



")







































 Биология
Биология Химия
ХимияПохожие презентации:






")



Митохондрии. Хлоропласты
1. Лекция 7
МитохондрииХлоропласты
2. Митохондрии – окраска по Альтману
3. Колокализация митохондрий и микротрубочек
Мтх – окраска родамином, МТ- окраска на альфа-тубулин4. Митохондрии в мышце - хондриом
5. Митохондрия – срез (ЭМ)
6. Митохондрии в живой клетке – окраска родамином 123
7. Компоненты митохондрий
Наружная мембрана: гладкая; содержит мало белков (отношениелипид/белок ~1:1); проницаема для веществ с молекулярным
весом менее 5 кД. За проницаемость отвечает белок – порин.
Внутренняя мембрана: связана с кристами; содержит много
белков (отношение липид/белок ~1:3); практически
непроницаема для любых веществ (имеет специальные белкипереносчики); в норме заряжена (- 200 мВ).
Мембрана крист: заряжена; содержит цепь переноса электронов и
АТФ-азу; соединяется с внутренней мембраной.
Межмембранные контакты ( митохондриальные поры) –
обеспечивают избирательный перенос белков из матрикса в
цитозоль и обратно.
Матрикс – имеет слабощелочной рН (рН=8); содержит белки цикла
Кребса, нуклеиновые кислоты (ДНК, РНК) и рибосомы
митохондрий; аккумулирует ионы кальция.
8. Структура митохондрий
9. Транспорт белков в митохондрии
Сигнальная последовательность митохондриальных белков, как правило, расположенана N-конце, и она отщепляется после пересечения молекулой белка митохондриальной
мембраны.
Белки переносятся в неупакованном (unfolded) состоянии.
Перенос через мембраны происходит с помощью специальных комплексов переносчиков.
Перенос в матрикс происходит через зоны контакта наружной и внутренней мембран,
однако комплексы на каждой мембране могут работать независимо.
Встраивание белка во внутреннюю мембрану (мембрану крист) требует двух сигнальных
последовательностей и происходит из матрикса митохондрий.
Некоторые белки попадают в митохондрии непосредственно из ЭПР через специальные
зоны контакта мембран ЭПР и митохондрий.
10. Динамика митохондрий
Митохондрии в клетке имеют непостоянную форму ифункционально связаны в единую сеть (быстро
обмениваются своим внутренним содержимым). Обмен
внутри сети происходит за счет слияний и делений, которые
обеспечиваются специальными белками .
Для сети митохондрий важен баланс активностей –
ингибирование слияния может быть компенсировано
подавлением деления. Физиологическая регуляция данных
процессов пока не изучена.
Общий объем митохондриальной сети связан с
интенсивностью дыхания. Наиболее сильные изменения при
стимуляции дыхания касаются белков крист.
В различных патологических состояниях может происходить
фрагментация митохондрий – разделение сети на
сравнительно небольшие органеллы.
11. Фрагментация митохондрий
Физиологическая рольфрагментации митохондрий
– удаление дефектных
компартментов органелл.
12. Регуляция слияния митохондрий
13. Основные функции митохондрий
Окислительное фосфорилирование: генерацияразности потенциалов в результате окисления
органических субстратов; синтез АТФ в матриксе и
его экспорт в цитозоль; поддержание низкого
уровня АДФ в клетке.
Регуляция уровня внутриклеточного кальция;
депонирование кальция.
Регуляция апоптоза – митохондрии содержат
«белки смерти», а также участвуют в активации
эффекторных каспаз.
Синтез некоторых митохондриальных белков,
липидов и цитоплазматическая наследственность.
14. Цепь переноса электронов и цикл трикарбоновых кислот
15. Перенос электронов в мембране митохондрии
16. Цепь переноса электронов
17. Работа внутренней митохондриальной мембраны
18. АТФ-аза митохондрий и прокариот
АТФ-аза – миниатюрный электромотор,вмонтированный в мембрану
перпендикулярно ее плоскости.
Ротор – гамма-субъединица; статор –
комплекс из трех альфа и трех бета
субъединиц.
Вращение ротора в одну сторону
приводит к гидролизу АТФ, а в
противоположную – к синтезу АТФ
(реверс возможен только при наличии
достаточного электрохимического
потенциала).
19. АТФ-АДФ транслоказа
Составляет около 10% от общего белка внутренней мембраны. Весьмаконсервативна, в особенности, канал для связывания нуклеотидов. У
человека представлена несколькими ткане-специфичными изоформами.
