

















 Биология
БиологияПохожие презентации:








")

Потенциал Действия
1.
Потенциал Действия2.
3.
4.7. Потенциал действияУравнение Гольдмана позволяет вычислить значение мембранного потенциала покоя, если
известна концентрация ионов внутри клетки и снаружи, а также проницаемость для этих ионов.
Реальная величина мембранного потенциала покоя ближе всего к значению равновесного
потенциала для ионов калия, которые проходят через постоянно открытые для них каналы.
Ситуация решительно изменяется при раздражении клетки, когда повышается натриевая
проницаемость и появляется деполяризующий рецепторный потенциал или
постсинаптический потенциал.
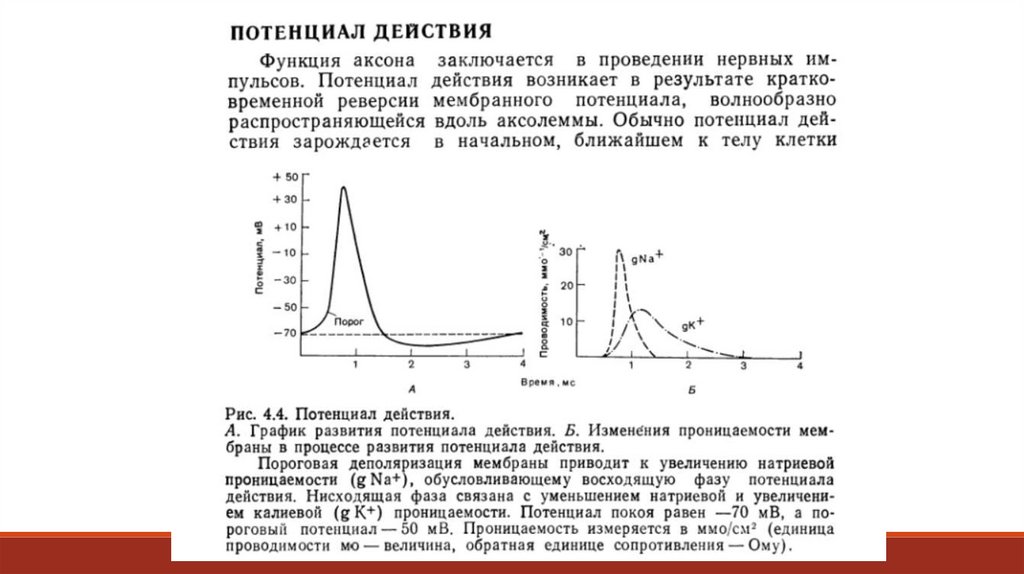
Потенциал действия возникает лишь при определённой величине деполяризующего сдвига,
например, с -65 мВ до -55 мВ. Если деполяризация будет меньше, то потенциал действия не
возникнет: такие деполяризующие сдвиги называются подпороговыми. Приведённые здесь
цифры относительны, в разных клетках они могут быть меньше или больше, но всегда
наименьший деполяризующий сдвиг, который вызовет появление потенциала действия,
определяется, как пороговый. Возникновение рецепторного или постсинаптического
потенциалов связано со сравнительно небольшим местным повышением натриевой
проницаемости мембраны.
4.
Вход ионов натрия в клетку и обусловленная этим местная деполяризацияприводят к возникновению локального электрического тока. Его
распространению вдоль мембраны препятствует электрическое
сопротивление самой мембраны, поэтому начавшаяся в каком-нибудь
месте пассивная деполяризация не может распространяться далеко –
пассивные электрические ответы всегда локальны.
Но, если сумма локальных деполяризующих сдвигов всё-таки сможет
деполяризовать мембрану триггерной зоны нейрона до критического
уровня, до порогового значения, то произойдёт активный и максимальный
ответ клетки по правилу "всё или ничего".
Деполяризация до критического значения ведёт к конформационным
изменениям внутренней стенки натриевых каналов и перемещению
полярных аминокислот. Вследствие этого открывается пора диаметром 0,3
– 0,5 нм через которую могут пройти катионы натрия (См. рис. 4.3).
Ток анионов через этот канал невозможен, поскольку его устье содержит
отрицательные заряды карбоксильных групп глутаминовой кислоты,
которые отталкивают отрицательные заряды анионов.
5.
Равновесный потенциал натрия +55 мВ, а каналы для него открываются при значениимембранного потенциала -55 мВ, поэтому ионы натрия поступают в клетку с высокой
скоростью: до 107 ионов/с через одиночный канал.
Плотность натриевых каналов варьирует от 1 до 50 на одном квадратном микрометре.
В результате этого за 0,2 – 0, 5 мс значение мембранного потенциала из отрицательного (-55
мВ) становится положительным (около +30 мВ), хотя и не достигает значения равновесного
натриевого потенциала.
