
















































 Биология
БиологияПохожие презентации:
")









Филогенез клеток и функции клеточных мембран
1.
РАЗДЕЛ 1.СИСТЕМА
КЛЕТКА
КАК
ЦЕЛОСТНАЯ,
ДИНАМИЧЕСКАЯ
Занятие 3:
1. Филогенез эукариотической клетки.
2. Обмен веществ, роль мембран (ионные каналы).
3. Молекулярные рецепторы.
2. Филогенез эукариотической клетки
3.
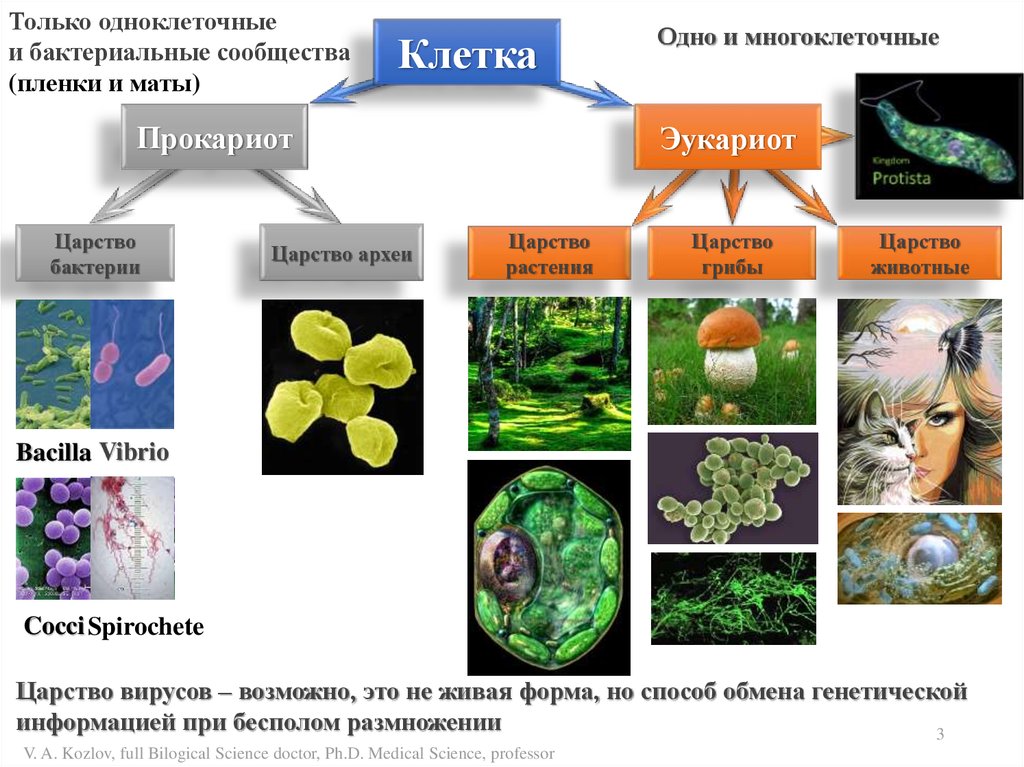
Только одноклеточныеи бактериальные сообщества
(пленки и маты)
Клетка
Прокариот
Царство
бактерии
Царство археи
Одно и многоклеточные
Эукариот
Царство
растения
Царство
грибы
Царство
животные
Bacilla Vibrio
Cocci Spirochete
Царство вирусов – возможно, это не живая форма, но способ обмена генетической
информацией при бесполом размножении
3
V. A. Kozlov, full Bilogical Science doctor, Ph.D. Medical Science, professor
4.
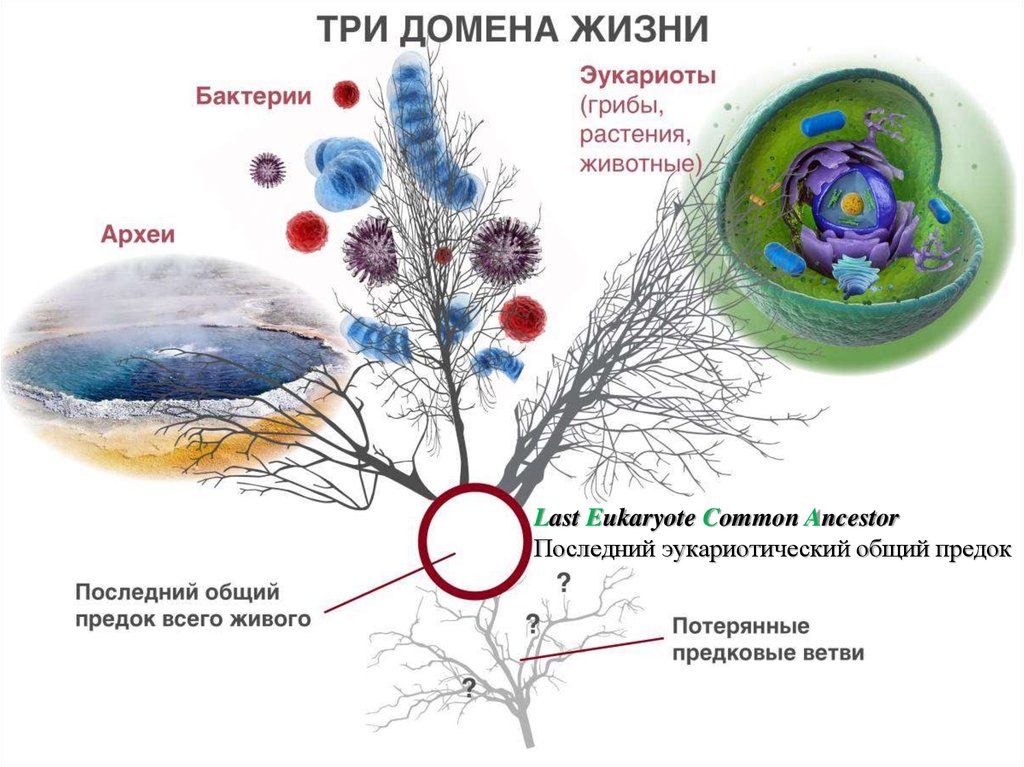
Last Eukaryote Common AncestorПоследний эукариотический общий предок
5.
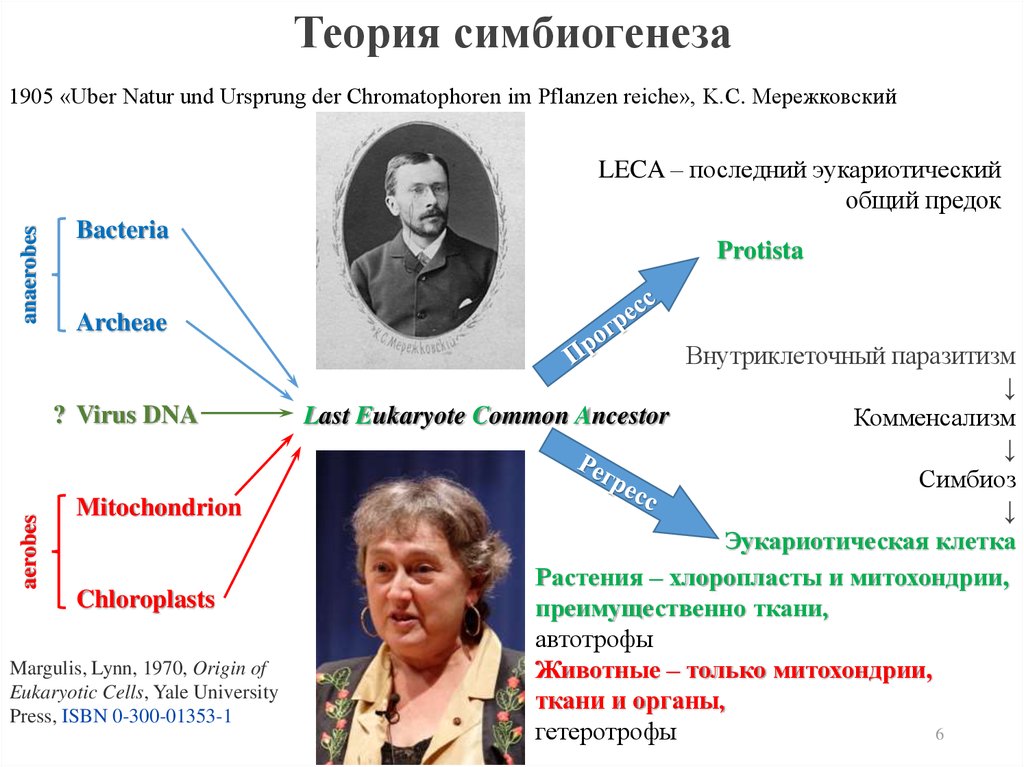
В 1905 K.С. Мережковский предположил, чтоэукариотическая клетка образовалась в
результате симбиогенеза – слияния
бактериальных клеток и митохондрий - у
животных и хлоропластов и митохондрий - у
грибов и растений.
