








































")




")
")

























")
















































")
 Биология
БиологияПохожие презентации:









")
Обмен углеводов
1. Обмен углеводов
UNIVERSITATEA DE STAT DE MEDICINĂ ȘI FARMACIE«NICOLAE TESTEMIȚANU»
CATEDRA BIOCHIMIE ȘI BIOCHIMIE CLINICĂ
Обмен углеводов
Svetlana Protopop
doctor în medicină,
conferențiar universitar
2. Углеводы
- полигидроксикарбонильныеорганические вещества.
•Общая формула – Сm(Н2О)n
•Углеводы составляют около 1% массы
сухого вещества в животных клетках, а в
клетках печени — до 5%.
•В растительных клетках углеводы
составляют до 90% сухой массы.
3. Функции углеводов
1. Энергетическая (4,3 ккал/г )2. Структурная (гликозаминогликаны,
целлюлоза)
3. Запасающая (гликоген, крахмал)
4. Защитная (слизь секретов)
5. Клеточное взаимодействие
6. Определяют специфичность белков
(гликопротеины; пр. – группы крови)
4. Классификация углеводов
•Моносахариды – общая формула(СН2О)n, где n ≥ 3.
•Олигосахариды – 2-10 остатков
моносахаридов.
•Полисахариды – более 10
остатков моносахаридов.
5. Классификация моносахаридов
• В зависимости от числа атомов углеродаразличают:
триозы (3С)
тетрозы (4С)
пентозы (5С)
гексозы (6С)
гептозы (7С)
• В зависимости от положения карбонильной
группы
Альдозы
Кетозы
6. Моносахариды
Триозы:Глицеральдегид и дигидроксиацетон
Пентозы:
Рибоза, дезоксирибоза
Гексозы:
Глюкоза, галактоза, фруктоза
7. Триозы
8. Пентозы
9. Гексозы
HO
C
CH2OH
H
C
OH
HO
C
H
H
C
H
C
C
O
HO
C
H
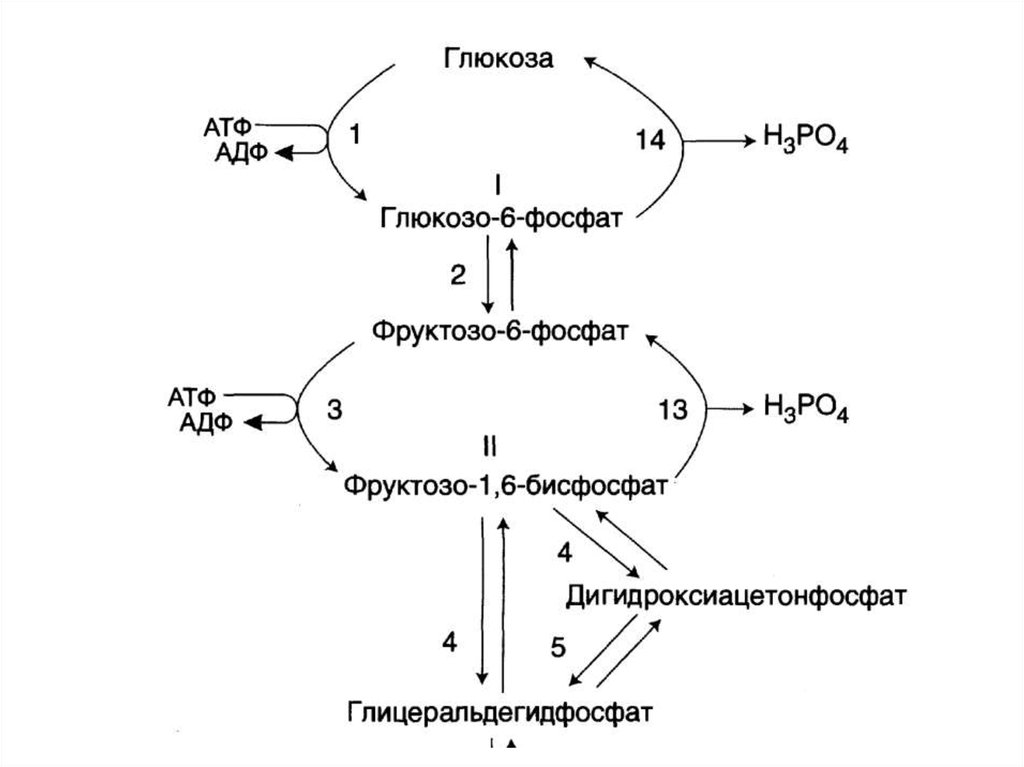
OH
H
C
OH
OH
H
C
OH
CH2OH
CH2OH
D-glucose
D-fructose
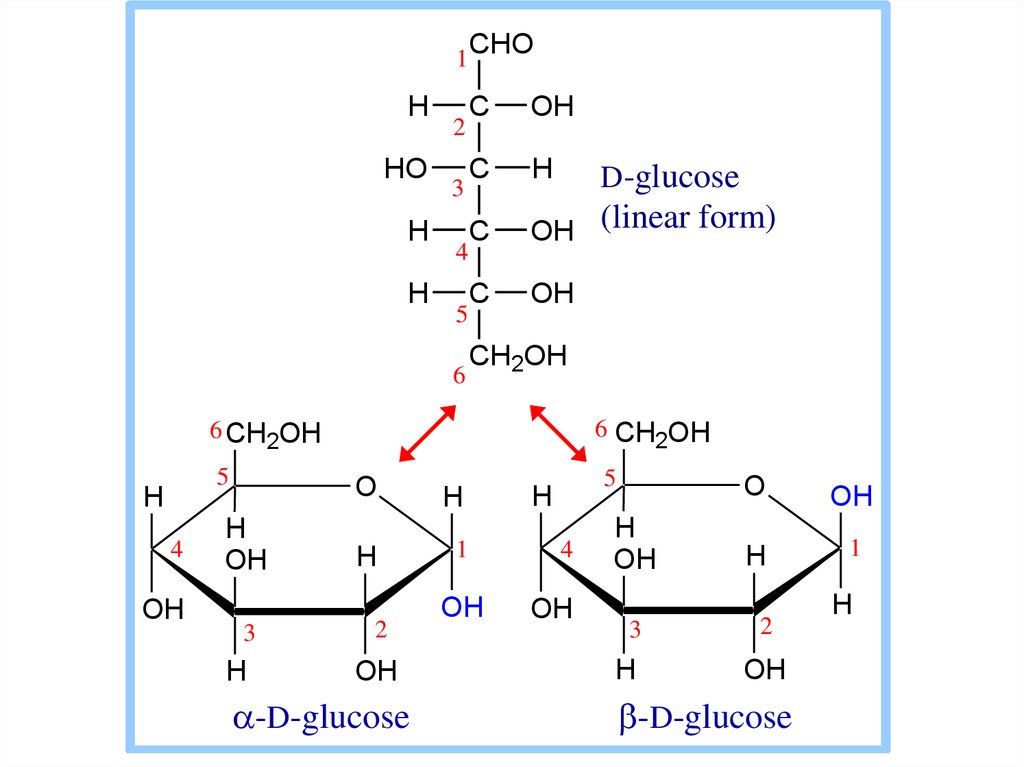
10. Гексозы
11.
1H
HO
H
H
2
3
4
5
6
CHO
C
OH
C
H
C
OH (linear form)
C
OH
D-glucose
CH2OH
6 CH2OH
6 CH2OH
5
H
4
OH
H
OH
3
H
O
H
H
1
2
OH
-D-glucose
OH
5
H
4
OH
H
OH
3
H
O
OH
H
1
2
OH
-D-glucose
H
12. Гексозы
CH2OH1
HO
H
H
2C
O
C
H
C
OH
C
OH
3
4
5
6
HOH2C 6
CH2OH
D-fructose (linear)
H
5
H
1 CH2OH
O
4
OH
HO
2
3
OH
H
-D-fructofuranose
13.
14. Олигосахариды
В зависимости от количества остатковмоносахаридов делятся на:
•Дисахариды
•Трисахариды
•Тетрасахариды и т.д.
Наиболее распространенные в природе –
дисахариды.
