



























 Биология
БиологияПохожие презентации:





")




Структура и организация генома, репликация ДНК
1.
Репликация ДНК: образование копий ДНК,передаваемых от клетки к клетке, от
родителя потомку
Структура и организация генома
Молекулярная модель ДНК
2. Доказательство генетической роли ДНК
• Наследственный материал должен удовлетворятьследующим требованиям:
– 1. Информативность: должен содержать информацию
о строении и функции всего организма
– 2. Передача: должен передаваться от родителей
потомству
– 3. Репликация: должен копироваться
• Для передачи потомству
– 4. Изменчивость: должен иметь способность
изменяться
• В соответствие с известным фенотипическим варьированием
в пределах вида
2
3. Идентификация молекулы ДНК в качестве генетического материала
• Данные многих генетиков, включаяГ.Менделя, согласуются с четырьмя
перечисленными свойствами
– Однако, химическая природа генетического
материала не может быть определена только на
основе генетических скрещиваний
• И в самом деле, идентификация ДНК в
качестве генетического материала
потребовала проведения серии
экспериментов
3
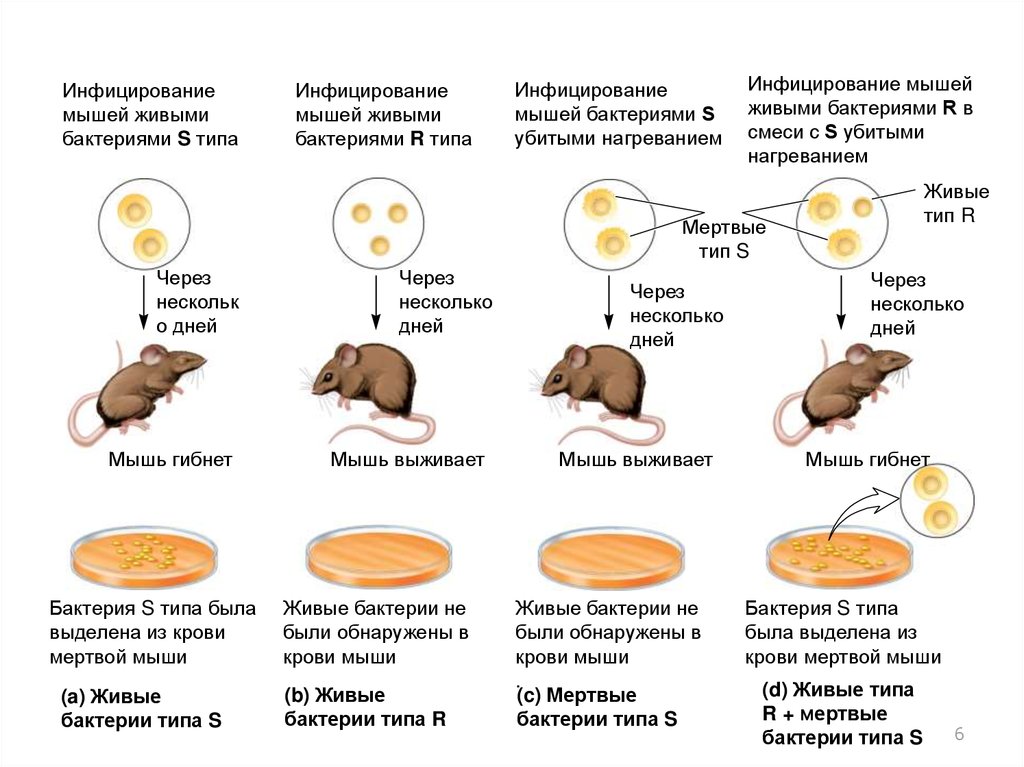
4. Эксперименты Ф.Гриффитца с бактерией Streptococcus pneumoniae
• Гриффитц (1928) изучал бактерию (пневмококка)сейчас известную как Streptococcus pneumoniae
• S. pneumoniae делится на два штамма
– S тип Гладкие
• Имеют полисахаридную капсулу, защищающую бактерию
от иммунной системы животных
• Образуют гладкие колонии на питательной среде
– R тип Шероховатые
• Не способны формировать капсулу
• Образуют колонии с шероховатым внешним видом
4
5.
В 1928, Гриффитц провел эксперименты с использованием двухштаммов S. pneumoniae: тип IIIS и тип IIR
1. При инфицировании мышей живыми бактериями IIIS
2. При инфицировании мышей живыми бактериями IIR
Мыши выживали
В крови живых бактерий не обнаруживали
3. При инфицировании мышей бактериями IIIS убитыми
нагреванием
Мыши погибали
Бактерии IIIS типа обнаруживали в крови мыши
Мыши выживали
В крови живых бактерий не обнаруживали
4. При инфицировании мышей живыми бактериями IIR в
смеси с IIIS убитыми нагреванием
Мыши погибали
Бактерии IIIS типа обнаруживали в крови мыши
5
6.
Инфицированиемышей живыми
бактериями S типа
Инфицирование
мышей живыми
бактериями R типа
Инфицирование
мышей бактериями S
убитыми нагреванием
Инфицирование мышей
живыми бактериями R в
смеси с S убитыми
нагреванием
Живые
тип R
Мертвые
тип S
Через
нескольк
о дней
Мышь гибнет
Бактерия S типа была
выделена из крови
мертвой мыши
(a) Живые
бактерии типа S
Через
несколько
дней
Мышь выживает
Живые бактерии не
были обнаружены в
крови мыши
(b) Живые
бактерии типа R
Через
несколько
дней
Мышь выживает
Живые бактерии не
были обнаружены в
крови мыши
.
(c) Мертвые
бактерии типа S
Через
несколько
дней
Мышь гибнет
Бактерия S типа
была выделена из
крови мертвой мыши
(d) Живые типа
R + мертвые
бактерии типа S
6
7.
Гриффитц пришел к выводу, что что-то измертвой бактерии S типа трансформировало
бактерии R типа в S тип
Он назвал данный процесс трансформацией
Субстанцию позволившую этому произойти
назвали трансформационным элементом
Гриффитц не знал, что это за вещество
Предположил, что молекулы с генетической
информацией устойчивы к высоким температурам
Большинство белков деградирует при нагревании
7
8. Эксперимент Херши и Чейза с бактериофагом T2
• В 1952, А.Херши и М.Чейз представили еще однодоказательство того, что ДНК ответственна за
передачу наследственной информации
Внутри
капсида
Они исследовали
бактериофага T2
С простой
организацией –
состоит из
ДНК внутри
капсидной
оболочки
Голова
Тело
Состоит
из белков
Хвостовые нити
ДНК и белка
Опорная плита
8
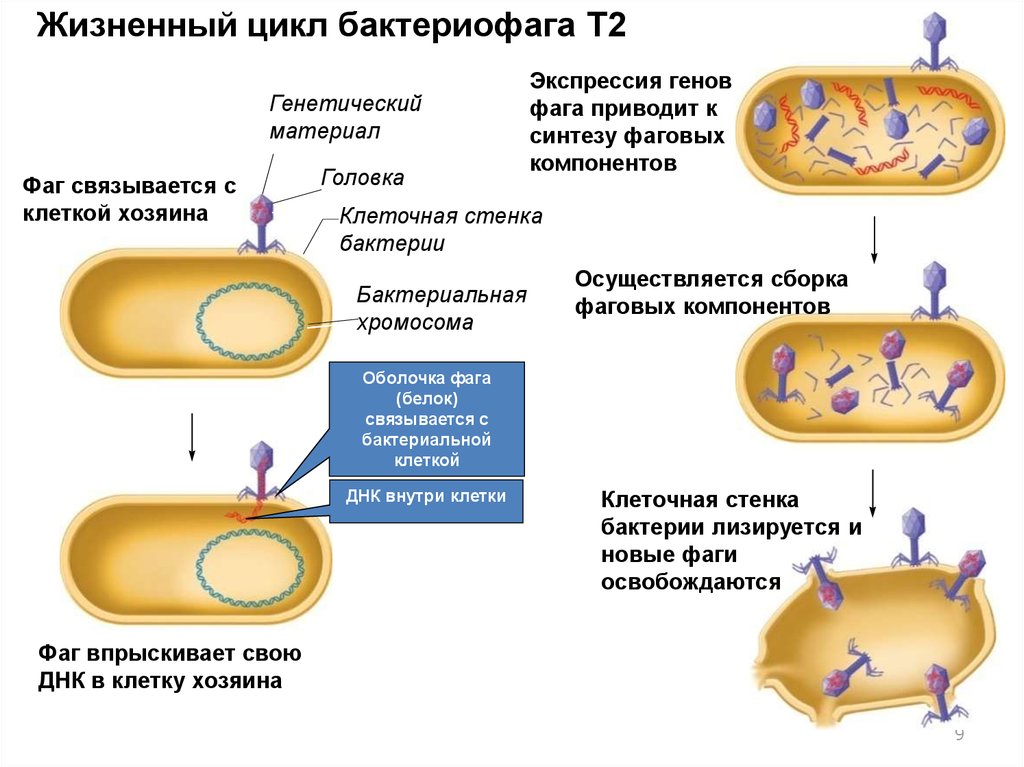
9.
