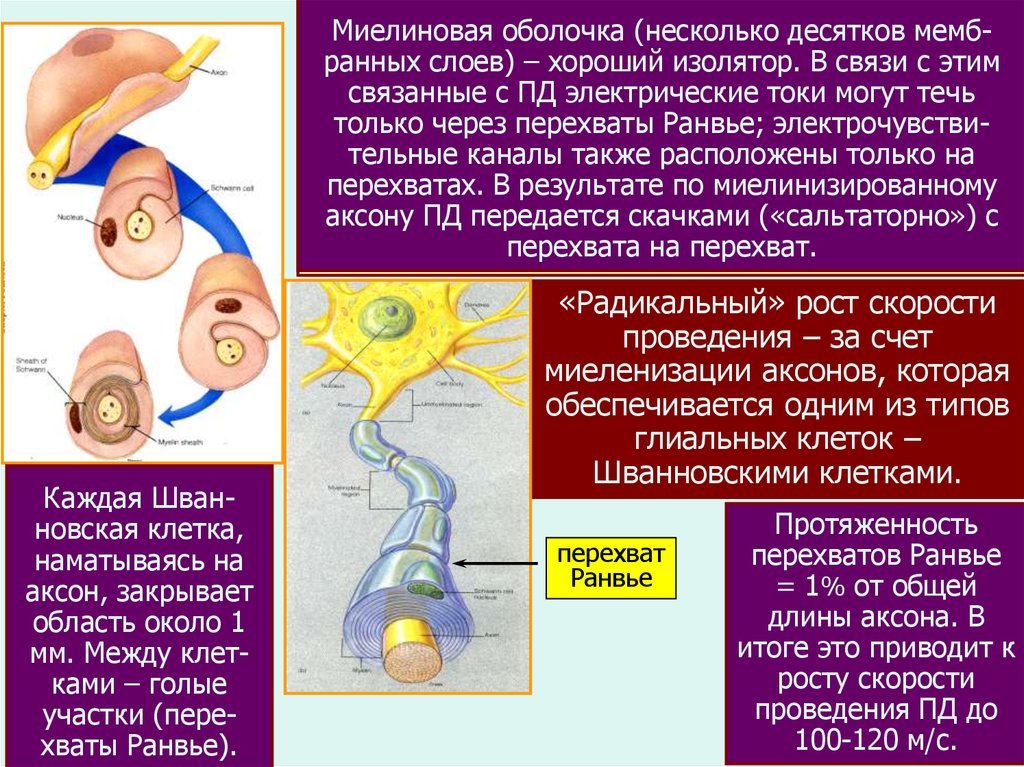
Миелиноваяоболочка
(нескольконизка
десятков
Скорость
такого
распространения
и не мембпреранныхуслоев)
– хороший
В связи
этим
вышает
человека
1-2 м/с изолятор.
(диаметр аксона
1-2 смкм).
ПД электрическиетем
токи
Но:связанные
чем толщес проводник-аксон,
< могут
его течь
только через перехваты Ранвье; электрочувствиэлектрич. сопротивление и легче происходит запуск
тельные каналы также расположены только на
ПД.
Это позволяет
увеливать
за счет
перехватах.
В результате
по скорость
миелинизированному
наращивания
диаметра аксона.
Рекорд
- гигантский с
аксону ПД передается
скачками
(«сальтаторно»)
аксон кальмара (d=0.5-1
мм,на
V=10
м/с).
перехвата
перехват.
Каждая Шванновская клетка,
наматываясь на
аксон, закрывает
область около 1
мм. Между клетками – голые
участки (перехваты Ранвье).
«Радикальный» рост скорости
проведения – за счет
миеленизации аксонов, которая
обеспечивается одним из типов
глиальных клеток –
Шванновскими клетками.
перехват
Ранвье
Протяженность
перехватов Ранвье
= 1% от общей
длины аксона. В
итоге это приводит к
росту скорости
проведения ПД до
100-120 м/с.
50.
Батрахотоксин: токсин кожи некоторыхлягушек-листолазов; модифицированный стероидный гормон насекомых (?).
Токсин проникает внутрь клетки и связывается с h-створками
в тот момент, когда они открыты. В результате электрочувствительные Na+-каналы не закрываются. Начинается тотальный вход Na+, проводящий к быстрой потере нейроном как
ПП, так и способности проводить ПД (одна лягушка – от 10 до
100 смертельных доз).
51.
