




 Химия
ХимияПохожие презентации:










Структура гетероциклов, нуклеозидов и нуклеотидов
1.
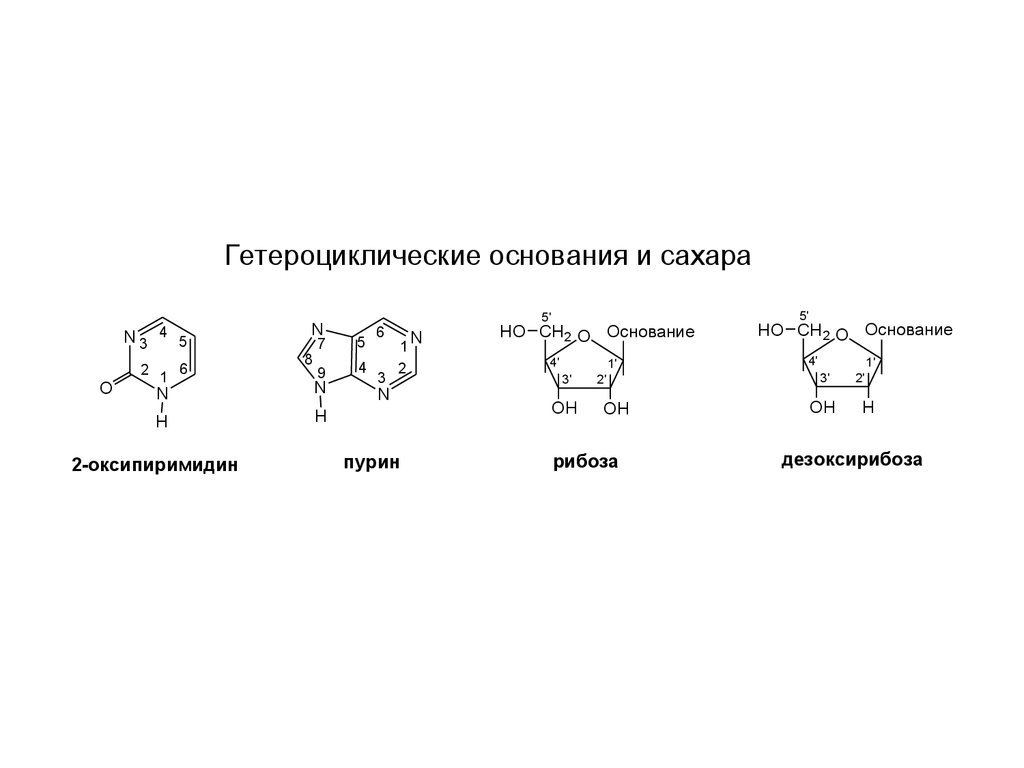
Гетероциклические основания и сахараN3
2
O
4
1
5
6
N
8
7
5
9
4
N
N
H
H
2-оксипиримидин
6
3
5'
1
2
N
пурин
N
HO CH2 O Основание
4'
1'
3'
OH
2'
OH
рибоза
5'
HO CH2 O Основание
4'
1'
3'
OH
2'
H
дезоксирибоза
2.
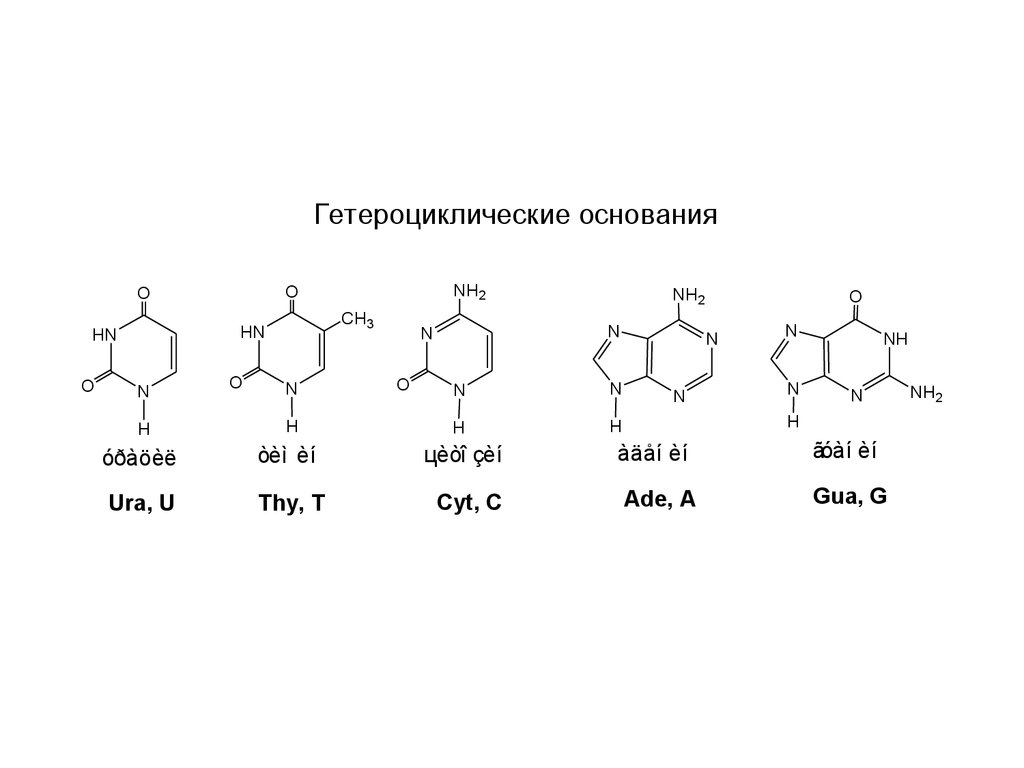
Гетероциклические основанияO
CH3
HN
HN
N
NH2
O
O
O
N
N
N
O
NH2
N
N
H
H
H
H
óðàöèë
òèì èí
цèòî çèí
Ura, U
Thy, T
Cyt, C
O
N
N
N
N
NH
N
H
àäåí èí
ãóàí èí
Ade, A
Gua, G
NH2
3.
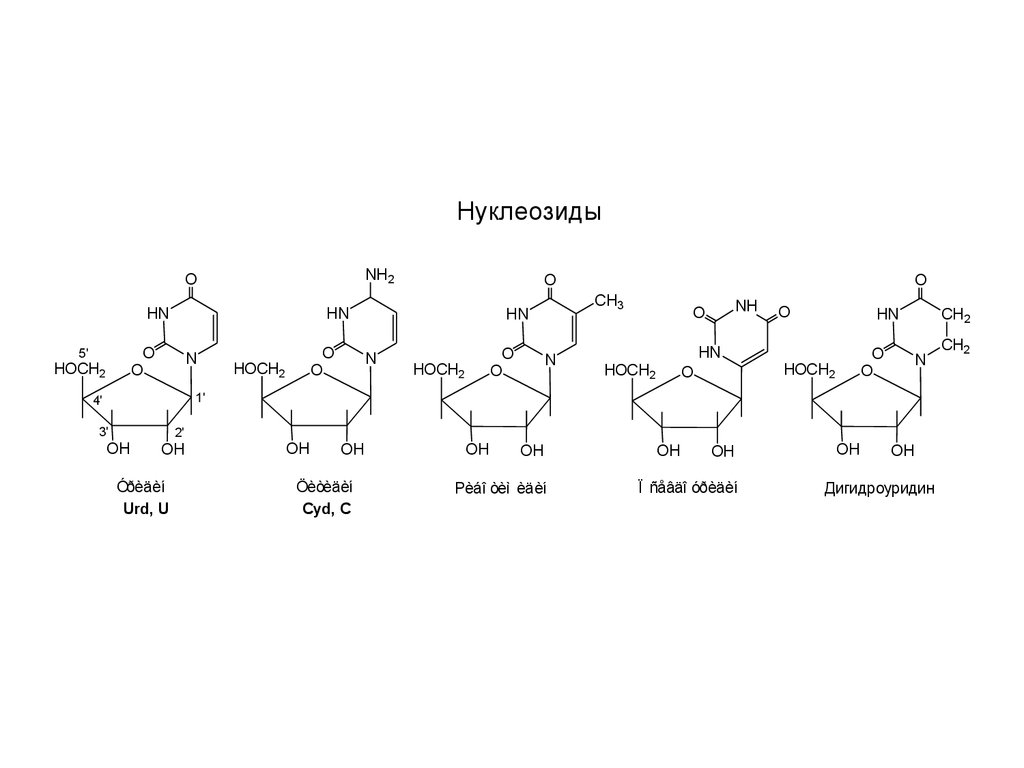
НуклеозидыNH2
O
HN
HN
O
O
5'
HOCH2
O
N
O
O
HOCH2
HN
N
O
O
HOCH2
O
CH3
N
NH
O
HN
HOCH2
O
O
CH2
HN
O
O
HOCH2
N
1'
4'
3'
2'
OH
OH
Óðèäèí
Urd, U
OH
OH
Öèòèäèí
Cyd, C
OH
OH
Рèáî òèì èäèí
OH
OH
Ï ñåâäî óðèäèí
OH
OH
Дигидроуридин
CH2
4.