Жизненный цикл бактериофага T2Генетический
материал
Фаг связывается с
клеткой хозяина
Головка
Экспрессия генов
фага приводит к
синтезу фаговых
компонентов
Клеточная стенка
бактерии
Бактериальная
хромосома
Осуществляется сборка
фаговых компонентов
Оболочка фага
(белок)
связывается с
бактериальной
клеткой
ДНК внутри клетки
Клеточная стенка
бактерии лизируется и
новые фаги
освобождаются
Фаг впрыскивает свою
ДНК в клетку хозяина
9
10.
• Краткое описание эксперимента Херши и Чейза:– Использование радиоизотопов для дифференциации ДНК и
белка
• 32P специфично меченая ДНК
• 35S специфично меченный белок
– Радиоактивно меченные фаги использовали для
инфицирования нерадиоактивные клетки Escherichia coli
– Через время, необходимое для заражения разделяли остатки
фаговых частиц и клетки E. coli
– Оценивали их радиоактивность по отдельности
Результат:
Отделенные белковые оболочки имели 35S изотоп
Бактерии содержали 32P изотоп
Некоторые из вновь образованных фаговых частиц содержали
32P изотоп, но ни одна не содержала в оболочке 35S изотоп
Вывод: для образования новых копий фага необходима ДНКфага, а белки не передаются им следовательно не несут
генетических функций
10
11. РНК является генетическим материалом у некоторых вирусов
В 1956, A. Gierer и G. Schramm выделили РНК извируса табачной мозаики (ВТМ)
Очищенная РНК
вызывала такие же
симптомы поражения
как и первоначальный
ВТМ вирус
11
12. Структура нуклеиновой кислоты
Нуклеотиды• ДНК и РНК – огромные молекулы
с несколькими уровнями
сложности
– 1. Нуклеотиды – основные
структурные единицы
нуклеиновой кислоты
– 2. Нуклеотиды связаны и
формируют линейную структуру
РНК и ДНК
– 3. Две цепи могут
взаимодействовать и
образовывать двуцепочечную
структуру
– 4. 3-D структура ДНК образуется
в результате спирализации
цепей. Взаимодействие ДНК с
белками образует хромосомы у
живых организмов
C
Одноцепочечная нить
A
TC
G
C
AA
TC
G
C
AA
TC
G
C
A A T
Двуцепочечная нить
C
G
C A
T GT
T A
A
G T
A C
GA
T
C A
T GT
T A
A
G
Трехмерная структура
12
13. Нуклеотиды
Нуклеотид это основной структурный элементДНК и РНК
Фосфатная
группа
Сахар
2
5′
HOCH2
H
O
H
H
P
H
HOCH2
H
H
3′
HO
H
4
5
2
6
3
H
N
Аденин (А)
N
H
2′
OH
D-Рибоза (в РНК))
6
5
7
H
8
N
9
4
N
1
H
3N
5
2
6
O
1N
Гуанин (Г))
2
O
Урацил (У)
в РНК
H
6
H
N
Тимин (Т) в ДНК
5
NH2
1
3N
H
NH 2
H
H
4
H
2
3
4
N
O
1′
4′
9
CH3
1N
H
OH
O
6
8
N
(в ДНК)
5′
H
D-Дезоксирибоза
O–
5
7
H
HO
O
N
2′
3′
O
OH
1′
4′
O–
Азотистые основания
Пурины
Пиримидины
NH
O
O
(двойное кольцо)
(однокольцевые)
H
4
1
N
3N
2
O
H
Цитозин (Ц)
Содержит три компонента
Фосфатную группу
Пентозный сахар
Азотистое основание
13
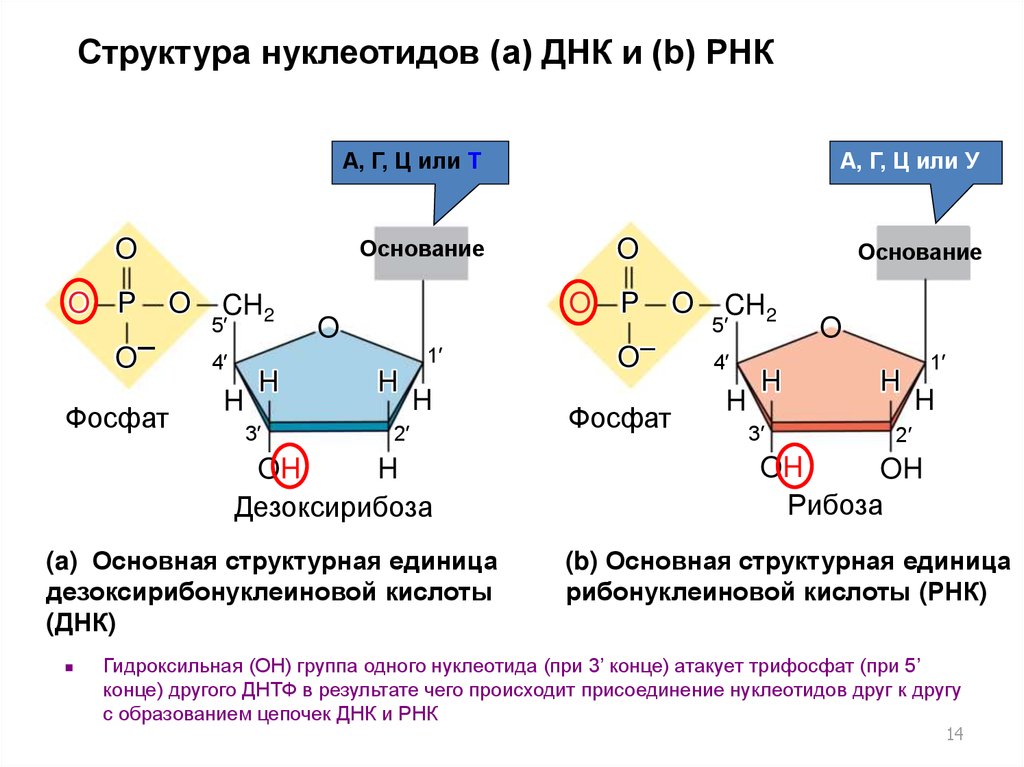
14.
Структура нуклеотидов (a) ДНК и (b) РНКA, Г, Ц или T
Основание
O
O P
O
O–
Фосфат
CH2
5′
4′
H
H
3′
O
O
O
1′
H
H
2′
OH
H
Дезоксирибоза
(a) Основная структурная единица
дезоксирибонуклеиновой кислоты
(ДНК)
A, Г, Ц или У
P
Основание
O
O–
Фосфат
CH2
5′
4′
H
H
3′
O
H
1′
H
2′
OH
OH
Рибоза
(b) Основная структурная единица
рибонуклеиновой кислоты (РНК)
Гидроксильная (ОН) группа одного нуклеотида (при 3’ конце) атакует трифосфат (при 5’
конце) другого ДНТФ в результате чего происходит присоединение нуклеотидов друг к другу
с образованием цепочек ДНК и РНК
14
15.
Основание + сахар нуклеозидПример
Аденин + рибоза = Аденозин
Аденин + дезоксирибоза = Дезоксиаденозин
Основание + сахар + фосфат(ы) нуклеотид
Пример
Аденозин монофосфат (АМФ)
Аденозин дифосфат (АДФ)
Аденозин трифосфат (АТФ)
15
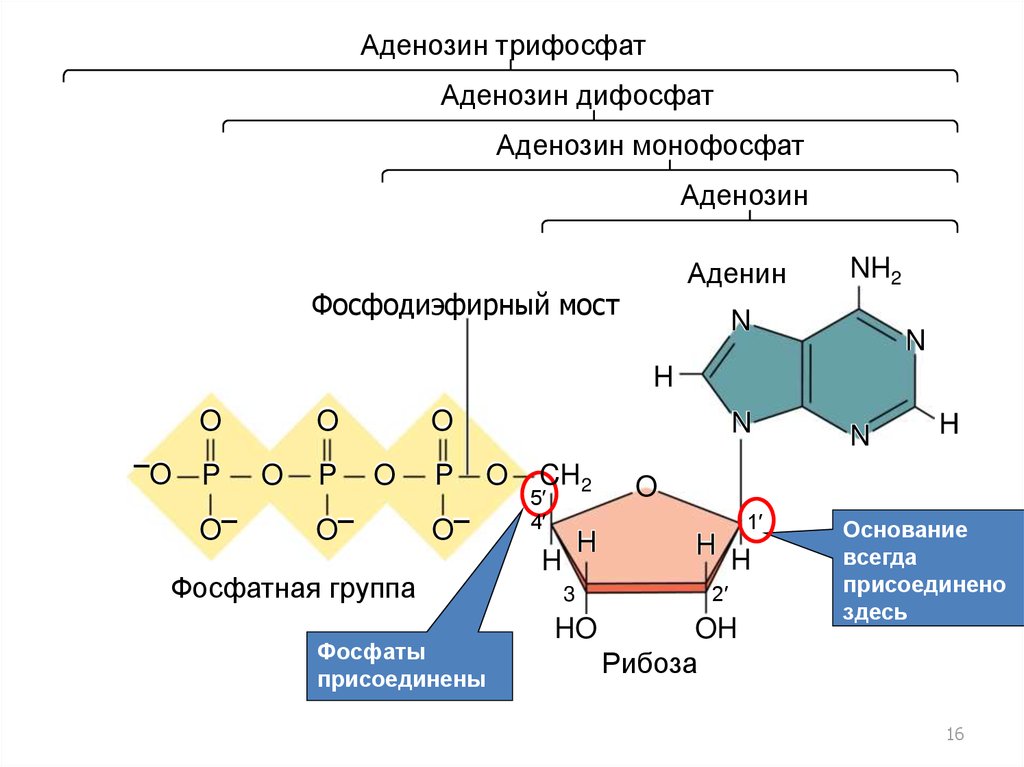
16.
