
















 и не работает в других (b)")





















 Биология
Биология Экология
ЭкологияПохожие презентации:








")

Биоразнообразие. Biodiversity
1.
БИОРАЗНООБРАЗИЕBIODIVERSITY
2.
КАК ИЗМЕРЯТЬ ВИДОВОЕРАЗНООБРАЗИЕ
(БИОРАЗНООБРАЗИЕ
BIODIVERSITY)
1. Видовое богатство (число видов без
случайных и временных вселенцев)
2. Гетерогенность (в одном 2 вида 99:1, в
другом 50:50) тем выше, чем больше
видов и чем более равномерно они
распределены. Функция ШенонаУинера
3. Широтные градиенты биоразнообразия
4.
ШИРОТНЫЕ ГРАДИЕНТЫ5. Какие факторы могут определять градиенты биоразнообразия
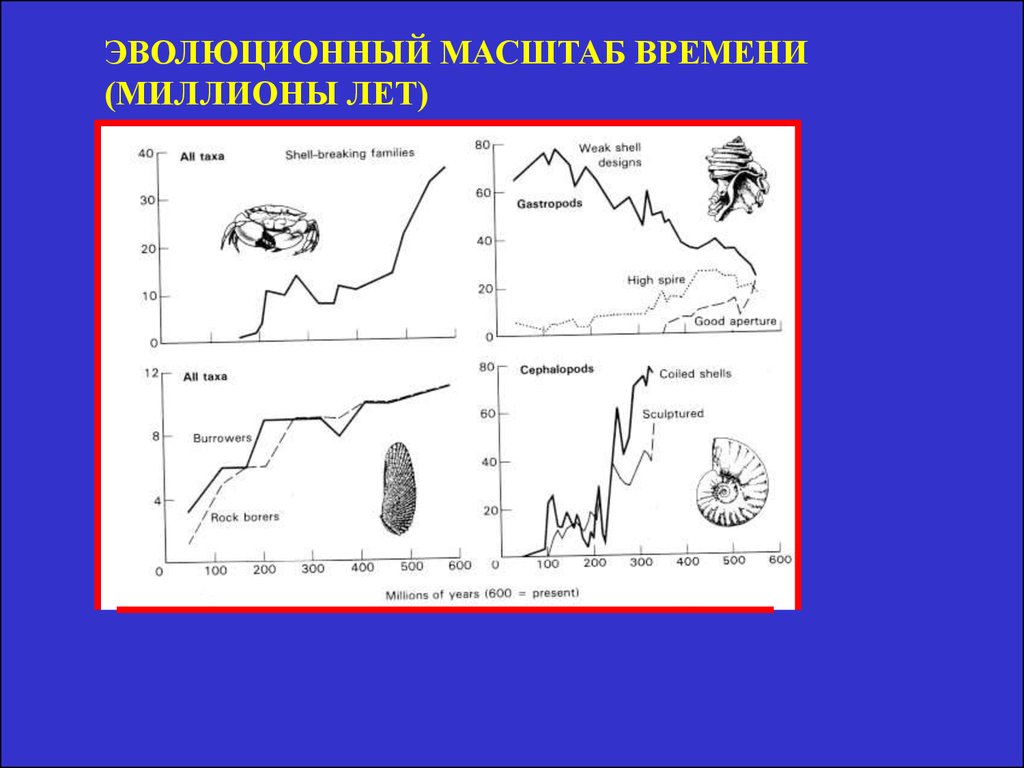
• Время в эволюционном масштабеТо есть в старых сообщества больше видов, чем в
молодых. Байкал – 580 видов бентоса, Б. Невольничье
Озеро – 4.
Число видов наземных
растений за 450 млн.л.
не достигает плато.
6.
ЭВОЛЮЦИОННЫЙ МАСШТАБ ВРЕМЕНИ(МИЛЛИОНЫ ЛЕТ)
7. Гипотетическая схема повышения биоразнообразия со временем.