Возможно, что клеточное ядро образовалось из
когда-то поглощенного такой симбиотической
клеткой ДНК содержащего вируса.
6.
Теория симбиогенеза1905 «Uber Natur und Ursprung der Chromatophoren im Pflanzen reiche», K.С. Мережковский
anaerobes
LECA – последний эукариотический
общий предок
Bacteria
Archeae
? Virus DNA
aerobes
Protista
Mitochondrion
Chloroplasts
Margulis, Lynn, 1970, Origin of
Eukaryotic Cells, Yale University
Press, ISBN 0-300-01353-1
Внутриклеточный паразитизм
↓
Last Eukaryote Common Ancestor
Комменсализм
↓
Симбиоз
↓
Эукариотическая клетка
Растения – хлоропласты и митохондрии,
преимущественно ткани,
автотрофы
Животные – только митохондрии,
ткани и органы,
гетеротрофы
6
7.
Р. Гупта предположил, что эукариотическая клетка возникла в результатесимбиоза грамотрицательной бактерии, относящейся к протеобактериям, и
археи. Этот симбиоз был создан в кислородной среде, богатой
антибиотиками, секретируемые другими микроорганизмами. Партнеры,
вступили в симбиоз, при условии, что:
1) археи – устойчивы к антибиотикам,
2) протеобактерии – толерантны к кислороду.
Внутриклеточный симбионт архея впоследствии был окружен клеточной
мембраной бактерий, защищающей его от действия кислорода, что
породило эндоплазматический ретикулум и ядерную мембрану (R.S. Gupta,
2005) . Gupta R.S. Molecular sequences and the early history of life // Microbial Phylogeny and Evolution / Ed. J. Sapp. Oxford
Univ. Press, 2005. Р. 160–183.
Возможно, что основой клеточного ядра стал ДНК содержащий вирус, в
который постепенно полностью мигрировали геномы протеобактерии и
археи. На заключительном этапе в симбиоз вошли митохондрии и
хлоропласты.
Radhey S. Gupta (Professor) Department of Biochemistry and
Biomedical Sciences (HSC- 4H2) 1200 Main Street West,
Hamilton, Ontario, Canada L8N 3Z5
V. A. Kozlov, full Bilogical Science doctor, Ph.D. Medical Science, professor
7
8.
Последний эукариотический общий предок имелзначительно более сложный геном, чем у современных
многоклеточных.
Многоклеточность осуществлялась через потерю ненужных
участков генома.
В процессе совмещения геномов произошло размыкание
кольцевой ДНК с преобразованием ее в линейную, возможно
это послужило причиной формирования так называемых
интронных участков ДНК, которых практически нет у
прокариот и которые на ранних этапах симбиогенеза были
нужны для «подгонки» геномов разных организмов друг к
другу.
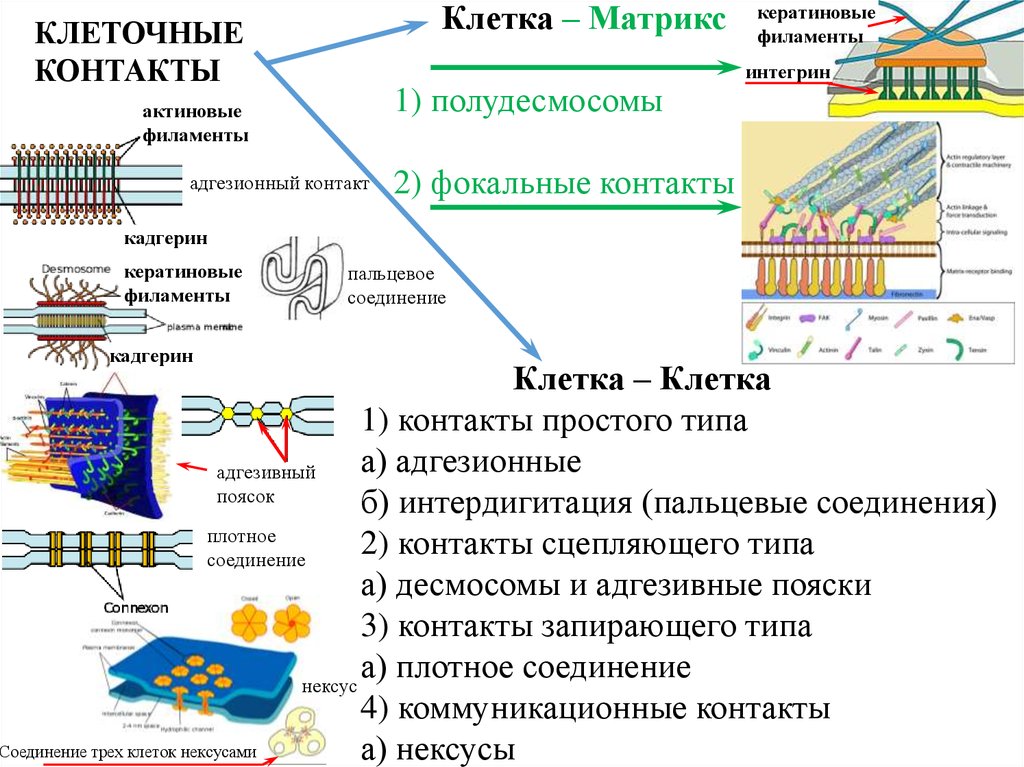
9. Типы соединения клеток
10.
Клетка – МатриксКЛЕТОЧНЫЕ
КОНТАКТЫ
1) полудесмосомы
актиновые
филаменты
адгезионный контакт
кератиновые
филаменты
интегрин
2) фокальные контакты
кадгерин
кератиновые
филаменты
кадгерин
пальцевое
соединение
Клетка – Клетка
1) контакты простого типа
а) адгезионные
адгезивный
поясок
б) интердигитация (пальцевые соединения)
плотное
2) контакты сцепляющего типа
соединение
а) десмосомы и адгезивные пояски
3) контакты запирающего типа
а) плотное соединение
нексус
4) коммуникационные контакты
Соединение трех клеток нексусами
а) нексусы
11.
12.
13.
Межклеточные контакты необходимы для1) соединения клеток
2) информационного обмена
а) межклеточного
б) с внешней средой
Для этого клетка создает
ионные каналы
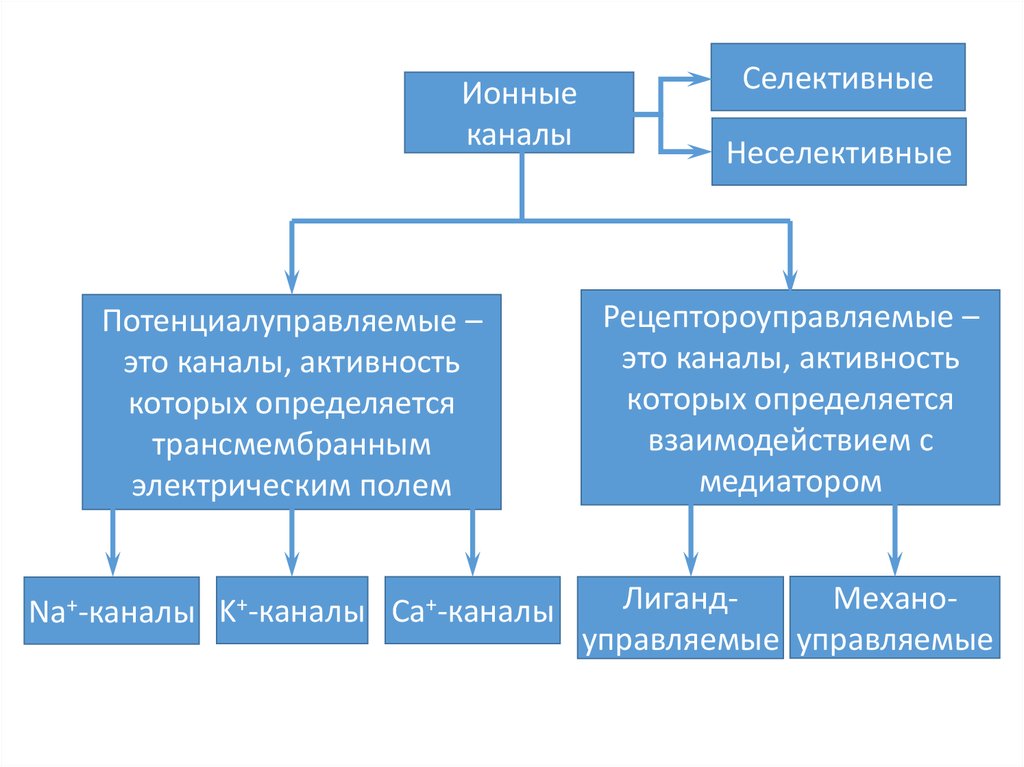
14. Ионные каналы
15.