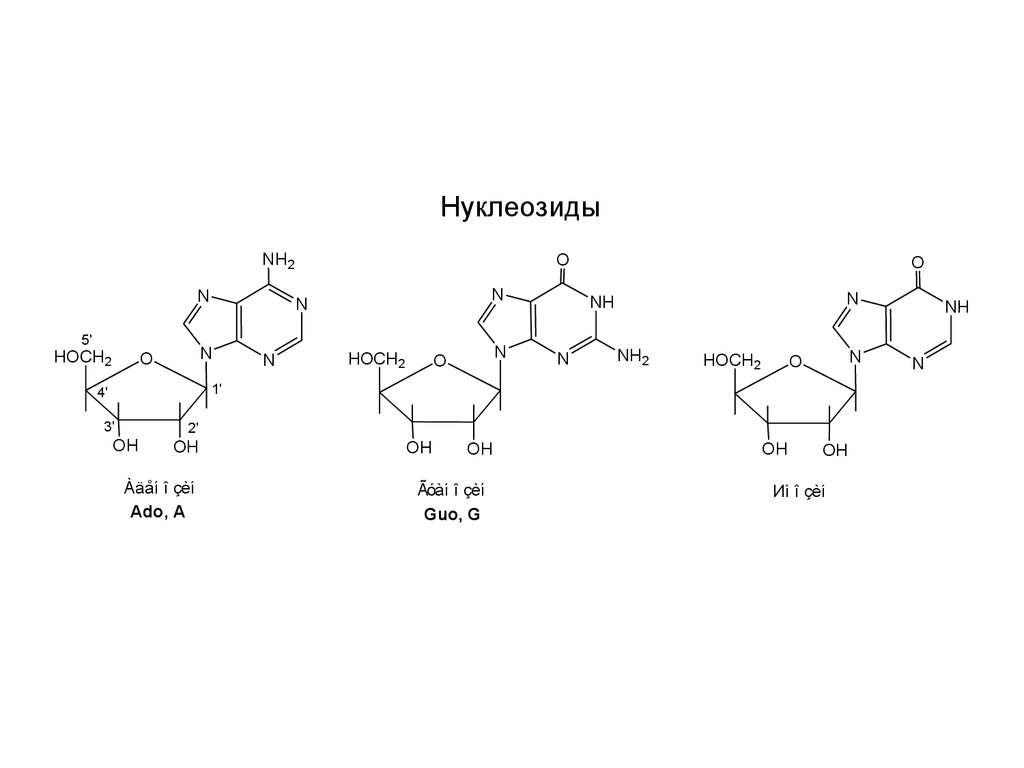
НуклеозидыNH2
N
5'
HOCH2
N
N
N
O
O
N
HOCH2
N
O
O
N
NH
N
NH2
HOCH2
N
O
1'
4'
3'
2'
OH
OH
Àäåí î çèí
Ado, A
OH
OH
Ãóàí î çèí
Guo, G
OH
OH
Иí î çèí
NH
N
5.
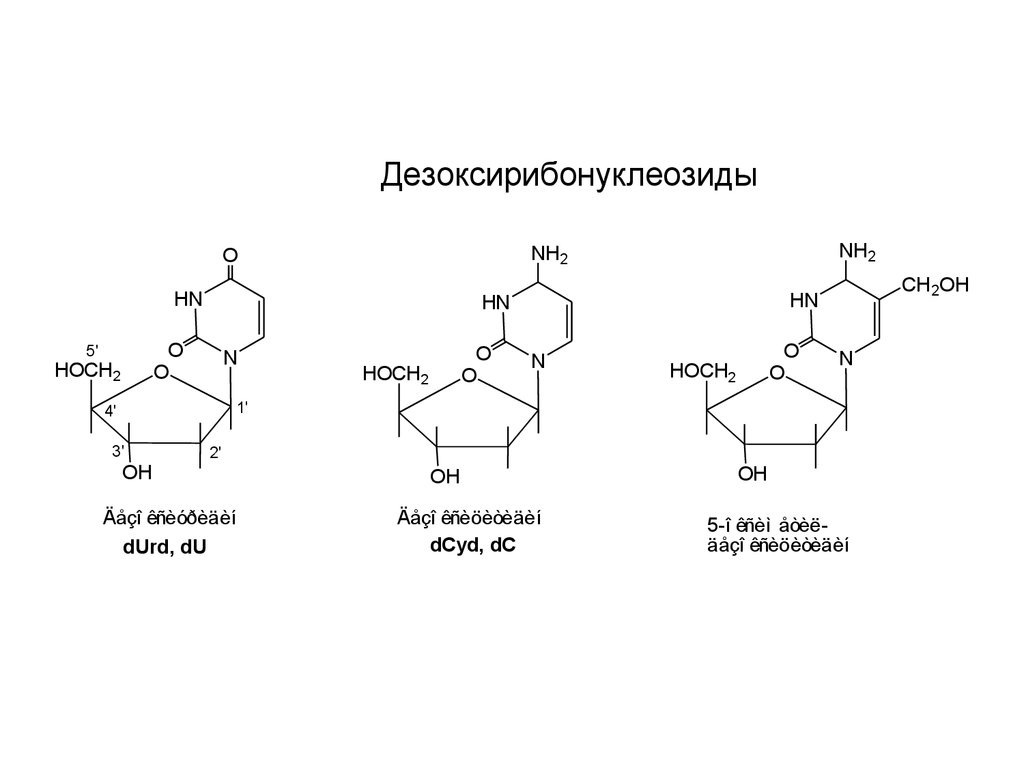
ДезоксирибонуклеозидыHN
HOCH2
N
O
O
HOCH2
CH2OH
HN
HN
O
O
5'
NH2
NH2
O
N
O
O
HOCH2
N
1'
4'
3'
2'
OH
Äåçî êñèóðèäèí
dUrd, dU
OH
Äåçî êñèöèòèäèí
dCyd, dC
OH
5-î êñèì åòèëäåçî êñèöèòèäèí
6.
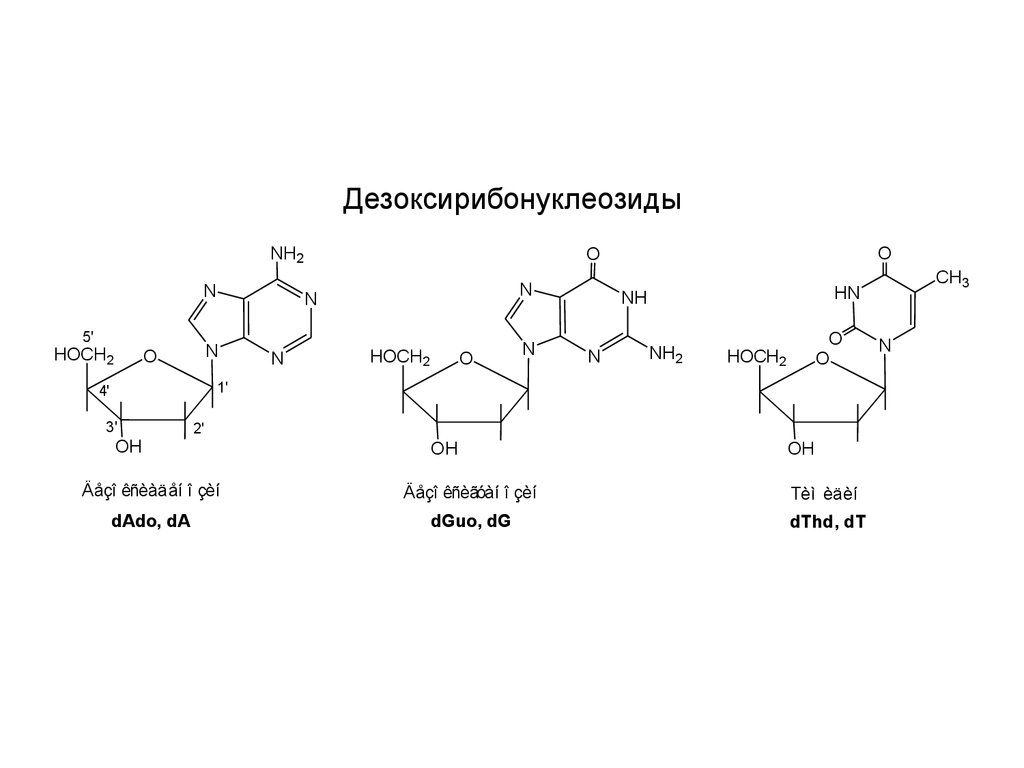
ДезоксирибонуклеозидыNH2
N
5'
HOCH2
N
N
N
O
N
O
O
HOCH2
O
N
NH
N
NH2
O
O
HOCH2
1'
4'
3'
2'
OH
OH
CH3
HN
OH
Äåçî êñèàäåí î çèí
Äåçî êñèãóàí î çèí
Тèì èäèí
dAdo, dA
dGuo, dG
dThd, dT
N
7.