20. Хемиосмотическая теория
Цепь переноса электронов переносит протоны из матриксамитохондрии в цитозоль в процессе снижения уровня
свободной энергии.
Митохондриальная АТФ-синтетаза переносит протон через
внутреннюю мембрану и синтезирует АТФ в матриксе. Ее
работа обратима, и направление переноса зависит от
величины электрохимического потенциала.
Внутренняя мембрана в остальном непроницаема для H +,
ОН- и большинства других ионов.
Для снабжения АТФ цитоплазмы внутренняя мембрана
имеет специальный переносчик для противоположного
переноса АДФ (внутрь) и АТФ (наружу).
21. Генерация энергии митохондриями
Универсальная форма энергии – электрохимическийпотенциал на мембране (ΔH+ у эукариот, ΔН+ или ΔNa+ у
прокариот).
Синтез АТФ происходит в матриксе за счет рассеивания
электрохимического потенциала.
Максимальный к.п.д. синтеза АТФ (3 пункта сопряжения:
НАДН+-дегидрогеназа + цитохромы b-c1 + цитохромоксидаза) –
около 50%.
Для поддержания баланса между матриксом митохондрий и
цитозолем через внутреннюю мембрану происходит:
- совместный перенос в матрикс ионов фосфата и водорода
- противоположный перенос АДФ (внутрь) и АТФ (наружу)
22. Регуляция окислительного фосфорилирования через систему обратных связей
Дыхательный контроль – в норме потребление кислородаограничивается концентрацией АДФ в цитоплазме и
потенциалом на митохондриальной мембране.
Ингибиторы дыхания – яды, которые подавляют перенос
электронов по цепи (азид, цианид, антимицин А, ротенон и др.).
Ингибитор АТФ-азы – олигомицин.
Разобщители (ускорители дыхания) – некоторые жирные
кислоты. Синтетические разобщители – ионофоры для Н+ (2,4динитрофенол, FCCP и др.) Максимальная скорость окисления
при разобщении ~ в 3 раза превышает нормальную.
23. Ингибиторы митохондрий
Разобщители – FCCP, 2,4-ДНФ, жирные кислоты. Повышаютпроницаемость мембраны для протонов. Сильнейшие яды.
Увеличивают скорость дыхания.
Ингибиторы АТФ-синтазы (нигерицин). Уменьшает скорость
дыхания.
Бонгкрековая кислота – ингибитор переноса АТФ/АДФ.
Сильнейший яд, защищает клетки от индуцированного
апоптоза. Уменьшает скорость дыхания.
.
24. Митохондрия как терморегулятор
Жирные кислоты – природные разобщители,которые регулируют соотношение окисления и
синтеза АТФ.
Митохондрии в клетках бурого жира млекопитающих
обеспечивают терморегуляцию и быстрый разогрев
организма. В них содержится специальный белок –
термогенин, который активируется за счет работы
нервной системы (симпатических нервов).
25. Энергетические затраты животной клетки
Работа K/Na–АТФазы – 15-60% энергии (в среднем около 35%).Поддержание протонного градиента на мембране крист
митохондрий – 20-40% энергии.
Биосинтез белка – 18-26%, иногда больше (в быстро растущих
клетках).
Репликация ДНК и транскрипция – до 10%.
Деградация белков с помощью убиквитина и протеасом –
точных данных нет.
Работа протеинкиназ и других АТФ-аз/ГТФ-аз, не упомянутых
выше – не более 1%.
26. Растительная клетка с хлоропластами и вакуолью
27. Схема организации хлоропласта
28. Хлоропласт, вид на срезе
Характерный размер – 1х5 мкм. Нередко хлоропласты образуютсеть, имея непрерывную наружную мембрану.
29. Хлоропласт, детали структуры
30. Мембраны хлоропласта
Наружная мембрана: гладкая; высокопроницаемая.Внутренняя мембрана: гладкая (не образует крист);
практически непроницаема для любых веществ, но содержит
много трансмембранных белков-переносчиков.
Мембрана гран – похожа на внутреннюю мембрану. Она
асимметрична по липидам и содержит в основном
фотосинтетические пигменты и АТФ-синтазу.
Все мембраны содержат наряду с фосфолипидами
специфические липиды: сульфолипиды, галактолипиды,
фосфатидилглицерин.
Происхождение мембран хлоропласта: все они являются
производными ЭПР.