Проверка истории,как фактора трудна.
поэтому сначала надо отбросить остальные более простые.
8. Пространственная гетерогенность
• МакАртур (1965) ввелпонятия:
Внутрибиотопическое
(альфа) р-зие и
Межбиотопическое (бета)
р-зие
• Вариант А
в тропиках увеличено бета р-е
• Вариант Б
в тропиках выше альфа р-е
Показано (MacArthur, 1969),
что и то, и другое: 2.7
больше на 2 га для птиц,7
раз больше на больших S
9. Простая гипотеза разнообразие животных зависит от разнообразия деревьев, проходит только для амфибий
10.
• Увеличение биоразнообразия в тропикахпоказано и для водных экосистем, где нет
гетерогенности.
• Таким образом, увеличение
пространственной гетерогенности, не
объясняет полностью полярнотропические градиенты.
11. Конкуренция
• Предполагают: Конкуренция играетбольшую роль в тропиках, тогда как в
высоких широтах численность
ограничена физическими факторами
среды
• Однако, по правилу Гаузе, числу видов
должно соответствовать число факторов,
по которым они расходятся.
• Для растений 4-5 факторов. Но число
видов на 2 порядка больше.
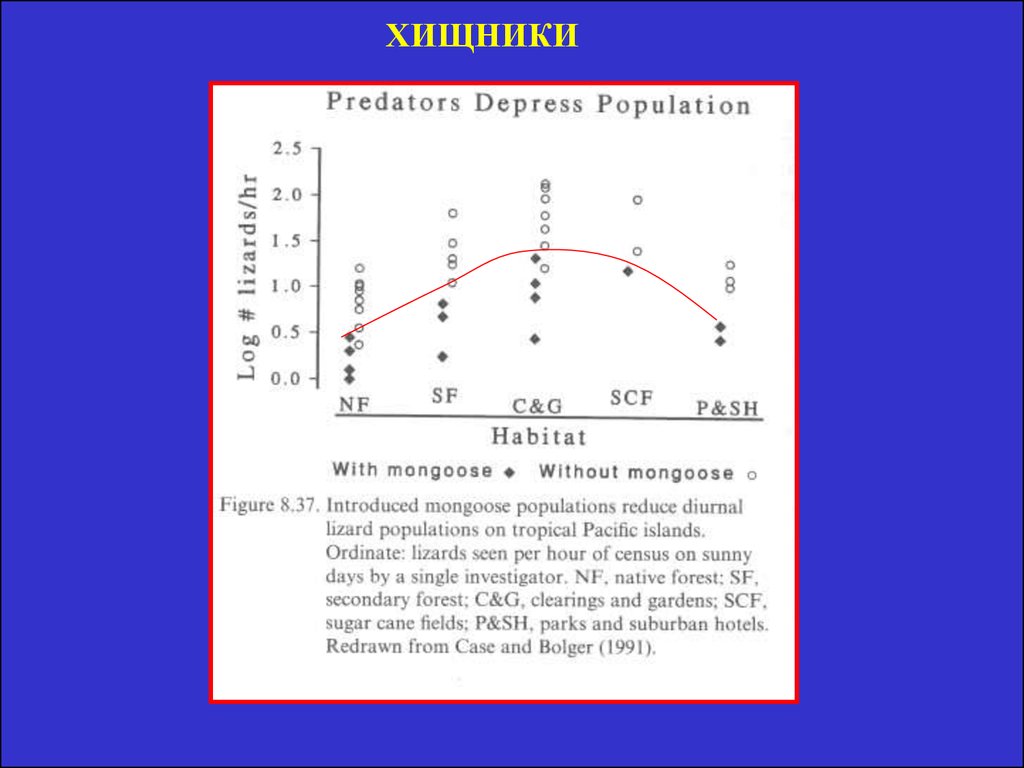
12. Хищничество
• Предполагают: Хищники держатпопуляции жертв на таком низком
уровне, что они не конкурируют. Это
позволяет встраиваться новым видам
жертв и появляться новым хищникам.