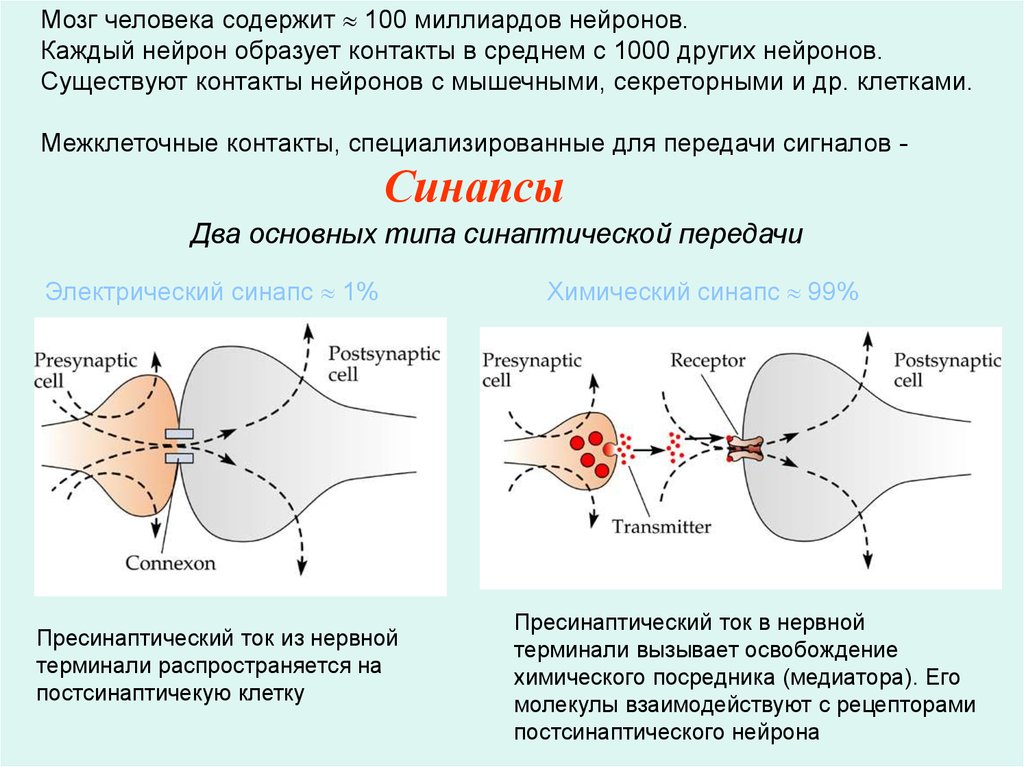
Мозг человека содержит 100 миллиардов нейронов.Каждый нейрон образует контакты в среднем с 1000 других нейронов.
Существуют контакты нейронов с мышечными, секреторными и др. клетками.
Межклеточные контакты, специализированные для передачи сигналов -
Синапсы
Два основных типа синаптической передачи
Электрический синапс 1%
Пресинаптический ток из нервной
терминали распространяется на
постсинаптичекую клетку
Химический синапс 99%
Пресинаптический ток в нервной
терминали вызывает освобождение
химического посредника (медиатора). Его
молекулы взаимодействуют с рецепторами
постсинаптического нейрона
52.
Структурные требования к электрическомумеханизму синаптической передачи
1.Тесное прилегание пре- и
постсинаптических мембран
2. Наличие системы каналов
щелевого контакта,
обеспечивающей быстрый
пассивный перенос ионов между
клетками (как правило)
двухсторонний.
В электрическом синапсе
сигнал ослабляется !
Главное преимущество –
высокая скорость передачи
53.
