































 Биология
БиологияПохожие презентации:








")
")
Естественный отбор, его формы и влияние на генофонд популяций
1.
Естественный отбор, егоформы и влияние на
генофонд популяций
2.
Естественный отборОтбор – это дифференциальное (неодинаковое) и
неслучайное (зависимое от приспособленности)
воспроизводство генотипов.
То есть в основе механизма отбора лежит неодинаковая
приспособленность, или неравная средняя
эффективность воспроизводства особей, имеющих
различия по некоторым генетически обусловленным
признакам фенотипа.
3.
Измерение приспособленностиАбсолютная приспособленность (F, fitness) – это величина,
являющаяся функцией от двух ключевых переменных:
выживаемость (В) и плодовитость (П)
F [потомков/поколение] = В × П
Выживаемость – процент оплодотворенных яиц, семян или
новорожденных, доживающих до стадии размножения (в среднем);
Плодовитость – число потомков, производимых (в среднем) одной
половозрелой особью.
Можно рассчитать F, не зная В и П, по формуле:
F = N1/N0
где N0 и N1 – число половозрелых носителей данного генотипа и
фенотипа в двух последовательных поколениях.
4.
Относительная приспособленность икоэффициент отбора
Относительная приспособленность выражается коэффициентом
приспособленности W.
W = F(1) /F(2)
Где F(1) и F(2) – коэффициенты абсолютной приспособленности
сравниваемых генотипов. В знаменатель обычно ставится значение
F для наиболее приспособленного генотипа, для которого W = 1.
Сила отсеивающего отбора измеряется коэффициентом отбора S.
S =1 – W
Чем выше S, тем быстрее будет происходить элиминация данного
генотипа.
Для наиболее приспособленного генотипа S = 0;
Для летальной мутации S = 1 (элиминация в первом поколении).
5.
Моделирование естественного отбораКак рассчитать изменение частот генотипов во времени?
Если частота встречаемости генотипа 1 в нулевом поколении
обозначена как p0, а альтернативного генотипа 2 как q0 (при p+q=1),
то частота встречаемости генотипа 1 в поколении n (pn)
вычисляется по формуле:
(pn) = (p0 × Wn)/(q0 + p0 × Wn)
Изменение частоты по данной формуле рассчитывают для менее
приспособленного генотипа (W < 1). При этом частоту более
приспособленного генотипа в поколении n можно рассчитать по
формуле:
qn = 1 – pn
6.
ЗадачаПусть в большой популяции бактерий появилась новая мутация гена А
(вариант А1). Ее частота (q) в популяции составляет 0,005
Новый вариант гена обеспечивает повышение скороcти размножения так,
что относительная приспособленность исходного генотипа оказывается =
= 0,9 по отношению к новому генотипу. Какова будет частота исходного
варианта гена (p) в популяции в 20-м, 100-м, 1000-м поколениях?
(p20) = (0,995 × 0,920)/(0,005 + 0,995 × 0,920) = 0,96
(p50) = (0,995 × 0,950)/(0,005 + 0,995 × 0,950) = 0,51
(p200) = (0,995 × 0,9200)/(0,005 + 0,995 × 0,9200) = 0,00000014
Если частота аллеля принимает значение p = 0 - это элиминация аллеля.
Если частота аллеля растет и достигает значения p = 1 – это фиксация
аллеля.
Если один из аллелей фиксируется вместо другого (предкового) аллеля, то
говорят, что произошло аллельное замещение.
7.
Предел демографического насыщенияэкологической ниши
Отбор, то есть изменение частот фенотипов и генотипов (аллелей)
в зависимости от различий в эффективности размножения идет
постоянно.
В то же время фиксация одних аллелей и элиминация других
(аллельное замещение) более вероятно в условиях достижения
предела демографического насыщения экологической ниши, т.е.
такого размера популяции, дальнейший рост которого невозможен
ввиду ограниченности количества ресурсов в занимаемом ареале.
То есть в популяции с предельным размером в 1000 особей
полной элиминации вредного генотипа можно ожидать с высокой
вероятностью после снижения его частоты до 1/1000.
8.
ЧастотаГрафическое отображение теоретической
модели изменения частот p и q
1
0,9
0,8
0,7
0,6
0,5
0,4
0,3
0,2
0,1
0
p
0
20
40
60
80
Поколение
Сферическая популяция в вакууме
q
100
9.
Эти расчеты часто не соответствуют идеально данным,получаемым на практике в полевых наблюдениях или в
экспериментах. Причины:
• Вклад случайных отклонений частот (дрейфа генов).
• Плейотропное действие гена: новая мутация может оказывать полезный
эффект в отношении одной функции, но вредный эффект в отношении какойто другой функции.
• Неполная пенетрантность и вариативная экспрессивность гена –
неустойчивое фенотипическое выражение одного и того же варианта гена у
разных особей, зависимое от средовых или случайных факторов.