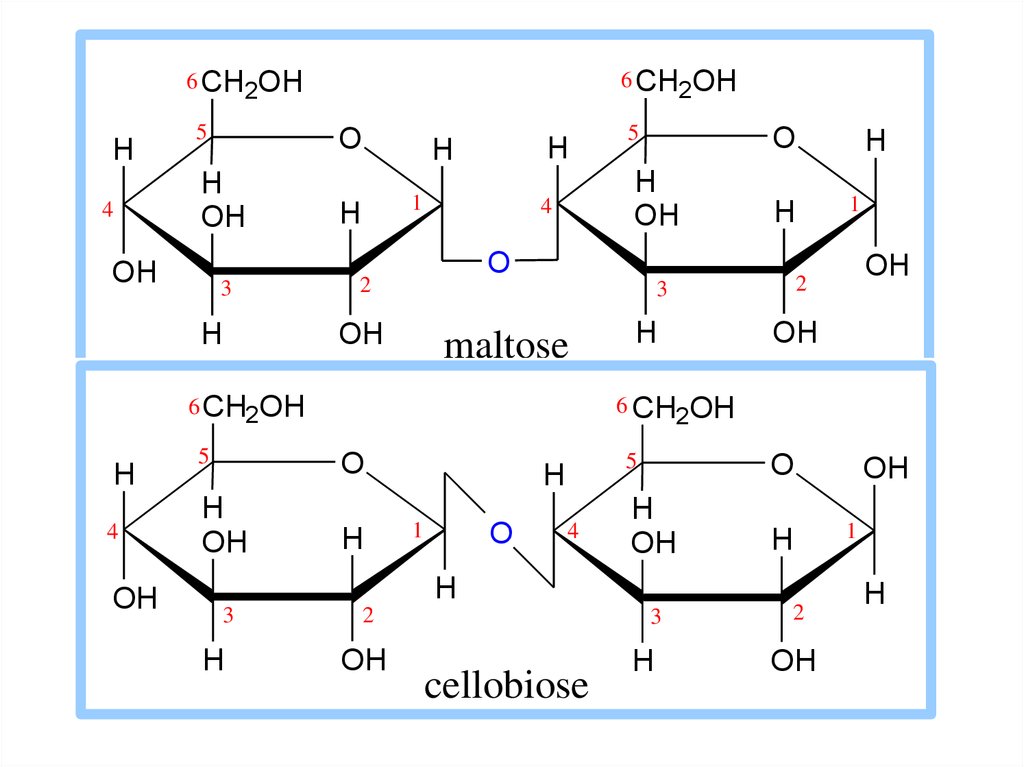
15. Дисахариды
• мальтоза, состоит из двух остатков -глюкозы;• лактоза – молочный сахар (β-глюкоза +
галактоза);
• сахароза – свекловичный сахар ( -глюкоза +
фруктоза).
16.
6 CH2OH6 CH2OH
H
5
O
H
OH
4
OH
3
H
H
H
1
H
4
H
4
OH
H
OH
H
OH
maltose
H
H
1
OH
OH
6 CH
2OH
H
H
1
O
4
5
O
H
OH
H
H
3
H
2
3
O
H
OH
O
O
2
6 CH2OH
5
5
2
OH
3
cellobiose
H
2
OH
OH
1
H
17.
CH2OHH
O H
OH
H
OH
H
OH
O
CH2OH
O
H
OH
CH2OH
H
OH
H
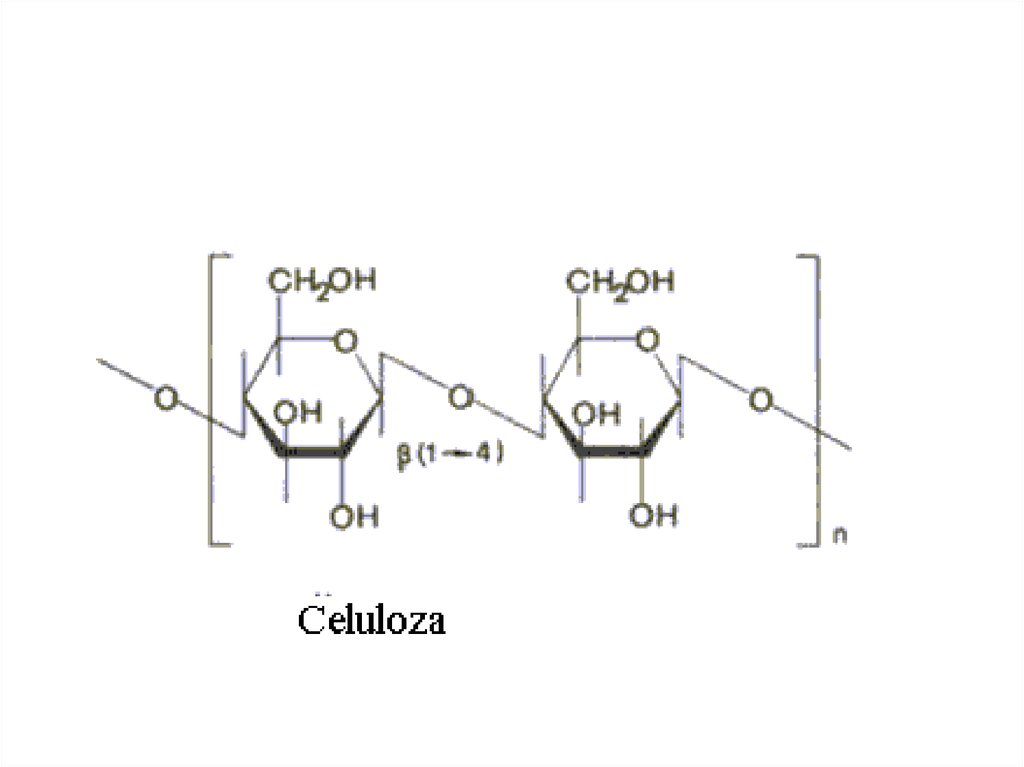
18. Полисахариды
1. Гомополисахариды:• крахмал (запасной углевод растений);
• гликоген (запасной углевод животных);
• целлюлоза (клеточная стенка растений);
2. Гетерополисахариды:
• Гиалуроновая кислота;
• Гепарин, гепаран-сульфат;
• Кератан-сульфат, дерматан-сульфат,
хондроитин-сульфат.
19.
20. Гликоген
21.
22. Гиалуроновая кислота
23. Гепарин
24. Хондроитин-4-сульфат
25. Кератан-сульфат
26. Переваривание углеводов
• Начинается в ротовой полости поддействием α-амилазы слюны, которая
расщепляет α-1,4-гликозидные связи в
крахмале и гликогене.
• Так как действие фермента в ротовой
полости кратковременно, крахмал
расщепляется до декстринов и небольшого
количества мальтозы.
• В кислой среде желудка действие амилазы
прекращается.
27. Переваривание углеводов
• Желудочный сок не содержит ферментов,расщепляющих углеводы.
• Переваривание углеводов продолжается в
тонком кишечнике под действием
панкреатической α-амилазы, которая
расщепляет декстрины до мальтозы.
• Образуется и небольшое количество
изомальтозы и разветвленные
олигосахариды.
• Целлюлоза не расщепляется в ЖКТ человека.
28. Переваривание углеводов
• Переваривание дисахаридов происходит поддействием кишечных ферментов дисахаридаз,
расположенных на наружной поверхности
мембраны энтероцитов.