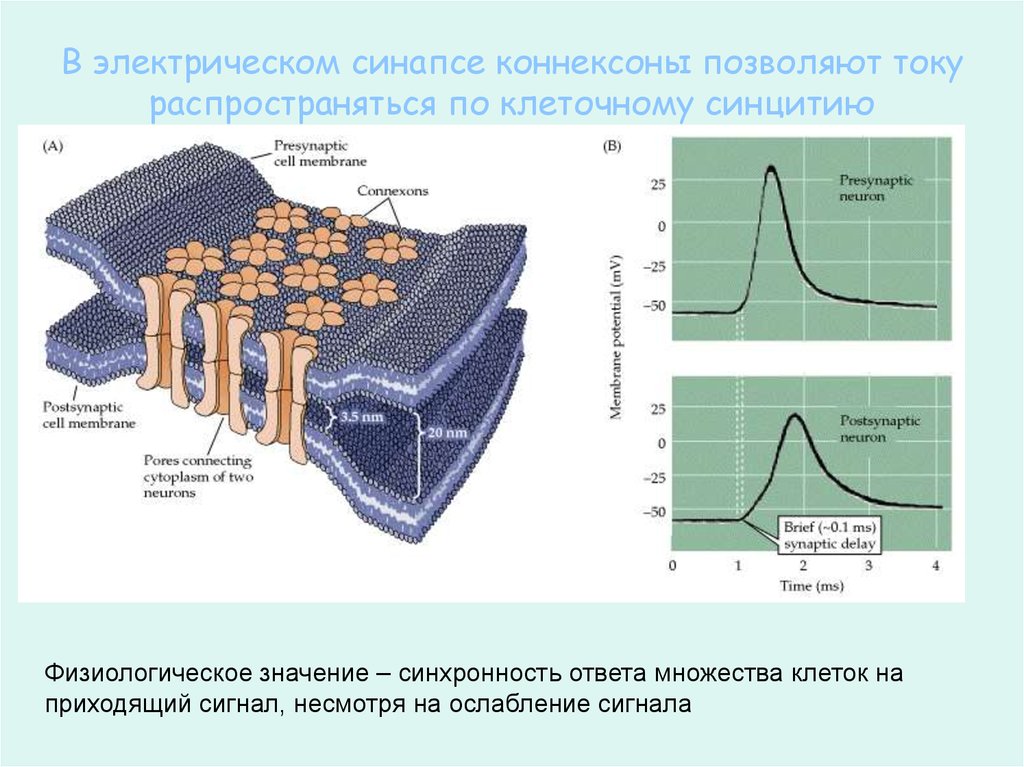
В электрическом синапсе коннексоны позволяют токураспространяться по клеточному синцитию
Физиологическое значение – синхронность ответа множества клеток на
приходящий сигнал, несмотря на ослабление сигнала
54.
Основные свойства электрическихсинапсов
• Менее распространены, чем химические ( .
• Прилегающие мембраны соединены щелевым контактом.
• Ток течет из одной клетки в другую в области щелевых
контактов через широкие каналы, образуемые белками коннексонами.
• Хотя сигнал при этом теряет в амплитуде, но зато сильно
выигрывает в скорости распространения, которая
лимитируется только диффузией.
• Сигналы могут распространяться в обоих направлениях.
• Основная функция – синхронизация электрической активности
в популяции близко расположенных нейронов.
• Не только ионы, но и вещества большего размера, например,
АТФ, могут распространяться этим путем.
55. Структурные требования к химическому механизму синаптической передачи
Взаимодействие трех структур:(а) пресинаптической –электрически
управляемая секреция медиатора;
(б) синаптической щели –
пространство шириной 40-50 нм, где
происходит свободная диффузия
медиатора;
(в) постсинаптической мембраной,
обладающей лиганд-активируемыми
каналами.
В химическом синпсе происходит
двукратная трансформация
сигнала: из электро в хемо и
снова в электро.
Сигнал усиливается, но
возникает синаптическая
задержка
56.
МедиаторыТребования к молекулярным свойствам медиаторов
• Высокая скорость диффузии, а значит низкий молекулярный вес
• Относительная простота и скорость синтеза (небольшое число стадий)
• Доступность исходных продуктов и наличие систем поступления их в нервную клетку
• Невысокие энергетические затраты («дешевизна») на синтез или обратный захват
нейроном
• Возможность повторного использования самого медиатора или непосредственных
продуктов его метаболизма
•В синапсе из окончания аксона при приходе потенциала действия выделяется
вещество-медиатор, которое может возбуждать либо тормозить активность клеткимишени.
Основные медиаторы
главный возбуждающий медиатор ЦНС –
глутаминовая кислота;
главный тормозный медиатор ЦНС – гаммааминомасляная кислота (ГАМК);
медиатор нервно-мышечных синапсов –
ацетилхолин;
медиаторы вегетативных синапсов – ацетилхолин и
норадреналин.
57.