Ионныеканалы
Селективные
Неселективные
Потенциалуправляемые –
это каналы, активность
которых определяется
трансмембранным
электрическим полем
Рецептороуправляемые –
это каналы, активность
которых определяется
взаимодействием с
медиатором
Na+-каналы K+-каналы Ca+-каналы
МеханоЛигандуправляемые управляемые
16. Общее представление о структуре и функциях ионных каналов
Каналообразующие белковые молекулы всех ионных каналов имеютнекоторые общие признаки строения и обычно представлены
большими трансмембранными белками с молекулярными
массами выше 250 кД.
• Состоят из нескольких субъединиц.
• Обычно важнейшие свойства каналов определяются их
α-субъединицей.
Она принимает участие в формировании
ионоселективной норы, сенсорного механизма
трансмембранной
разности
потенциалов
(ворот канала) и имеет места связывания для
экзогенных и эндогенных лигандов.
• Другие субъединицы, входящие в структуру ионных каналов, играют
вспомогательную роль, модулирующую свойства каналов.
17.
Трехмерная пространственная структураканалообразующей молекулы белка
располагается
в цитоплазматической
формирует
мембране
устья канала, обращенные к наружной и внутренней
сторонам мембраны,
пору, заполненную водой
«ворота» канала
от ее размеров и заряда зависят
селективность и проницаемость
ионного канала.
Проницаемость канала для данного
иона определяется 1) размерами,
2) величиной заряда и
3) гидратной оболочкой.
образуются участком пептидной цепи, который может легко
изменять свою конформацию и определять открытое или
закрытое состояние канала.
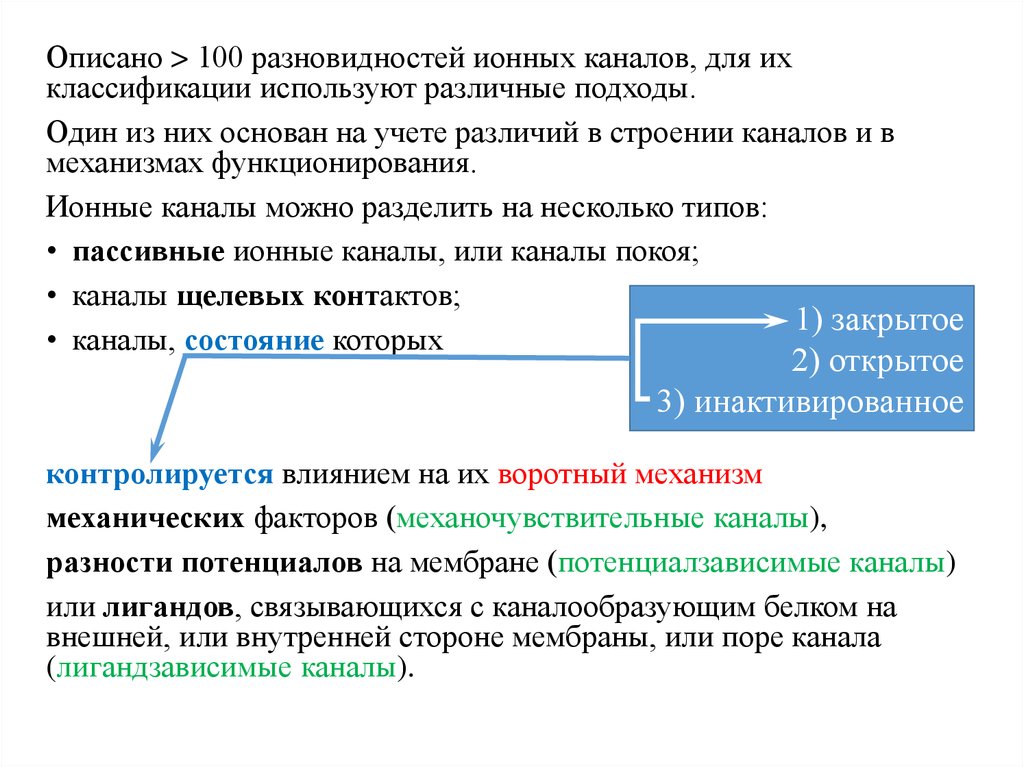
18.
Описано > 100 разновидностей ионных каналов, для ихклассификации используют различные подходы.
Один из них основан на учете различий в строении каналов и в
механизмах функционирования.
Ионные каналы можно разделить на несколько типов:
• пассивные ионные каналы, или каналы покоя;
• каналы щелевых контактов;
1) закрытое
• каналы, состояние которых
2) открытое
3) инактивированное
контролируется влиянием на их воротный механизм
механических факторов (механочувствительные каналы),
разности потенциалов на мембране (потенциалзависимые каналы)
или лигандов, связывающихся с каналообразующим белком на
внешней, или внутренней стороне мембраны, или поре канала
(лигандзависимые каналы).
19.
Пассивные каналыканалы цитоплазматических мембран
нервных волокон и их окончаний,
клеток поперечно-полосатой, гладкой
мускулатуры, миокарда и др. тканей
Формируют и поддерживают потенциал
покоя цитоплазматической мембраны клетки
могут быть открытыми (активными) в покоящихся клетках,
т.е. в отсутствие каких-либо воздействий
не являются строго селективными,
через них мембрана клетки может «протекать» для нескольких
ионов, например К+ и CI–; К+ и Na+; Ca2+.
Поэтому второе название этих каналов – каналы утечки.
20.
Каналы щелевых (плотных) контактов клеток• Каналы щелевых контактов формируются в области контакта двух
соседних клеток, очень близко (3,5 нм) прилежащих друг к другу.
• В мембране каждой контактирующей клетки шесть субъединиц белков
коннексинов формируют гексагональную структуру – коннексон, в центре
которой образуется пора или ионный канал (следующий слайд).
• В точке контакта в мембранах обеих клеток образуется зеркальная
структура из двух коннексонов, и ионный канал между ними становится
общим.
Через такие ионные каналы из клетки в клетку могут перемещаться
1) различные минеральные ионы, в том числе Са2+,
2) низкомолекулярные органические вещества.
Каналы щелевых контактов клеток обеспечивают передачу информации
между клетками миокарда, гладких мышц, сетчатки глаза, нервной системы.
21.
Мембраныдвух соседних
клеток
3,5 нм
Коннексон –
функциональный
комплекс из
коннексинов
Коннексин –
белок
Щелевой
контакт
Канал
Зеркальная структура из двух
коннексонов
22.
Ионные каналыбывают
механозависимые
потенциалзависимые
лигандзависимые
открываются и закрываются
в ответ на
механическую
деформацию
изменение потенциала
покоя мембраны
взаимодействие
низкомолекулярного
медиатора с рецепторным
участком канала
23.
Потенциалзависимые каналы• Состояние этих каналов управляется силами электрического поля,
создаваемого величиной разности потенциалов ( ) на мембране.
• Они могут находиться в неактивном (закрытом), активном (открытом) и
инактивированном состояниях, которые контролируются состоянием
активационных и инактивационных ворот, зависящим от на мембране.
• В покоящейся клетке потенциал зависимый канал находится обычно в
закрытом состоянии, из которого он может быть открыт или активирован.
• Вероятность его самостоятельного открытия невысока, в состоянии покоя в
мембране открыто небольшое количество этих каналов.
• Деполяризация мембраны (уменьшение ) активирует канал, повышая
вероятность его открытия. Для открытия канала необходимо достижение
критического уровня деполяризации.
• Функцию активационных ворот выполняет электрически заряженная
аминокислота (лизин или аргинин, расположенные в каждой третьей
позиции на данном участке молекулы), закрывающая вход в устье канала.