• Эффекты соотбора – влияние сцепления с другими генами, подверженными
отбору в большей степени, чем анализируемый ген. Сцепление с вредным
геном будет приводить к совместной элиминации, а сцепление с полезным
геном – к совместной фиксации.
• Эпистаз: неодинаковое проявление одного и того же варианта гена и его
адаптивной ценности в разном генетическом контексте (комбинации всех
остальных генов в генотипе конкретного организма). Генетический контекст
при половом размножении высоко индивидуален и определяется конкретной
комбинацией генов родительских особей в конкретной паре.
• Непостоянство условий среды, ведущее к переменчивости адаптивной
ценности каждого варианта генотипа в разных поколениях.
• Повторное появление мутаций или их приток из соседних популяций.
10.
Значение плотности популяции и еерегуляции в процессе эволюции
• До момента достижения демографического предела
экологической ниши эволюционный шанс сохраняется за всеми
генотипами, в принципе способными (хуже или лучше) к
воспроизведению.
• Следовательно, в условиях сниженной плотности популяции
конкурентный отбор ослабляется, в условиях повышенной
плотности – усиливается.
• Факторы, препятствующие достижению критических значений
плотности (паразиты, хищники, пожары и пр.) могут работать
как факторы, ослабляющие конкуренцию и способствующие
сохранению в популяции разнообразия генотипов, хотя они же
одновременно являются и факторами отбора на сопротивляемость. Тем самым, они могут благоприятствовать поддержанию
эволюционного потенциала популяции.
11.
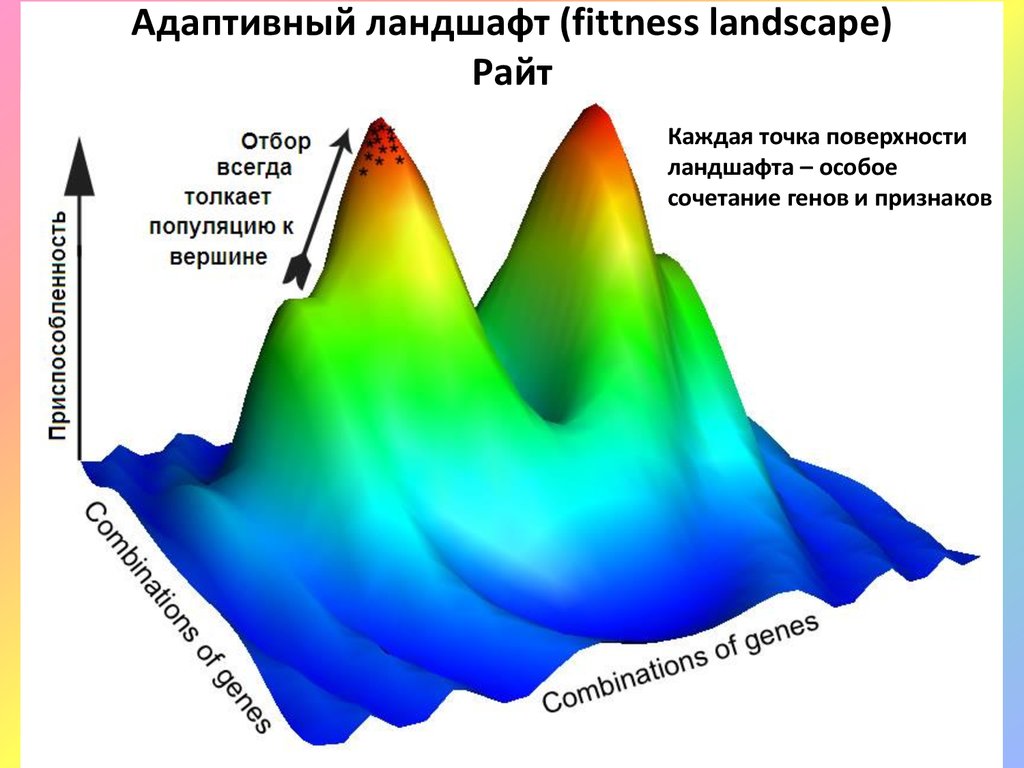
Адаптивный ландшафт (fittness landscape)Райт
Каждая точка поверхности
ландшафта – особое
сочетание генов и признаков
12.
Условие эффективности отбора (Райт): отбор будетпредсказуемо влиять на частоты генотипов, только в
том случае если Ne>>(1/S).
Ne – эффективная численность популяции
(численность особей, участвующих в репродукции).
При Ne≤(1/S) частоты генотипов могут меняться стохастично
(дрейф генов), так что различные генотипы будут либо
сосуществовать, либо может произойти случайная утрата
(элиминация) или фиксация любого, в том числе и менее
адаптивного («вредного») варианта гена.
Выводы:
1) чем выше эффективная численность популяции, тем
эффективнее отбор;
2) чем малочисленнее популяция и чем нейтральнее признак,
тем выше роль дрейфа.