Такая стремительная деполяризация является саморегенерирующей: чем больше входит
натрия в клетку и чем больше сдвиг мембранного потенциала, тем больше открывается
натриевых каналов и тогда ещё больше натрия входит в клетку:
6.
По мере приближения величины мембранногопотенциала к значению равновесного натриевого
потенциала движущая сила для ионов натрия слабеет,
но одновременно растёт движущая сила, заставляющая
выходить из клетки ионы калия, каналы для которых
постоянно открыты.
Когда мембранный потенциал становится
положительным, натриевые потенциалзависимые
каналы закрываются, а ток калия из клетки резко
увеличивается.
В связи с этим происходит реполяризация, т.е.
восстановление первоначального значения
мембранного потенциала (иногда выходной ток калия
приводит даже к кратковременной следовой
гиперполяризации).
7.
Две фазы потенциала действия – деполяризация и реполяризация образуют пик или спайкпотенциала действия (Рис. 4.6). Само открытие натриевых каналов происходит необычайно
быстро, в течение не более, чем 10 микросекунд (т.е. миллионных долей секунды),
открытыми они остаются несколько миллисекунд, затем быстро закрываются, причём на
некоторое время конформация канального белка становится такой, что его невозможно
активировать, а значит и открыть каналы.
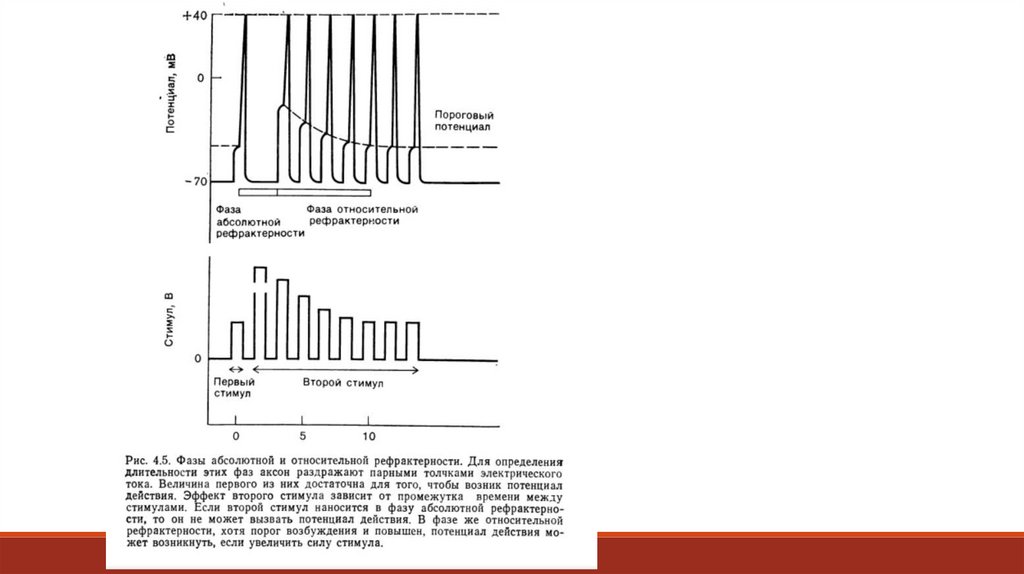
Это состояние называется рефрактерностью, около 1 мс она абсолютна, а затем
относительна: при абсолютной рефрактерности нельзя открыть каналы никаким
действием, при относительной их нельзя активировать пороговой деполяризацией, но
можно надпороговой.
Общая продолжительность рефрактерного состояния определяет максимальную частоту
возбуждения нейрона. Например, если рефрактерный период продолжается 2 мс, то за 1с
нейрон может возбуждаться максимум 500 раз (1 с = 1000 мс: 2мс = 500).
8.
Некоторые нейроны могут возбуждаться чаще, чем 500/с, другие – реже: всоответствии с этим первые можно назвать более лабильными, чем вторые.
Проблему лабильности или функциональной подвижности клеток в конце XIX –
начале ХХ века исследовал российский физиолог Н.Е.Введенский, он же ввёл в
употребление понятие о мере лабильности, как о наибольшем числе
электрических осцилляций, которое может воспроизвести за секунду нерв или
мышца.
Так, например, нерв, по данным Введенского, способен возбуждаться до 500/с, а
мышца – лишь до 200/с, т.е. нерв является более лабильным объектом, чем
мышца.
9.
10.