• Лизин и аргинин, как полярные аминокислоты, являются сенсором на
мембране; при достижении критического уровня деполяризации мембраны
заряженная часть молекулы сенсора смещается в сторону липидного
микроокружения каналообразующей молекулы и ворота открывают вход в
устье канала (см. следующий слайд) .
24.
Потенциал-зависимый канал можетнаходиться в трех последовательных
состояниях:
2
1) закрытый;
2) открытый;
3) инактивированный
Состояние канала зависит
1
3
Время, мс
от величины потенциала мембраны
Переход 1-2-3 осуществляется скачком,
Потенциал-зависимый канал
Переход 3-1 за «длительный» период
времени
25.
Скорость открытия активационных ворот может бытьочень высокой
и низкой.
По этому показателю потенциалзависимые ионные каналы делят на
быстрые (например, быстрые потенциалзависимые Na+ каналы) и
медленные (например, медленные потенциалзависимые Ca2+ каналы).
Скорость срабатывания канала зависит от подвижности
активационных ворот.
Быстрые каналы открываются мгновенно (мкс) и остаются
открытыми в среднем в течение 1 мс. Их активация сопровождается
быстрым лавинообразным возрастанием проницаемости канала для
определенных ионов – пропускная способность открытого канала
поразительна: ток ионов происходит со скоростью до 100 000 000
ионов/с.
Медленные каналы открываются вместе с быстрыми, но скорость
движения активационных ворот в них значительно меньше, поэтому и
движение ионов через них начинается позднее.
26.
Активационные ворота – последовательность аминокислот в виде плотного шарика(клубка) на нити, располагающегося у выхода устья канала. При изменении знака
заряда на мембране с (+) на (–) шарик закрывает выход из устья, канал становится
непроницаемым (закрытым) для иона, но канал может быть активирован пороговым
или большим стимулом.
Инактивационные ворота – также последовательность аминокислот. Инактивация
сопровождается прекращением перемещения ионов через канал и может протекать
так же быстро, как активация, или медленно — в течение секунд и даже минут. Все
это время канал не может быть открыть ни при какой силе возбуждения, пока не
перейдет закрытое состояние.
27.
Существуют вещества, блокирующие работу потенциалзависимыхионных каналов. Таким образом, возможно еще одно состояние
ионного канала – блокированное. Такие вещества получили название
блокаторов ионных каналов.
Одним из первых блокаторов было описано вещество тетродотоксин
(образуется в организме рыбы фугу), блокатор потенциалзависимых
натриевых каналов. В эксперименте при его введении в организм
животных отмечались потеря чувствительности, расслабление
мышц, обездвиженность, остановка дыхания и смерть.
В клинике используют лидокаин, новокаин, прокаин — вещества,
при введении которых в организм в небольших дозах развивается
блокада потенциалзависимых натриевых каналов нервных волокон и
блокируется передача в ЦНС сигналов от болевых рецепторов, это
местные обезболивающие средства.
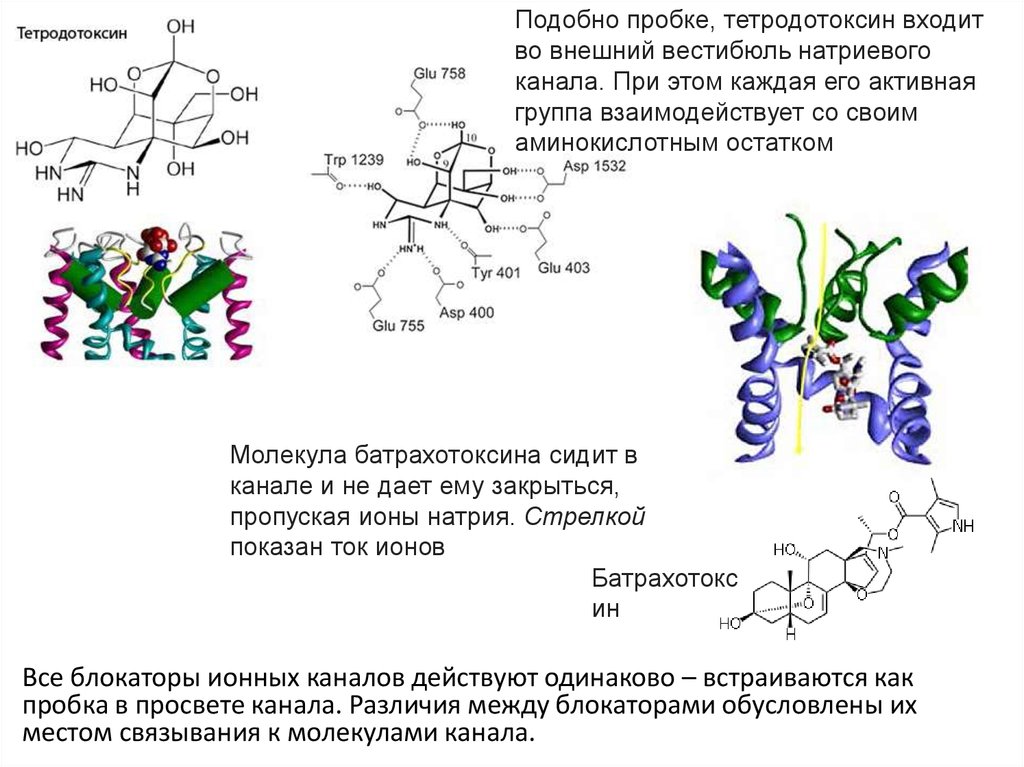
28.
Подобно пробке, тетродотоксин входитво внешний вестибюль натриевого
канала. При этом каждая его активная
группа взаимодействует со своим
аминокислотным остатком
Молекула батрахотоксина сидит в
канале и не дает ему закрыться,
пропуская ионы натрия. Стрелкой
показан ток ионов
Батрахотокс
ин
Все блокаторы ионных каналов действуют одинаково – встраиваются как
пробка в просвете канала. Различия между блокаторами обусловлены их
местом связывания к молекулами канала.
29.
Порообразующая часть ионного канала может представлять собой1) единый полипептид, организованный в виде нескольких идентичных
трансмембранных доменов,
2) нескольких белковых субъединиц, которые могут быть как одинаковы
(гомоолигомер), так и неодинаковы (гетероолигомер) по своей структуре.
Каждая порообразующая субъединица или домен образована несколькими
трансмембранными α-спиральными сегментами с N- и С-терминальными
доменами белка, направленными внутриклеточно или внеклеточно.
Один из трансмембранных сегментов субъединицы потенциал-активируемых
каналов содержит уникальный набор положительных зарядов и
функционирует как сенсор потенциала (Armstrong, Hille, 1998, Hille, 2001).
Практически все каналы в составе образующих пору канала субъединиц
имеют регуляторные домены, связывающиеся с различными регуляторными
молекулами.
Каналы обладают свойством селективно пропускать ионы, которое
реализуется в самом узком месте канала – селективном фильтре.
30.
Потенциал зависимый каналСтроение Na+-ионного канала клеточной мембраны:
а – двухмерное строение α-субъединицы ионного канала клеточной мембраны;
б – слева – натриевый канал, состоящий из 4 α -субъединиц и двух -субъединиц
(вид сбоку); справа – вид натриевого канала сверху.
Цифрами I. II. III. IV отмечены домены α -субъединицы
31.
Нарушение работы ионных каналов часто приводят кзаболеваниям — каналопатиям. Основная причина
таких нарушений — наследственные мутации,
влияющие на структуру канала.
Примеры каналопатий:
- муковисцидоз
- сердечные аритмии
- синдром Бругада (натриевый канал кардиомиоцитов)
- синдром Тимоти (аномалии кальциевых каналов)
- генерализованная эпилепсия
32.
Работа ионного канала(разным цветом выделены разные белки, образующие домены)
пропускная способность открытого канала поразительна:
ток ионов происходит со скоростью до 100 000 000 ионов/с
33.
Лигандзависимые ионные каналыФормируются белковыми макромолекулами, одновременно
выполняющими функцию
ионных каналов
и селективных рецепторов лигандов.
Поэтому за ними закрепились разные синонимические названия,
например: синаптический рецептор, лигандзависимый канал.
В отличие от потенциалзависимых ионных каналов, открытие
которых осуществляется в ответ на уменьшение ,
лигандзависимые ионные каналы открываются (активируются)
при взаимодействии участка узнавания лиганда белка рецептора
молекулы с лигандом — веществом, к которому рецепторный
участок узнавания имеет высокое сродство.
34.
2 , , , белковые субъединицы,образующие систему распознавания ионов
Гидрофильная полость
Центр связывания
лиганда (трансмиттера)
Клеточная мембрана
Гидрофобная поверхность
Лиганд-зависимый канал
35.