Какой размер должна иметь популяция для эффективного
отбора аллели гена с S = 0,1?
13.
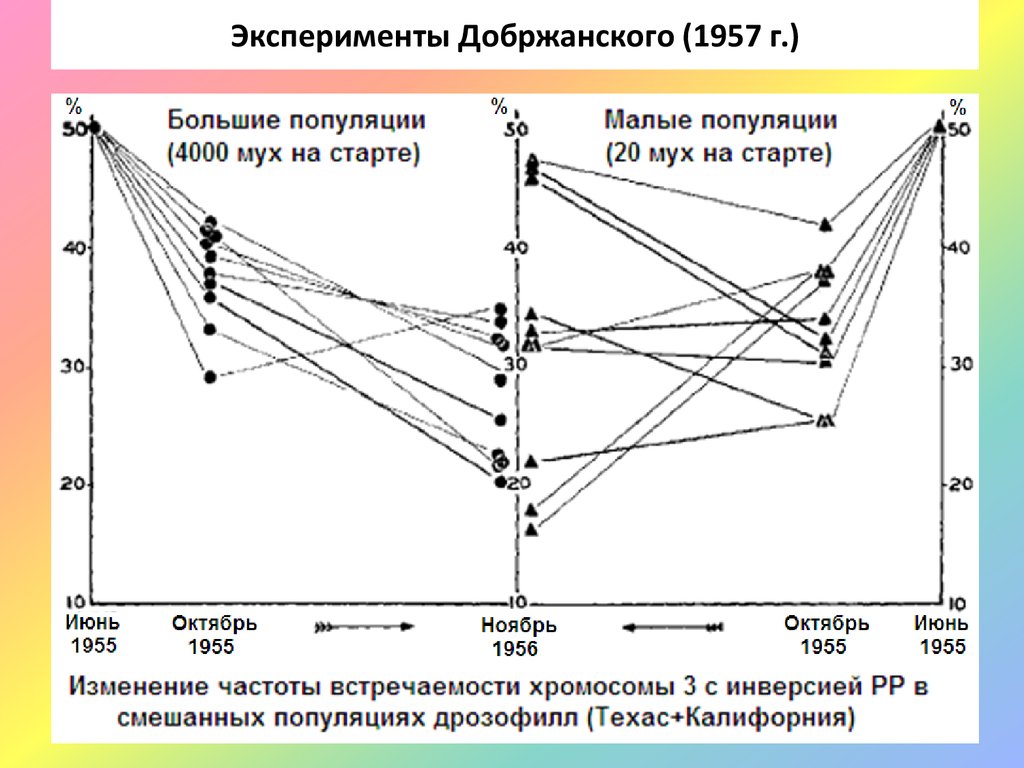
Эксперименты Добржанского (1957 г.)14.
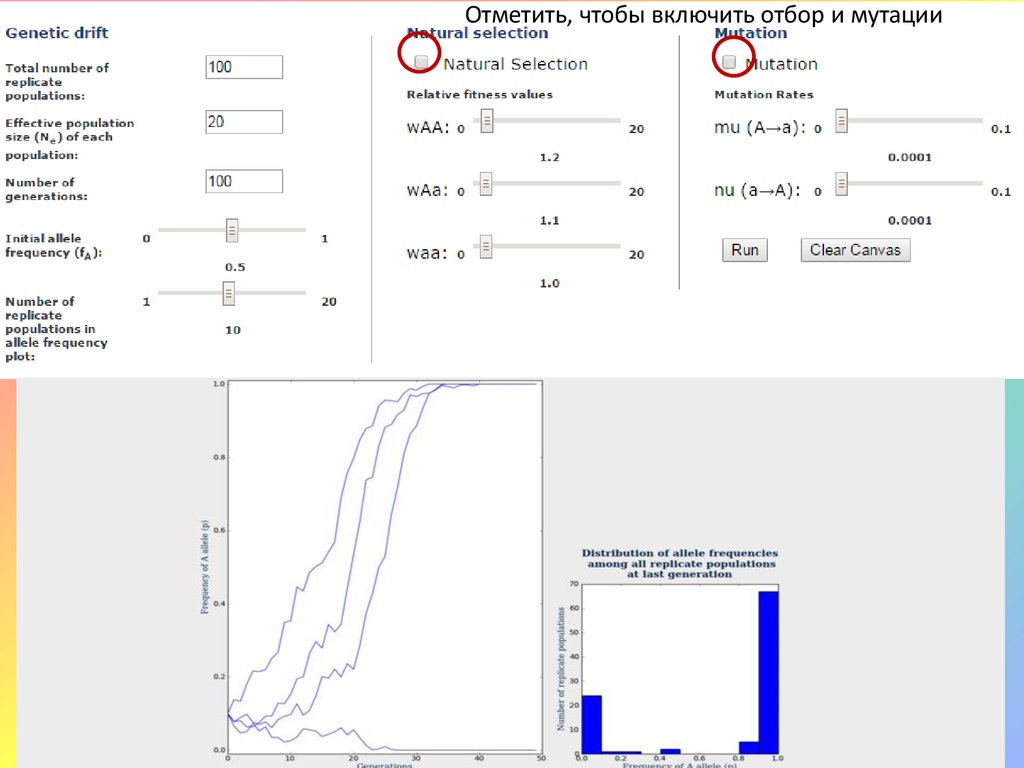
Виртуальная симуляция: Мутации + дрейф + отборhttp://trilby.uis.georgetown.edu:8000/simulations/driftselection/
Посмотреть динамику при параметрах:
1) Кол-во популяций – 100, размер популяций – 2000, количество генераций –
100, частота аллеля А – 0,2. Отбора и мутаций нет. Насколько выполняется
закон Харди-Вайнберга?