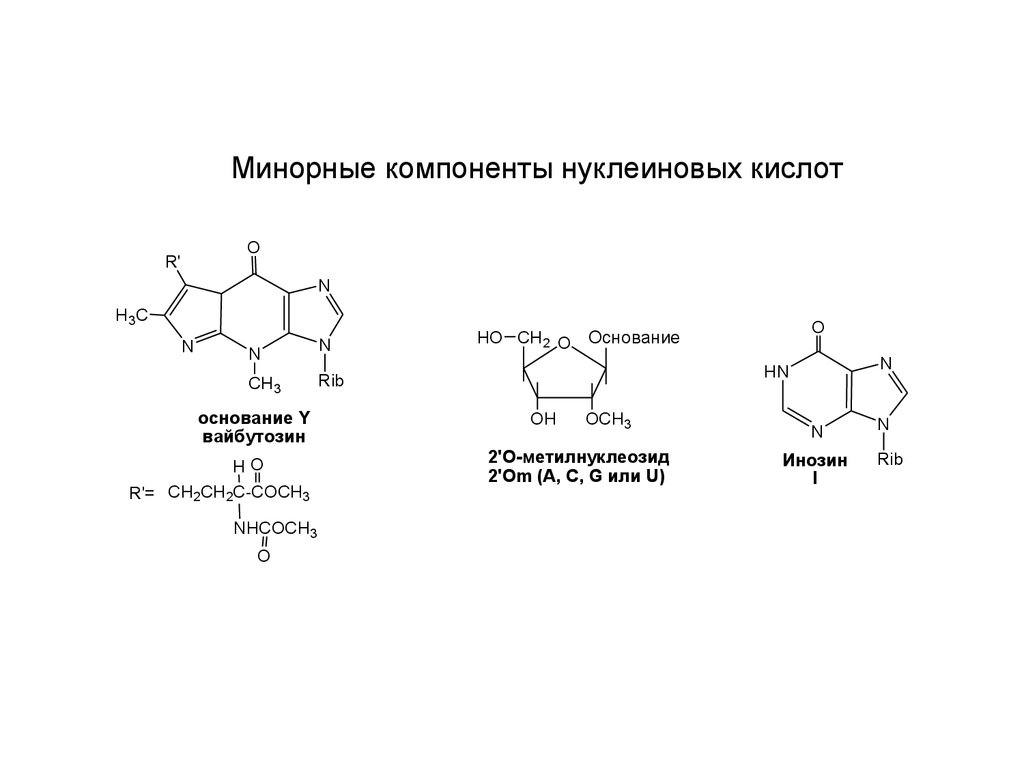
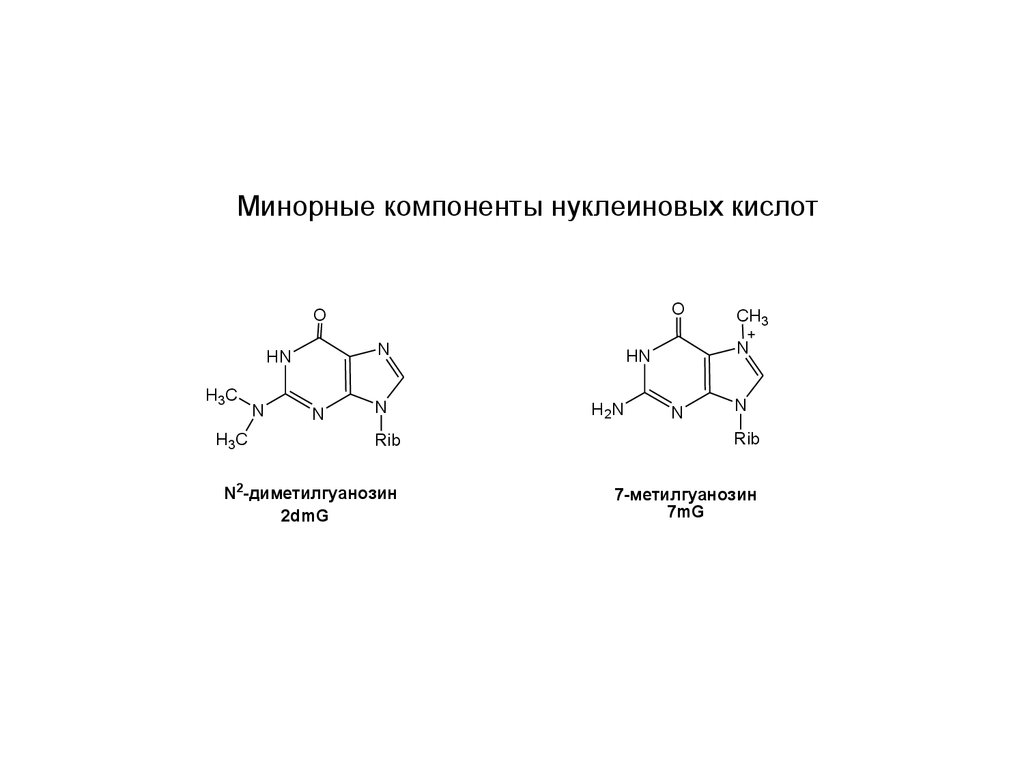
Минорные компоненты нуклеиновых кислотO
R'
N
H 3C
N
N
CH3
основание Y
вайбутозин
HO
R'= CH2CH2C-COCH3
NHCOCH3
O
N
O
HO CH2 O Основание
N
HN
Rib
OH
OCH3
2'O-метилнуклеозид
2'Оm (A, C, G или U)
N
Инозин
I
N
Rib
8.
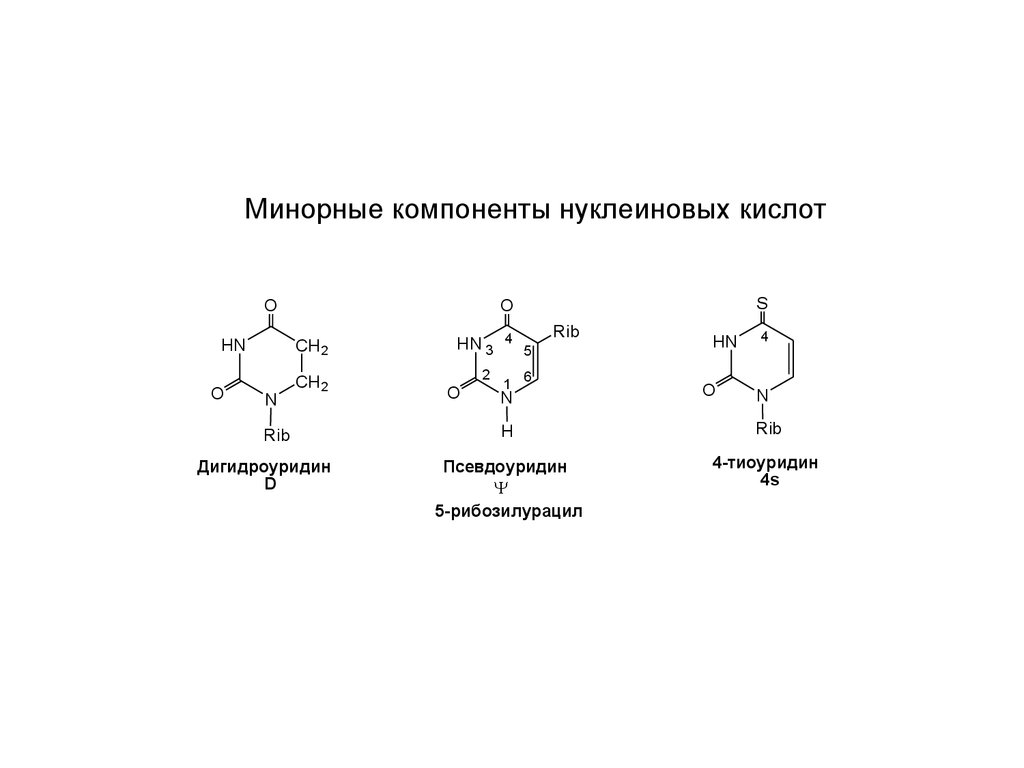
Минорные компоненты нуклеиновых кислотHN
O
CH2
N
S
O
O
CH2
Rib
Дигидроуридин
D
HN 3 4 5
2
O
1
Rib
6
N
HN
O
4
N
H
Rib
Псевдоуридин
5-рибозилурацил
4-тиоуридин
4s
9.
Минорные компоненты нуклеиновых кислотO
O
N
HN
H3C
N
N
N
N
HN
H2 N
CH3
N
N
Rib
Rib
N2-диметилгуанозин
2dmG
7-метилгуанозин
7mG
H3C
10.