• Пайн (1966) показал, что отсутствие
морской звезды приводит к сокращению
числа видов с 15 до 8, так как мидии всех
вытесняют.
• Число ключевых (key-stone) видов
хищников в тропиках должно больше. Но
доказательств нет.
13.
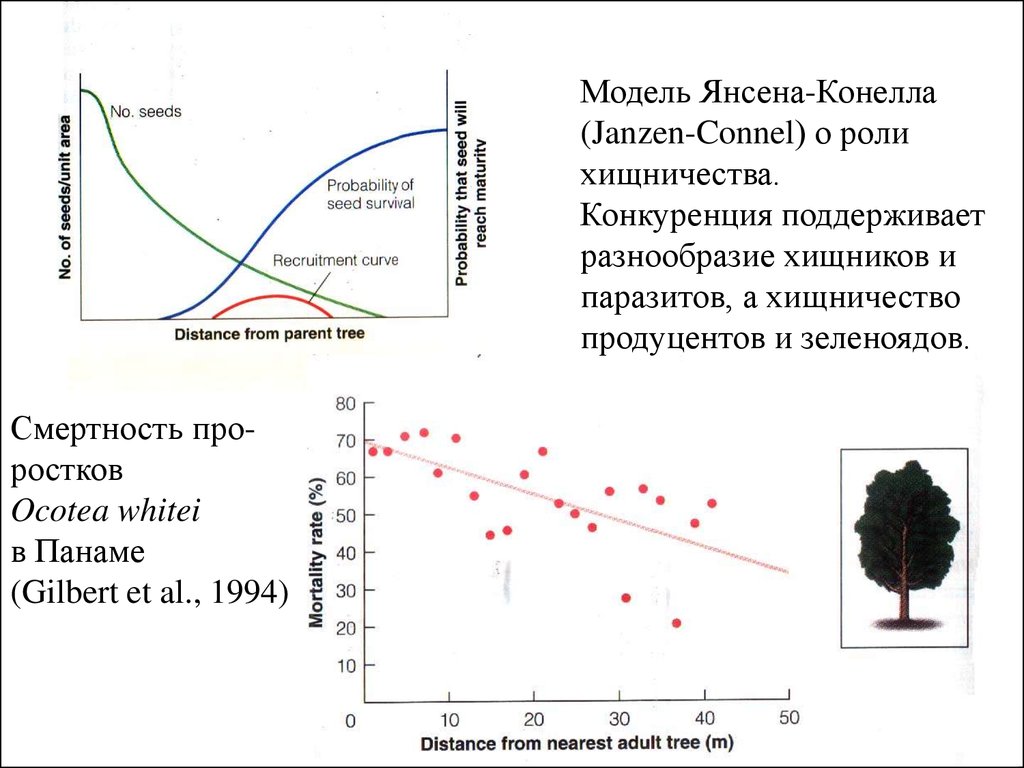
Модель Янсена-Конелла(Janzen-Connel) о роли
хищничества.
Конкуренция поддерживает
разнообразие хищников и
паразитов, а хищничество
продуцентов и зеленоядов.
Смертность проростков
Ocotea whitei
в Панаме
(Gilbert et al., 1994)
14.
ХИЩНИКИ15. Климат и его флуктуации
• Более стабильный и мягкий климат – большевидовое богатство (Sanders, 1968).
• Сообщества с низким разнообразием могут
быть связаны с суровым (горячие источники)
или непредсказуемыми условиями (пустыни)
• Сандерс (1968) сравнивал донные
глубоководные сообщества по богатству
достигающие тропических мелководий. Другие
(Abele, Walters, 1979) объясняют
гетерогенностью.
16.