2) Кол-во популяций – 100, размер популяций – 20, количество генераций – 100.
Отбора и мутаций нет. Насколько выполняется закон Харди-Вайнберга?
Почему различается доля популяций, в которых произошла элиминация и
фиксация аллелей?
3) Те же начальные условия (2 варианта). Подключаем отбор, с коэффициентом
относительной приспособленности генотипов АА и Аа = 1,2. Как отличается
динамика от модели без отбора? Как различаются варианты между собой?
4) Те же начальные условия (2 варианта). Подключаем отбор, с коэффициентом
относительной приспособленности генотипов аа и АА = 1, генотипа Аа = 2. Как
распределяются частоты аллелей в популяциях?
5) К каждому из вариантов подключаем мутации с одинаковыми или разными
вероятностями. Начальную частоту аллеля А ставим равной 0, число
генераций - 1000. Каков результат, как он отличается от ситуации без
мутаций?
15.
Отметить, чтобы включить отбор и мутации16.
• Дрейф особенно влиятелен для популяций или генотипов с малойчисленностью.
Малочисленные популяции могут возникать при следующих
обстоятельствах:
- Разрыв ареала вследствие геологических процессов (вулканы,
появление новых рек, горных цепей, ледников и т. д.).
- Отселение небольшой части популяции на изолированные ареалы,
прежде не заселенные данным видом в процессе миграции
(заселение островов, отдаленных лесных массивов и т. д.). Местная
популяция будет формироваться на основе того небольшого набора
аллелей, который имеется у этой исходно небольшой группы особей –
«эффект основателя» (Райт).
- Период спада в популяционных волнах (периодические колебания
численности популяции, обусловленные различными экологическими
факторами и их сочетаниями, в том числе сокращение количества
ресурсов, сокращение ареала, пригодного для существования данного
вида ). Эффект резкого сокращения генетического разнообразия в
условиях резкого спада численности популяции назван «эффектом
бутылочного горлышка» (Майр).
17.
Каждая точка поверхностиландшафта – особое
сочетание генов и
признаков
Удержание популяции в зоне вершины адаптивного пика зависит от наложения
интенсивностей 3-х основных процессов:
1) генетической изменчивости (мутации + рекомбинации + поток генов),
2) отбора и 3) дрейфа генов.
18.
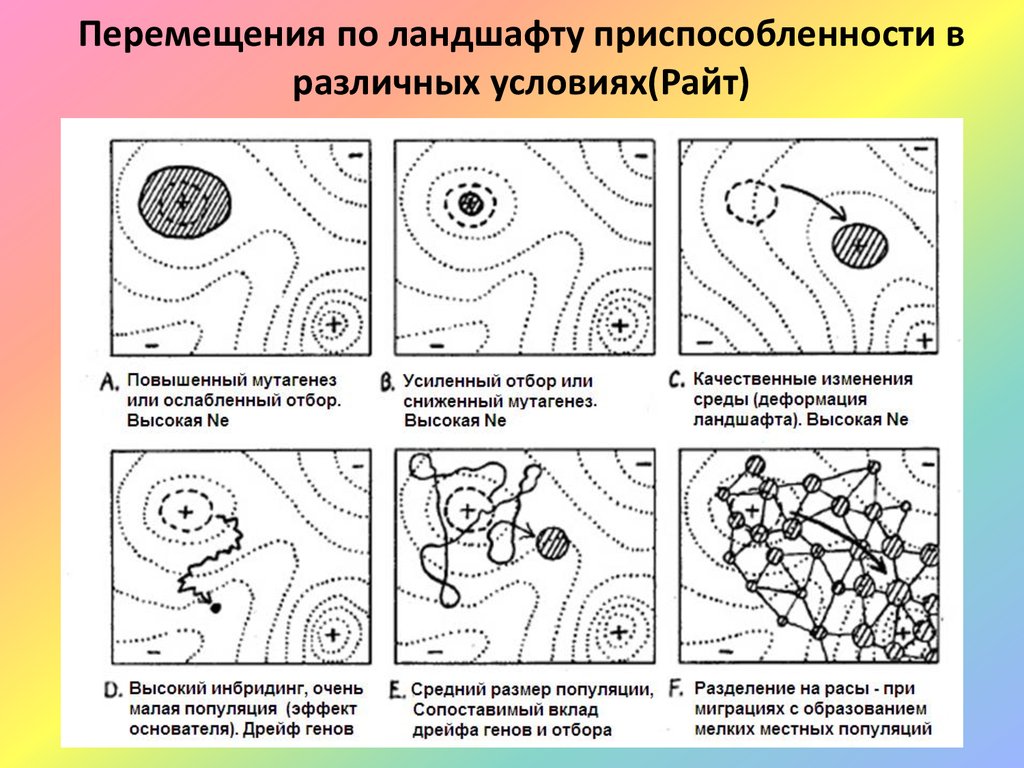
Перемещения по ландшафту приспособленности вразличных условиях(Райт)
19.