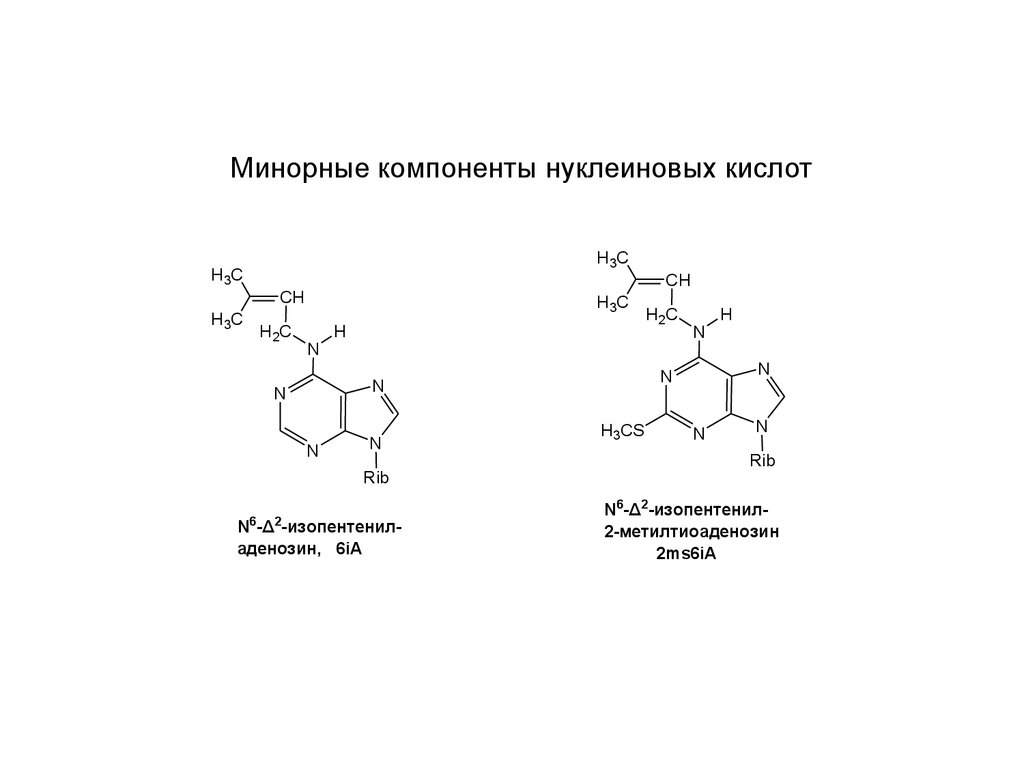
Минорные компоненты нуклеиновых кислотH3C
H 3C
CH
H 3C
H2C
H3C
N
H
N
N
Rib
6
2
N -Δ -изопентениладенозин, 6iA
H2C
N
H3CS
H
N
N
N
N
CH
N
N
Rib
N6-Δ2-изопентенил2-метилтиоаденозин
2ms6iA
11.
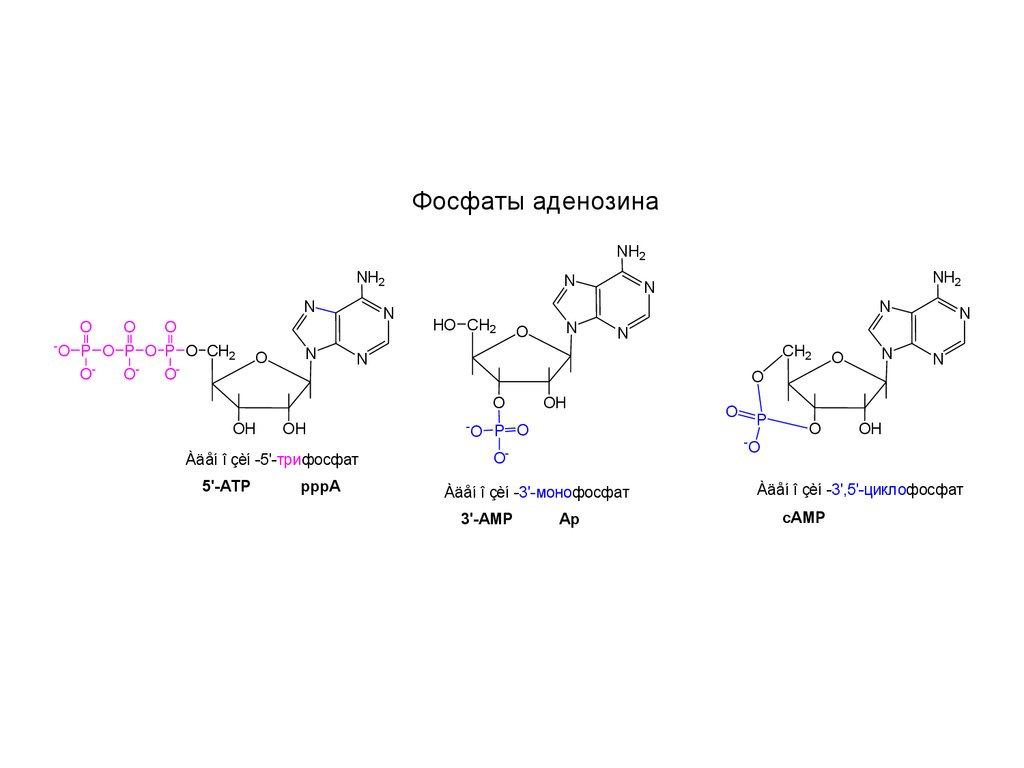
Фосфаты аденозинаNH2
NH2
N
O
-
O
N
O
O P O P O P O CH2
OOO-
O
N
N
N
HO CH2
O
OH
Àäåí î çèí -5'-трифосфат
5'-ATP
pppA
N
CH2
O
N
O
-
OH
O P O
O
P
-
OÀäåí î çèí -3'-монофосфат
3'-AMP
Ap
N
N
N
O
OH
NH2
N
O
N
OH
O
Àäåí î çèí -3',5'-циклофосфат
сAMP
12.
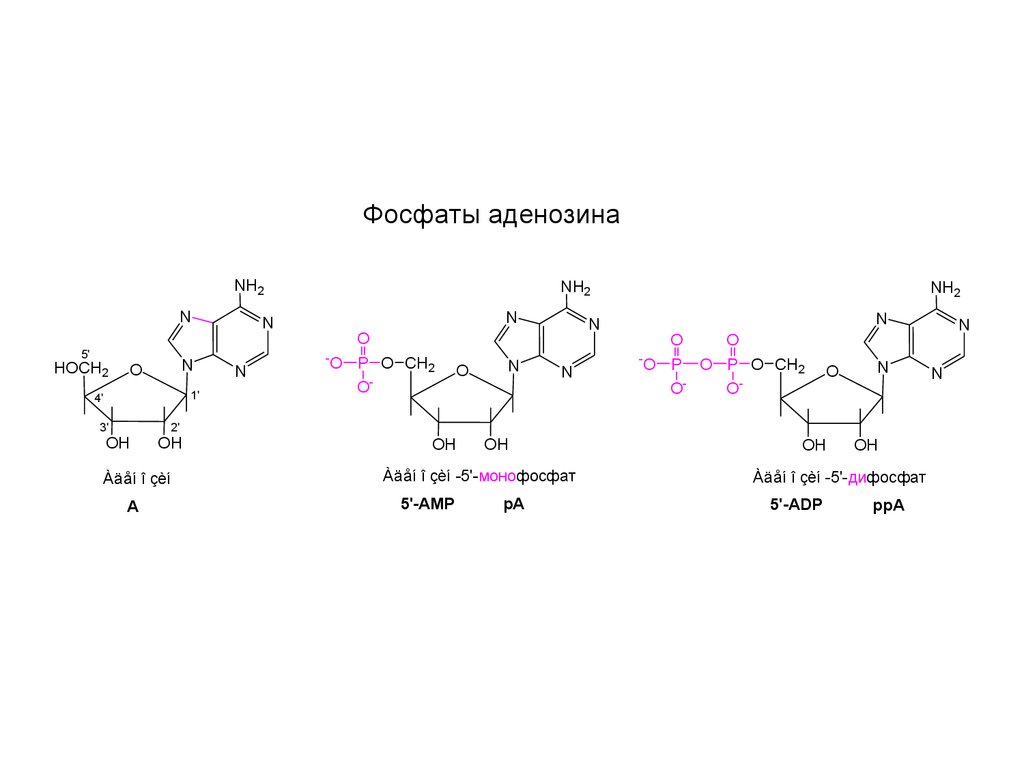
Фосфаты аденозинаNH2
N
5'
HOCH2
N
O
1'
4'
NH2
N
N
N
O P O CH2
O-
O
N
N
N
O
-
NH2
N
O
-
O P O P O CH2
OO-
O
N
2'
3'
OH
OH
Àäåí î çèí
A
OH
OH
Àäåí î çèí -5'-монофосфат
5'-AMP
pA
N
O
OH
OH
Àäåí î çèí -5'-дифосфат
5'-ADP
ppA
N
13.