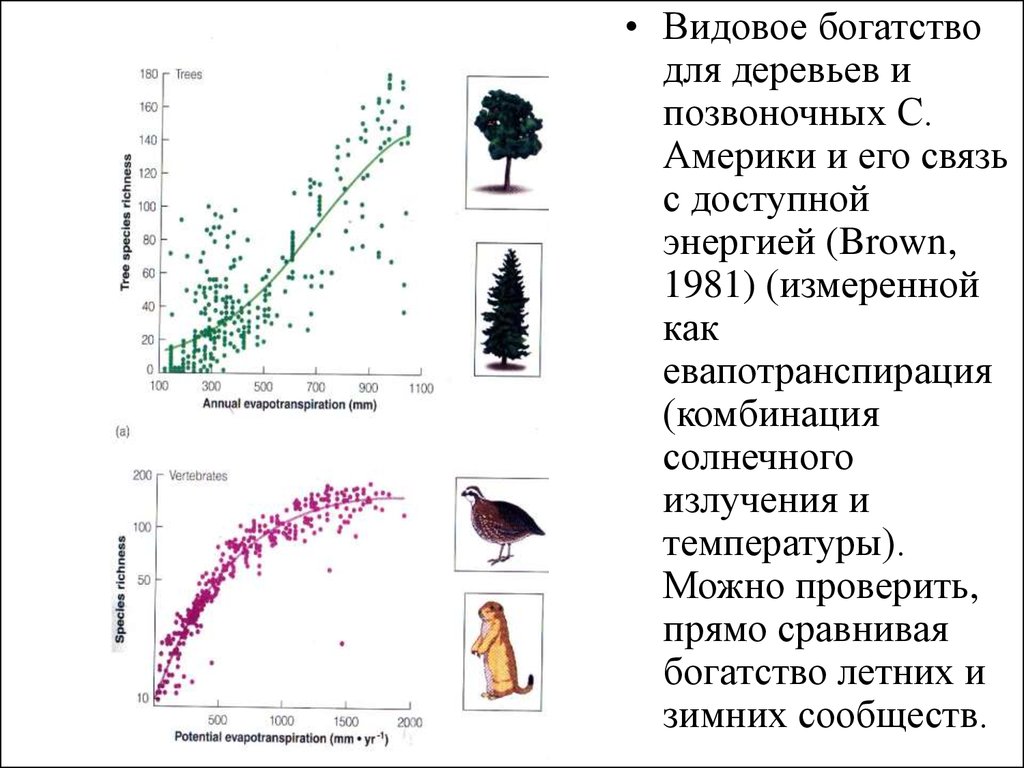
• Видовое богатстводля деревьев и
позвоночных С.
Америки и его связь
с доступной
энергией (Brown,
1981) (измеренной
как
евапотранспирация
(комбинация
солнечного
излучения и
температуры).
Можно проверить,
прямо сравнивая
богатство летних и
зимних сообществ.
17. Продуктивность
• Гипотеза, чем выше продуктивностьсообщества, тем выше разнообразие не
проходит.
• Самые богатые растительные
сообщества не на самых богатых почвах
18.
ОБИЛИЕ РЕСУРСОВ (ПРОДУКТИВНОСТЬ)19. Модификация гипотезы влияния продуктивности.
• Более длительный сезон вегетации –более дробное временное деление
ресурсов. Комбинация стабильности и
продуктивности (Connel, Orias, 1964).
20.