Аденозин трифосфатАденозин дифосфат
Аденозин монофосфат
Аденозин
Аденин
Фосфодиэфирный мост
NH2
N
N
H
O
–O
P
O–
O
O
P
O
O
O–
P
N
O
O–
Фосфатная группа
Фосфаты
присоединены
здесь
CH2
5′
4′
H
H
3
HO
N
O
H
1′
H
2′
OH
Рибоза
Основание
всегда
присоединено
здесь
16
17.
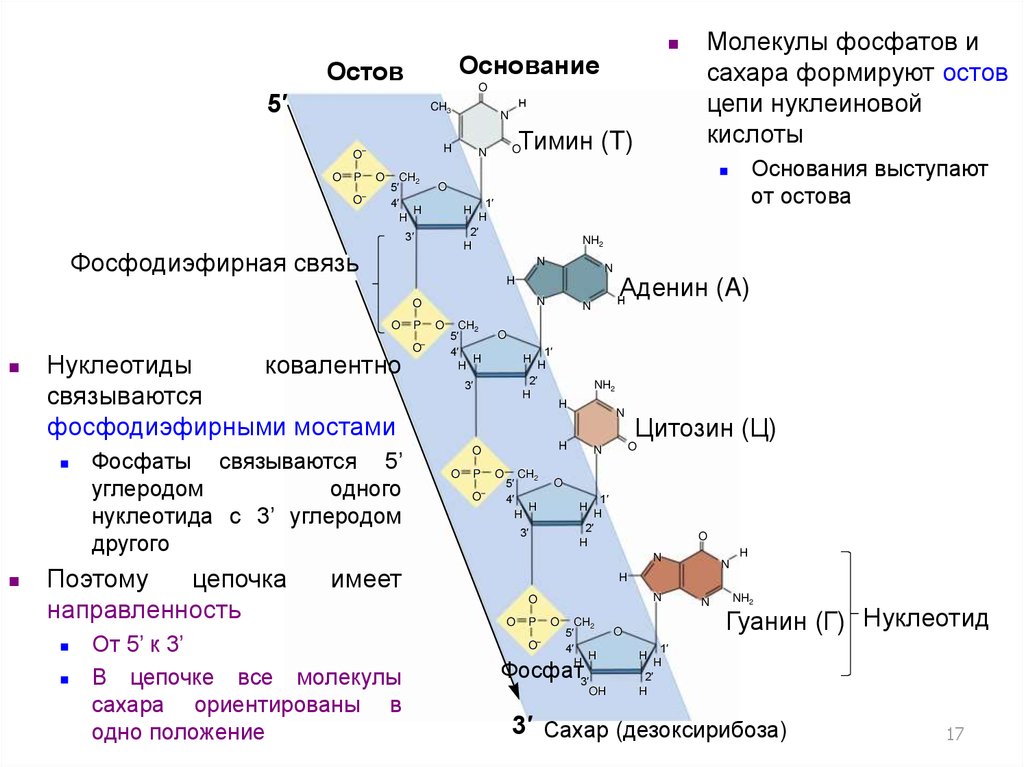
ОснованиеОстов
O
5′
CH3
P
O–
N
H
O–
O
O
CH2
5′
4′
H
H
Тимин (T)
O
N
1′
H
2′
H
H
Фосфодиэфирная связь
NH2
N
O
Нуклеотиды
ковалентно
связываются
фосфодиэфирными мостами
Фосфаты связываются 5’
углеродом
одного
нуклеотида с 3’ углеродом
другого
Поэтому
цепочка
направленность
N
Аденин (A)
H
P
O–
O
CH2
5′
4′
H
H
O
1′
H
2′
H
H
3′
P
O–
NH2
H
N
H
O
O
N
O
CH2
5′
4′
H
H
Цитозин (Ц)
O
N
O
1′
H
2′
H
H
3′
O
H
N
имеет
От 5’ к 3’
В цепочке все молекулы
сахара ориентированы в
одно положение
Основания выступают
от остова
N
O
Молекулы фосфатов и
сахара формируют остов
цепи нуклеиновой
кислоты
O
3′
N
H
N
O
O
P
O–
O
CH2
5′
4′
H
H
Фосфат3′
OH
O
N
NH2
Гуанин (Г) Нуклеотид
1′
H
2′
H
H
3′ Сахар (дезоксирибоза)
17
18. Открытие структуры ДНК
В 1953 Джеймс Ватсон и Френсис Крик открылидвуспиральную структуру ДНК
Научные основы для
прорыва были обеспечены
исследованиями:
Linus Pauling
Rosalind Franklin и Maurice
Wilkins
Erwin Chargaff
(a) Watson и Crick
(b) Оригинальная
18
двуспиральная модель ДНК
19. Двуспиральная модель ДНК
Основные структурные особенностиДве цепочки закручиваются вокруг общей оси
10 оснований и 3.4 нм на один виток вокруг оси
Две цепочки являются антипараллельными
Одна имеет 5’ - 3’ направление, а другая 3’ - 5’
Спираль правозакрученная
Двуцепочечная структура стабилизирована
1. Водородными связями между комплементарными
основаниями
A связывается двумя водородными связями с T
Ц связывается тремя водородными связями с Г
2. Расположение оснований
В ДНК основания расположены таким образом, что их
плоские части соприкасаются друг с другом
19
20.
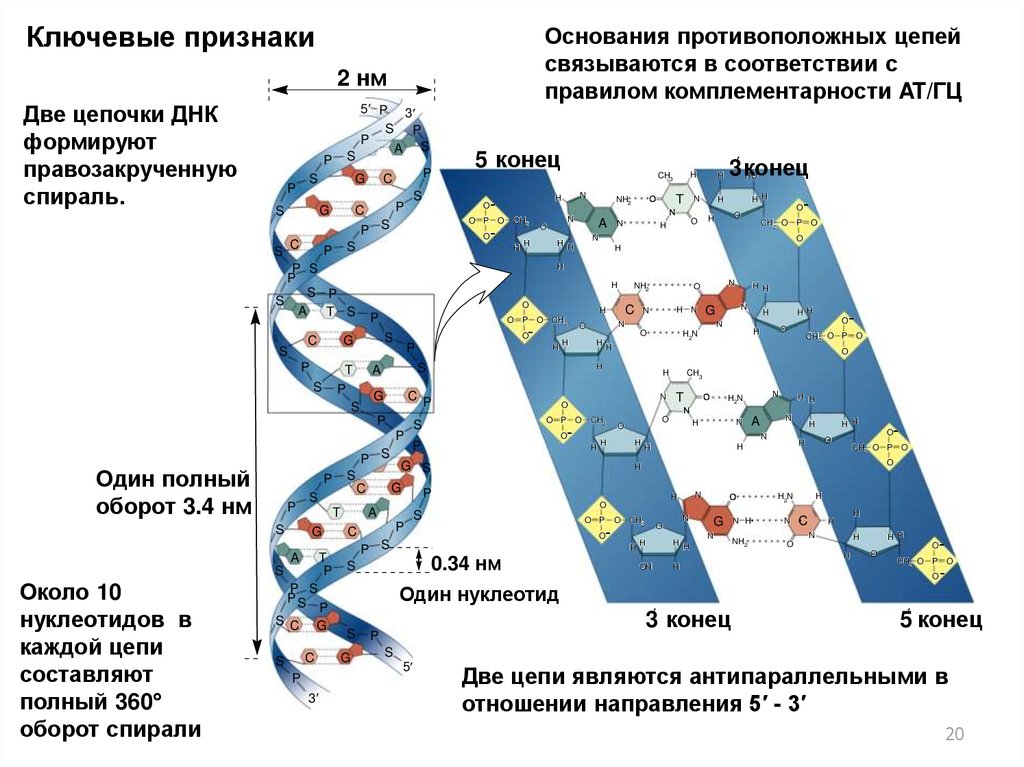
Ключевые признакиОснования противоположных цепей
связываются в соответствии с
правилом комплементарности AT/ГЦ
2 нм
Две цепочки ДНК
формируют
правозакрученную
спираль.
S
P
P
5 P 3
S P
P
A S
S
G
G
S
P
C
S
P
C
O
C
C
H
P
O P
S
P
P
S
G
C P
P S
S
A
T
P
S
Около 10
нуклеотидов в
каждой цепи
составляют
полный 360°
оборот спирали
G
C
S
P
3
-
CH2
-
CH2 O P
NH2
C
O
H
H N
N
O
O
H
O
P
P S
H
O
H
T
H
N
H2N
N
H
H H
N
A
N
-
O
O
H
H
H
H H
H
CH2 O P O
O
H
N
H
S
O P
-
O
Один нуклеотид
G
O
H
H
H
H2N
N
CH2
O
0.34 нм
P
-
CH2 O P
CH3
O
O
H
S
O
O
O
P
P
H H
H
H
H
CH2
H
N
G
H2N
N
-
H H
N
O
O P
N
O
O
C
G
O
O
H
N
O
A
S
H H
H
H
H
S
G
C
P S
PS
P
S C
H
G S
S
T
G
O
H
HO
O
H
O
H
A
P
S
N
N
3 конец
N
H
O
P
P
H
A
O
P
S
Один полный
оборот 3.4 нм
N
O
T
NH2
H
T
P
N
H
H
G
S
-
S
P
P S
P
S P
S
A T S
S
O
CH2
H
CH3
H
-
O P O
P S
S
5 конец
N
H
N H
N
NH2
O
C
H
H
N
H
OH
H
H H
H
O
-
O
CH2 O P
O
3 конец
-
O
5 конец
S
5
Две цепи являются антипараллельными в
отношении направления 5′ - 3′
20
21.