31. Компоненты хлоропласта
Строма имеет щелочной рН. Она содержит нуклеиновыекислоты (ДНК, РНК) и рибосомы хлоропласта; содержит в
высокой концентрации метаболические ферменты;
аккумулирует крахмал (амилоид). В ней происходит
«темновой» этап фотосинтеза (фиксация СО2)
Тилакоиды – система мембран, образующих стопки – граны;
на свету заряжена (-200 мВ).
Граны содержат фотосистемы, цепь переноса электронов и
систему синтеза АТФ
Матрикс тилакоида имеет кислый рН.
32. Основы фотосинтеза
Молекула хлорофиллаВозможные преобразования энергии кванта
света в молекулярном ансамбле:
1. флуоресценция
2. Ферстеровский резонансный перенос
энергии
3. перенос электронов
Строение фотосистемы:
1. антенный комплекс – хлорофиллы (100-400
молекул) + каротиноиды + белки
2. фотохимический реакционный центр
(трансмембранный белок с двумя
специальными молекулами хлорофилла)
33. Световые реакции фотосинтеза
Фотосистема II – комплекс из примерно 20 белков с большимколичеством трансмембранных доменов. Она генерирует
кислород, протоны и восстановленный пластохинон. В результате
работы фотосистемы II на мембране накапливается
электрохимический потенциал, достаточный для синтеза АТФ.
Основную роль в фотолизе воды и выделении кислорода играют
четыре атома марганца и две молекулы хлорофилла (Р-680),
расположенные в центре антенны. Окисленный Р-680 является
самым сильным из известных окислителей – он имеет
окислительно-восстановительный потенциал 1,3 В. Для фотолиза
воды необходим потенциал около 1 В (1000 мВ).
Фотосистема I – переносит электроны от пластоцианина к
ферредоксину и генерирует восстановленный НАДФ. В ее состав
входит димер хлорофилла (Р-700) и пять молекул акцепторов
электронов. Основной комплекс состоит из двух субъединиц с
большим числом трансмембранных доменов. Антенна
фотосистемы I содержит хлорофилл (100-200 молекул) и
каротиноиды (20-50 молекул).
34. Перенос электронов в мембране хлоропласта
Перенос протонов поперек мембраны граны хлоропласта аналогичен переносупоперек внутренней мембраны в митохондриях. Источником энергии в
хлоропласте являются две светособирающие антенны. К.п.д. – около 40%.
35. Цепь переноса электронов и фотосистемы
36. Световые реакции
37. Световые и темновые реакции фотосинтеза
Световые реакции – зависят от постоянного потока фотонов ипротекают на мембранах тилакоидов.
Они включают в себя генерацию радикалов, работу электронтранспортной цепи по разделение зарядов, восстановление
НАД-фосфата, синтез АТФ, фотолиз воды и генерацию
кислорода.
Темновые реакции – превращение двуокиси углерода в
трехуглеродные сахаро-фосфаты. Эти реакции продолжаются
некоторое время в темноте, пока не истощается пул АТФ и
восстановленного НАД-фосфата.
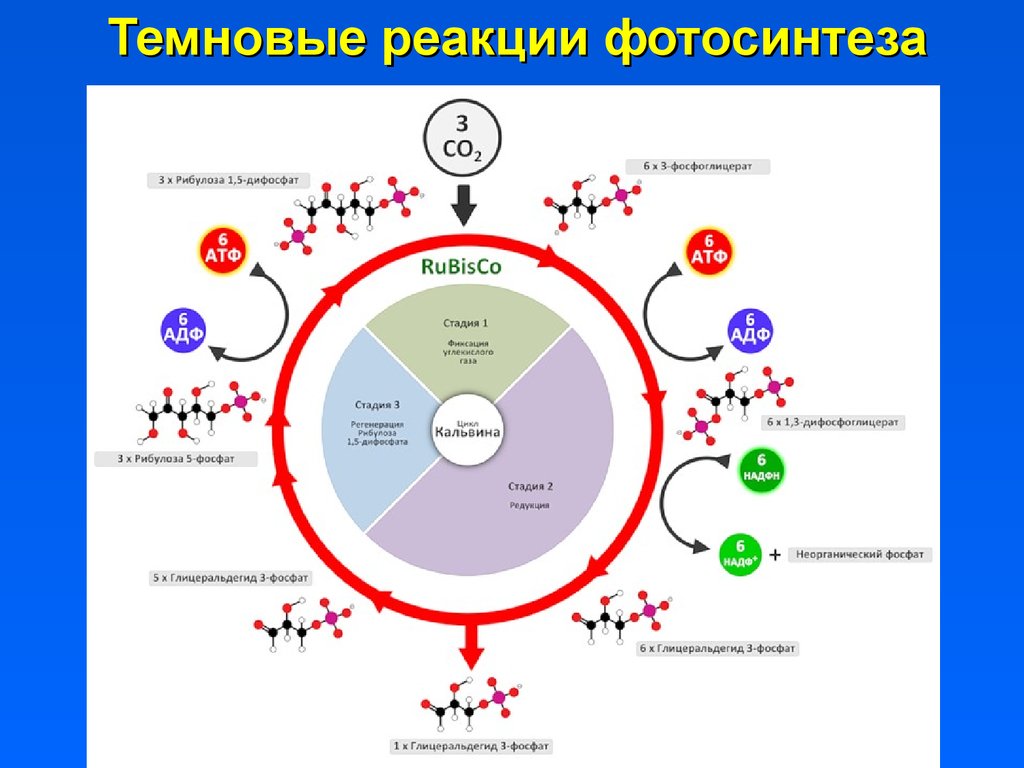
38.