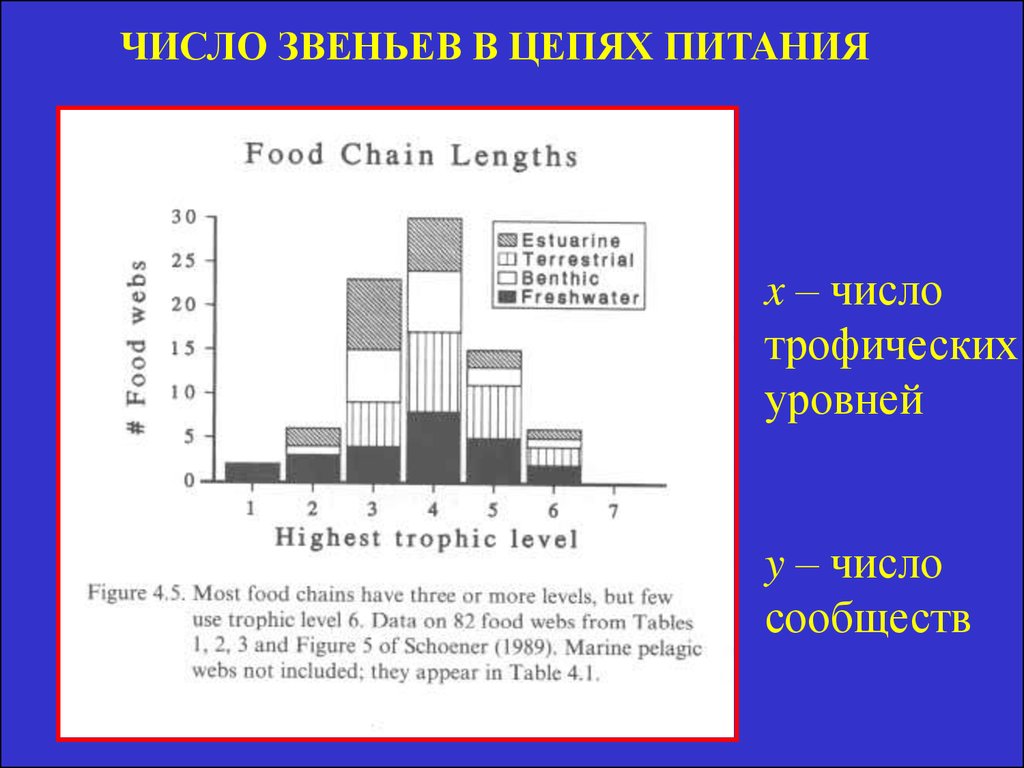
ЧИСЛО ЗВЕНЬЕВ В ЦЕПЯХ ПИТАНИЯx – число
трофических
уровней
y – число
сообществ
21. Нарушения
• В равновесии доминируют самыеконкурентные.
• Хищничество, пастьба, изменения
погоды, катастрофы (пожары) нарушают
равновесие.
• Часто плохо и редко плохо. Intermediate
disturbance hypothesis (Grime, 1973 и др.)
22.
• Оценка роли нарушений зависит отпространственной шкалы. Для
небольших S – разнообразие стадий
сукцессии. Но на большей шкале –
включает все виды.
23. Гипотеза «среднего уровня нарушений» работает в одних биотопах (а) и не работает в других (b)
24. Примеры сообществ, где гипотеза не работает