Мальтоза мальтаза
2 α-глюкозы
Сахароза
Лактоза
Трегалоза
сахараза
лактаза
трегалаза
α-глюкоза + β-фруктоза
β-галактоза + α-глюкоза
2 α-глюкозы
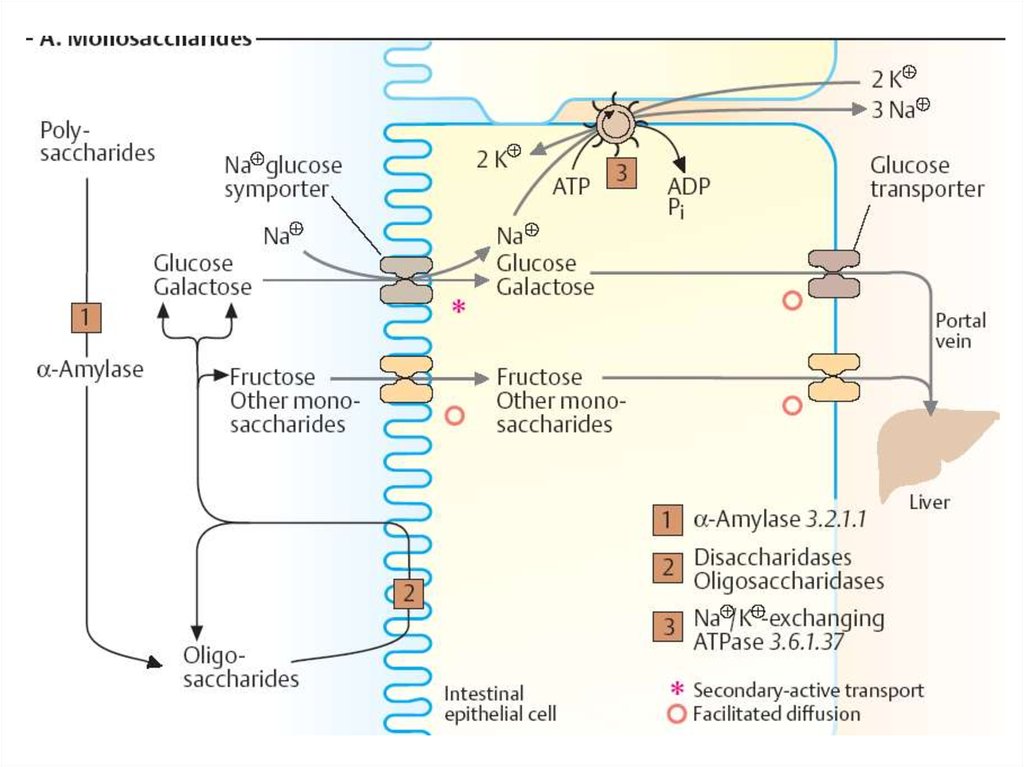
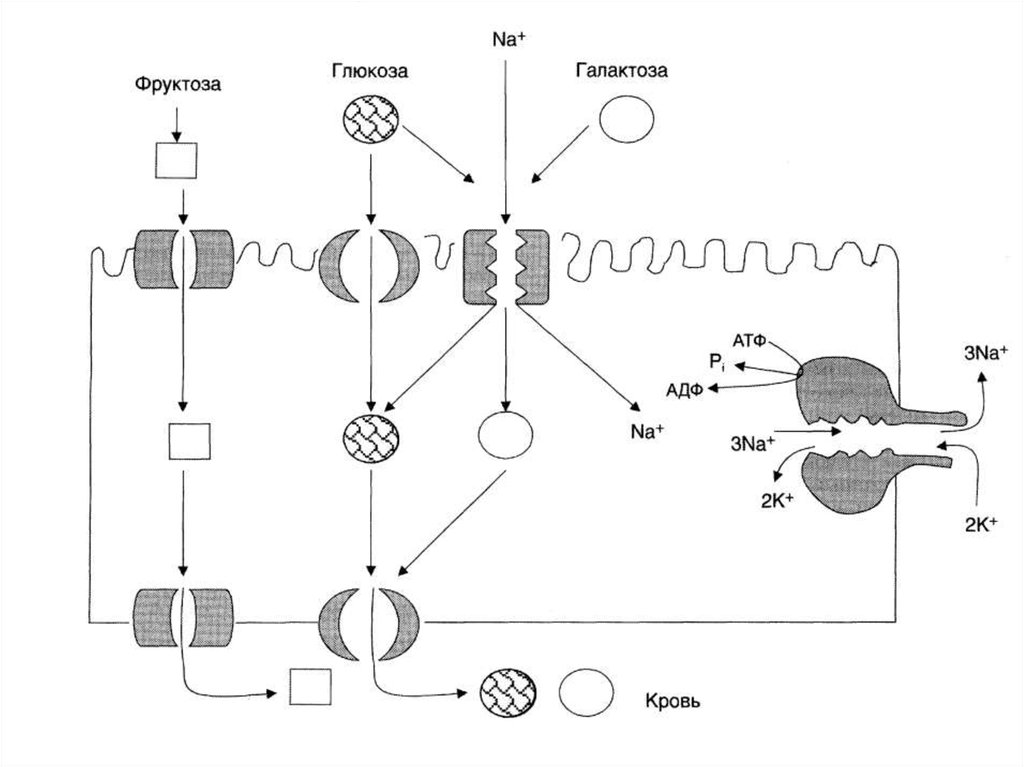
29. Всасывание моносахаридов в тонком кишечнике
• Облегченная диффузия – при высокойконцентрации глюкозы.
• Вторично-активный транспорт (Na+, АТФ)
• Происходит симпорт Na+ и глюкозы из
кишечника в энтероцит посредством белкапереносчика глюкозы. Na+ транспортируется
по градиенту концентрации, а глюкоза –
против градиента концентрации.
30. Всасывание моносахаридов в тонком кишечнике
• После всасывания глюкоза покидаетэнтероцит через мембрану, обращенную к
кровеносному капилляру, с помощью
облегченной диффузии.
• Na+ выходит в просвет кишечника в обмен на
ионы К+ за счет работы Na+ ,К+- АТФ-азы.
31.
32.
33. Транспорт глюкозы из крови в клетки
• Происходит путем облегченной диффузии поградиенту концентрации, за исключением
мышц и жировой ткани, где диффузия
стимулируется инсулином.
• Транспортеры глюкозы (ГЛЮТ):
• ГЛЮТ-1 – транспорт глюкозы в эритроциты
• ГЛЮТ-2 – транспорт глюкозы из энтероцитов
в кровь; транспорт глюкозы в гепатоциты и βклетки поджелудочной железы
34. Транспорт глюкозы из крови в клетки
• ГЛЮТ-3 – клетки нервных тканей• ГЛЮТ-4 – транспорт глюкозы в мышцы и
жировую ткань (инсулинзависимый).
• ГЛЮТ-4 находится в цитозольных везикулах.
Инсулин стимулирует транспорт везикул,
содержащих ГЛЮТ-4, на плазматическую
мембрану. Происходит слияние везикул с
мембраной и встраивание транспортеров в
мембрану.
• Увеличение количества ГЛЮТ-4 приводит
к усилению транспорта глюкозы в клетку.
35. Нарушения переваривания и всасывания углеводов
• Непереносимость лактозы:Причина – недостаточность лактазы
• наследственная
• приобретенная
• Клинические проявления:
• Осмотическая диарея
• Метеоризм
• Спазмы и боли в животе.
• Реже – непереносимость сахарозы и
трегалозы
36. Использование глюкозы в тканях
Гликолиз
Синтез гликогена
Синтез пентоз
Синтез другим моносахаридов →
гетерополисахариды
• Синтез заменимых аминокислот
• Синтез липидов
37. Фосфорилирование глюкозы
38. Отличия глюкокиназы и гексокиназы
ЛокализацияСубстрат
Сродство к
субстрату
Регуляция
Инсулин
Глюкокиназа
Печень
Глюкоза
Низкое –↑КМ
=10 ммоль/л)
-
индуцирует
Гексокиназа
Др. органы
+ др. гексозы
Высокое –↓КМ
= 0,1 ммоль/л
Ингибитор –
Г-6-Р
-
39. Обмен гликогена
• Гликогеногенез (синтезгликогена)
• Гликогенолиз (распад
гликогена, мобилизация
гликогена)
• Гликоген запасается в
печени и в мышцах.
• 5% от массы печени
• 1% от массы мышц
• 450 г гликогена
40. Синтез гликогена
• Происходит интенсивно в печени и вмышцах после приема пищи.