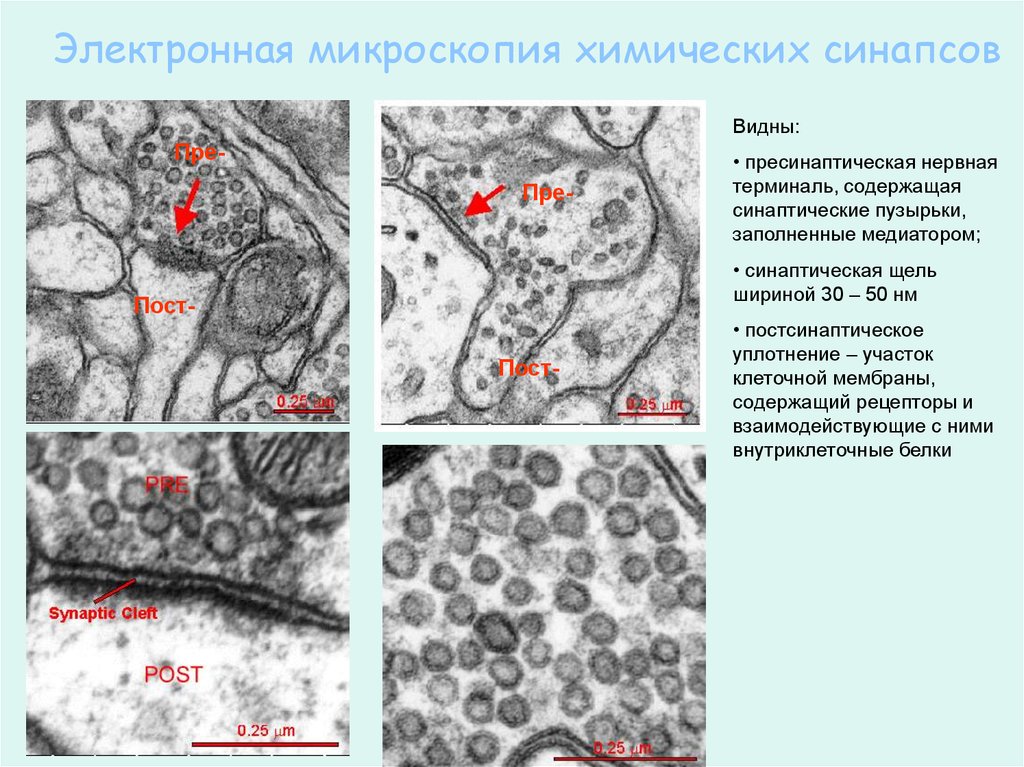
Электронная микроскопия химических синапсовВидны:
ПреПре-
• пресинаптическая нервная
терминаль, содержащая
синаптические пузырьки,
заполненные медиатором;
• синаптическая щель
шириной 30 – 50 нм
ПостПост-
• постсинаптическое
уплотнение – участок
клеточной мембраны,
содержащий рецепторы и
взаимодействующие с ними
внутриклеточные белки
58.
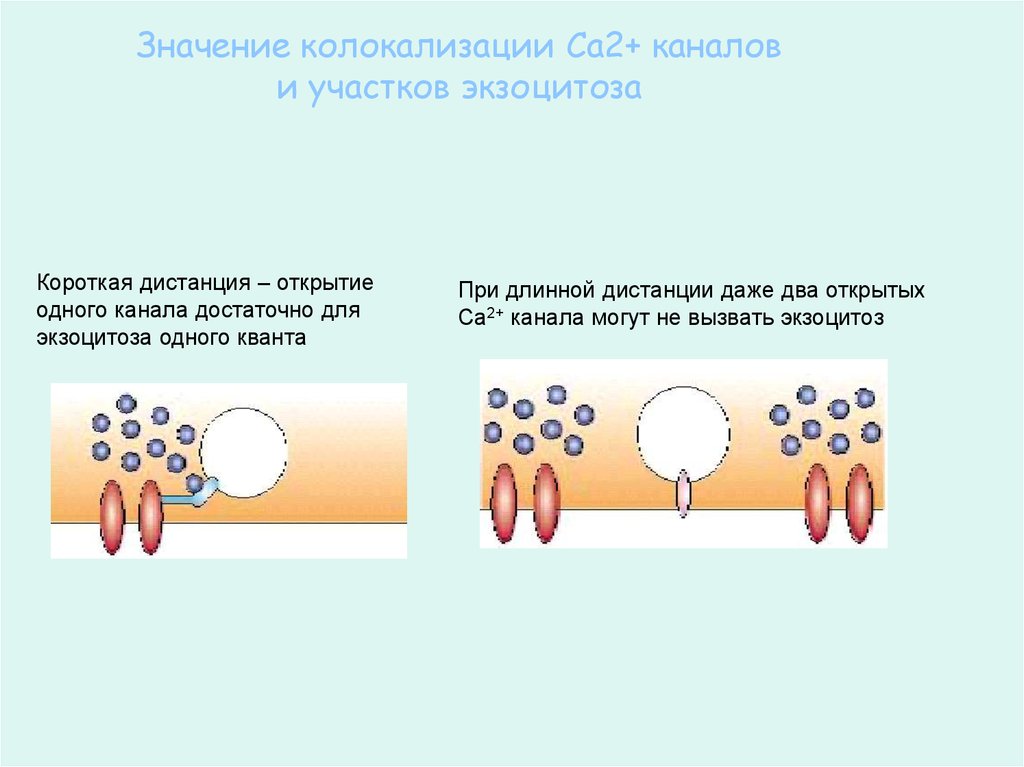
Значение колокализации Са2+ каналови участков экзоцитоза
Короткая дистанция – открытие
одного канала достаточно для
экзоцитоза одного кванта
При длинной дистанции даже два открытых
Са2+ канала могут не вызвать экзоцитоз
59.