• Донные с-варучьев Южного
о-ва Новой
Зеландии
(температура,
течение, дно)
• Растительные сва (Частота
пожаров в
Канзасе)
25. Локальное и региональное разнообразие
• Локальное разнообразие определяетсяшкалой при которой виды сообщества
могут взаимодействовать.
• Шкала регионального разнообразия в
сотни раз больше.
26.
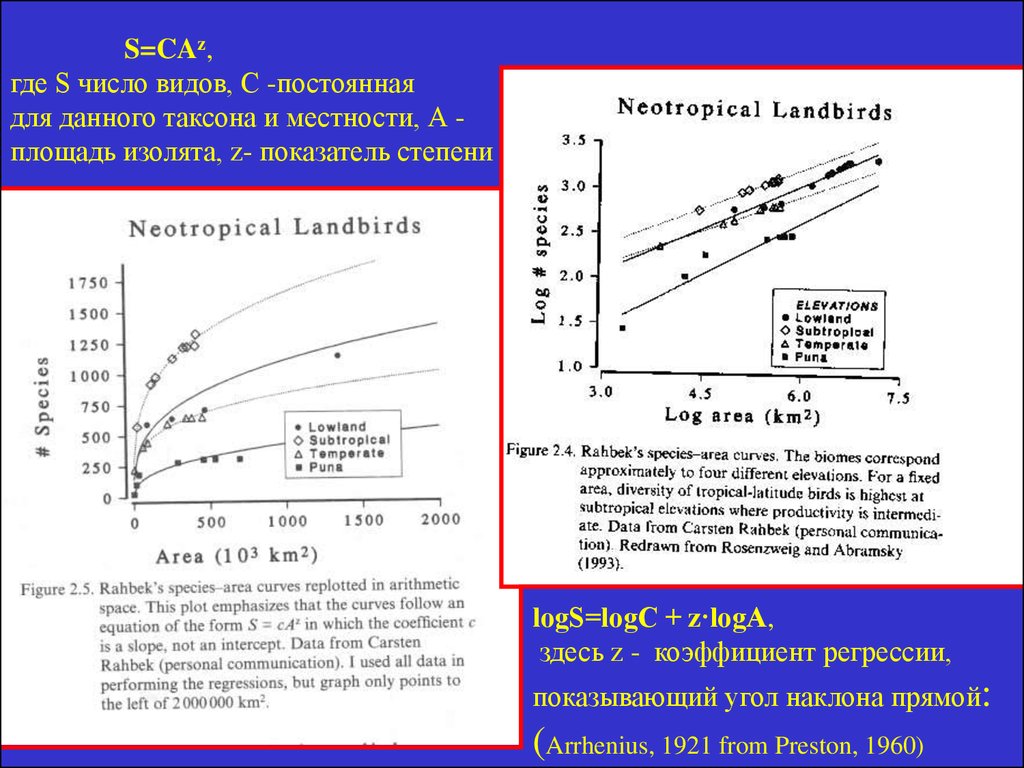
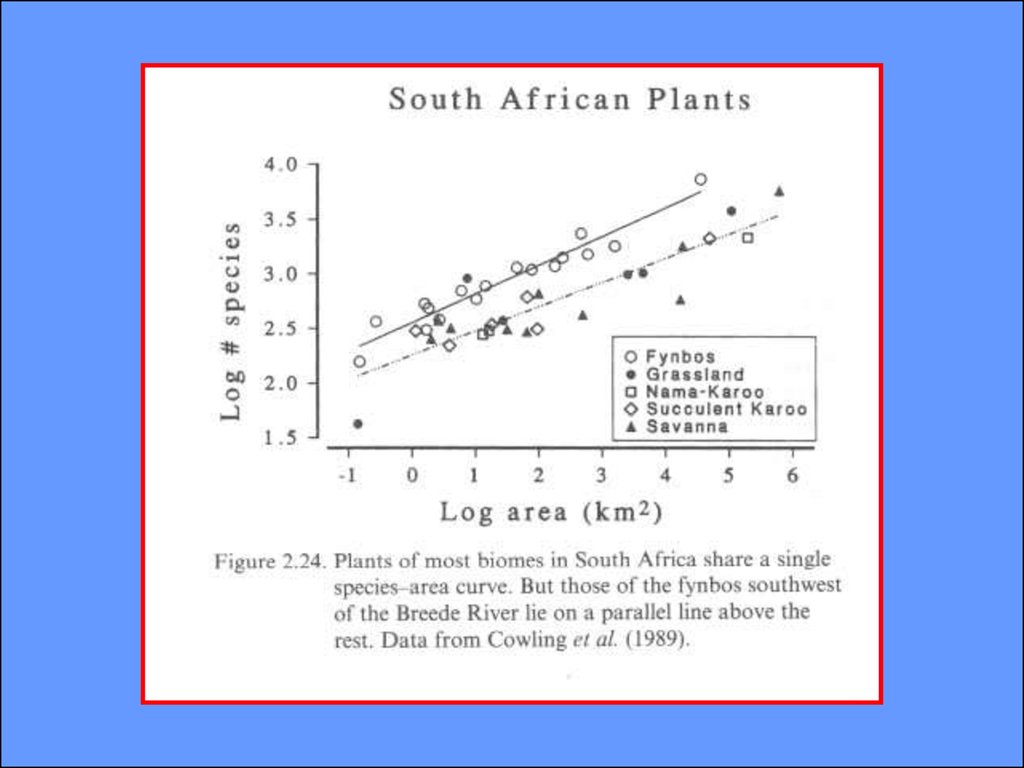
СВЯЗЬ ЧИСЛА ВИДОВ С ПЛОЩАДЬЮ ИЗОЛЯТАS=CAz,
где S число видов, С -постоянная
для данного таксона и местности, А площадь изолята, z- показатель степени, или
коэффициент регрессии,
показывающий угол наклона прямой:
lоgS=lоgC + z·lоgA
27.