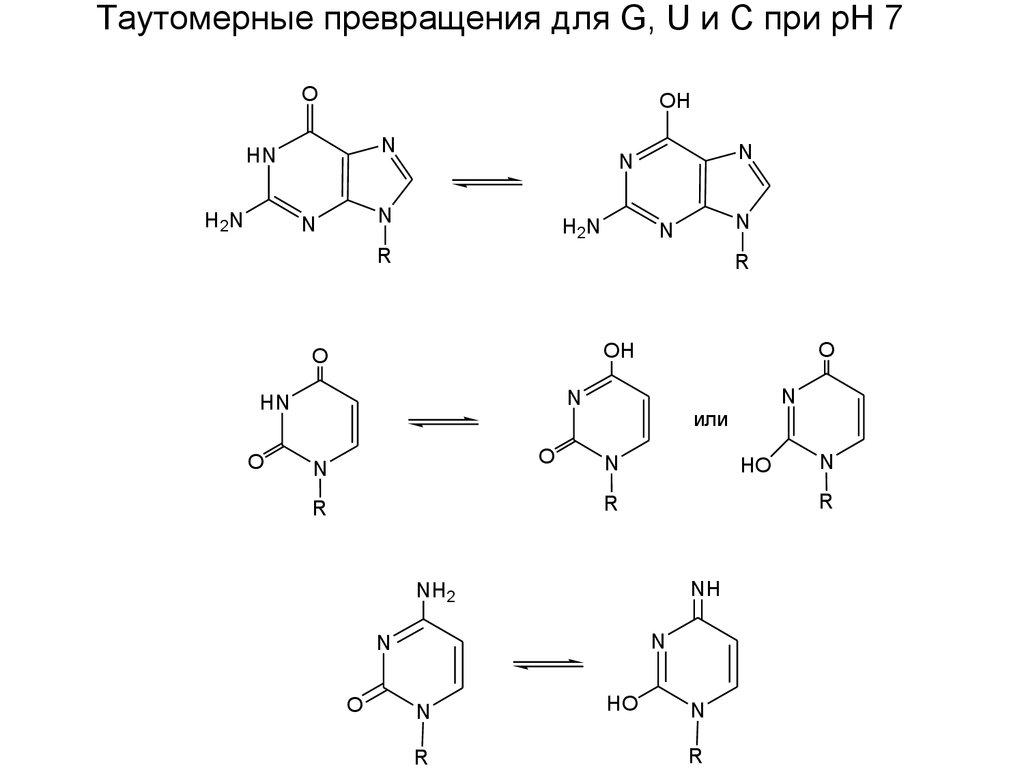
Таутомерные превращения для G, U и С при рН 7O
OH
N
HN
H2N
N
N
N
N
H2N
N
N
R
R
N
HN
O
O
OH
O
O
N
или
N
HO
NH
NH2
N
N
O
N
R
HO
N
R
R
R
N
N
R
14.
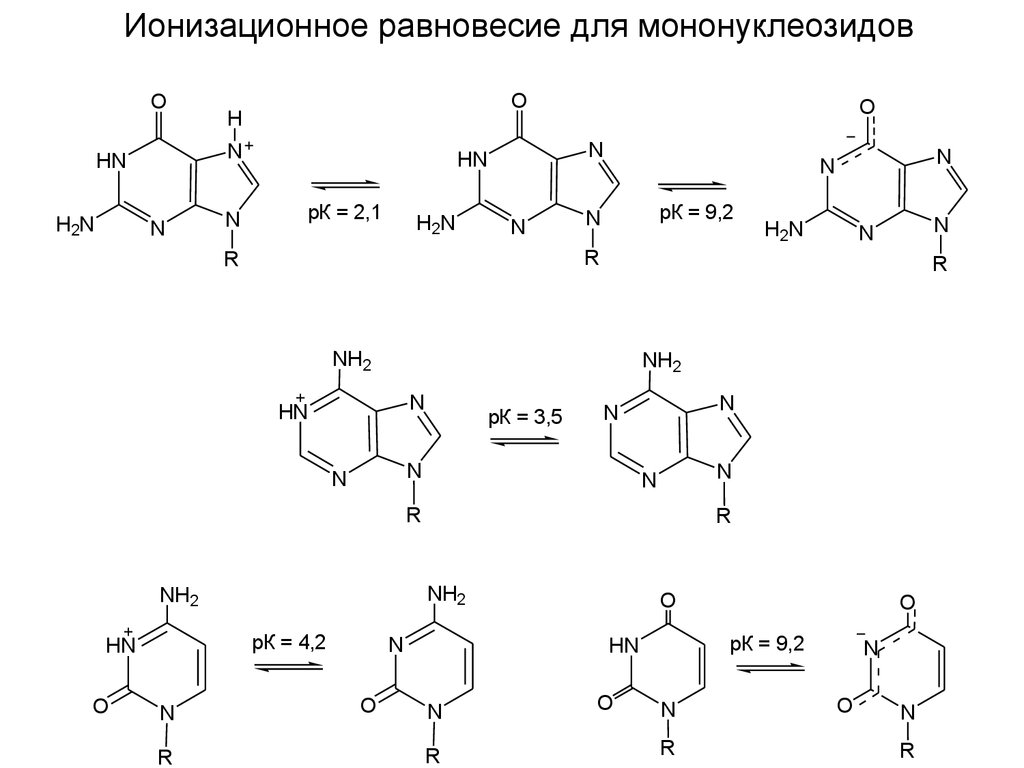
Ионизационное равновесие для мононуклеозидовO
H
N
HN
H2N
O
N
N
N
HN
рК = 2,1
H2N
O
N
рК = 9,2
N
NH2
N
HN
рК = 3,5
N
N
N
N
N
N
R
рК = 4,2
R
R
NH2
NH2
N
N
N
R
NH2
O
H2N
R
R
HN
N
N
N
O
O
рК = 9,2
HN
N
R
O
O
N
R
N
O
N
R
15.
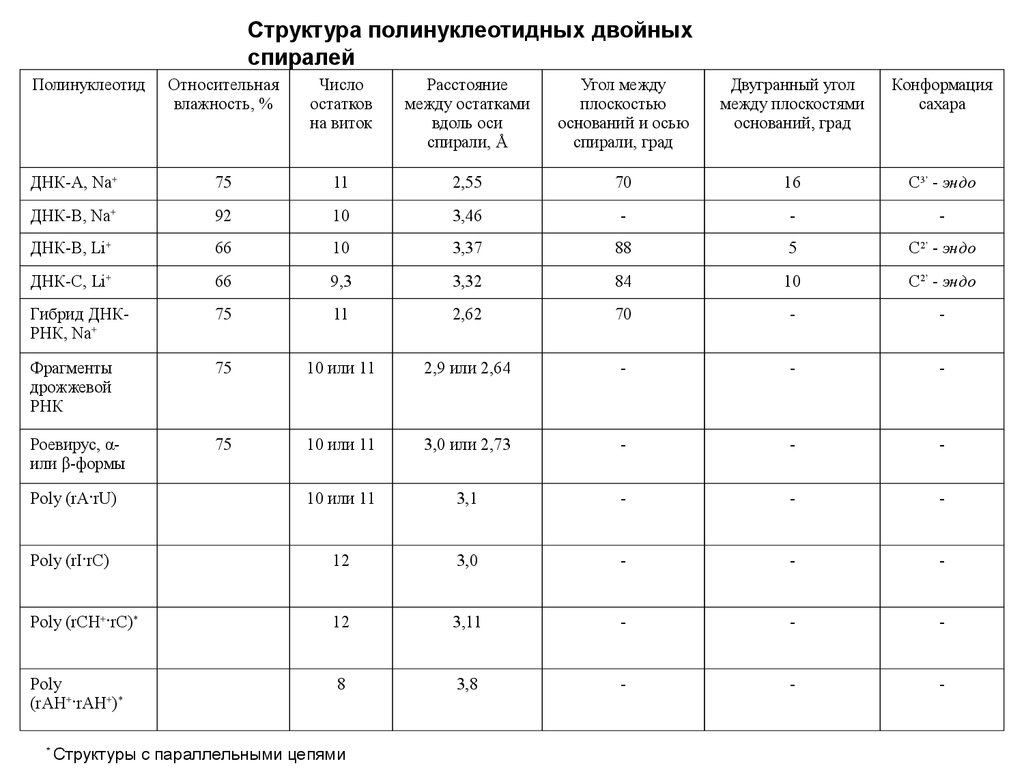
Структура полинуклеотидных двойныхспиралей
Полинуклеотид
Относительная
влажность, %
Число
остатков
на виток
Расстояние
между остатками
вдоль оси
спирали, Å
Угол между
плоскостью
оснований и осью
спирали, град
Двугранный угол
между плоскостями
оснований, град
Конформация
сахара
ДНК-А, Na+
75
11
2,55
70
16
С3’ - эндо
ДНК-B, Na+
92
10
3,46
-
-
-
ДНК-B, Li+
66
10
3,37
88
5
С2’ - эндо
ДНК-C, Li+
66
9,3
3,32
84
10
С2’ - эндо
Гибрид ДНКРНК, Na+
75
11
2,62
70
-
-
Фрагменты
дрожжевой
РНК
75
10 или 11
2,9 или 2,64
-
-
-
Роевирус, αили β-формы
75
10 или 11
3,0 или 2,73
-
-
-
Poly (rA·rU)
10 или 11
3,1
-
-
-
Poly (rI·rC)
12
3,0
-
-
-
Poly (rCH+·rC)*
12
3,11
-
-
-
Poly
(rAH+·rAH+)*
8
3,8
-
-
-
*
Структуры с параллельными цепями
16.