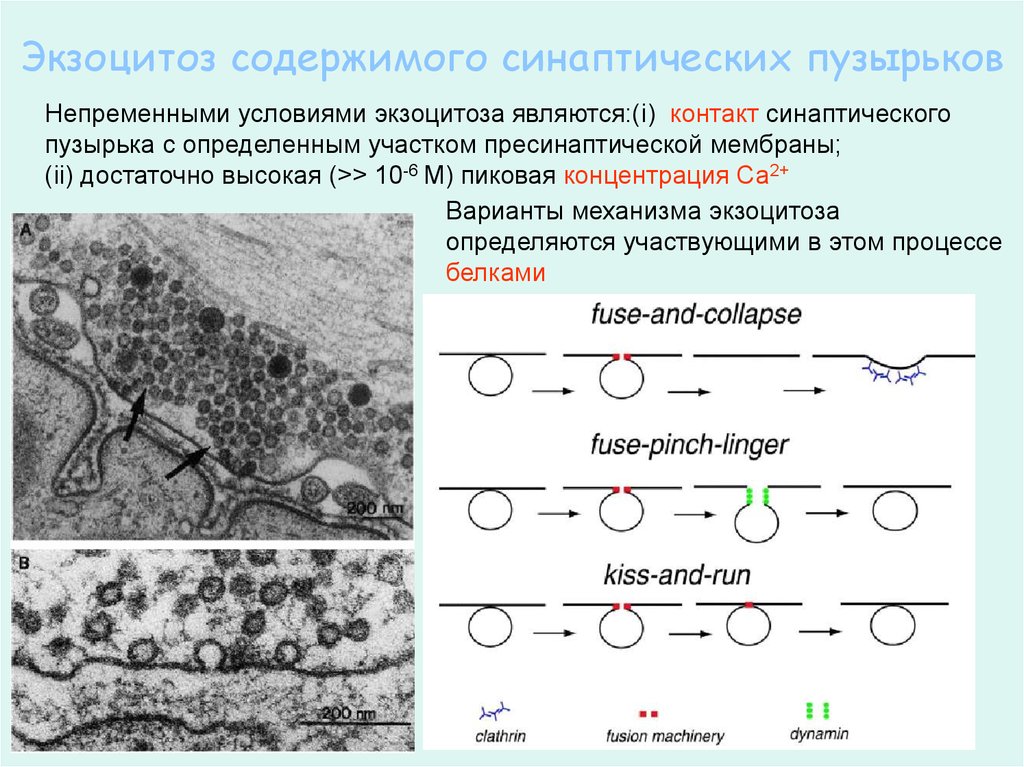
Экзоцитоз содержимого синаптических пузырьковНепременными условиями экзоцитоза являются:(i) контакт синаптического
пузырька с определенным участком пресинаптической мембраны;
(ii) достаточно высокая (>> 10-6 M) пиковая концентрация Са2+
Варианты механизма экзоцитоза
определяются участвующими в этом процессе
белками
60.