Глюкоза
Глюкокиназа
Гексокиназа
АТФ
Глюкоза1-фосфат
ФосфоглюкоГлюкозамутаза
6-фосфат
АДФ
УДФ-глюкозопирофосфорилаза
УТФ
УДФ-глюкоза
Н4Р2О7
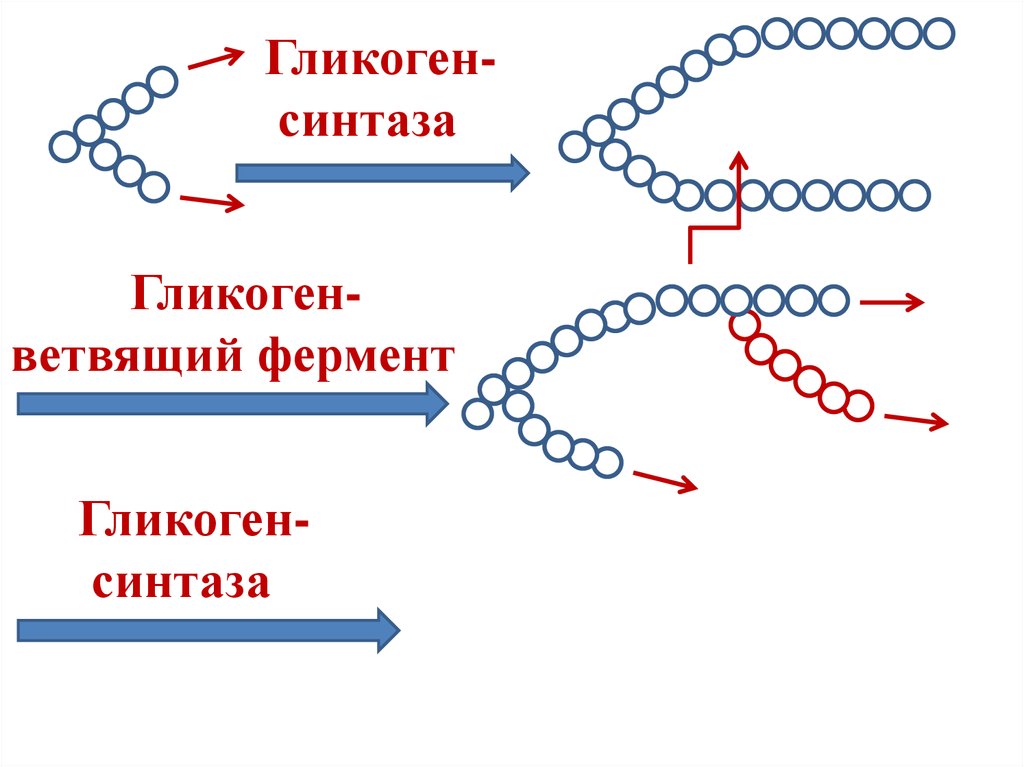
41.
Гликоген (n глюкоз) + УДФ-глюкозаГликогенсинтаза
Гликоген (n+1 глюкоза) + УДФ
Гликогенсинтаза образует только
α-1,4-гликозидные связи.
α-1,6-гликозидные связи образуются
гликоген-ветвящим ферментом
(амило-1,4→1.6-гликозилтрансфераза).
42.
ГликогенсинтазаГликогенветвящий фермент
Гликогенсинтаза
43. Образование гликоген-затравки
+Гликозилтрансферазная
активность гликогенина
+ UDP
44. Гликоген
45. Регуляция биосинтеза гликогена
• Регуляторным ферментом синтезагликогена является гликогенсинтаза.
• Инсулин активирует гликогенсинтазу
(дефосфорилирование фермента).
• Адреналин, глюкагон ингибируют
гликогенсинтазу (фосфорилирование
фермента).
46. Регуляция гликогенсинтазы
47. Регуляция гликогенсинтазы
48. Регуляция синтеза гликогена в мышцах
49. Гликогенолиз (мобилизация гликогена)
Гликоген (n глюкоз) + Н3РО4Гликогенфосфорилаза
Фосфоглюкомутаза
ГлюкозаГликоген (n-1 глюкоз) + 1-фосфат
ГлюкозоГлюкоза- 6-фосфатаза
6-фосфат
Глюкоза
Печень
Мышцы
Гликолиз
Н2О
Н3РО4
Кровь
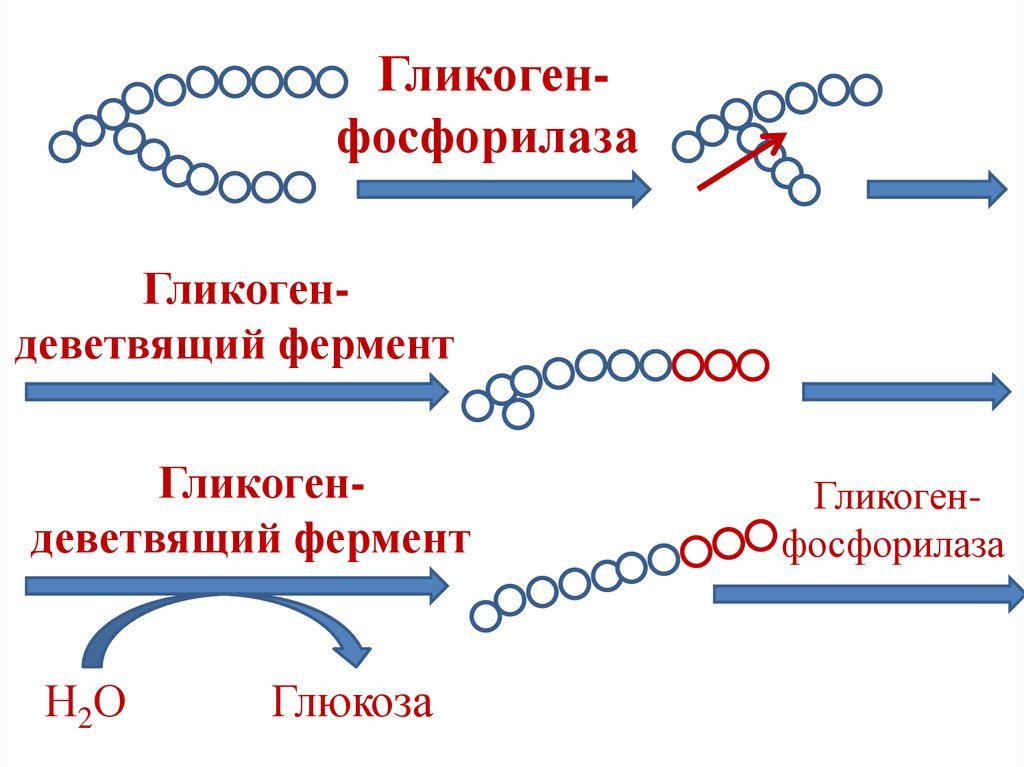
50. Гликогенолиз
• Гликоген-фосфорилаза расщепляеттолько α-1,4-гликозидные связи.
• α-1,6-гликозидные связи расщепляются
гликоген-деветвящим ферментом
(олиго-1,6→1,4-гликантрансфераза).
• Фермент обладает:
1. трансферазной активностью;
2. 1→6-гликозидазной активностью.
51.
ГликогенфосфорилазаГликогендеветвящий фермент
Гликогендеветвящий фермент
Н2О
Глюкоза
Гликогенфосфорилаза
52. Регуляция гликогенолиза
• Регуляторным ферментом распада гликогенаявляется гликогенфосфорилаза.
• Адреналин, глюкагон активируют
гликогенфосфорилазу (фосфорилирование
фермента).
• Инсулин ингибирует гликогенфосфорилазу
(дефосфорилирование фермента).
53. Регуляция гликогенфосфорилазы
54. Гликогеновые болезни
• Наследственные болезни, обусловленныеснижением или отсутствием активности
ферментов, участвующих в распаде или
синтезе гликогена.
• Гликогенозы
• Агликогенозы
55. Болезнь Гирке (I тип)
• Причина – наследственный дефект глюкозо6-фосфатазы.• Печеночная форма.
• Гепатомегалия, увеличение почек
• Отставание в развитии
• Судороги
• Гипогликемия
• Гипертриглицеридемия
• Гиперурикемия
• Ацидоз
56. Болезнь Помпе (II тип)
• Причина – наследственный дефектлизосомальной α-1,6-гликозидазы.
• Смешанная форма
• Генерализованное накопление гликогена в
лизосомах, а затем в цитозоле.