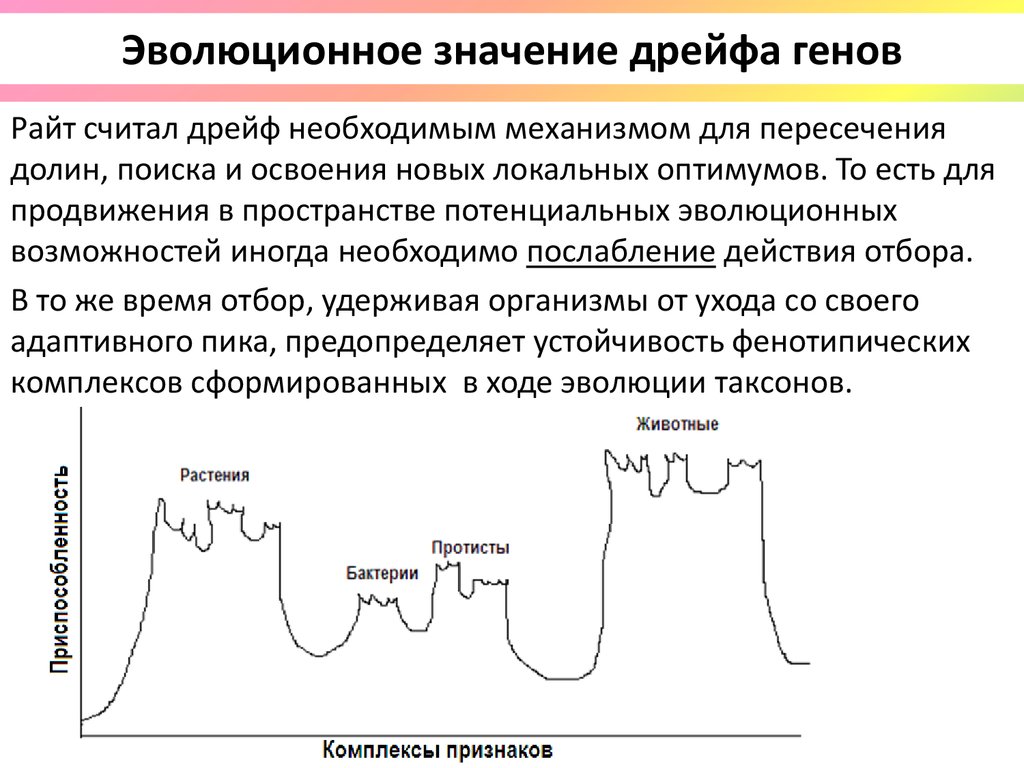
Эволюционное значение дрейфа геновРайт считал дрейф необходимым механизмом для пересечения
долин, поиска и освоения новых локальных оптимумов. То есть для
продвижения в пространстве потенциальных эволюционных
возможностей иногда необходимо послабление действия отбора.
В то же время отбор, удерживая организмы от ухода со своего
адаптивного пика, предопределяет устойчивость фенотипических
комплексов сформированных в ходе эволюции таксонов.
20.
Динамика адаптивного ландшафтаНеобходимо иметь в виду, что адаптивный ландшафт не
остается постоянным. Форма этого ландшафта изменяется не
только в ответ на изменения чисто внешней среды, но и на
собственные
изменения
в
генетике
организмов,
фиксирующиеся в процессе эволюции. Поэтому какие-то
переходы, невозможные в один период времени, становятся
возможными в последующем, равно как и наоборот, прежде
возможные переходы могут в последующем оказаться
невозможными.
Почему нынешние обезьяны не превращаются в
цивилизованные виды, как это было с предками человека?
21.
Правило выживания «наиболее плоских» (surviving of the flattest):при наличии в популяции конкурирующих субпопуляций,
вынужденных адаптироваться к часто и непредсказуемо
меняющейся среде, в эволюционной перспективе выживут скорее
не те, которые достигают наибольших скоростей размножения
(адаптивный пик более «высокий»), а те, которые сохраняют
жизнеспособность при более широком разнообразии мутаций и
диапазоне условий.