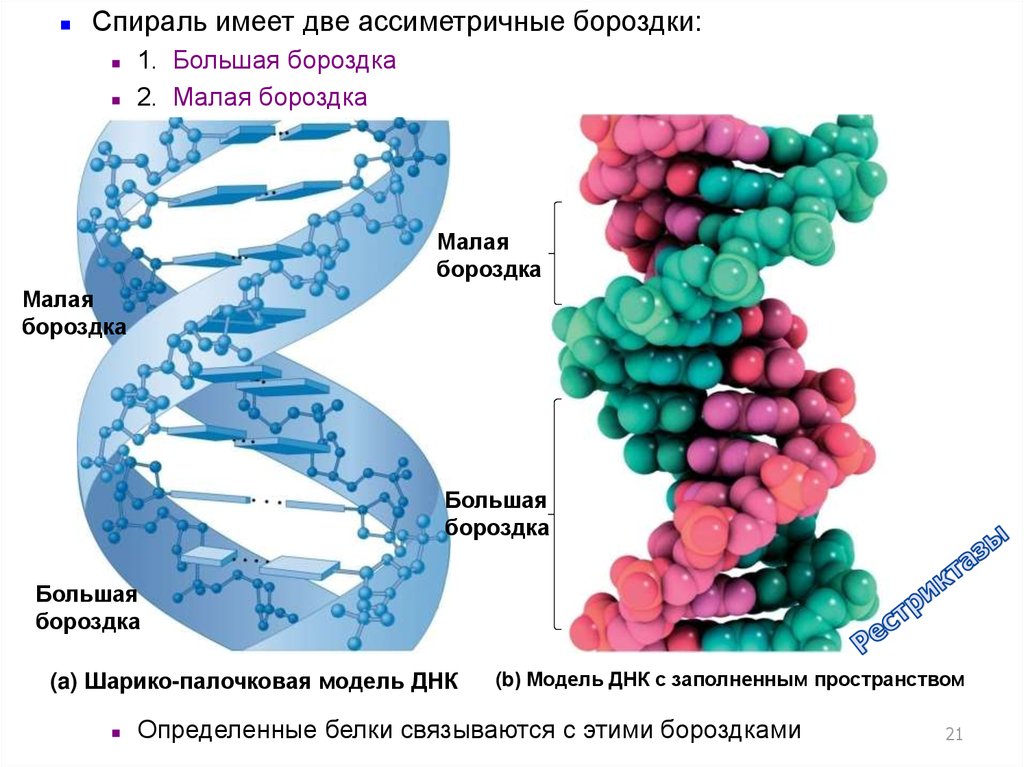
Спираль имеет две ассиметричные бороздки:1. Большая бороздка
2. Малая бороздка
Малая
бороздка
Малая
бороздка
Большая
бороздка
Большая
бороздка
(a) Шарико-палочковая модель ДНК
(b) Модель ДНК с заполненным пространством
Определенные белки связываются с этими бороздками
21
22. Трехмерная структура ДНК
Радиальные петли(300 нм в диаметре)
Метафазная
хромосома
30 нм фибры
Нуклеосомы
(11 нм в диаметре)
ДНК
(диаметр 2 нм)
Гистоновый
ДНК обвивается
белок
вокруг гистоновых
белков
Каждая хроматида
(700 нм в диаметре)
Для размещения внутри живой клетки
спираль ДНК должна быть компактизована
в 3-D конформацию
Это осуществляется с помощью ДНКсвязывающих белков
22
23.
2324.
РЕПЛИКАЦИЯ ДНККогда Уотсон и Крик предложили модель двойной спирали ДНК, то эта
модель должна была содержать в себе идею того как
воспроизводится ген.
То есть каким образом наследственная информация хранителем
которой является молекула ДНК воспроизводится в ряду поколений
и передаётся от материнской клетки к дочерней.
Их модель была хороша именно тем, что она объясняла этот факт. То
есть она содержала в себе идею воспроизведения гена.
В основе воспроизведения гена лежит процесс удвоения или
репликации ДНК.
24
25. Репликация ДНК
Репликация ДНК – этопроцесс удвоения
генетического материала
Репликация проходит
очень быстро и очень
аккуратно в S-периоде
клеточного цикла
Репликация
основывается на
комплементарности
цепочек ДНК
AT/ГЦ правило
Чаргаффа
Цепочки ДНК
разъединяются,
каждая цепь служит
матрицей для синтеза
новой цепи
а) Механизм репликации ДНК
Идентичные
последовательности
оснований
b) Результат репликации
26.
2627.
Уотсон и Крик предложили следующее:Поскольку ДНК содержит две нити, две цепи связанные между собой
более слабыми водородными связями, то можно сообщив молекуле
энергию разорвать водородные связи (этот процесс называется
плавлением или денатурацией ДНК).
При этом освобождаются две цепи и у них обнажаются азотистые
основания которые в них содержатся. В результате этого можно
каждую из этих цепей использовать как матрицу для синтеза новой
цепи.
Этот синтез управляется двумя основными принципами, которые и
лежат в основе структуры ДНК – это комплементарность и
антипараллельность.
27
28.
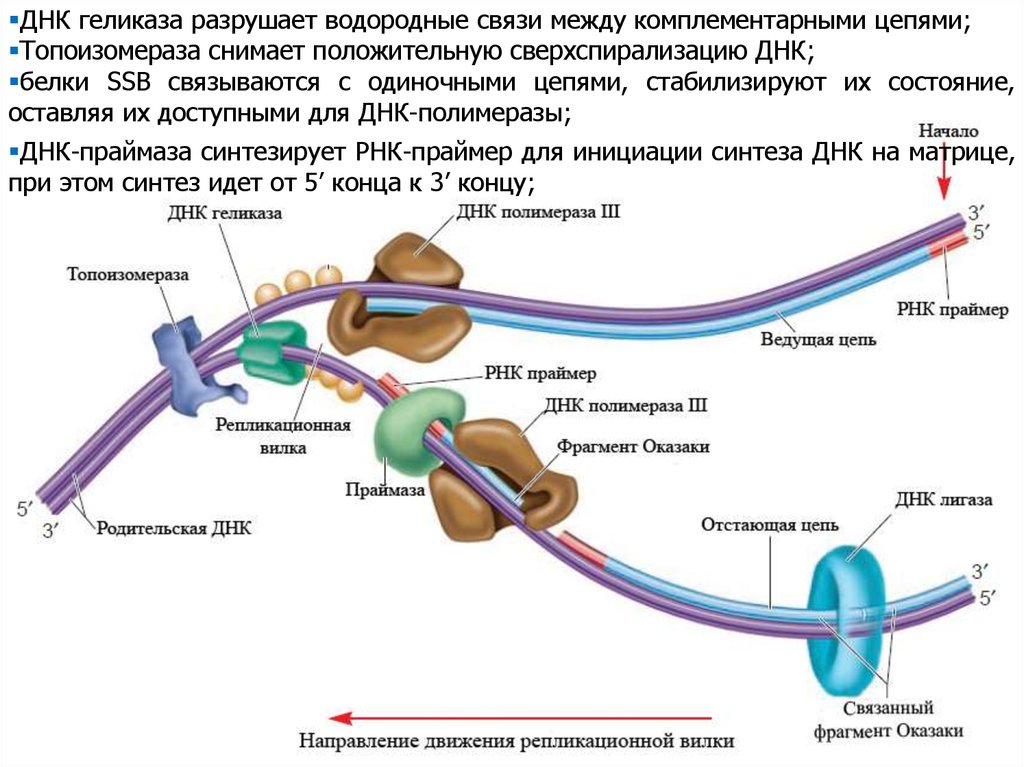
ДНК геликаза разрушает водородные связи между комплементарными цепями;Топоизомераза снимает положительную сверхспирализацию ДНК;
белки SSB связываются с одиночными цепями, стабилизируют их состояние,
оставляя их доступными для ДНК-полимеразы;
ДНК-праймаза синтезирует РНК-праймер для инициации синтеза ДНК на матрице,
при этом синтез идет от 5’ конца к 3’ концу;
28
29.