S=CAz,где S число видов, С -постоянная
для данного таксона и местности, А площадь изолята, z- показатель степени
lоgS=lоgC + z·lоgA,
здесь z - коэффициент регрессии,
показывающий угол наклона прямой:
(Arrhenius, 1921 from Preston, 1960)
28.
29.
30.
31.
БОГАТСТВО ОСТРОВНОЙ ФАУНЫ (MacArthur, Wilson, 1963)32. Гипотетическое соотношение локального и регионального видового богатства в зависимости от насыщенности сообщества
33. Экспериментальная проверка
1. Птицы 15 га – 200-700 ,ч. набл. (Pearson, 1977)
2. Разные сообщества на
разных континентах (Caley,
Schluter, 1997)
34.
35.
ЭНТРОПИЙНАЯ МЕРА РАЗНООБРАЗИЯШЕННОНА-УИВЕРА
H=- PilogPi
36.
37.
38.
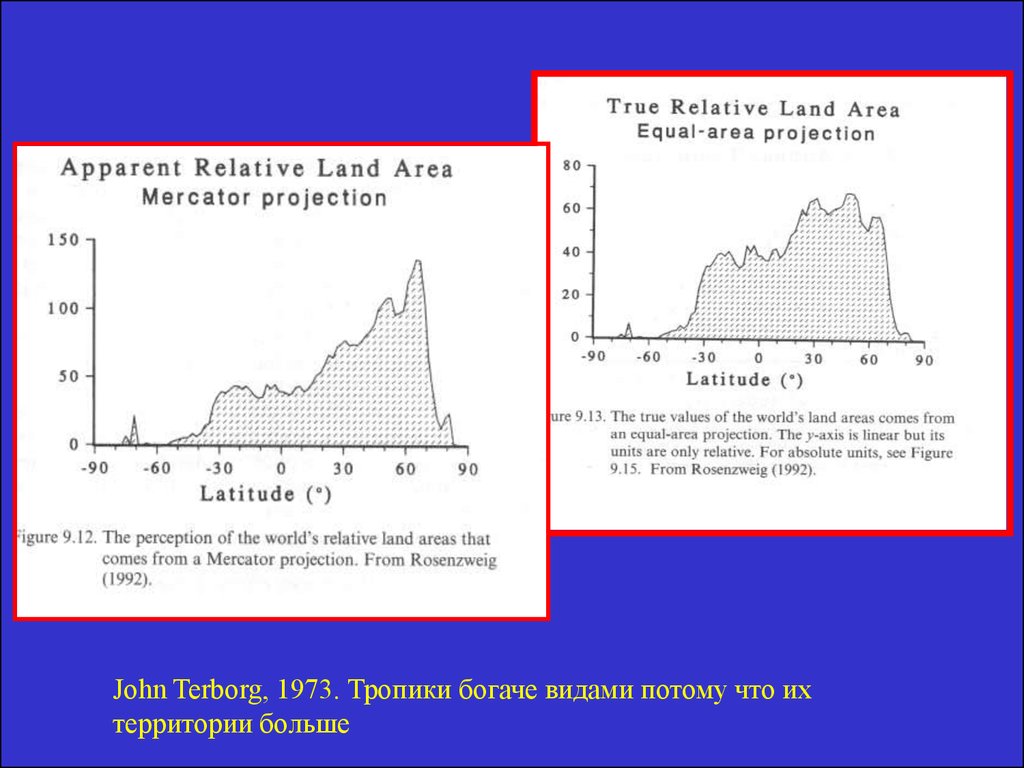
John Terborg, 1973. Тропики богаче видами потому что ихтерритории больше
39.
ЭКОЛОГИЧЕСКИЙ МАСШТАБ ВРЕМЕНИ40.