14 нанометровАминоконцевой
домен
12
нанометров
Лигандсвязывающий
домен
Антагонист
рецептора
Трансмембранный домен
Молекулярная (белковая) структура лигандзависимого ионного канала
(разным цветом выделены разные белки, образующие домены)
36.
Лигандзависимые ионные каналы обычно локализованы впостсинаптических мембранах нервных клеток и их отростков, а
также мышечных волокон.
Типичными примерами лигандзависимых ионных каналов являются
каналы пре- и постсинаптических мембран, активируемые
ацетилхолином, глутаматом, аспартатом, -аминомасляной
кислотой, катехоламинами, глицином и др. синаптическими
нейромедиаторами.
Обычно название канала (рецептора) отражает тип нейромедиатора,
который в естественных условиях является его лигандом. Так, если
это каналы нервно-мышечного синапса, в котором используется
нейромедиатор ацетилхолин, то используется термин
«ацетилхолиновый рецептор», если он является к тому же
чувствительным к никотину, то его называют
никотинчувствительным, или просто н-ацетилхолиновым,
рецептором (н-холинорецептор, N-АЦХ).
37.
Лигандзависимые каналы могут изменять проницаемость длякатионов Na+ и К+ или для К+ и анионов СI–. Такая
избирательность связывания лиганда и изменения ионной
проницаемости генетически закреплена в пространственной
структуре макромолекулы.
Если взаимодействие медиатора и рецепторной части
макромолекулы, формирующей ионный канал, непосредственно
изменяет проницаемость канала, то в течение нескольких
миллисекунд это меняет проницаемости постсинаптической
мембраны для минеральных ионов и величину постсинаптического
потенциала, например, ацетилхолиновые и глутаматные каналы.
Такие каналы названы быстрыми и локализуются, например, в
постсинаптической мембране аксо-дендритных возбуждающих
синапсов и аксосоматических тормозных синапсов.
38.
Существуют медленные лигандзависимые ионные каналы.Их открытие реализуется через цепь последовательных событий:
первичный медиатор (лиганд, нейромедиатор) активирует G-белок →
он взаимодействует с GTP → который запускает синтез вторичных
медиаторов (посредников) во внутриклеточной передаче сигнала →
которые фосфорилируют ионный канал → это меняет его
проницаемость для минеральных ионов → и, как следствие, величину
постсинаптической .
Эта цепь событий осуществляется уже за сотни миллисекунд.
С такими медленными лигандзависимыми ионными каналами мы
встретимся при изучении механизмов регуляции работы сердца,
гладких мышц.
39.
АцетилхолинК+-канал
Синоатриальный
узел миокарда
Гиперполяризация
Связь ионного канала с рецептором
Ацетилхолин возбуждает m2-рецептор, это через систему доменных белков
приводит к открытию К+-канала синоатриального узла, входу К+ в клетку,
гиперполяризации ее мембраны и снижением ее возбудимости.
40.
Особым типом лигандзависимых каналов являются каналы, локализованные вмембранах эндоплазматического ретикулума гладкомышечной клетки.
Их лигандом является вторичный посредник внутриклеточной передачи сигнала
инозитолтрифосфат – ИФ3.
Описаны ионные каналы, характеризующиеся некоторыми структурными и
функциональными свойствами, присущими как потенциалзависимым, так и
лигандзависимым ионным каналам. Они являются потенциалНЕчувствительными
ионными каналами, состояние воротного механизма которых контролируется
циклическими нуклеотидами (цАМФ и цГМФ).
Циклические нуклеотиды связываются с внутриклеточной СООН-терминалью
каналообразующей молекулы белка и активируют канал. Эти каналы менее
избирательны для катионов и регулируют проницаемость для других ионов.
Так, Са2+, поступая через активированные каналы из внеклеточной среды, блокируют
проницаемость каналов для Na+.
Такие каналы встречаются в палочках сетчатки глаза, их проницаемость для Са2+ и
Na+ определяется уровнем цГМФ.
Лигандзависимые ионные каналы широко представлены в мембранных структурах,
обеспечивающих синаптическую передачу сигналов от ряда сенсорных рецепторов в
ЦНС; передачу сигналов в синапсах нервной системы и от нейронов к эффекторным
клеткам.
41. Рецепторы
РЕЦЕПЦИЯ – процесс восприятия и трансформации(преобразования) энергии внешнего раздражителя в
нервный импульс или сложную последовательность
внутриклеточных процессов, посредством
мембранных и/или цитоплазматических процессов.
Функцию рецепции выполняют специальные
образования – рецепторы.
42.
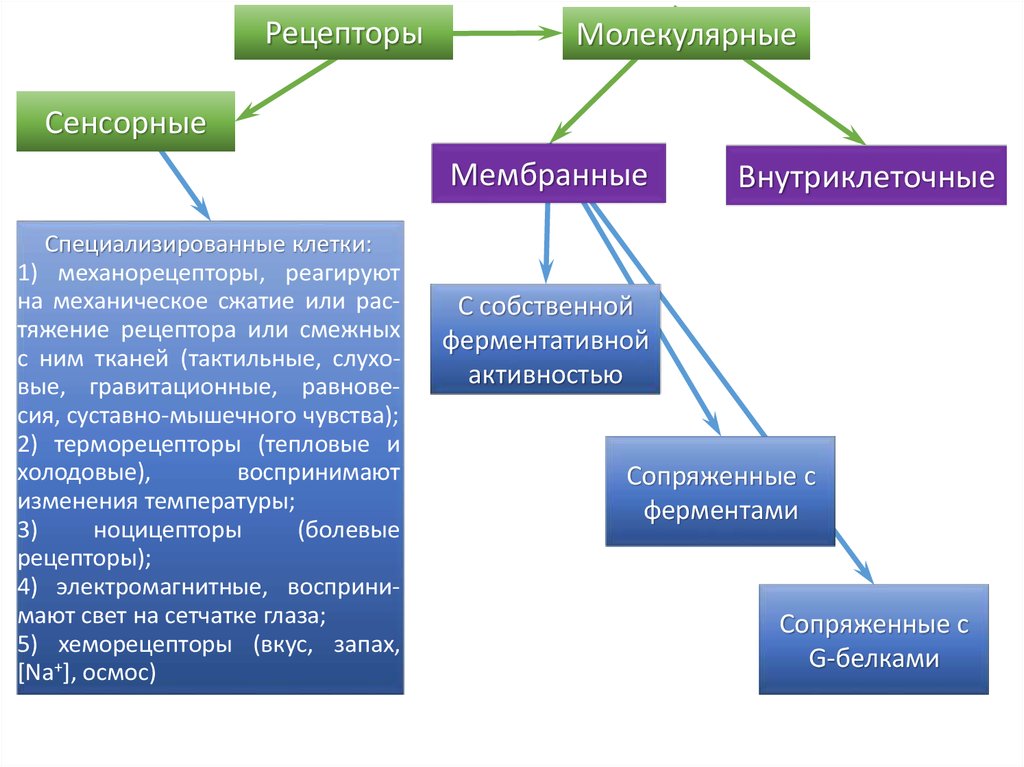
РецепторыМолекулярные
Сенсорные
Мембранные
Специализированные клетки:
1) механорецепторы, реагируют
на механическое сжатие или растяжение рецептора или смежных
с ним тканей (тактильные, слуховые, гравитационные, равновесия, суставно-мышечного чувства);
2) терморецепторы (тепловые и
холодовые),
воспринимают
изменения температуры;
3)
ноцицепторы
(болевые
рецепторы);
4) электромагнитные, воспринимают свет на сетчатке глаза;
5) хеморецепторы (вкус, запах,
[Na+], осмос)
Внутриклеточные
С собственной
ферментативной
активностью
Сопряженные с
ферментами
Сопряженные с
G-белками
43.