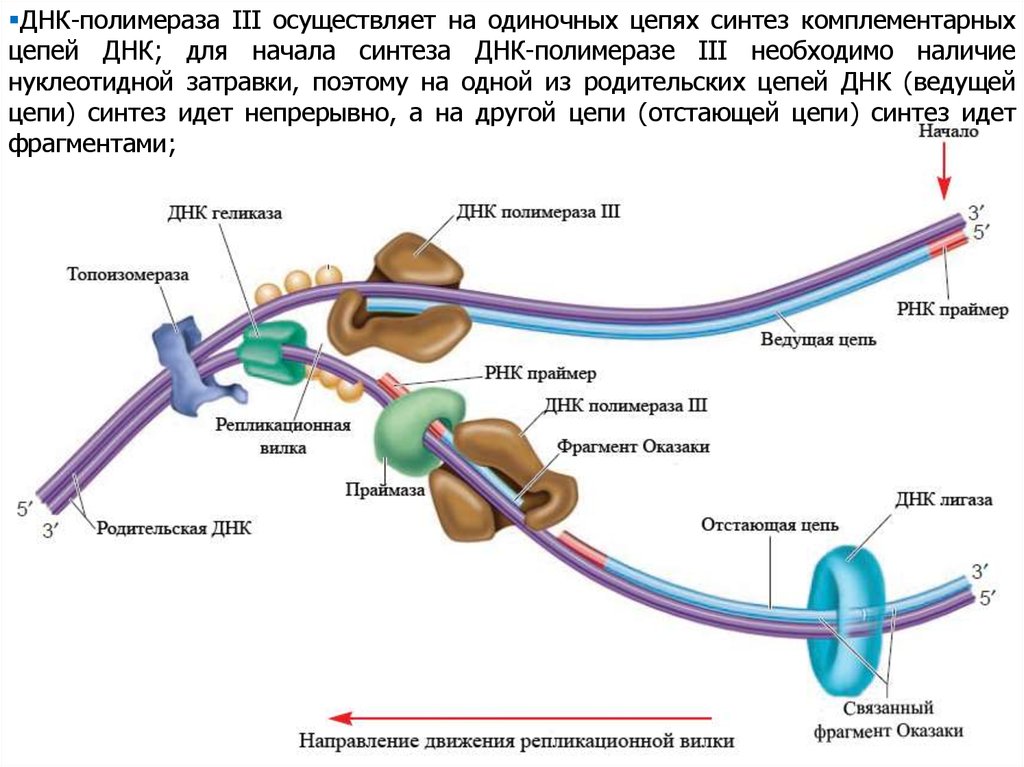
ДНК-полимераза III осуществляет на одиночных цепях синтез комплементарныхцепей ДНК; для начала синтеза ДНК-полимеразе III необходимо наличие
нуклеотидной затравки, поэтому на одной из родительских цепей ДНК (ведущей
цепи) синтез идет непрерывно, а на другой цепи (отстающей цепи) синтез идет
фрагментами;
29
30.
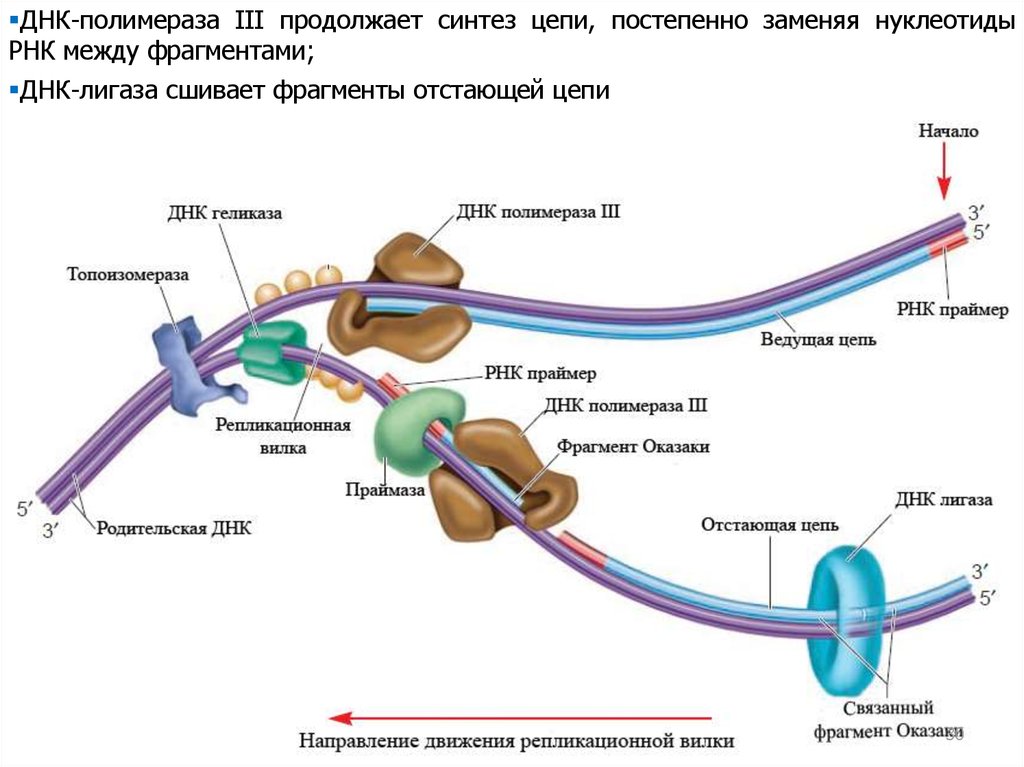
ДНК-полимераза III продолжает синтез цепи, постепенно заменяя нуклеотидыРНК между фрагментами;
ДНК-лигаза сшивает фрагменты отстающей цепи
30
31. Структура РНК
5′Остов
O
H
H
N
Урацил (У)
O–
O
P
O
–
O
5′
4′
CH2
H
H
O
N
O
1′
H
2′
OH
H
H
3′
NH2
N
Фосфодиэфирная
связь
N
O
O
P
O
5′
4′
CH2
H
В среднем длина
цепи РНК от
нескольких сотен
до нескольких
тысяч нуклеотидов
При синтезе РНК в
качестве матрицы Нуклеотид
РНК
используется
только одна цепь
ДНК
1′
H
2′
OH
H
NH2
H
N
H
P
O
Аденин (А)
H
O
O
H
N
O
3′
N
H
O–
Структура РНК во
многом схожа со
структурой ДНК
Основания
O
–
5′
4′
CH2
O
H
H
Цитозинe (Ц)
O
N
H
1′
H
Гуанин (Г)
2′
OH
3′
O
H
N
N
H
N
O
O
P
O
–
O
5′
4′
Фосфат
3′
CH2
H
H
3′
OH
N
NH2
O
H
1′
H
2′
OH
Сахар (рибоза)
31
32.
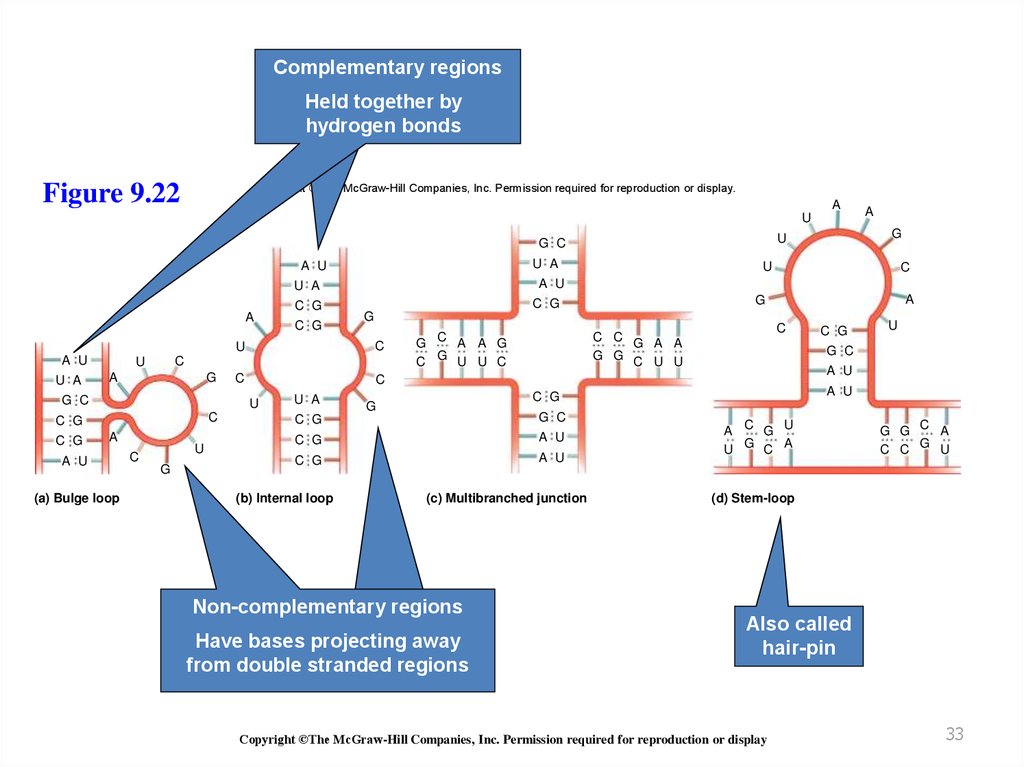
Обычно РНК молекулы являютсяодноцепочечными, но могут формировать
короткие двуцепочечные участки
Вторичные структуры образуются в следствие
комплементарного спаривания оснований
This allows short regions to form a double helix
RNA double helices typically
AсУиЦсГ
Are right-handed
Have the A form with 11 to 12 base pairs per turn
Different types of RNA secondary structures are
possible
Refer to Figure 9.22
Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display
32
33.
Complementary regionsHeld together by
hydrogen bonds
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Figure 9.22
A
U
U A
A U
A U
U A
A
U
C
G
C
C
U
C
C G
A
A U
(a) Bulge loop
C G
U
C
G
U A
C
G
A A G
G
C
U U C
A
C
C C
G A A
G G
C U U
G C
C G
A U
C G
A U
(c) Multibranched junction
Non-complementary regions
Have bases projecting away
from double stranded regions
C G
U
G C
A U
A U
C G
G
C G
(b) Internal loop
C
G
C G
G
C
U
G C
C G
C G
U
A U
U A
A
G
U
G C
A
C
U
A
G
G
U
C A
C
G G
A
C C G U
(d) Stem-loop
Also called
hair-pin
Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display
33
34.