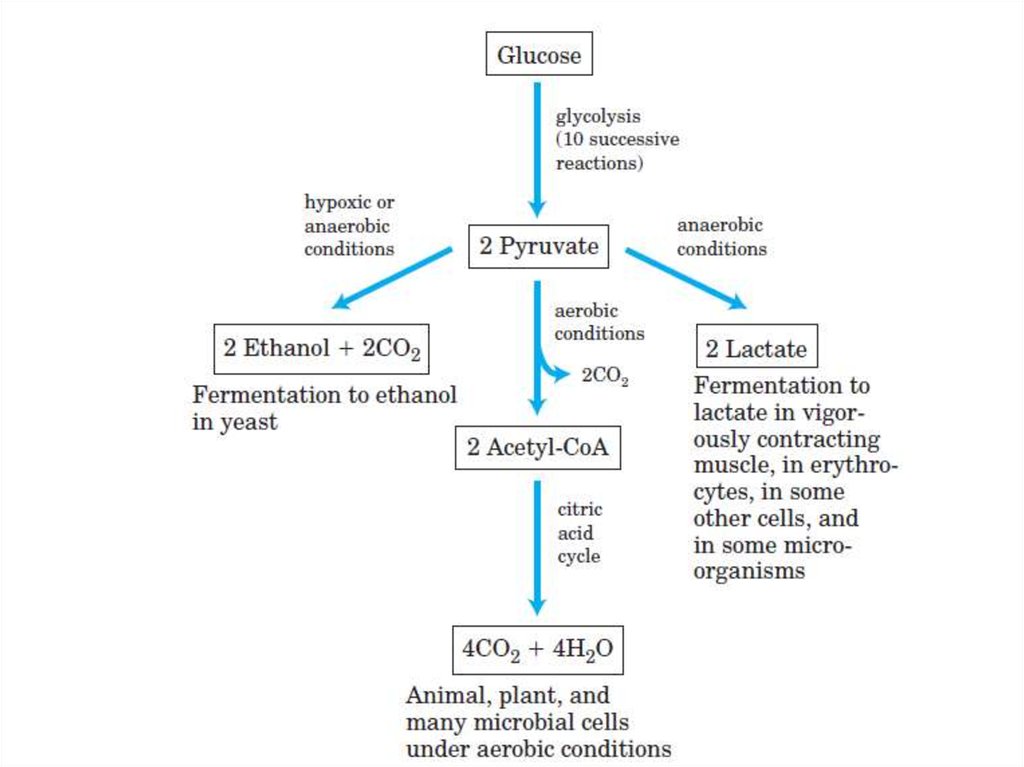
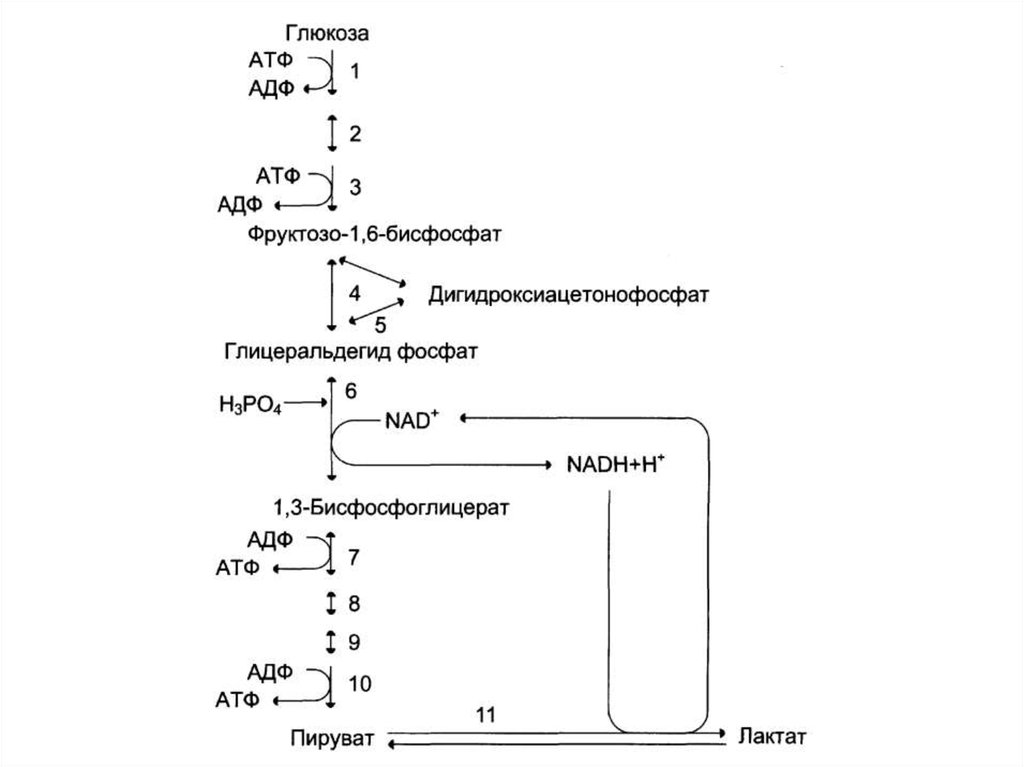
57. Гликолиз
• Окисление глюкозы (образование энергии)• Аэробное окисление глюкозы (до СО2 и Н2О).
• Анаэробный распад глюкозы (до лактата).
58.
59. Гликолиз
60.
61. Гликолиз
62. Гликолиз
63. Гликолиз
64. Гликолиз
65. Гликолиз
66.
67. Суммарная реакция анаэробного гликолиза
Глюкоза + 2АТФ + 2НАД+ + 2Н3РО4 +2
+ 4АДФ + 2НАДН+2Н+
2Лактат + 2АДФ +
2
+ 2НАДН+2Н+ + 4АТФ + 2НАД+ + 2Н2О
Глюкоза + 2АДФ + 2Н3РО4
+
2Лактат + 2АТФ
2Н2О
68. Схема аэробного гликолиза
69.
Глюкоза- 2АТФ
8 АТФ
2 ГА-3-Р
4 АТФ
2НАДН
6 АТФ
24 АТФ
38 АТФ
2 Пируват
2 Ацетил-КоА
2 цикла
Кребса
2НАДН
6 НАДН
2ФАДН2
2 ГТФ
70. Челночные системы
• Механизмы переноса водородов от НАДНиз цитозоля в митохондрии.
• Малат-аспартатный челночный механизм
(сердце, почки, печень)
• Глицеролфосфатный челночный механизм
(скелетные мышцы, мозг, печень)
71. Малат-аспартатная челночная система
Малат-аспартатная челночнаяЦитозоль
система Митохондрия
НАДН+Н+
Оксалоацетат
НАД+
Малат
М
ОА
НАД+
НАДН+Н+
Глу
Глу
ДЦ
α-кг
Асп
Асп
α-кг
3АТФ
72. Малат-аспартатная челночная система
73.
• Энергетический баланс аэробногогликолиза при использовании
малат-аспартатного челночного
механизма
38 АТФ
74. Глицеролфосфатная челночная система
Глицеролфосфатная челночнаяЦитозоль
система Митохондрия
НАДН+Н+
Дигидроксиацетонфосфат
ФАД
Глицерол3-фосфат ДГ
НАД+
Глицерол3-фосфат
ФАДН2
ДЦ
2АТФ
75.
• Энергетический баланс аэробногогликолиза при использовании
глицеролфосфатного челночного
механизма
36 АТФ
76. Глюконеогенез
• Синтез глюкозы из неуглеводных веществ(лактат, пируват, гликогенные
аминокислоты, глицерол).
• Происходит в печени и коре почек.
• Протекает интенсивно при длительном
голодании, при интенсивной мышечной
нагрузке.
77. Глюконеогенез
• Протекает за счет обратимыхреакций гликолиза, но 3
необратимые реакции гликолиза
заменяются на другие реакции –
обходные пути глюконегенза.
78. Схема гликолиза и глюконеогенеза
79.
80.
81. Обходные пути глюконегенза
82. Обходные пути глюконегенза
83. Обходные пути глюконегенза
84. Обходные пути глюконегенза
Фруктоза-1,6-дифосфат + Н2О →Фруктоза-6-фосфат + Н3РО4
Фермент – фруктоза-1,6-дифосфатаза
Глюкоза-6-фосфат + Н2О → Глюкоза
+ Н3РО4
Фермент – глюкоза-6-фосфатаза
85. Суммарная реакция глюконеогенеза
2 Пиуват + 4АТФ + 2ГТФ + 2НАДН+2Н+ ++ 4Н2О
Глюкоза + 4АДФ + 2ГДФ +
+ 6Н3РО4 + 2НАД+
86. Глюкозо-лактатный цикл (цикл Кори)
МышцыПечень
Кровь
Глюкоза
Глюкоза
Пируват
Пируват
Лактат
Лактат
87. Глюкозо-аланиновый цикл
ПеченьМышцы
Кровь
Глюкоза
Глюкоза
Пируват
Пируват
Аланин
Аланин
88. Причины лактоацидоза
• Гипоксии (активация анаэробного гликолиза)• Патология печени (нарушение глюконеогенеза)
• Наследсвенные дефекты ферментов
глюконеогенеза (пример – глюкозо-6фосфатаза)
• Нарушение ПДК (дефекты ферментов или
гиповитаминозы)
• Лекарства (бигуаниды ингибируют
глюконеогенез)
89.