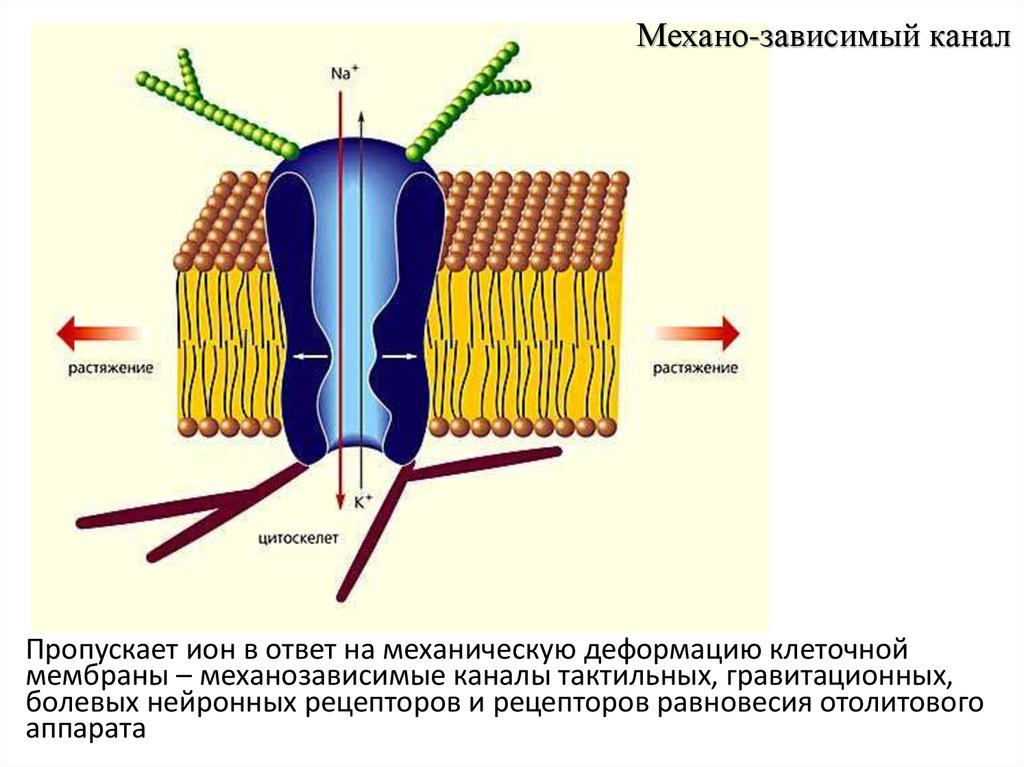
Механо-зависимый каналПропускает ион в ответ на механическую деформацию клеточной
мембраны – механозависимые каналы тактильных, гравитационных,
болевых нейронных рецепторов и рецепторов равновесия отолитового
аппарата
44. Сенсорные рецепторы
45.
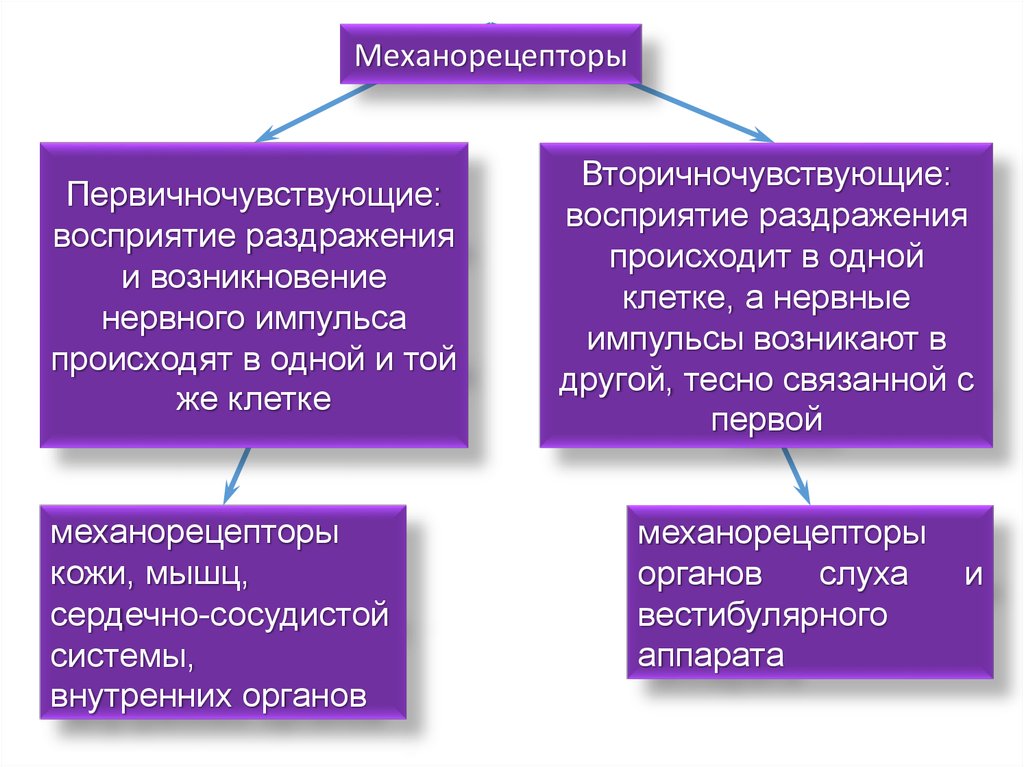
МеханорецепторыПервичночувствующие:
восприятие раздражения
и возникновение
нервного импульса
происходят в одной и той
же клетке
механорецепторы
кожи, мышц,
сердечно-сосудистой
системы,
внутренних органов
Вторичночувствующие:
восприятие раздражения
происходит в одной
клетке, а нервные
импульсы возникают в
другой, тесно связанной с
первой
механорецепторы
органов
слуха
вестибулярного
аппарата
и
46.
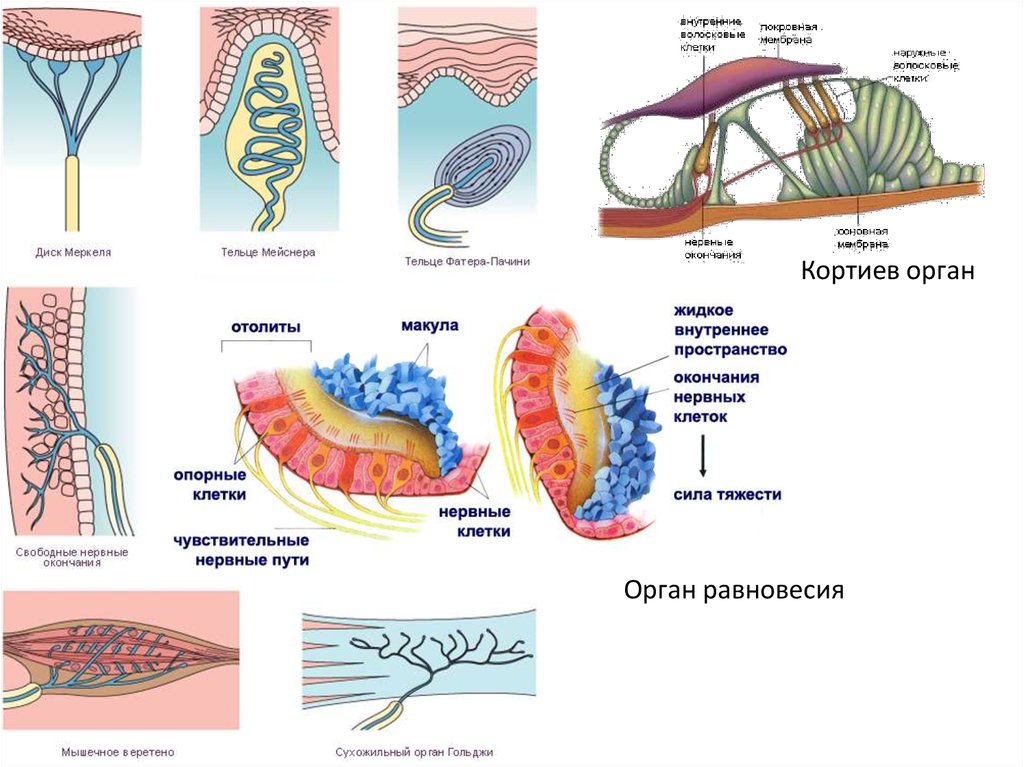
Кортиев органОрган равновесия
47.