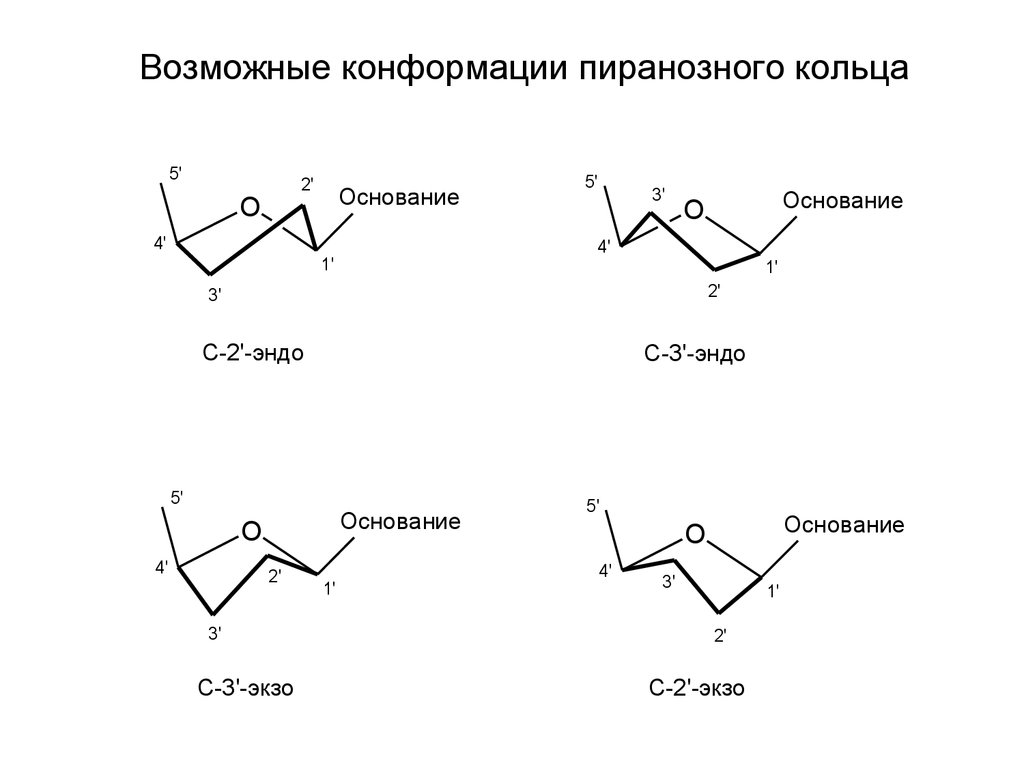
Возможные конформации пиранозного кольца5'
2'
O
Основание
4'
5'
3'
4'
1'
1'
2'
3'
C-2'-эндо
C-3'-эндо
5'
Основание
O
4'
Основание
O
2'
3'
C-3'-экзо
1'
5'
Основание
O
4'
3'
1'
2'
C-2'-экзо
17.
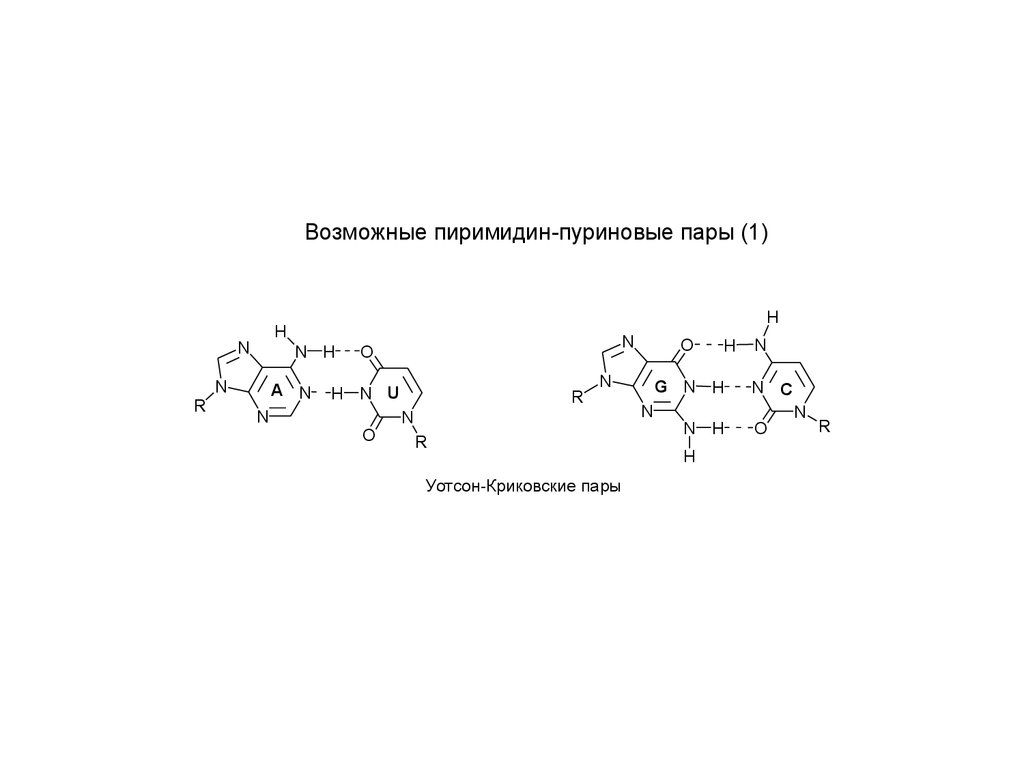
Возможные пиримидин-пуриновые пары (1)N
R
H
H
N H
N
A N
N
N
O
H N U
O
R
O
N
N
R
Уотсон-Криковские пары
H
G N H
N
N H
H
N
N C
O
N
R
18.
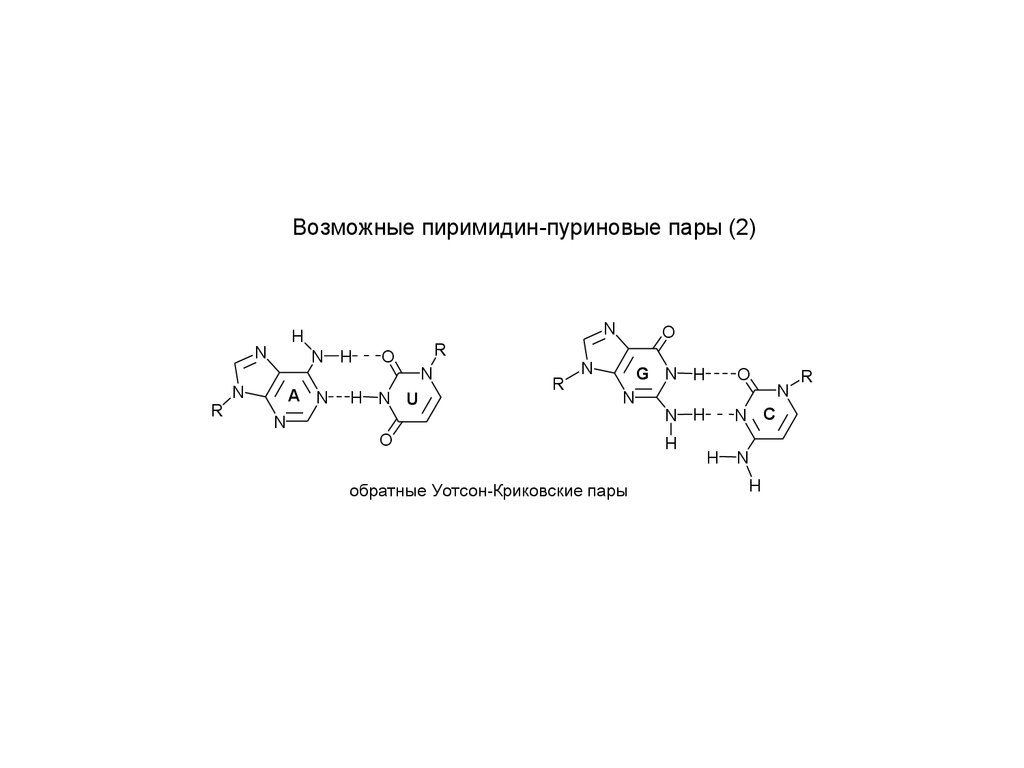
Возможные пиримидин-пуриновые пары (2)N
N H
N
R
N
H
A N
N
O
H N U
R
N
R
O
N
G N H
N
O
обратные Уотсон-Криковские пары
O
N C
N H
H
H
N
H
N
R
19.