Дарвин: «Выживает не самый сильный и
не самый умный, а тот, кто лучше всех
приспосабливается к изменениям.»
22.
Формы и направления отбораВ отношении вновь возникающих мутаций отбор может работать:
1) На их элиминацию – такой отбор называют отрицательным,
отсеивающим, очищающим. Мутации, подвергающиеся
отрицательному отбору называются вредными.
2) На их распространение и фиксацию – такой отбор называют
положительным,
поддерживающим.
Сами
мутации
рассматривают как полезные.
Отсеивающий отбор преобладает на протяжении эволюции. Отбор
сказывается на геномных последовательностях тем больше, чем они
более функционально значимы.
Следствие: наиболее значимые области генома как правило
наиболее консервативны.
Высококонсервативные (древние) области составляют около
10-15 % генома человека. Малозначимые геномные области
эволюционируют в нейтральном режиме и быстро изменяются.
23.
Наглядные примеры положительного отбора• Распространение мутаций устойчивости к ядам и антибиотикам у
синантропных видов и бактерий, опухолевых клеток.
• Распространение мутации устойчивости к прионной болезни куру
у аборигенов Папуа-Новой Гвинеи (заражались при обряде
каннибалического поедания мозга соплеменников).
К семинару еще примеры
24.
Балансирующий отборБалансирующий отбор – это такой отбор, результатом которого
оказывается не аллельное замещение, а, наоборот, сохранение и
повышение разнообразия генов и генотипов в популяции в ходе
микроэволюционного процесса.
3 типа случаев балансирующего отбора:
1) Отбор в разнородной среде или ситуация адаптивного
конфликта – освоение нескольких альтернативных стратегий
достижения репродуктивного успеха.
2) Частотно-зависимый отбор.
3) Преимущество гетерозигот.
25.
Отбор в разнородной среде или в условияхадаптивного конфликта
Организмы одной популяции сталкиваются с множеством
требований среды, которые могут противоречить друг другу,
создавая ситуацию адаптивного конфликта.
Балансирующий отбор поддерживает в популяции разнообразие
аллелей генов, которые дают возможность использовать несколько
альтернативных стратегий выживания, обеспечивая поддержание
устойчивости всей популяции в условиях действия нескольких
агрессивных факторов, влияние которых может быть
неравномерным в пространстве и во времени.
Примеры: 1) балансирование между мышечной силой и ловкостью;
2) светлые и темные морфы у насекомых, грызунов.
3) гены гистосовместимости: разные аллели могут определять
более выраженный иммунитет по отношению к разным инфекциям.
К семинару еще примеры.
26.
Частотно-зависимый отборЧастотно-зависимый отбор – ситуация, когда полезность гена
находится в зависимости (как правило, обратной) от его частоты в
популяции.
Примеры: 1) бейтсовская мимикрия (маскировка особей
неядовитых видов под ядовитых) – такая маскировка сработает,
только если обманщики редки по сравнению с действительно
ядовитыми.
2) Объяснение соотношения полов 1:1: когда самок больше,
выгоднее быть самцом, а когда больше самцов – выгоднее быть
самкой. В результате сохраняются и Х и У хромосомы, в
соответствующем
соотношении.
К семинару другие примеры.
27.
Мюллеровская мимикрияHeliconius
28.
Преимущество гетерозиготЕсли гетерозиготы имеют преимущество в воспроизводстве по
сравнению с каждым вариантом гомозигот, то каждый из
аллельных вариантов генов будет поддерживаться в популяции.
Пример 1: Присутствие нормального и мутантного аллелей
гемоглобина в человеческих популяциях регионов, где
распространен возбудитель малярии.
Гомозигота АА – нормальные эритроциты, восприимчивость к
малярии.
Гомозигота аа – летальный генотип (эритроциты не переносят
кислород).
Аа
–
серповидно-клеточная
анемия:
деформированные
эритроциты, сниженная кислородная емкость крови, устойчивость к
малярии.
29.
Преимущество гетерозиготПример 2:
2 варианта хромосомы 1 (1A и 1B) у гребенчатого тритона.
До рождения развиваются только генотипы 1A1B;
1A1A и 1B1B гибнут на стадии эмбрионального развития.
Каждая из хромосом содержит делеции разных генов,
обязательных для протекания нормального развития зародыша.
Результат: оба варианта хромосомы сохраняются в популяции.
30.
Отбор в эволюции количественных признаковНорма реакции – спектр значений, которые может принимать
количественный признак при заданном генотипе (с учетом
возможных различий в условиях протекания онтогенеза).
Каждый вид (или популяция) организмов характеризуется
собственной нормой реакции количественных признаков.
Типы отбора (по влиянию на норму реакции):
• Стабилизирующий
• Движущий
• Дизруптивный
• Дестабилизирующий
• Балансирующий (в т. ч. частотно зависимый)
Единицы отбора – особи (иногда группы особей).
Следствие отбора – сохранение или изменение нормы реакции и
генофонда популяции, определяющего эту норму реакции.
31.