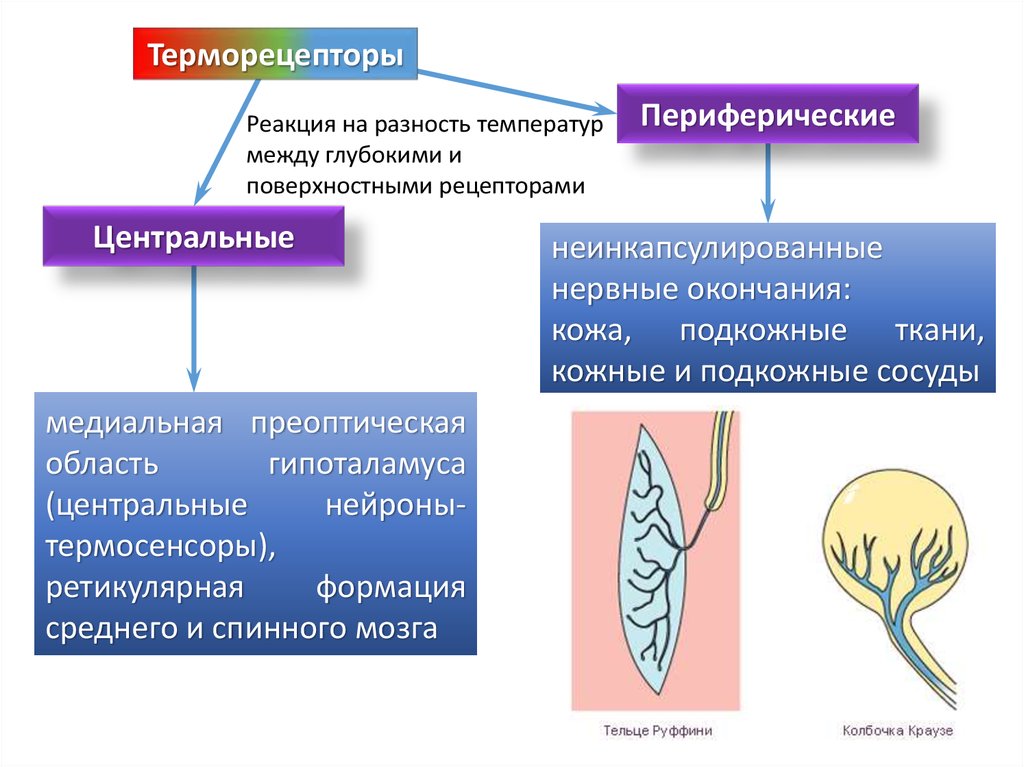
ТерморецепторыРеакция на разность температур
между глубокими и
поверхностными рецепторами
Центральные
медиальная преоптическая
область
гипоталамуса
(центральные
нейронытермосенсоры),
ретикулярная
формация
среднего и спинного мозга
Периферические
неинкапсулированные
нервные окончания:
кожа, подкожные ткани,
кожные и подкожные сосуды
48.
Реакция на превышение порога раздражения. 1 тип:механо- и теплове рецепторы; 2тип - хеморецепторы,
Ноцицепторы
Свободные нервные окончания безмиелиновые (медленные нервные волокна)
и миелиновые (быстрые нервные волокна)
49.
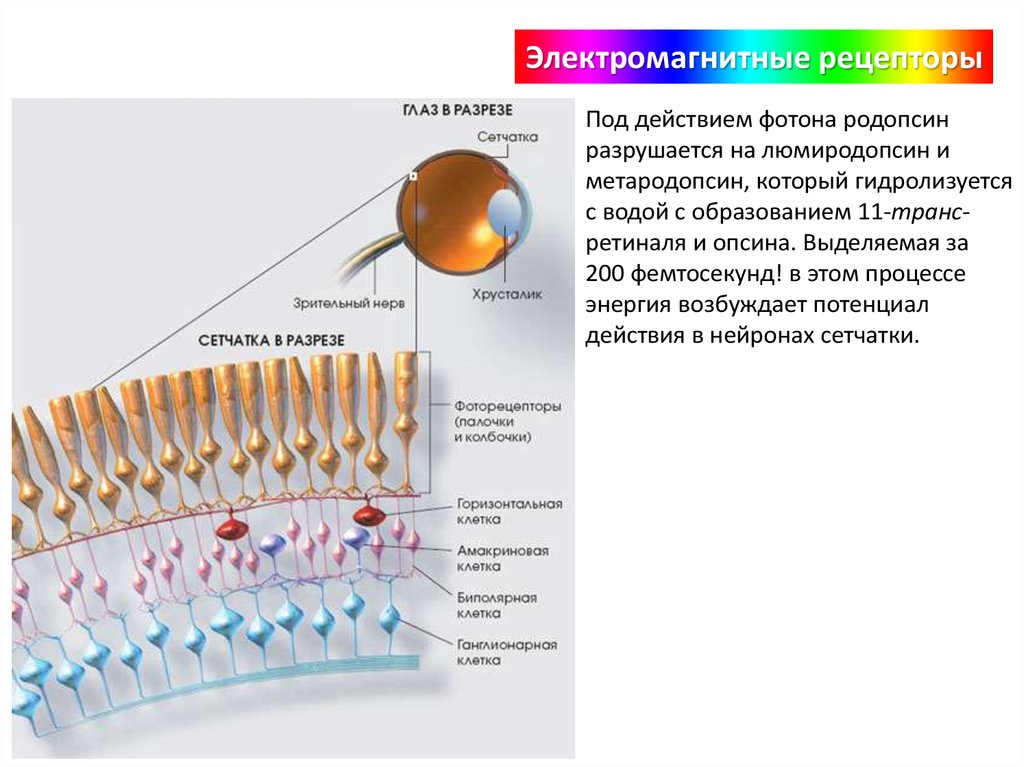
Электромагнитные рецепторыПод действием фотона родопсин
разрушается на люмиродопсин и
метародопсин, который гидролизуется
с водой с образованием 11-трансретиналя и опсина. Выделяемая за
200 фемтосекунд! в этом процессе
энергия возбуждает потенциал
действия в нейронах сетчатки.
50. 3D организация сетчатки
51.
Центральные (медуллярные)хеморецепторы расположены
непосредственно в ростральных
отделах вентральной дыхательной
группы, в структурах голубого пятна
(locus coeruleus), ретикулярных ядрах
шва ствола мозга, реагируют на
водородные ионы в окружающей их
межклеточной жидкости мозга
Хеморецепторы
Периферические (артериальные) хеморецепторы расположены в
каротидных тельцах в области бифуркации общих сонных артерий и в
аортальных тельцах в области дуги аорты, реагируют как на изменение
концентрации водородных ионов, так и парциального давления кислорода в
артериальной крови.
Обонятельные (запахи – опасные, нейтральные, приятные, феромоны).
Вкусовые (горькое, кислое, сладкое, соленое, умами).
Натриевые (расположены в плотной субстанции нефронов).
52.
ОсморецепторыРасположены в гипоталамусе и печени.
Реагируют на изменение осмотического давления плазмы
крови. Между осморецепторами печени и гипоталамуса, а
также нефронами почек существуют прямые
рефлекторные связи.
53. Молекулярные рецепторы
54.
55.
Структура рецепторова) ионотропного и б) связанного с G-белком
56.
Мембранные рецепторы сопряженные с ионнымиканалами
Механизм работы:
Активация рецептора
Открытие ионного канала
Изменение электрического потенциала клеточной
мембраны
Изменение функционального состояния клетки
57.
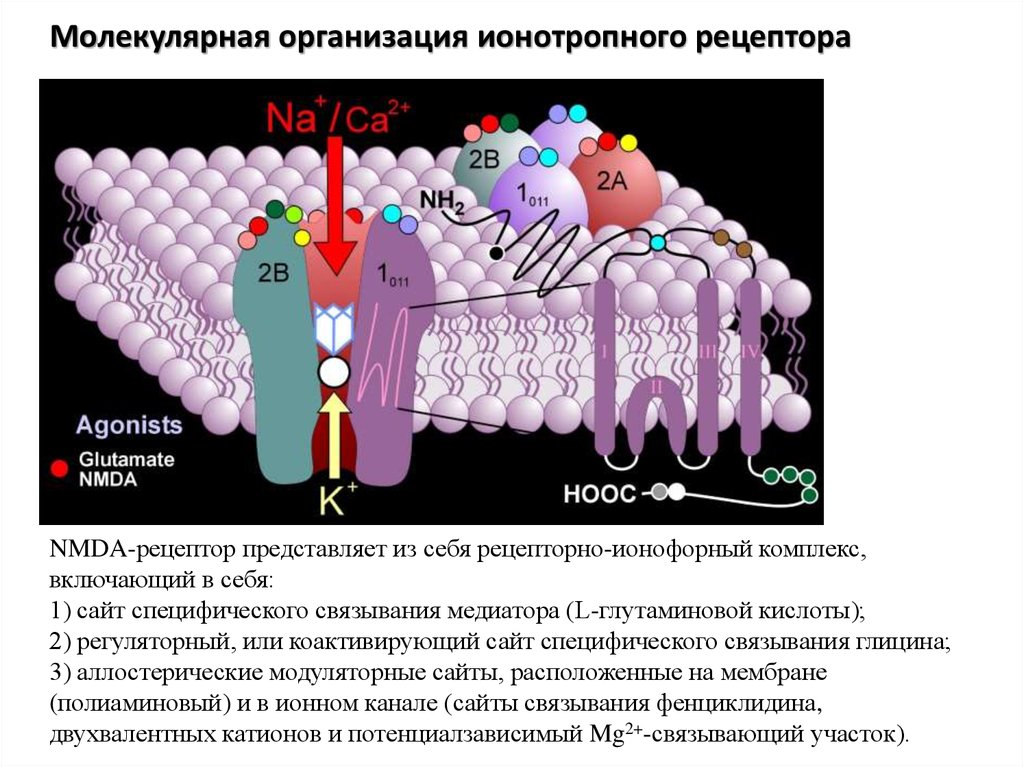
Молекулярная организация ионотропного рецептораNMDA-рецептор представляет из себя рецепторно-ионофорный комплекс,
включающий в себя:
1) сайт специфического связывания медиатора (L-глутаминовой кислоты);
2) регуляторный, или коактивирующий сайт специфического связывания глицина;
3) аллостерические модуляторные сайты, расположенные на мембране
(полиаминовый) и в ионном канале (сайты связывания фенциклидина,
двухвалентных катионов и потенциалзависимый Mg2+-связывающий участок).