Возможные пиримидин-пуриновые пары (3)H
N
N H
A
N
N
O
H
N
U
N
R
H
O
N
R
N
A.U Хугстеновская пара
A
N
.
N H
N
N
R
N
O
H
N
U
O
R
A U обратная Хугстеновская пара
O
H
N
A
N
N
N
.
N
N
R
N C
H
H
N
H
R
A С обратная Хугстеновская пара
20.
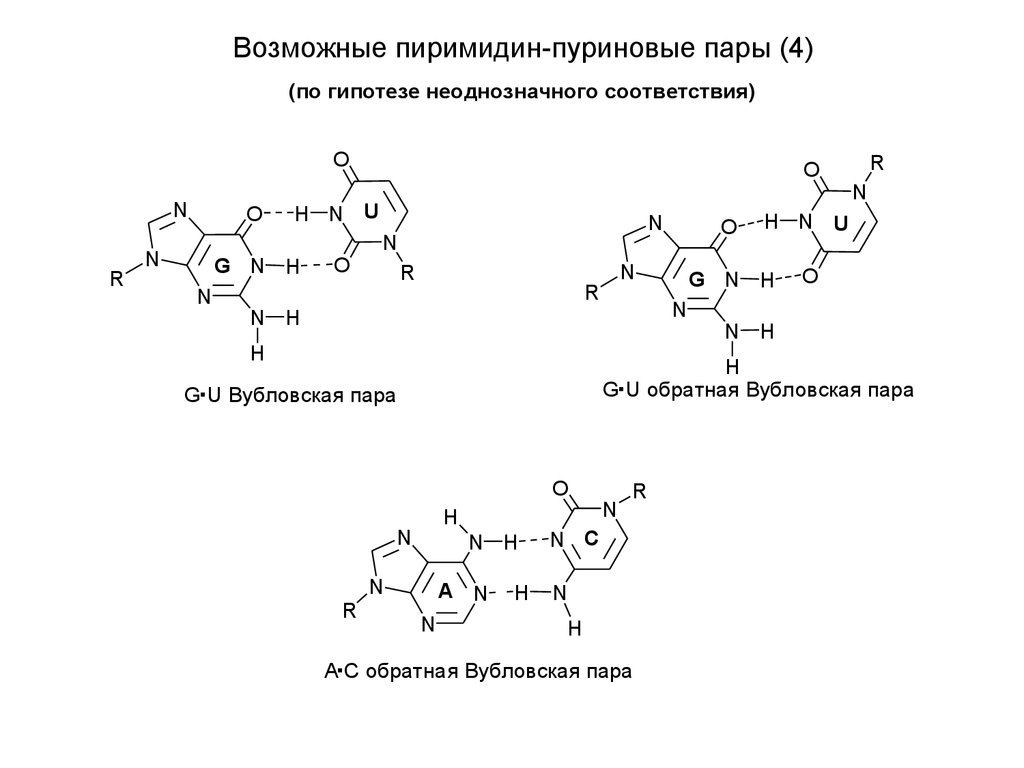
Возможные пиримидин-пуриновые пары (4)(по гипотезе неоднозначного соответствия)
O
N
R
O
N
H N U
G N H
N
.
O
O
N
N
R
N
R
O
H
N
.
O
N H
H
.
G U обратная Вубловская пара
G U Вубловская пара
R
N
H N U
G N H
N
N H
H
O
R
N H
N
A N
N
H
N
R
N C
N
H
A С обратная Вубловская пара
21.
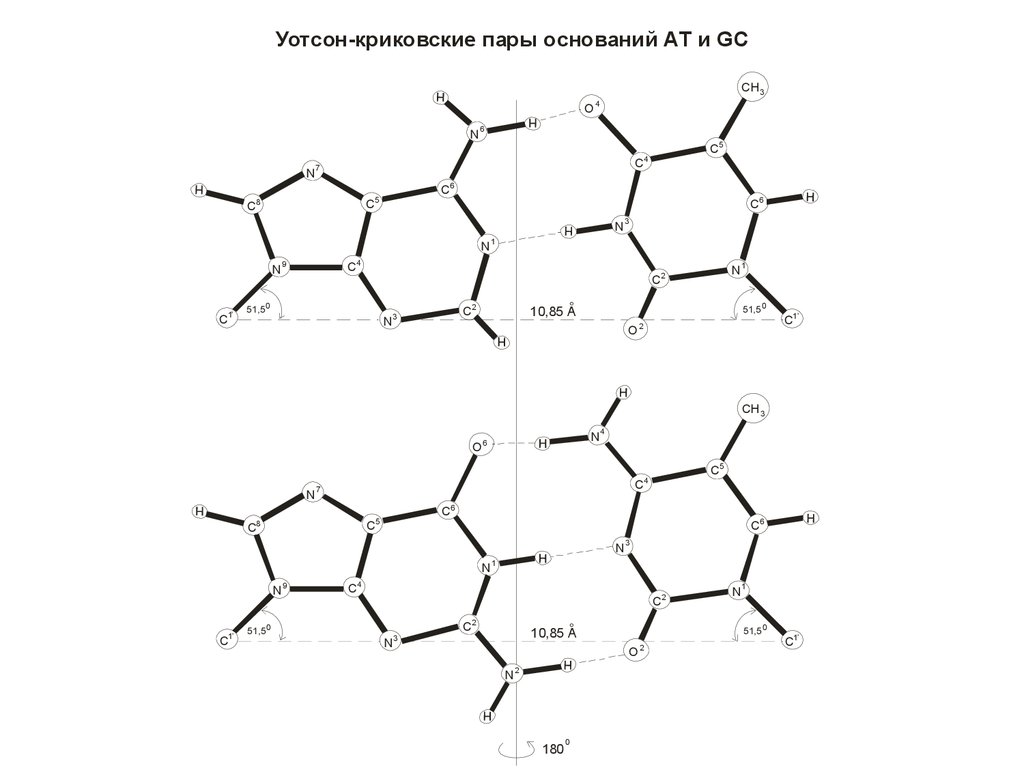
Уотсон-криковские пары оснований АТ и GCCH 3
О4
N
N
6
C
7
C6
C5
C8
C6
N
N
N
C1’
3
1
C4
9
C5
4
N
C2
51,5 0
N
°
10,85 А
C2
3
1
51,5 0
О
2
C1’
CH 3
N
О6
N
C
4
7
C
8
C6
5
C6
N
N
N
1’
C
9
C5
C4
3
1
C4
C2
C2
51,5 0
N
°
10,85 А
3
2
180
0
1
51,5 0
О2
N
N
C1’
22.
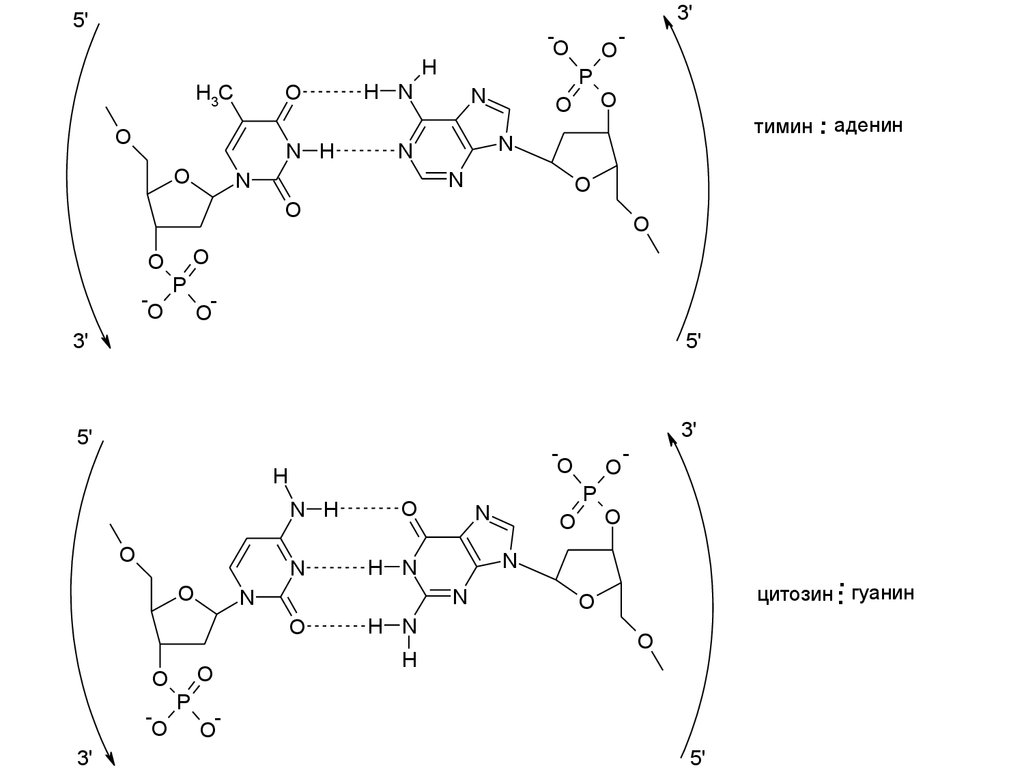
3'5'
O
H
H3C
O
O
N H
O
H N
-
P
N
O
O
тимин
N
N
N
N
O
O
O
O
O
O
O
P
O
3'
5'
5'
3'
O
H
N H
O
N
O
O
O
3'
P
O
H
-
O
O
N
N
H N
O
P
N
H N
N
O
O
.. аденин
...