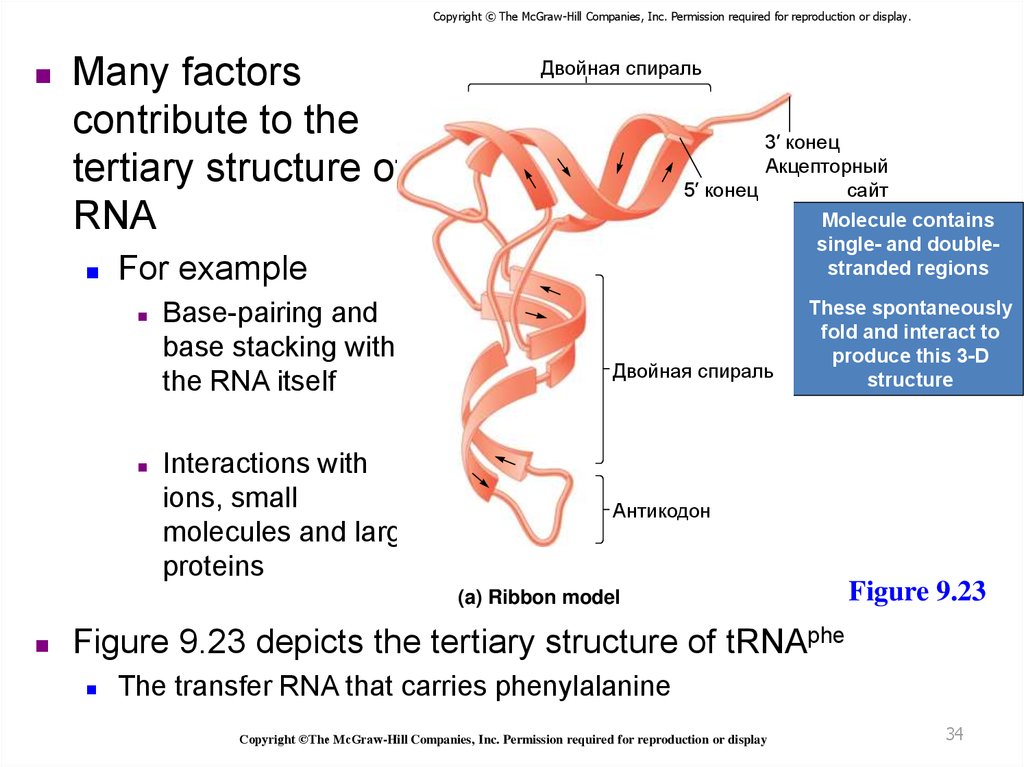
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.Many factors
contribute to the
tertiary structure of
RNA
Двойная спираль
3′ конец
Акцепторный
сайт
5′ конец
Molecule contains
single- and doublestranded regions
For example
Base-pairing and
base stacking within
the RNA itself
Interactions with
ions, small
molecules and large
proteins
Двойная спираль
These spontaneously
fold and interact to
produce this 3-D
structure
Антикодон
(a) Ribbon model
Figure 9.23
Figure 9.23 depicts the tertiary structure of tRNAphe
The transfer RNA that carries phenylalanine
Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display
34
35.
Транскрипция гена36. ВВЕДЕНИЕ
• ДНК хранит информацию– Для построения механизма (организма) обеспечивающего
– Не является самодостаточной молекулой
• Для построения и поддержания организма
информация ДНК должна быть декодирована
– Конечная цель: Образование активных молекул РНК и
белков, которые выполняют структурную функцию в клетке и
катализируют химические реакции
• Два шага: транскрипция и трансляция
37. Центральная догма генетики
Репликация ДНК: образование копий ДНК,передаваемых от клетки к клетке, от
родителя потомку
ДНК хромосомы: хранит генетическую
информацию в виде генов
Транскрипция: образование РНК копий генов
мРНК: временная копия гена, содержащая
информацию о синтезируемом полипептиде
Трансляция: образования полипептида на
основе информации мРНК
Функциональные РНК:
тРНК,
рРНК,
мРНК и др.
Полипептид: становится частью
функционального белка
38.
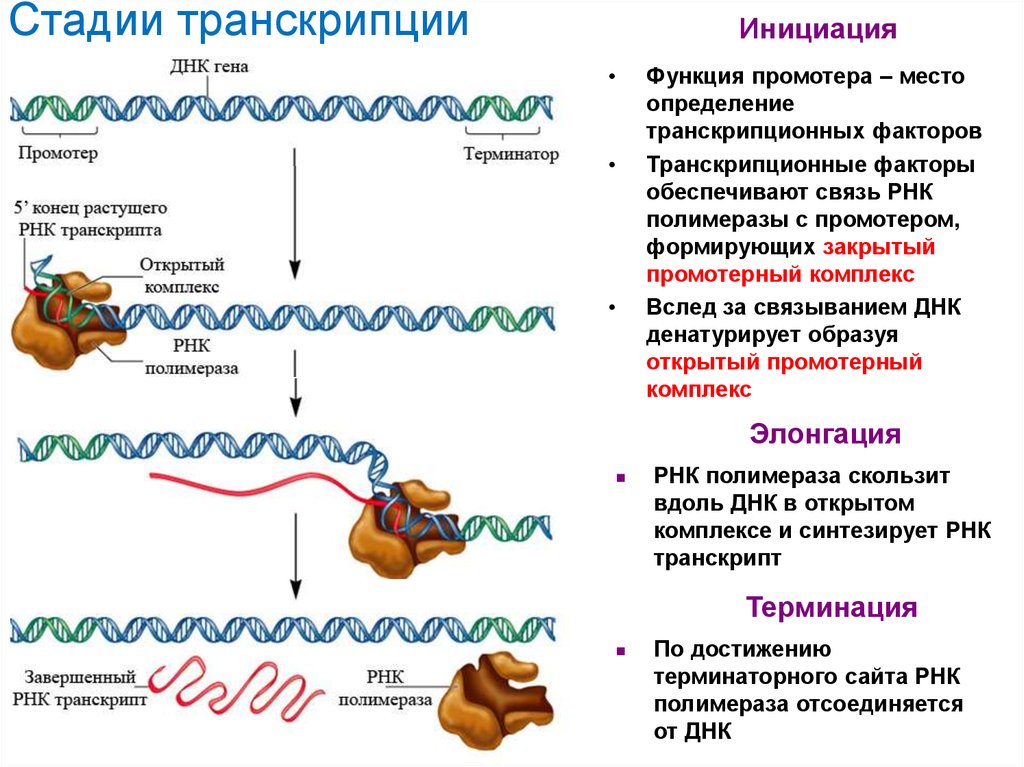
Стадии транскрипцииИнициация
Функция промотера – место
определение
транскрипционных факторов
Транскрипционные факторы
обеспечивают связь РНК
полимеразы с промотером,
формирующих закрытый
промотерный комплекс
Вслед за связыванием ДНК
денатурирует образуя
открытый промотерный
комплекс
Элонгация
РНК полимераза скользит
вдоль ДНК в открытом
комплексе и синтезирует РНК
транскрипт
Терминация
По достижению
терминаторного сайта РНК
полимераза отсоединяется
от ДНК
39. Структура гена
Ген может быть определен как дискретный участок ДНКтранскрибируемый в РНК
В процессе экспрессии гена, последовательности ДНК
окружающие его определяют:
Ген так же называют транскрипционной единицей
Молекула РНК образующаяся на основе гена называется
транскрипт
Силу экспрессии гена
Время экспрессии
Место (ткань или тип клеток) экспрессии гена
Транскрипция осуществляется большим ферментативным комплексом
называемым РНК полимераза
В большинстве случаев РНК полимераза связывается со специфической
последовательностью ДНК перед геном, называемой промотером
Промотер привлекает РНК полимеразу к гену и «сообщает» ферменту о гене
40.