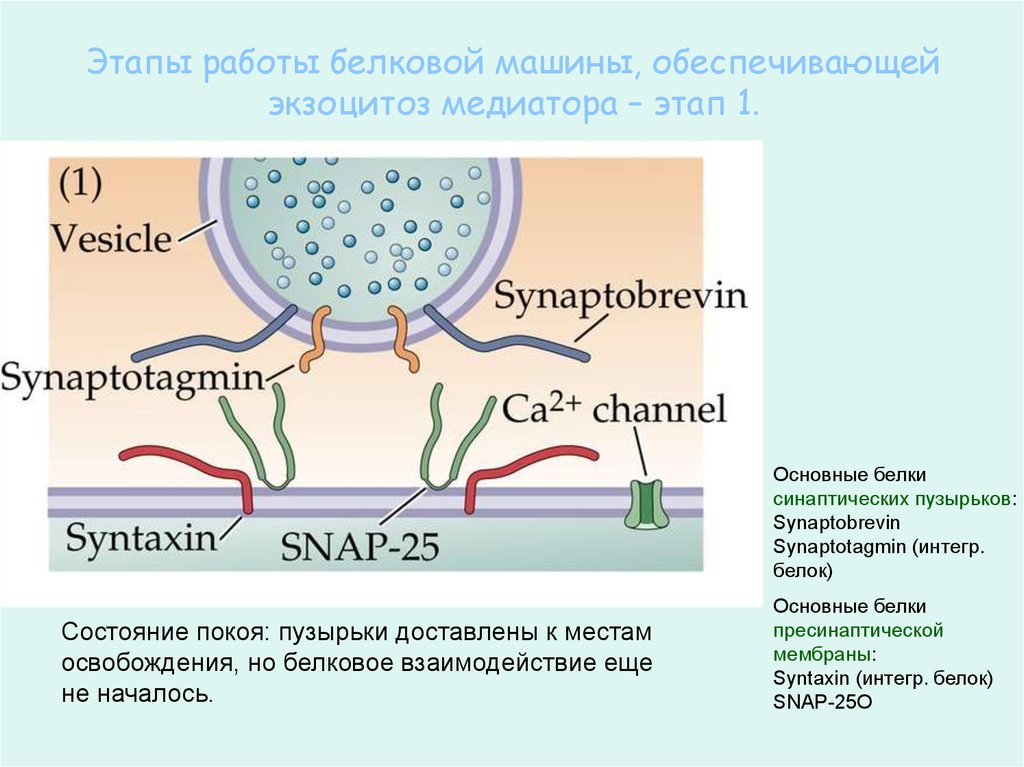
Этапы работы белковой машины, обеспечивающейэкзоцитоз медиатора – этап 1.
Основные белки
синаптических пузырьков:
Synaptobrevin
Synaptotagmin (интегр.
белок)
Состояние покоя: пузырьки доставлены к местам
освобождения, но белковое взаимодействие еще
не началось.
Основные белки
пресинаптической
мембраны:
Syntaxin (интегр. белок)
SNAP-25О
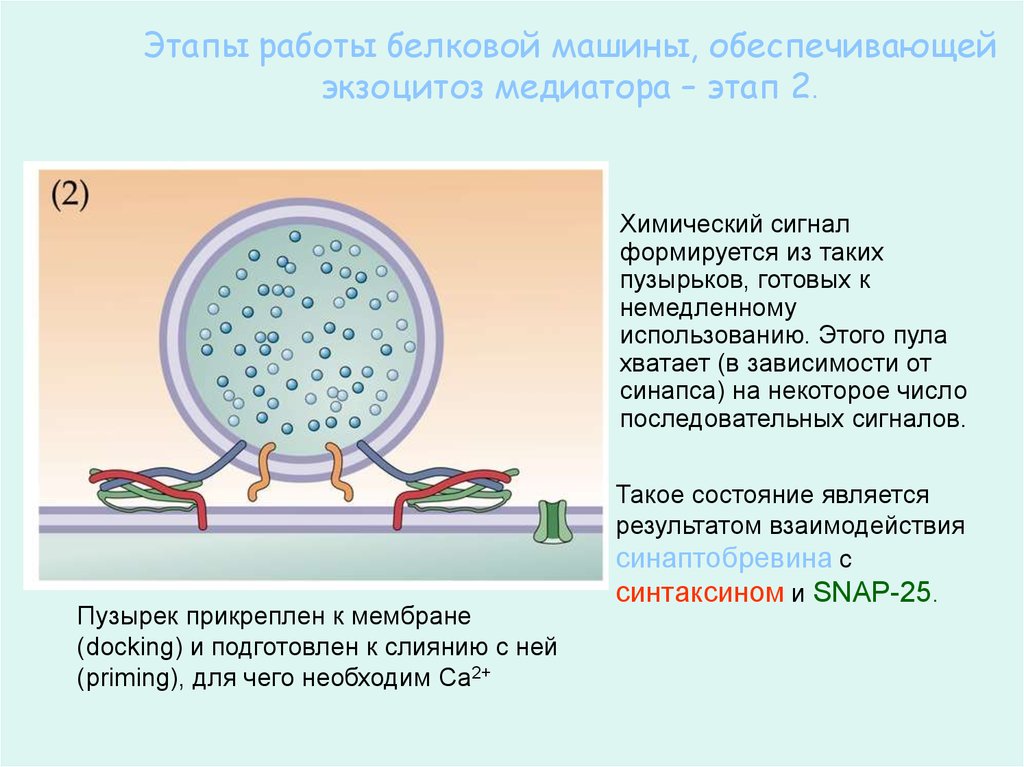
61.
Этапы работы белковой машины, обеспечивающейэкзоцитоз медиатора – этап 2.