90. Регуляция гликолиза и глюконеогенеза
• Осуществляется на уровне ферментов,катализирующих необратимые
реакции гликолиза и глюконеогенеза.
• Активаторы гликолиза являются
ингибиторами глюконеогенеза и
наоборот.
91. Регуляция гликолиза и глюконеогенеза
• Аллостерическая регуляция• Регуляция энергетическим статусом
клетки
• Ковалентная регуляция –
фосфорилирование/дефосфорилирование
• Индукция/репрессия ферментов
92. Регуляция гликолиза и глюконеогенеза
ГлюкозаГексокиназа
-
Глюкозо-6фосфатаза
Глюкозо-6фосфат
+
В печени глюкокиназа индуцируется инсулином
93. Регуляция гликолиза и глюконеогенеза
Фруктозо-6фосфатФруктозо-1,6Фосфофруктодифосфатаза
киназа
+
-
Фруктозо-1,6дифосфат
Фруктозо-2,6-дифосфат
АМФ
-
+
АТФ, НАДН
94. Регуляция гликолиза и глюконеогенеза
ФосфоенолРЕПкарбоксипируват
киназа
Пируваткиназа
-
+
АТФ, НАДН,
ацетил-КоА, ЖК
Оксалоацетат
Пируват
Пируваткарбоксилаза
-
+Ф-1,6-диР,
АМФ
95. Гормональная регуляция гликолиза и глюконеогенеза
• Инсулин (абсорбтивный период) приводит кдефосфорилированию регуляторных
ферментов гликолиза и глюконеогенеза –
гликолиз активируется, а глюконеогенез
ингибируется.
• Глюкагон (голодание) приводит к
фосфорилированию регуляторных
ферментов гликолиза и глюконеогенеза –
гликолиз ингибируется, а глюконеогенез
активируется.
96. Гормональная регуляция гликолиза и глюконеогенеза
• Кортизол индуцирует регуляторныеферменты глюконеогенеза:
• Фосфоенолпируваткарбоксикиназа
• Фруктозо-1,6-бифосфатаза
97.
98.
Спиртовое брожение99. Пентозофосфатный путь окисления глюкозы
Метаболическая роль:1. Образование пентоз (для синтеза
нуклеотидов, нуклеиновых кислот,
кофакторов).
2. Образование НАДФН:
А. Восстановительные синтезы – синтез
жирных кислот, холестерола, стероидных
гормонов, аминокислот;
В. Обезвреживание токсичных веществ.
100. Пентозофосфатный путь окисления глюкозы
Локализация:
Печень
Жировая ткань
Кора надпочечников
Эритроциты
Молочная железа в период лактации
Семенники
101. Пентозофосфатный путь окисления глюкозы
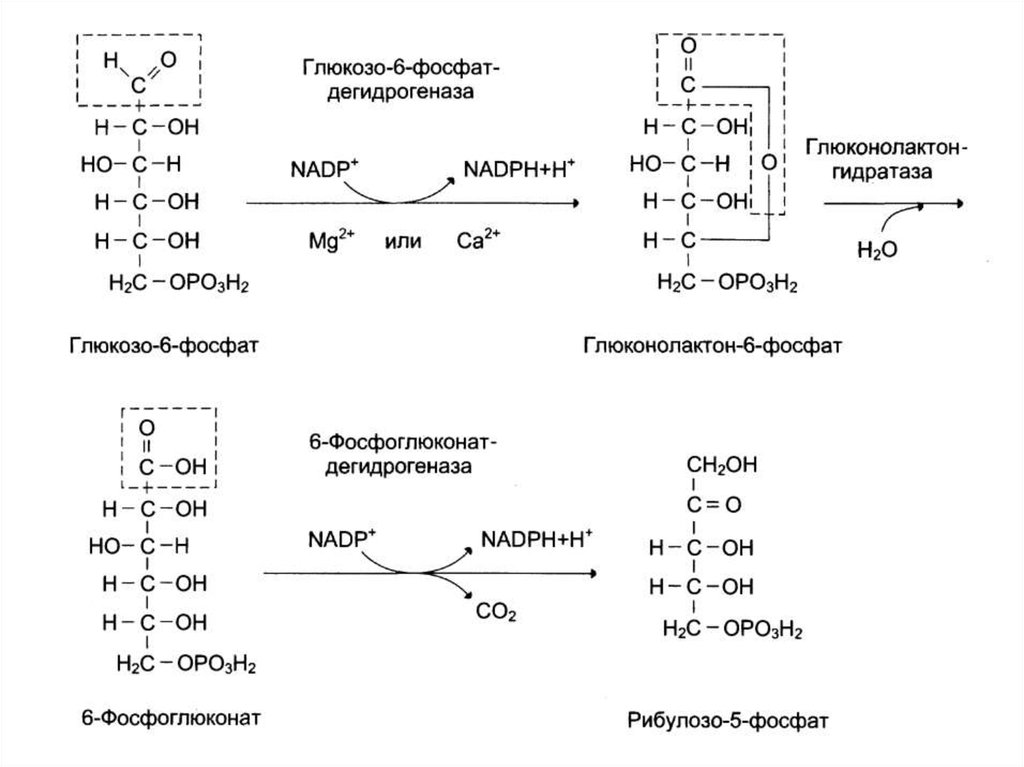
Этапы:1. Окислительный этап – превращение гексоз
в пентозы
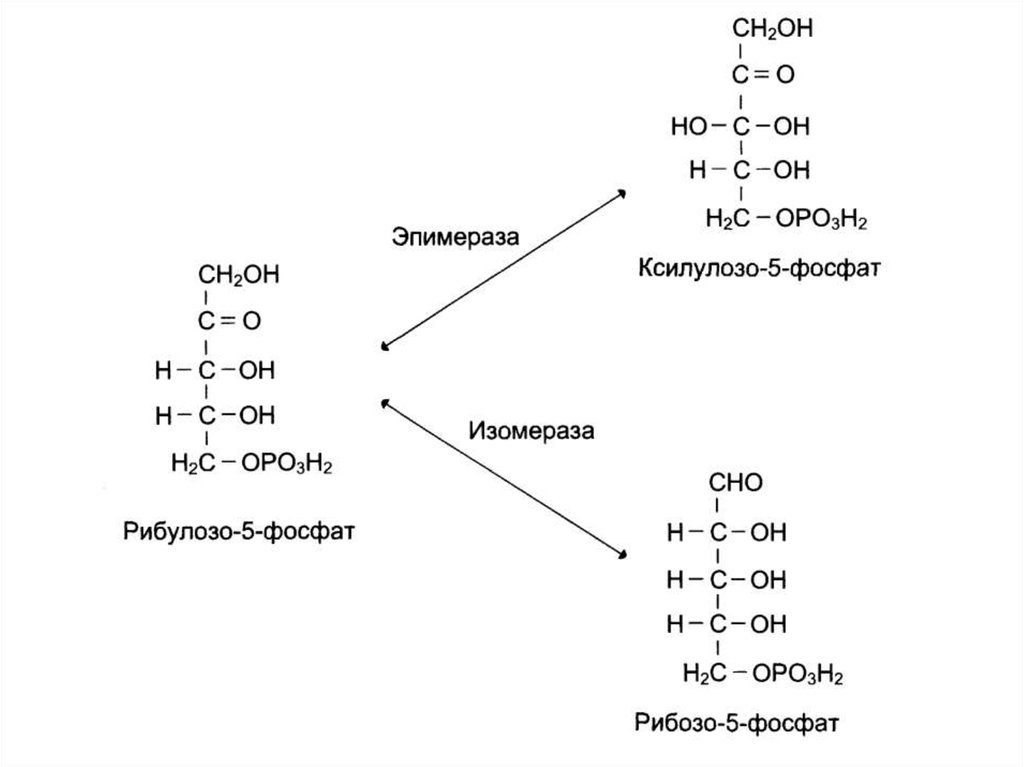
2. Неокислительный этап – превращение
пентоз в гексозы
102.
103.
104. Неокислительный этап пентозофосфатного пути окисления глюкозы
105. Неокислительный этап пентозофосфатного пути окисления глюкозы
106. Схема пентозофосфатного пути
107.