цитозин гуанин
O
O
O
5'
23. Структура G·С пары в составе Z-ДНК
HH
O
N
HC
N
C
C
HC
H 2C
3'
O CH
HC
O
5'
CH2
O
C
N
H
N
C
N
N
H
C
N
H
C
C
O
C H
N
HC
H2 C
5'
O
3'
CH
CH
O
CH2 O
24. Для двунитевых ДНК имеется еще одна специфическая особенность. При наличии в составе ДНК фрагментов, в которых чередуются пуриновые или пи
Для двунитевых ДНК имеется еще одна специфическая особенность.При наличии в составе ДНК фрагментов, в которых чередуются
пуриновые или пиримидиновые нуклеотиды, например, d(GC)n, при
достаточно высокой ионной силе наблюдается переход этого фрагмента
в левоспиральную конформацию, причем повторяющимся фрагментом в
этом случае оказывается динуклеотид d(pGpC) Линия, соединяющая
между собой фосфаты, не является винтовой, а имеет зигзагообразную
форму, в связи с этим образующаяся пространственная структура
называется Z-ДНК. В составе этой структуры G имеет син-конформацию,
а С – анти-конформацию. Фактически полному обороту спирали
соответствует 6 динуклеотидов d(pGpC), т. е. на полный оборот спирали
приходится 12 пар нуклеотидов. Фрагмент дезоксирибозы для С имеет
2’-эндо-, а для G – 2’-экзо-конформацию. На предыдущем слайде
приведена структура одной G·С пары в составе Z-ДНК.
25.
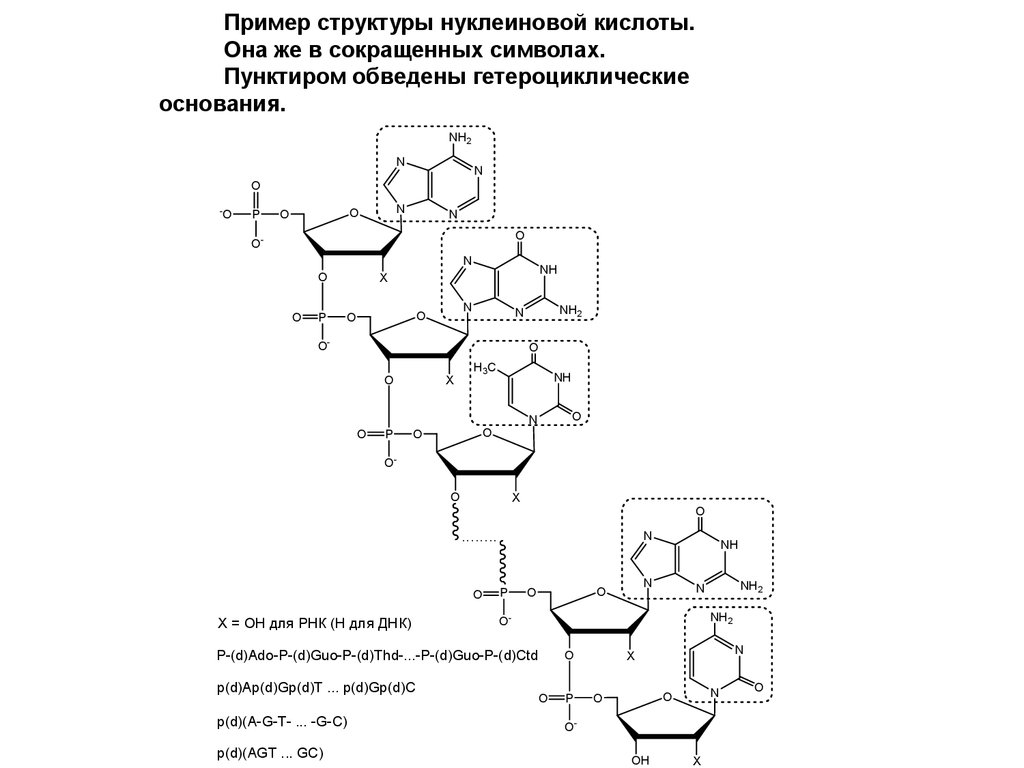
Пример структуры нуклеиновой кислоты.Она же в сокращенных символах.
Пунктиром обведены гетероциклические
основания.
NH2
N
N
O
-
O
P
N
O
O
N
O
ON
O
O
P
NH
X
N
O
O
NH2
N
O-
O
O
O
P
X
H3C
NH
O
O
N
O
OO
X
O
........
O
X = OH для РНК (H для ДНК)
N
P
p(d)(A-G-T- ... -G-C)
p(d)(AGT ... GC)
NH2
N
NH2
O-
P-(d)Ado-P-(d)Guo-P-(d)Thd-...-P-(d)Guo-P-(d)Ctd
p(d)Ap(d)Gp(d)T ... p(d)Gp(d)C
N
O
O
NH
O
O
P
N
X
N
O
O
OOH
X
O
26.
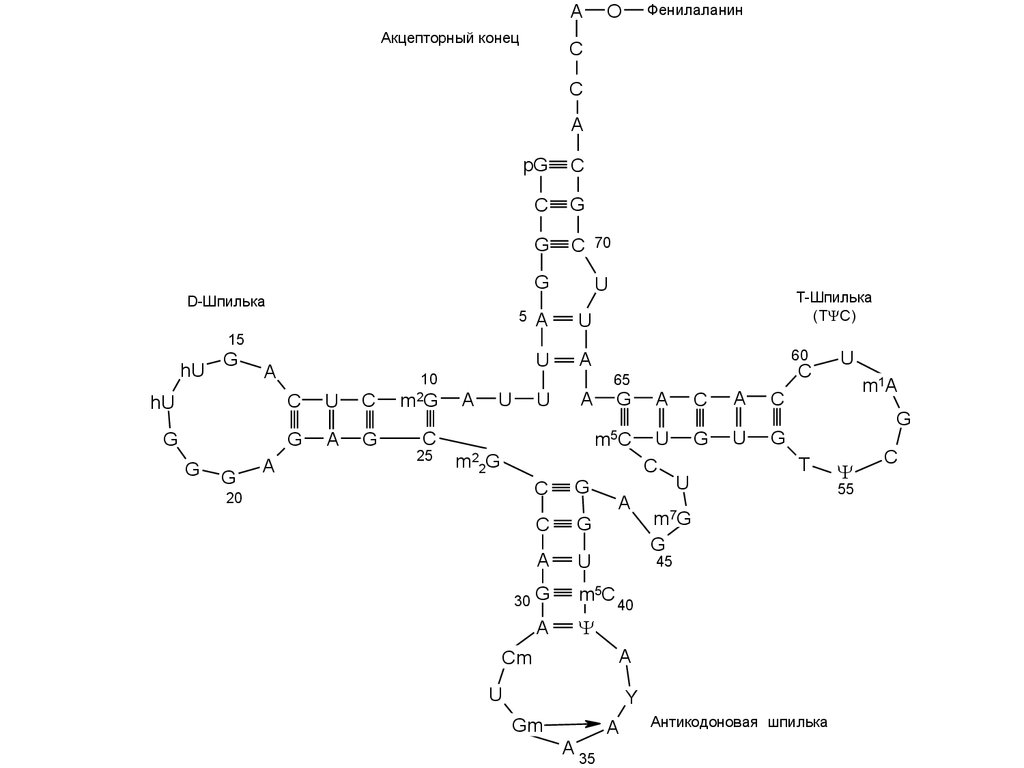
AАкцепторный конец
O
Фенилаланин
C
C
A
pG
C
C
G
G
C
G
D-Шпилька
5
70
U
A
U
T-Шпилька
(T C)
U
A
60
15
hU
G
A
10
C
hU
G
G
G
G
A
U
A
C
m2G
G
C
25
C
65
A
U
U
G
A
C
A
C
m5C
U
C
G
U
G
A
m22G
20
C
G
C
G
A
U
30 G
A
A
U
T
m7G
G
m5C 40
A
U
Y
Gm
A 35
A
m1A
G
45
Cm
U
Антикодоновая шпилька
55
C
27.
28. Взаимодействия, стабилизирующие третичную структуру тРНК
RH
O
N
N
N
R
N
N
R
N
N H
O
N
O
N H
N H
N
N H
H N
G 19
C 48
H
H
O
N
R
R
N
N H
N H
H
G 18
N
O
N
N
N
H
G 15
H N
N
H
O
H
O
R
C 56