Химический сигнал
формируется из таких
пузырьков, готовых к
немедленному
использованию. Этого пула
хватает (в зависимости от
синапса) на некоторое число
последовательных сигналов.
Пузырек прикреплен к мембране
(docking) и подготовлен к слиянию с ней
(priming), для чего необходим Са2+
Такое состояние является
результатом взаимодействия
синаптобревина с
синтаксином и SNAP-25.
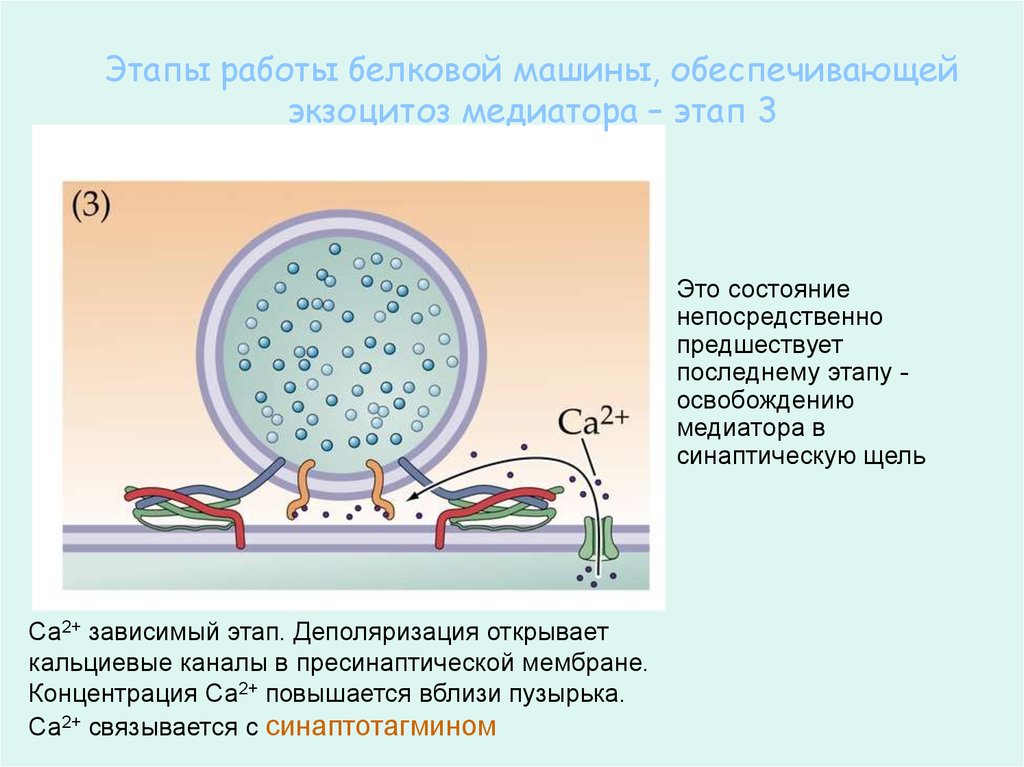
62.
Этапы работы белковой машины, обеспечивающейэкзоцитоз медиатора – этап 3
Это состояние
непосредственно
предшествует
последнему этапу освобождению
медиатора в
синаптическую щель
Са2+ зависимый этап. Деполяризация открывает
кальциевые каналы в пресинаптической мембране.
Концентрация Са2+ повышается вблизи пузырька.