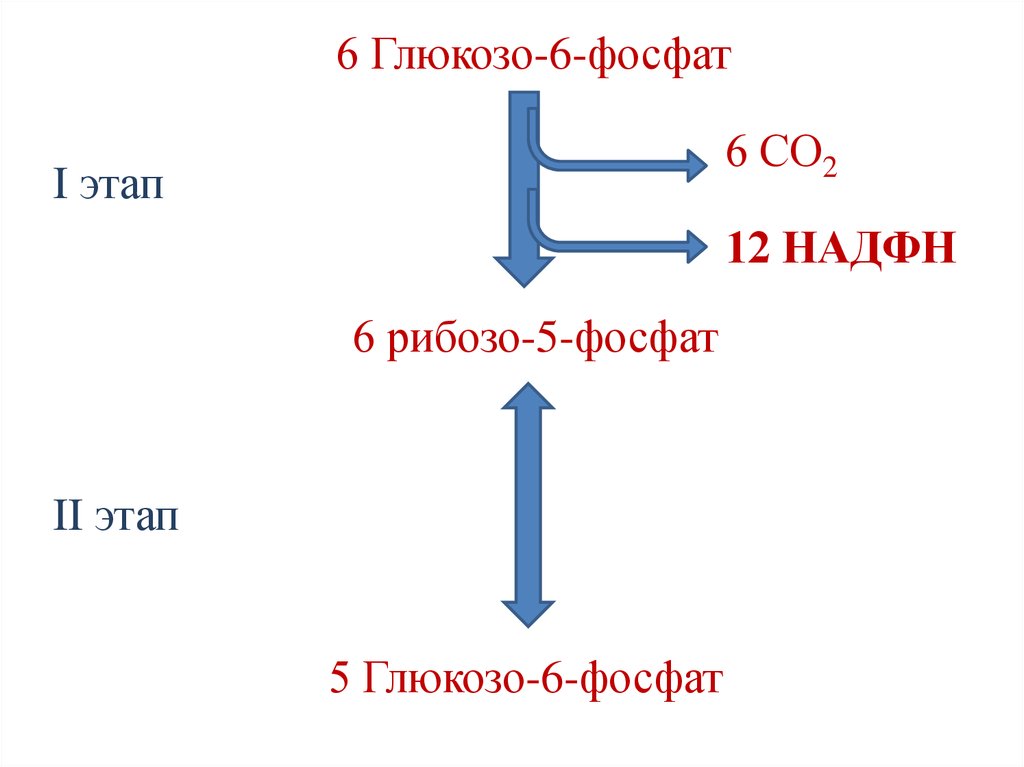
6 Глюкозо-6-фосфат6 СО2
I этап
12 НАДФН
6 рибозо-5-фосфат
II этап
5 Глюкозо-6-фосфат
108. Варианты протекания пентозофосфатного пути
1. Если потребности тканей в НАДФН ирибозе примерно одинаковы, протекает
только I этап пентозофосфатного пути.
Суммарная реакция:
6 глюкозо-6-фосфат + 12НАДФ+ + 6Н2О →
6 рибозо-5-фосфат + 12(НАДФН+Н+) + 6 СО2
109. Варианты протекания пентозофосфатного пути
2. Если потребности тканей в НАДФН выше,чем потребности в рибозе, протекают оба
этапа пентозофосфатного пути.
Суммарная реакция:
глюкозо-6-фосфат + 12НАДФ+ + 2Н2О →
12(НАДФН+Н+) + 6 СО2
110. Варианты протекания пентозофосфатного пути
3. Если потребности тканей в рибозе выше,чем в НАДФН, протекает только II этап в
обратном направлении.
Суммарная реакция:
5 фруктозо-6-фосфат → 6 рибозо-5-фосфат
111. Дефект глюкозо-6-фосфат дегидрогеназы в эритроцитах
• Уменьшается концентрация НАДФН вэритроцитах → окислительный стресс
• Происходит окисление –SH групп
гемоглобина с образованием перекрестных
дисульфидных связей и агрегация
протомеров гемоглобина → образуются
тельца Хайнца.
• Нарушается пластичность мембраны
эритроцитов → гемолиз.
112. Тельца Хайнца
113. Метаболизм фруктозы
• I путь – печень, почки, кишечник• II путь – мышцы
114. Наследственные нарушения метаболизма фруктозы
Ф-1-РФруктокиназаальдолаза
Фруктоза
Ф-1-Р
ДГАР + ГА
1. Недостаточность фруктокиназы –
эссенциальная фруктозурия →
выделение фруктозы с мочой
115. Наследственные нарушения метаболизма фруктозы
Ф-1-РФруктокиназа
альдолаза
Фруктоза
Ф-1-Р
ДГАР + ГА
2. Недостаточность фруктозо-1-фосфатальдолазы
непереносимость фруктозы →
рвота, диарея, боли в животе, кома, судороги,
нарушение функции печени и почек,
гипогликемия, гипофосфатемия, гиперурикемия
метаболический ацидоз (кетоновые тела).
116. Метаболизм галактозы
117. Наследственные нарушения метаболизма галактозы
Гал-1-Р-уридилГалактокиназа трансферазаГалактоза
Гал-1-Р
УДФ-гал
1
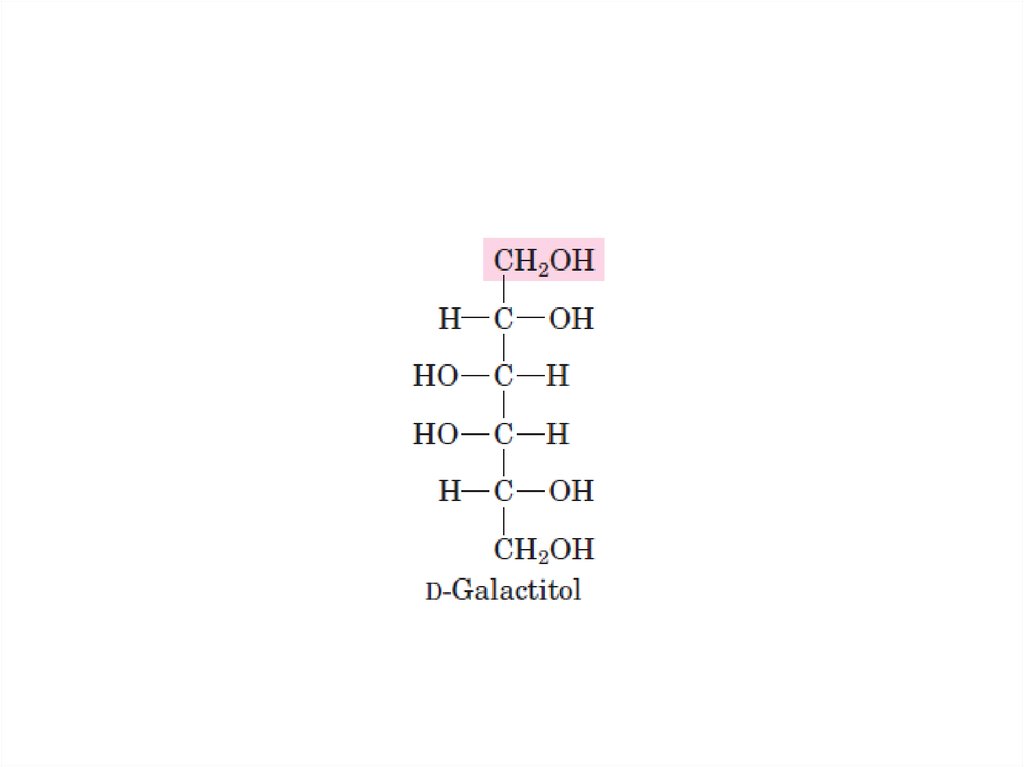
1. Недостаточность галактокиназы –
галактоземия →
галактоземия, галактозурия,
катаракта (восстановление галактозы
в галактитол).
118.
119. Наследственные нарушения метаболизма галактозы
Гал-1-Р-уридилГалактокиназа трансферазаГалактоза
Гал-1-Р
УДФ-гал
2
2. Недостаточность галактозо-1-фосфатуридил-трансферазы – галактоземия
(непереносимость галактозы) →
галактоземия, галактозурия, катаракта,
гипогликемия, умственная отсталость,
цирроз печени, гепатомегалия
120. Регуляция концентрации глюкозы в крови
• Нормальное содержание глюкозы в крови –3,3-5,5 ммоль/л
• После приема пищи (в течение 1 часа) –
повышается до 8 ммоль/л
• Спустя 2 часа – уровень глюкозы
возвращается к норме
121. Изменение концентрации глюкозы в течение суток
122. Влияние инсулина
Кровь↑ Глюкоза
Гликоген
Глюкоза
+
+
+
Глюкозо-6-Р
Инсулин
+
-
Пируват
Ткани
-
123. Влияние инсулина на обмен углеводов
• Стимулирует транспорт глюкозы из крови вткани (транслокация ГЛЮТ4 на мембрану).