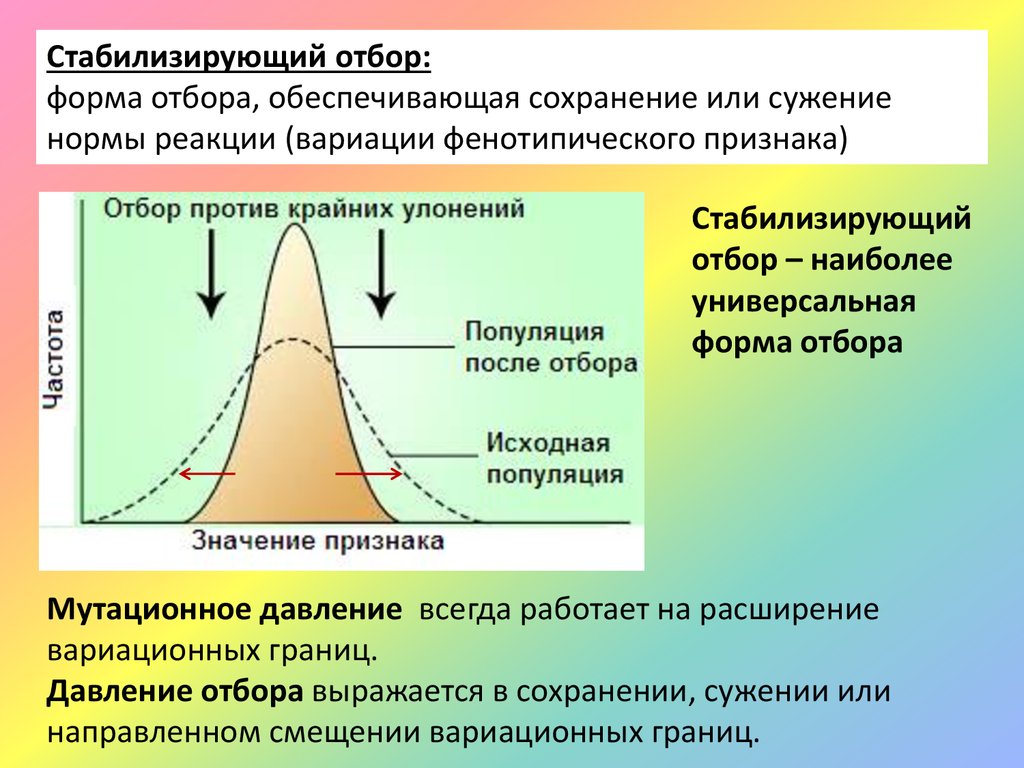
Стабилизирующий отбор:форма отбора, обеспечивающая сохранение или сужение
нормы реакции (вариации фенотипического признака)
Стабилизирующий
отбор – наиболее
универсальная
форма отбора
Мутационное давление всегда работает на расширение
вариационных границ.
Давление отбора выражается в сохранении, сужении или
направленном смещении вариационных границ.
32.
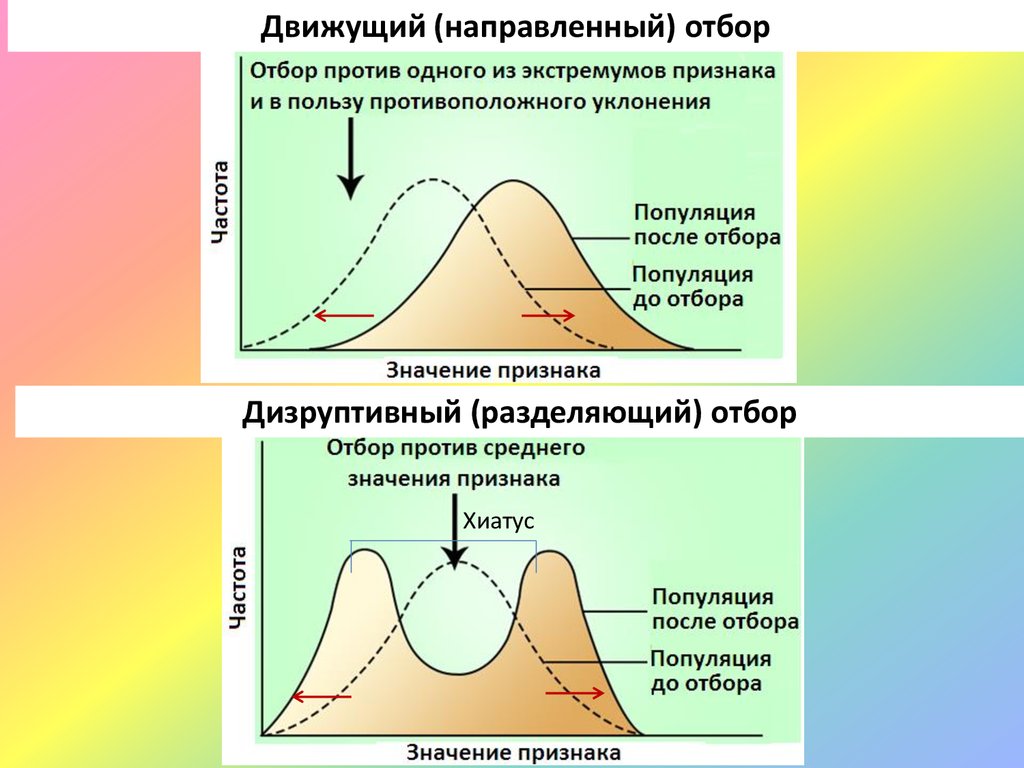
Движущий (направленный) отборДизруптивный (разделяющий) отбор
Хиатус
33.