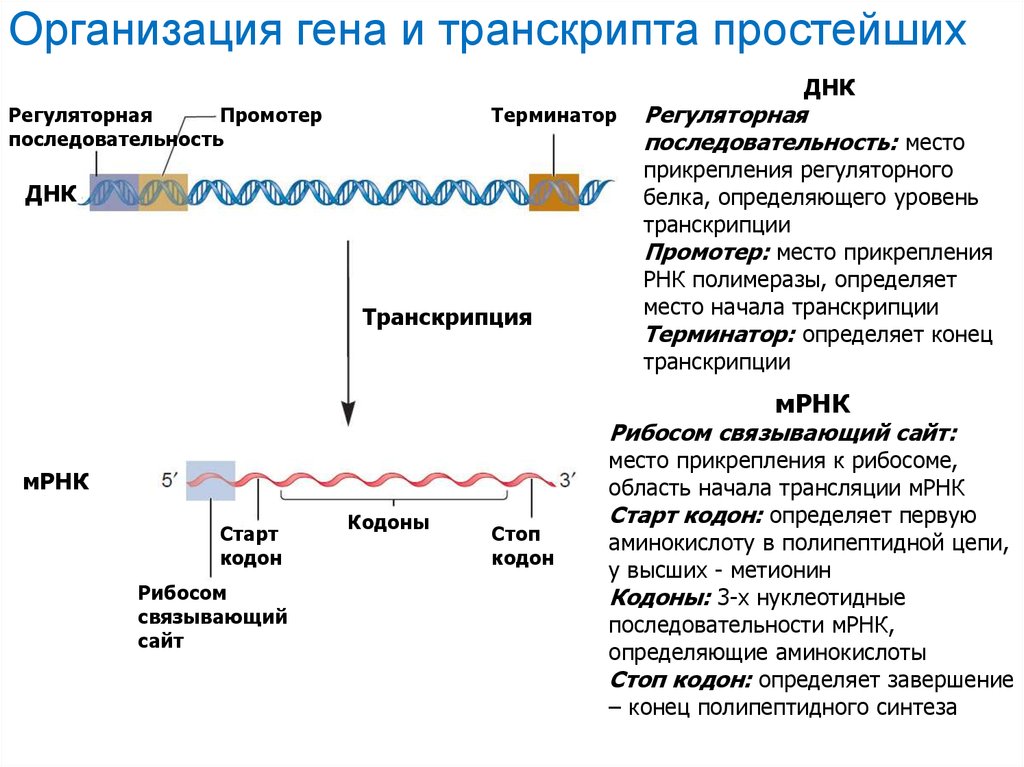
Организация гена и транскрипта простейшихДНК
Регуляторная
Промотер
последовательность
Терминатор
ДНК
Транскрипция
Регуляторная
последовательность: место
прикрепления регуляторного
белка, определяющего уровень
транскрипции
Промотер: место прикрепления
РНК полимеразы, определяет
место начала транскрипции
Терминатор: определяет конец
транскрипции
мРНК
Рибосом связывающий сайт:
место прикрепления к рибосоме,
мРНК
Старт
кодон
Рибосом
связывающий
сайт
Кодоны
Стоп
кодон
область начала трансляции мРНК
Старт кодон: определяет первую
аминокислоту в полипептидной цепи,
у высших - метионин
Кодоны: 3-х нуклеотидные
последовательности мРНК,
определяющие аминокислоты
Стоп кодон: определяет завершение
– конец полипептидного синтеза
41.
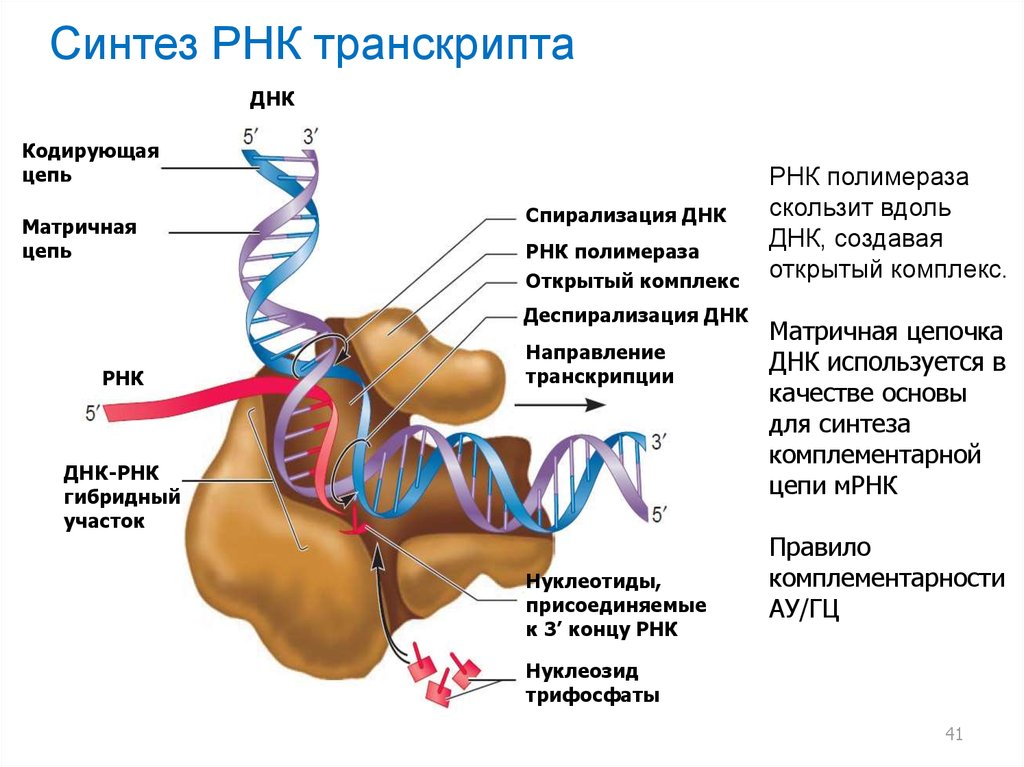
Синтез РНК транскриптаДНК
Кодирующая
цепь
Матричная
цепь
Спирализация ДНК
РНК полимераза
Открытый комплекс
Деспирализация ДНК
РНК
Направление
транскрипции
ДНК-РНК
гибридный
участок
Нуклеотиды,
присоединяемые
к 3’ концу РНК
РНК полимераза
скользит вдоль
ДНК, создавая
открытый комплекс.
Матричная цепочка
ДНК используется в
качестве основы
для синтеза
комплементарной
цепи мРНК
Правило
комплементарности
АУ/ГЦ
Нуклеозид
трифосфаты
41
42. Полипептидный синтез
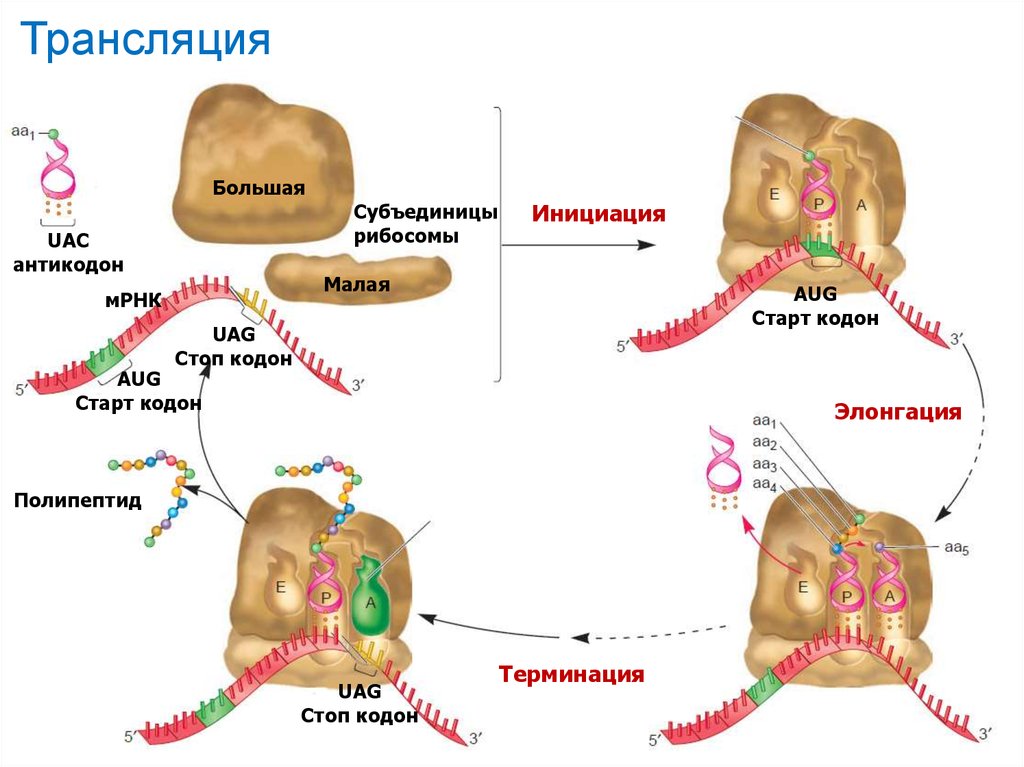
Процесс синтеза белка на матричной мРНК называется трансляцией, его
основные этапы:
активизация определенной аминокислоты и её присоединение к тРНК,
образование молекулярного комплекса: аминокислота-фермент аминоацил-тРНКсинтетаза -тРНК;
образование пептидных связей между отдельными аминокислотами,
происходящее с участием рибосом: рибосома присоединяется к 5’ концу
молекулы мРНК и считывает информацию от 5’ конца к 3’ концу; тРНК, несущие
аминокислоту внутри рибосомы связываются своими антикодонами с
соответствующими кодонами мРНК; в результате выстраивается цепочка
аминокислот в соответствии с закодированной генетической информацией мРНК;
когда рибосома доходит до стоп-кодона на мРНК, синтез белковой молекулы
прекращается и она отделяется от рибосомы. При этом освободившаяся мРНК и
рибосома могут быть снова вовлечены в процесс биосинтеза новой белковой
молекулы. При значительной длине мРНК на ней одновременно могут работать
несколько рибосом (полисом), находящихся на разных стадиях синтеза
соответствующего белка;
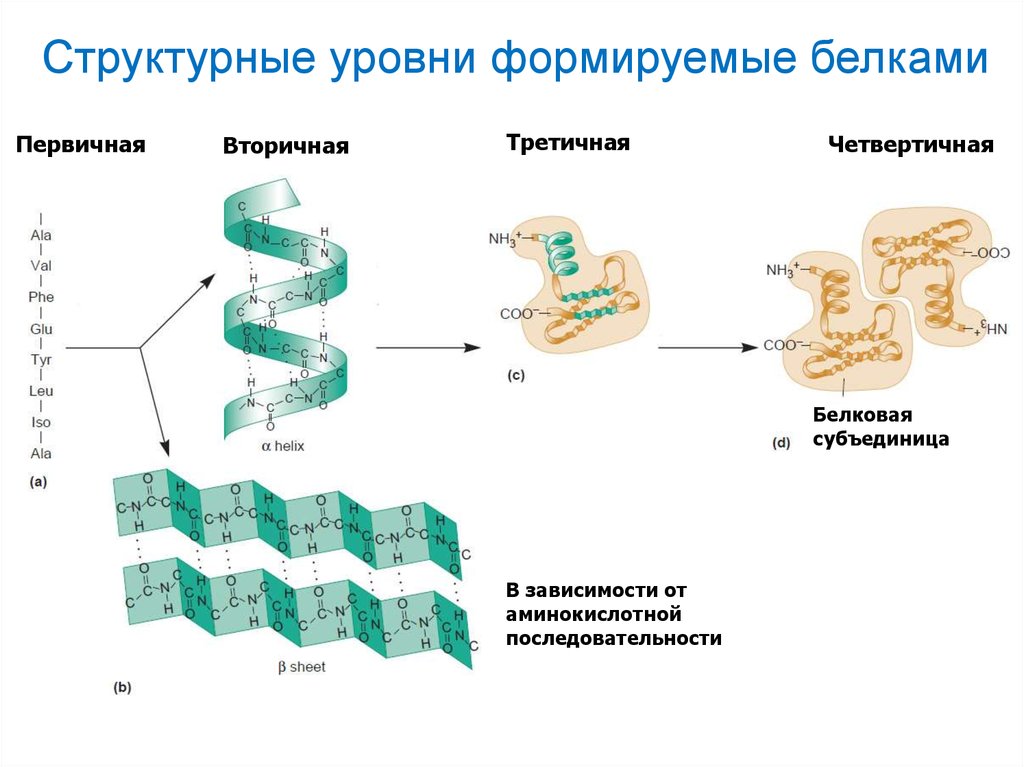
после освобождения от рибосомы белковая молекула приобретает вторичную
структуру, скручиваясь в альфа или бета-спирали; третичную структуру белки
приобретают, когда отдельные спирали объединяются в глобулы; четвертичная
структура белка представляет собой ассоциации из различных глобул; эти
преобразования белковых молекул происходят с участием особых ферментов –
шаперонов.