Са2+ связывается с синаптотагмином
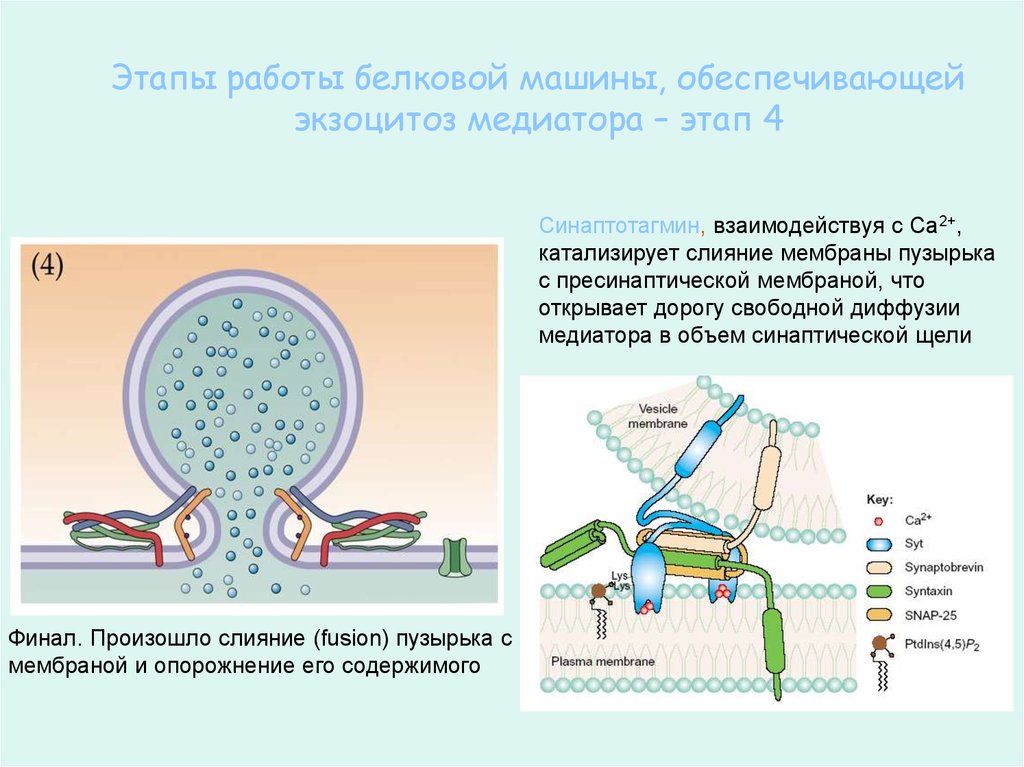
63.
Этапы работы белковой машины, обеспечивающейэкзоцитоз медиатора – этап 4
Синаптотагмин, взаимодействуя с Са2+,
катализирует слияние мембраны пузырька
с пресинаптической мембраной, что
открывает дорогу свободной диффузии
медиатора в объем синаптической щели
Финал. Произошло слияние (fusion) пузырька с
мембраной и опорожнение его содержимого
64.
Разнообразие морфологии нейронов65.
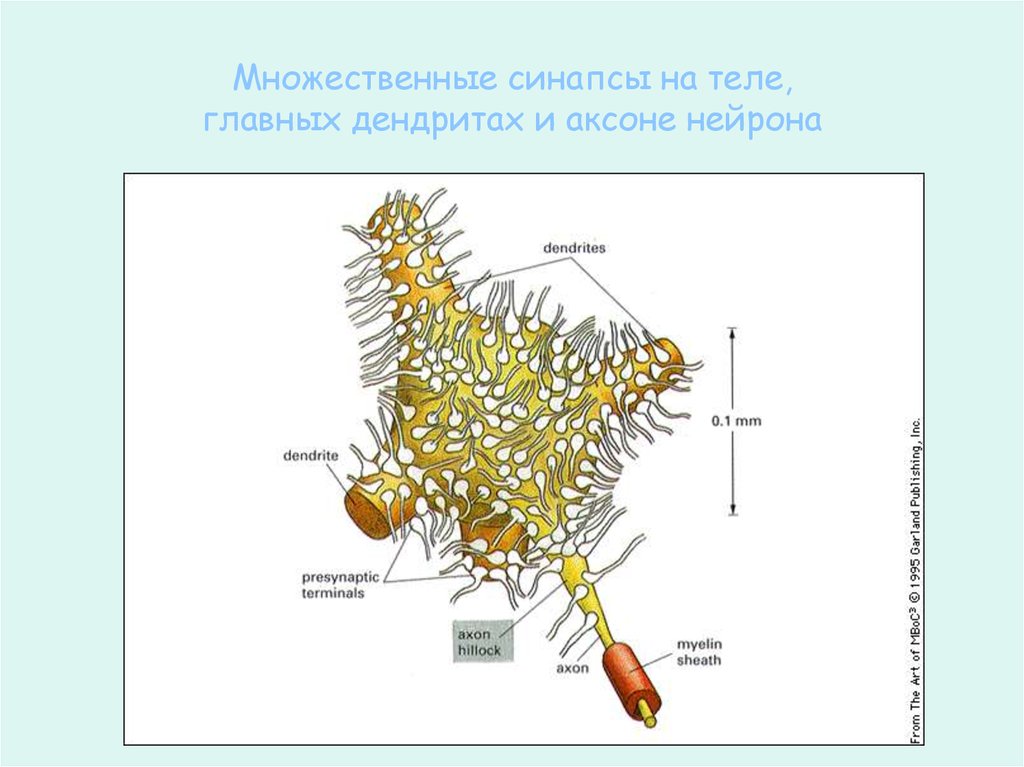
Множественные синапсы на теле,главных дендритах и аксоне нейрона