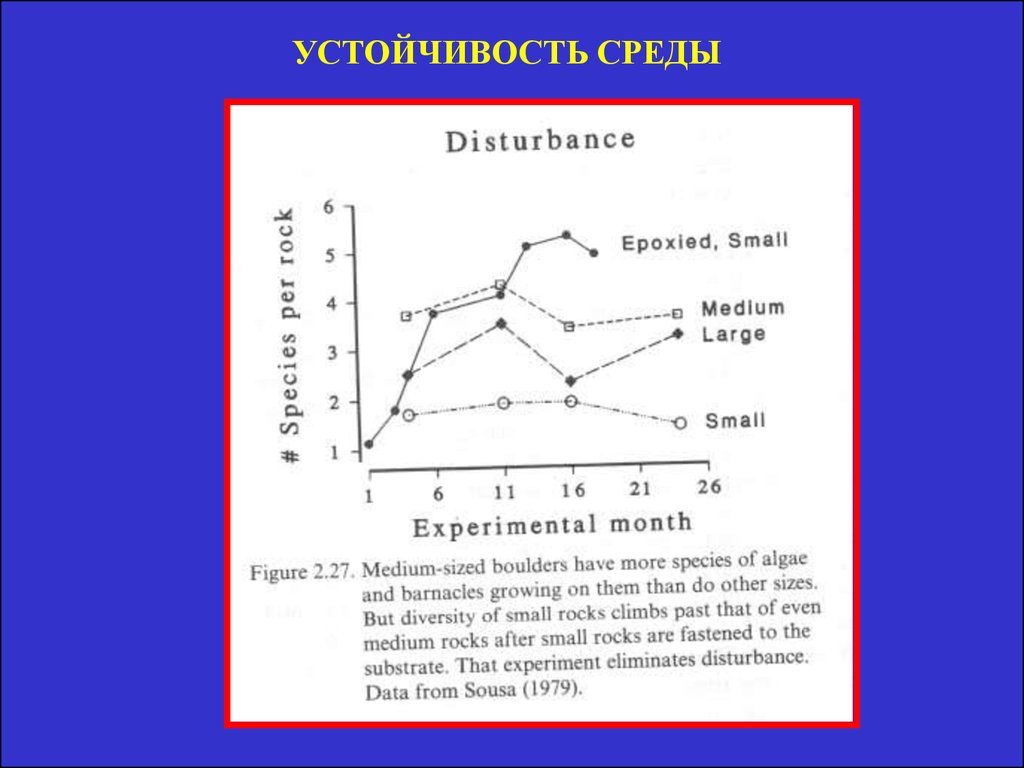
УСТОЙЧИВОСТЬ СРЕДЫ41.
ГЕТЕРОГЕННОСТЬ СРЕДЫ42.
КОНКУРЕНЦИЯ43.
ГИПОТЕЗЫ, ОБЪЯСНЯЮЩИЕ РАЗНООБРАЗИЕВИДОВ В СООБЩСТВАХ
1. РАЗМЕР ТЕРРИТОРИИ
3. ВРЕМЯ. ЭВОЛЮЦИОННЫЙ МАСШТАБ
4. ВРЕМЯ. ЭКОЛОГИЧЕСКИЙ МАСШТАБ
5. УСТОЙЧИВОСТЬ КЛИМАТА И СПЕЦИАЛИЗАЦИЯ
6. ПРЕДСКАЗУЕМОСТЬ КЛИМАТА И СПЕЦИЛИЗАЦИЯ
7. ГЕТЕРОГЕННОСТЬ ПРОСТРАНСТВА
8. ПРОДУКТИВНОСТЬ
9. УСТОЙЧИВОСТЬ ПЕРВИЧНОЙ ПРОДУКЦИИ
8. КОНКУРЕНЦИЯ
10. РАЗРЕЖИВАНИЕ (ВЛИЯНИЕ КЛИМАТА)
11. РАЗРЕЖИВАНИЕ (ВЛИЯНИЕ ХИЩНИКОВ)
44.
45.
ЭКОЛОГИЧЕСКИЙ МАСШТАБВРЕМЕНИ (ГОДЫ)
Динамика структуры сообщества
грызунов Черных земель,
Калмыкия