Следствия действия стабилизирующегоотбора
• Отсев новых вредных мутаций (очищающий отбор)
• Закрепление новых компенсаторных мутаций по отношению к
ранее приобретенным вредным мутациям
• Закрепление новых генов, повышающих помехоустойчивость
генных сетей, обеспечивающих реализацию программы
онтогенеза.
• Закрепление определенных комбинаций генов, благоприятствуя
выживанию особей с наиболее удачными группами сцепления.
Стабилизирующий отбор может приводить к обеднению генофонда
в одних условиях (высокая стабильность и предсказуемость среды),
и к обогащению генофонда в других условиях (высокая
непредсказуемость среды). Во втором случае стабилизирующий
отбор обеспечивает повышение адаптационной и эволюционной
пластичности популяций.
34.
Дестабилизирующий отбор• Если условия среды меняются достаточно резко, чтобы создать
ситуацию включения сильного движущего (или дизруптивного)
отбора в отношении каких-то признаков организма, - внезапно
получают поддержку те мутации, которые снижают устойчивость
реализации онтогенеза, взламывающие систему
помехоустойчивости. В результате разброс значений признака
начинает расти – это и есть дестабилизирующий отбор.
• Дестабилизирующий отбор не может действовать долго, рано
или поздно он снова сменяется стабилизирующим,
фиксирующим новую норму реакции - это явление
перемежающегося дестабилизирующего (движущего) и
стабилизирующего отбора обозначено термином
«эволюционные качели».
35.
Эксперименты Беляева (1960-е)Лис отбирали на снижение
агрессивности (эксперимент
продолжается уже 50 лет)
Объяснение: онтогенетическое
развитие фенотипа регулируется теми
же гормонами, которые управляют и
развитием видовых особенностей
организации мозга и поведения. Отбор
привел к концентрированию мутаций,
вызывающих отклонение в системе
нейро-гуморальной регуляции
развития лис, что и дестабилизировало
видовые морфологические признаки.
36.
Еще 2 специфические формы отбора• Половой отбор – выдвигается в качестве предполагаемого
механизма эволюции вторичных половых признаков
самцов и самок, демонстративных форм поведения и
других признаков, определяющих не успех в выживании, а
преуспевание в размножении.
• Межгрупповой отбор – гипотеза, предлагаемая для
объяснения эволюции признаков кооперации и
альтруизма. Группа может быть как одновидовой (семья,
стая и т.п.), так и включать организмы нескольких видов,
формирующих симбиотический комплекс (напр., человек
и его микробиом). Чем теснее взаимозависимость
организмов и чем жестче среда, тем эффективнее такой
отбор.
37.
Эволюционная функция отбора• Отбор в биологических системах основан на
преимущественном размножении наиболее
приспособленных, т.е. несущих наиболее
оптимизированную, адекватную информацию о способе
выживания в занимаемой эконише среди конкурентов.
Соответственно, он обеспечивает:
1) Сброс «вредной», отягчающей информации
(=генетического груза).
2) Распространение (вертикальное и/или горизонтальное)
«полезной» информации, которая далее будет сохраняться
механизмом наследственности.
3) Аккумулирование «полезной» информации в генотипах,
приводящее к появлению принципиально новых фенотипов
на основе комбинаторики (творческая роль отбора).
38.
Изменчивость?
Фенотип <-> Генотип
?
Формирование качественно
новых генов (генотипов) и
признаков (фенотипов)
ОТБОР
Наследственность
(в соответствии с
законами Менделя)
Дрейф генов
Поток генов
Формирование частотной представленности генов и генотипов в генофонде
популяции.
Полноценное исследование закономерностей, управляющих
процессами изменчивости стало возможным только после
развития методов молекулярной биологии, благодаря
которым эволюционная биология претерпевает в последние
десятилетия революционные преобразования.
39.
Материал для дополнительного чтенияhttp://elementy.ru/news/431149
http://elementy.ru/news/431708
http://elementy.ru/news/431741
http://elementy.ru/novosti_nauki/432833
http://elementy.ru/novosti_nauki/431619
http://elementy.ru/novosti_nauki/430258
http://elementy.ru/novosti_nauki/431876
http://elementy.ru/novosti_nauki/432701
http://elementy.ru/news/430526