Темновые реакции фотосинтеза39. Развитие хлоропластов
Хлоропласты (например, в побегах) развиваютсяпод действием света из пропластид, лишенных
гран (т.н. агранулярные хлоропласты).
40. Дифференцировка хлоропластов
Пропластиды – бесцветные двумембранные структуры, диаметром0,4-1 мкм, содержащие ДНК хлоропластов. Они содержатся в
яйцеклетках растений.
Хлоропласты возникают из пропластид на свету (после прорастания
семян). При формировании хлоропластов в пропластидах возникают
складки внутренней мембраны и формируется трехмембранная
система, содержащая граны.
Из пропластид могут возникать амилопласты, где запасается крахмал
(картофель). На свету амилопласты могут превращаться в
хлоропласты.
Хлоропласты могут дифференцироваться в хромопласты под
воздействием ядерных продуктов. Хромопласты накапливают
каротиноиды.
41. Деление хлоропластов
Деление хлоропластов происходит медленно. Оно регулируетсянесколькими группами белков – динамин, FtsZ (аналог
прокариотического белка) и пластидные кольца. Каждая группа
белков образует соответствующие полимерные кольца.
42. Сравнение митохондрий и хлоропластов
Первичные источники энергииЦепь переноса электронов на мембране
Протонная помпа – синтез АТФ
Две мембраны/три мембраны
Автономная система биосинтеза белка
(ДНК, РНК, рибосомы)
43. Митохондрии и хлоропласты: нуклеиновые кислоты
Митохондрия: кольцевые молекулы ДНК – 16 кБ, кодируют 13 белков(часть рибосомных, электрон-транспортной системы и субъединица
АТФ-азы), 22 тРНК и 2 рРНК. Митохондрии растений имеют геном в 10
раз больше. Имеют собственный генетический код. ~10 кольцевых
молекул на одну митохондрию.
Хлоропласт: кольцевые (иногда линейные) молекулы ДНК – 120-170
кБ, содержат около 120 генов (часть рибосомных белков, некоторые
ферменты фотосинтеза, все т-РНК, 3 р-РНК). Для трансляции
используется ядерный генетический код, но и-РНК сильно
трансформируются (аномальный процессинг). ~100 молекул на
молодой хлоропласт, при старении число снижается до ~10/
Рибосомы органелл: в митохондриях 55S (меньше бактериальных); в
хлоропластах – 70S (гомологичны бактериальным)
44. Митохондрии и хлоропласты: полуавтономность
Митохондрия: может функционировать без собственной ДНК,однако эффективность дыхания резко снижается.
У человека ряд наследственных заболеваний (встречаются
крайне редко и передаются только по женской линии) связан с
мутациями в митохондриальных белках.
Нарушение транспорта белков в митохондрии приводит к
снижению эффективности их работы.
Хлоропласт: не может функционировать без собственной ДНК.
Транспорт белков в хлоропласты обеспечивается развитой
системой шаперон-подобных белков. Его нарушение приводит
к резкому снижению эффективности фотосинтеза.
45. Митохондрии и хлоропласты - наследственность
Митохондрии и хлоропласты наследственностьПроисхождение органелл – симбиоз (слияние клеток и последующая
редукция генома с переносом части генов в ядро). Митохондрии – из
бактерий (типа риккетсий), хлоропласты – из цианобактерий.
Наследование митохондрий происходит только по материнской линии
у животных, растений и грибов. Механизмы: разбавление или
исключение отцовских митохондрий на стадии оплодотворения.
Наследование хлоропластов происходит только по материнской
линии у цветковых растений.
У растений митохондриальная и хлоропластная ДНК часто
обмениваются участками друг с другом.