43.
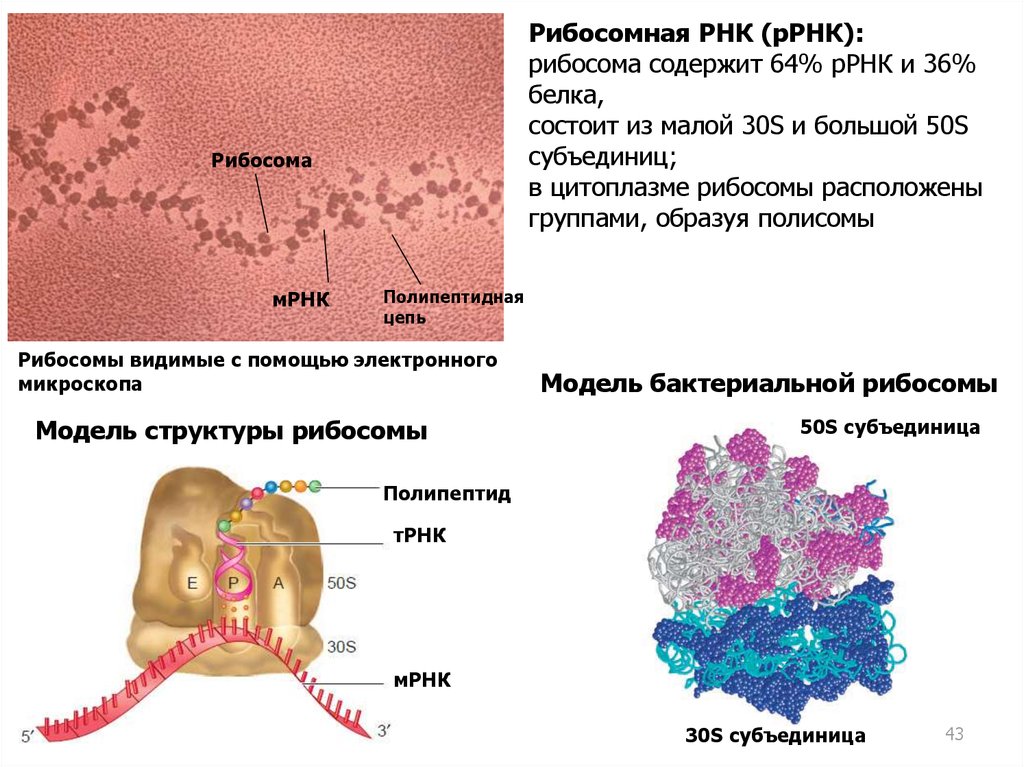
Рибосомная РНК (рРНК):рибосома содержит 64% рРНК и 36%
белка,
состоит из малой 30S и большой 50S
субъединиц;
в цитоплазме рибосомы расположены
группами, образуя полисомы
Рибосома
мРНК
Полипептидная
цепь
Рибосомы видимые с помощью электронного
микроскопа
Модель структуры рибосомы
Модель бактериальной рибосомы
50S субъединица
Полипептид
тРНК
мРНК
30S субъединица
43
44.
ТрансляцияБольшая
UAC
антикодон
Субъединицы
рибосомы
Инициация
Малая
мРНК
AUG
Старт кодон
UAG
Стоп кодон
AUG
Старт кодон
Элонгация
Полипептид
UAG
Стоп кодон
Терминация
44
45.
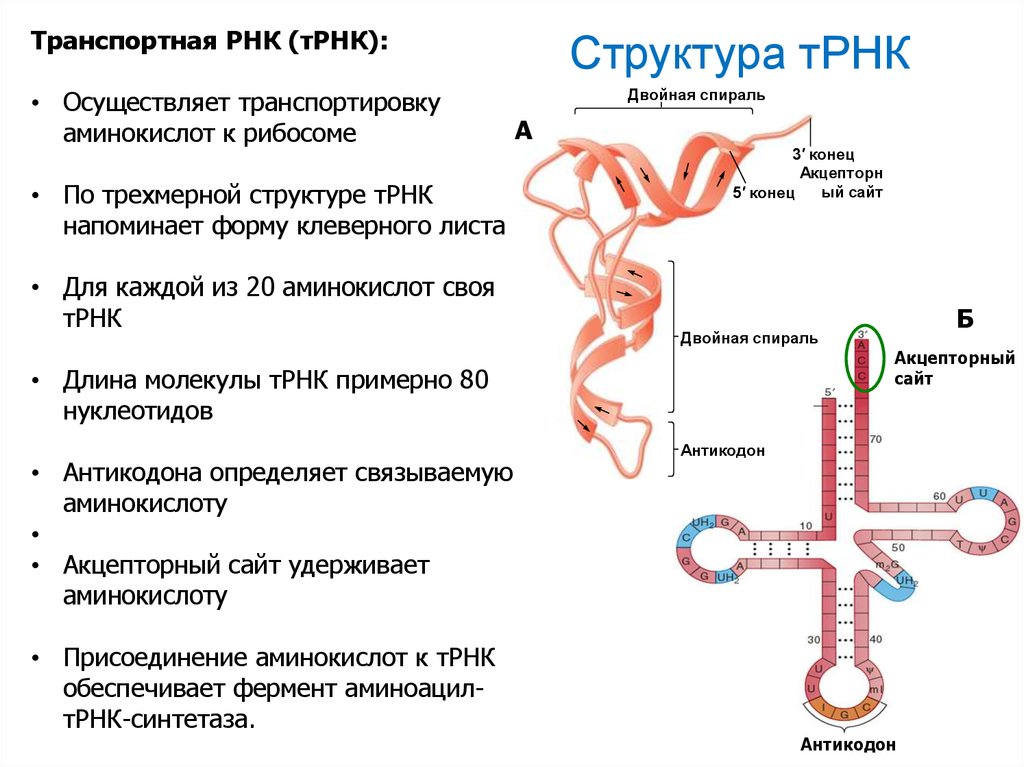
Транспортная РНК (тРНК):• Осуществляет транспортировку
аминокислот к рибосоме
• По трехмерной структуре тРНК
напоминает форму клеверного листа
• Для каждой из 20 аминокислот своя
тРНК
Структура тРНК
Двойная спираль
А
3′ конец
Акцепторн
ый сайт
5′ конец
Двойная спираль
• Длина молекулы тРНК примерно 80
нуклеотидов
• Антикодона определяет связываемую
аминокислоту
• Акцепторный сайт удерживает
аминокислоту
• Присоединение аминокислот к тРНК
обеспечивает фермент аминоацилтРНК-синтетаза.
Б
Акцепторный
сайт
Антикодон
Антикодон
46.
Структурные уровни формируемые белкамиПервичная
Вторичная
Третичная
Четвертичная
Белковая
субъединица
В зависимости от
аминокислотной
последовательности
47.
Генетический кодГенетический код триплетный каждая из 20 входящих в состав
белков аминокислот кодируется
тремя нуклеотидами (кодоном)
Ген. код вырожденный - одна
аминокислота может быть
закодирована не одним, а
несколькими триплетами
нуклеотидов, например, метионин
кодируется одним кодоном АУГ, а
валин - четырьмя кодонами ГУА,
ГУЦ, ГУГ, ГУУ
Ген. код неперекрывающийся -
началом старта считывания любого
гена является кодон АУГ, задающий
состав нуклеотидов последующих
триплетов
Ген. код универсален для
практически всех живых
организмов, от бактерий до
человека