58.
Мембранные рецепторы сопряженные с G-белкамиМеханизм работы:
Активация рецептора
Активация G–белка
Активация фермента, катализирующего образование
второго посредника
Образование второго посредника
Активация протеинкиназы
Фосфорелирование эффекторного белка
Изменение функционального состояния клетки
59.
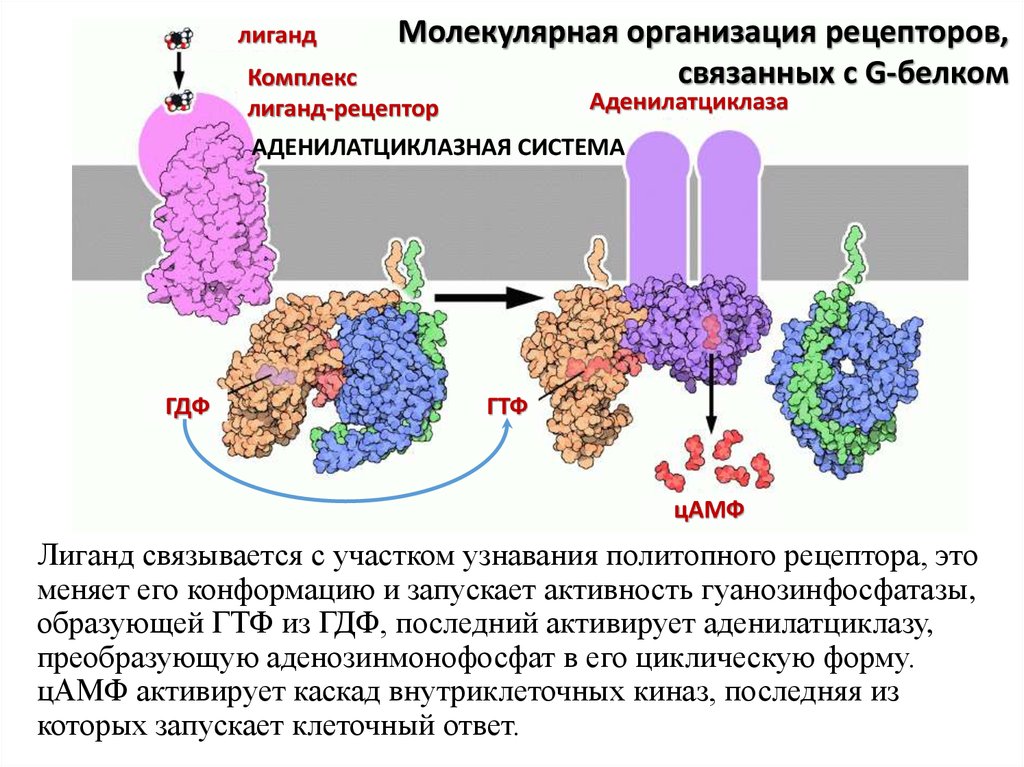
лигандМолекулярная организация рецепторов,
связанных с G-белком
Комплекс
лиганд-рецептор
Аденилатциклаза
АДЕНИЛАТЦИКЛАЗНАЯ СИСТЕМА
ГДФ
ГТФ
цАМФ
Лиганд связывается с участком узнавания политопного рецептора, это
меняет его конформацию и запускает активность гуанозинфосфатазы,
образующей ГТФ из ГДФ, последний активирует аденилатциклазу,
преобразующую аденозинмонофосфат в его циклическую форму.
цАМФ активирует каскад внутриклеточных киназ, последняя из
которых запускает клеточный ответ.
60.
Молекулярная организация рецепторов,связанных с G-белком
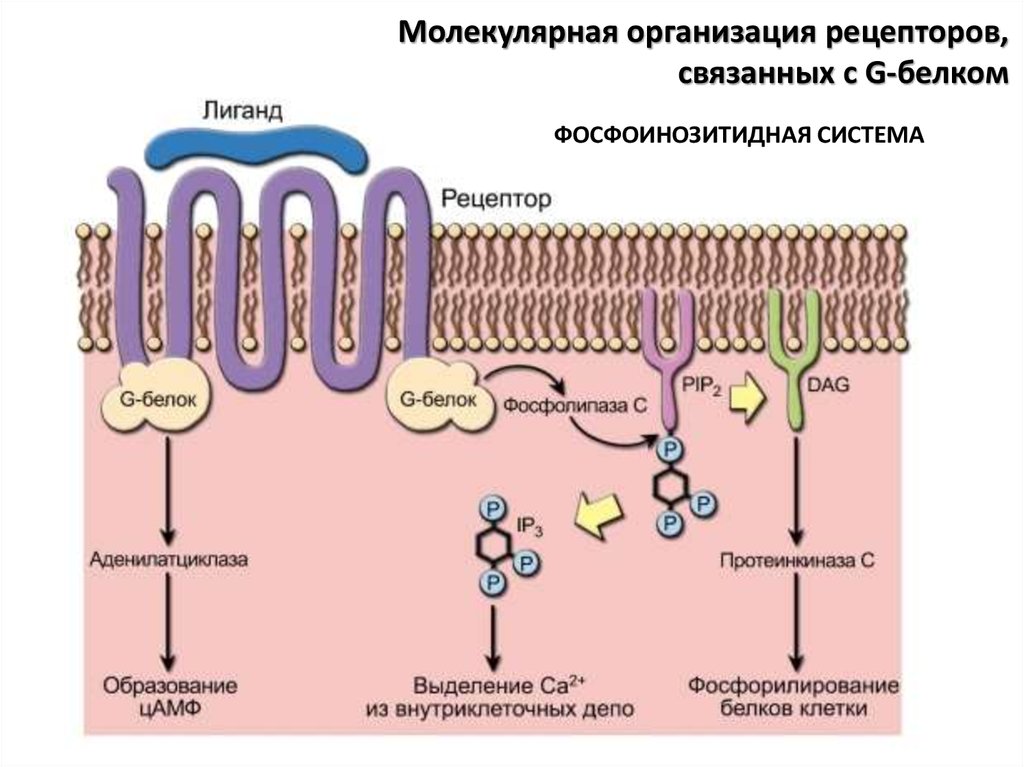
ФОСФОИНОЗИТИДНАЯ СИСТЕМА
61.
G-белки двух типов:GS – стимулирующие клеточный ответ
GI – ингибирующие клеточный ответ
Лиганды (первичные посредники) – гидрофильные амины,
аминоподобные вещества, пептиды:
ацетилхолин, гистамин, катехоламины (дофамин, норадреналин,
адреналин), серотонин, гормоны гипофиза
Вторичные посредники (мессенджеры):
циклические нуклеотиды (цАМФ, цГМФ – гуанилатциклазная
система),
диацилглицерол (ДАГ)
инозитолдифосфат (ИТФ2)
инозитолтрифосфат (ИТФ3)
62.
63.
Молекулярная организация монотопных рецепторовс собственной ферментативной активностью
Лиганды:
факторы роста
нервов,
тромбоцитов,
инсулин и др.,
64.
Внутриклеточные (ядерные) рецепторы –факторы транскрипции
Только гидрофобные молекулы!
65.
Лиганды ядерных рецепторов (жирорастворимыевещества):
дийодтирозин, трийодтирозин, ретиноиды,
стероидные гормоны (холекальциферолы,
глюкокортикоиды, минералокортикоиды,
дигидротестостерон, прогестины, эстрогены)