• Индуцирует глюкокиназу.
• Активирует синтез гликогена (гликогенсинтаза).
• Ингибирует распад гликогена
(гликогенфосфорилаза).
• Активирует регуляторные ферменты гликолиза.
• Ингибирует регуляторные ферменты
глюконеогенеза.
124. Влияние глюкагона и адреналина
• Активируют распад гликогена(гликогенфосфорилаза).
• Ингибируют синтез гликогена
(гликогенсинтаза).
• Активируют регуляторные ферменты
глюконеогенеза.
• Ингибируют регуляторные ферменты гликолиза.
• Активируют глюкозо-6-фосфатазу, что приводит
к дефосфорилированию глюкозы и ее секреции
в кровь.
125. Влияние кортизола
• Индуцирует регуляторные ферментыглюконеогенеза.
• Активирует катаболизм мышечных
белков и триглицеридов жировой
ткани, обеспечивая глюконеогенез
субстратами (аминокислоты,
глицерол).
126. Регуляция уровня глюкозы в крови в абсорбтивном постабсорбтивном периоде
Выделяется инсулин.Около 60% глюкозы поступает в печень
(воротная вена).
• 2/3 превращается в гликоген.
• 1/3 окисляется для получения энергии и
превращается в жиры.
127. Регуляция уровня глюкозы в крови в абсорбтивном постабсорбтивном периоде
Около 40% поступает в общий кровоток.• 2/3 поглощается мышцами (превращается в
гликоген) и жировой тканью (превращается в
жиры).
• 1/3 поступает в другие ткани (окисление).
• При нормальном ритме питания уровень
глюкозы в крови поддерживается в основном за
счет синтеза и распада гликогена.
128. Регуляция уровня глюкозы в крови при длительном голодании
Выделяется глюкагон.• Активируется распад гликогена (в течение
суток запасы гликогена исчерпываются).
• Усиливается глюконеогенез (из лактата,
аминокислот, глицерола).
• Выделяется кортизол (индуцирует регуляторные
ферменты глюконеогенеза).
129. Регуляция уровня глюкозы в крови при длительном голодании
• При голодании глюкоза не используетсямышцами и жировой тканью (из-за низкого
содержания инсулина).
• Глюкоза сберегается для головного мозга и
других глюкозозависимых клеток.
130. Регуляция уровня глюкозы в крови при мышечной нагрузке
• Вначале источником глюкозы для мышцявляется гликоген, запасенный в мышцах (при
интенсивной мышечной нагрузке 100 г
гликогена используется в течение 15 минут).
• После исчерпания запасов гликогена мышц,
глюкоза поступает в мышцы из крови (распад
гликогена и глюконеогенез в печени).
131. Сахарный диабет
Причина – абсолютная илиотносительная недостаточность
инсулина.
Основные типы:
1. Инсулинзависимый (I тип)
2. Инсулиннезависимый (II тип)
132. Сахарный диабет I типа
Причина – абсолютный дефицитинсулина:
• Аутоиммунная деструкция -клеток
(пусковые механизмы – вирусные
инфекции: оспа, краснуха, корь,
цитомегаловирус, паротит, аденовирус).
• Генетическая предрасположенность.
Начало – молодой возраст.
133. Сахарный диабет II типа
Причина – относительный дефицитинсулина:
• Инсулинорезистентность (дефект
рецептора инсулина, пострецепторного
аппарата).
• Нарушение секреции инсулина.
• Пусковые механизмы – ожирение,
малоподвижный образ жизни, стресс.
Начало – после 40 лет.
134. Проявления сахарного диабета
• Гипергликемия, глюкозурия• Кетонемия, кетонурия
• Повышение концентрации жирных
кислот
• Повышение концентрации
аминокислот
• Ацидоз
• Уремия
135. Клинические проявления сахарного диабета
• Полиурия (из-за повышенияконцентрации глюкозы, кетоновых
тел, мочевины).
• Полидипсия (из-за полиурии).
• Полифагия (энергетический голод
клеток).
• I тип – больные худеют, II тип –
ожирение.
136. Стероидный диабет
Причина – гиперсекреция кортизола.• Стимуляция глюконеогенеза.
• Стимуляция катаболизма мышечных
белков и триглицеридов жировой ткани
(аминокислоты, глицерол используются
для синтеза глюкозы).
137. Тесты для оценки углеводного обмена
Пероральный тест толерантности кглюкозе:
• Определяют концентрацию глюкозы в
крови натощак (12-14 часов после
приема пищи).
• Нагрузка глюкозой (50 г глюкозы в 250
мл воды).
• Определяют концентрацию глюкозы
через 2 часа.
138.
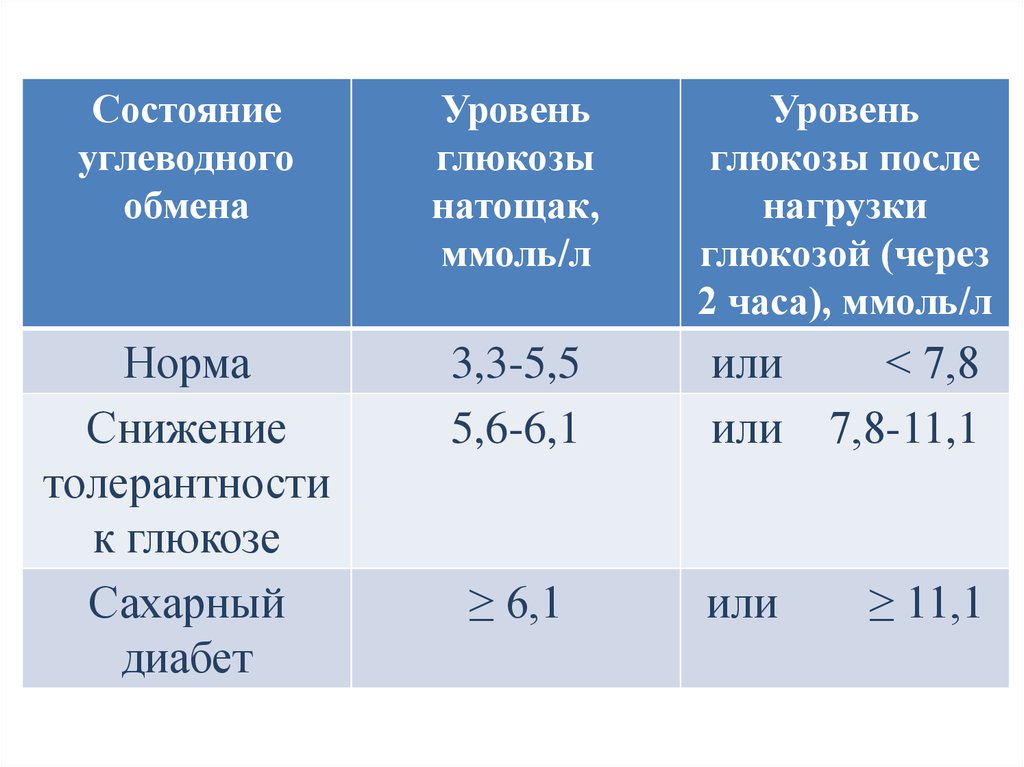
Состояниеуглеводного
обмена
Уровень
глюкозы
натощак,
ммоль/л
Уровень
глюкозы после
нагрузки
глюкозой (через
2 часа), ммоль/л
Норма
Снижение
толерантности
к глюкозе
Сахарный
диабет
3,3-5,5
5,6-6,1
или
< 7,8
или 7,8-11,1
≥ 6,1
или
≥ 11,1
139. Концентрация инсулина и С пептида
• Инсулин и С пептид секретируютсяподжелудочной железой в эквимолярных
концентрациях.
• В норме отношение инсулина к С пептиду в
крови составляет 1/3 (т.к. 2/3 инсулина
задерживается печенью).
• Определение пептида С используются для
оценки секреторной функции
поджелудочной железы.
140. Гликозилированный гемоглобин (Hb A1c)
• В норме составляет 4-6% от общегосодержания гемоглобина.
• Является индикатором уровня глюкозы
крови.
• При сахарном диабете увеличивается в 2-3
раза.