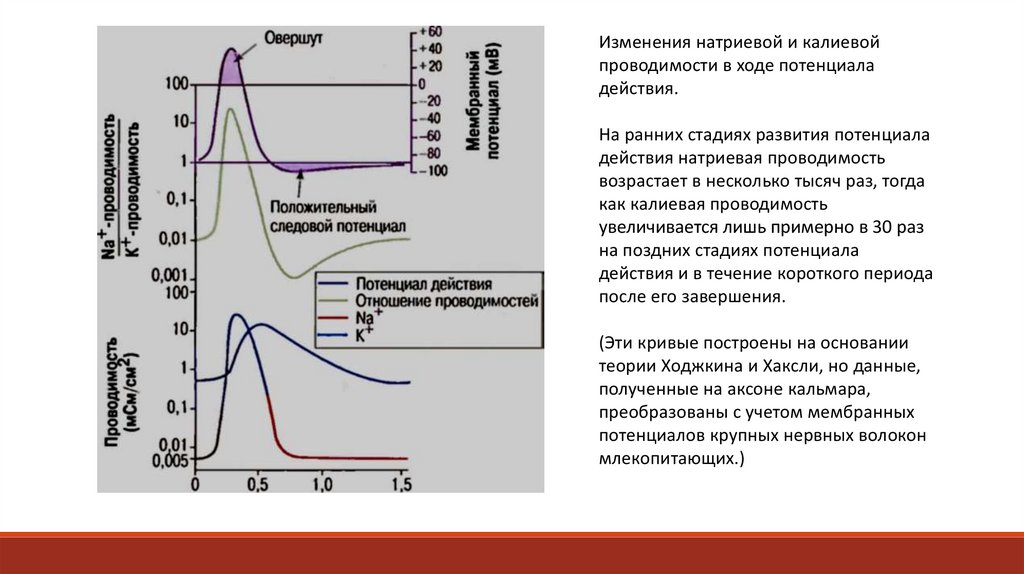
Изменения натриевой и калиевойпроводимости в ходе потенциала
действия.
На ранних стадиях развития потенциала
действия натриевая проводимость
возрастает в несколько тысяч раз, тогда
как калиевая проводимость
увеличивается лишь примерно в 30 раз
на поздних стадиях потенциала
действия и в течение короткого периода
после его завершения.
(Эти кривые построены на основании
теории Ходжкина и Хаксли, но данные,
полученные на аксоне кальмара,
преобразованы с учетом мембранных
потенциалов крупных нервных волокон
млекопитающих.)
11.
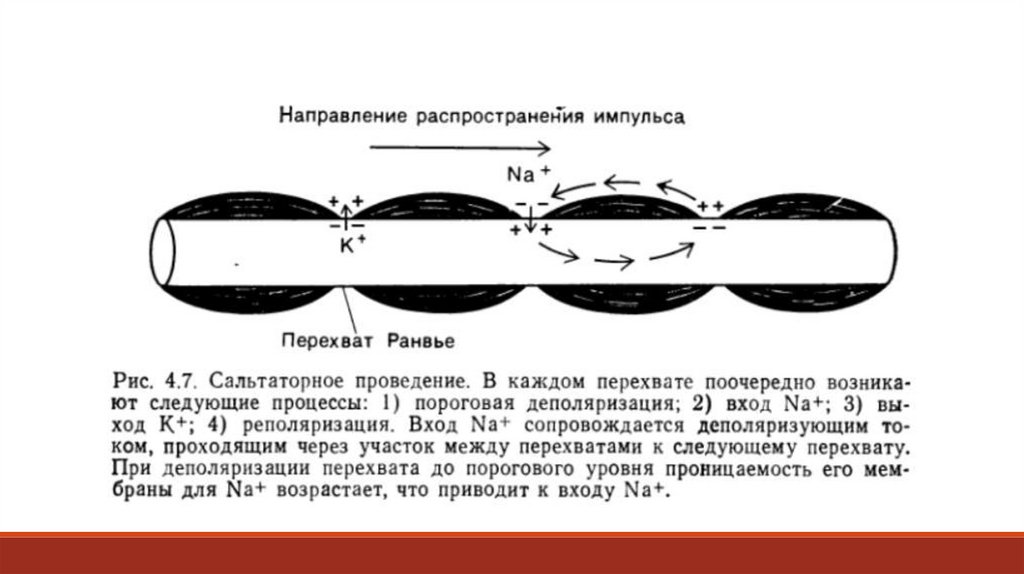
4.8. Механизм проведения потенциалов действияЧем более сложные задачи решает мозг, тем большее количество нейронов ему необходимо. Однако вся масса
нейронов должна поместиться в ограниченном черепом и позвоночным каналом пространстве, а потому
нервные клетки должны быть небольшими, а их отростки – достаточно тонкими. Но, как известно, чем тоньше и
длиннее проводник, тем большее сопротивление он окажет распространяющемуся по нему току.
Действующее напряжение в нейроне (V) не может быть больше, чем амплитуда потенциала действия, т.е
приблизительно 100-120 мВ, а ток (I), согласно закону Ома, прямо пропорционален напряжению и обратно
пропорционален сопротивлению: I = V/R Из этого следует, что потенциал действия обычным для проведения
электричества способом не может распространяться далеко. Очень тонкая мембрана аксона, окруженная
электропроводной средой, имеет очень высокую ёмкость, что тормозит распространение электрического
сигнала.
Если сказать проще: тонкий цитоплазматический отросток –
это очень плохой проводник. Но, несмотря на это,
потенциалы действия распространяются по аксону с
высокой скоростью, достигающей 100 м/с. Как это
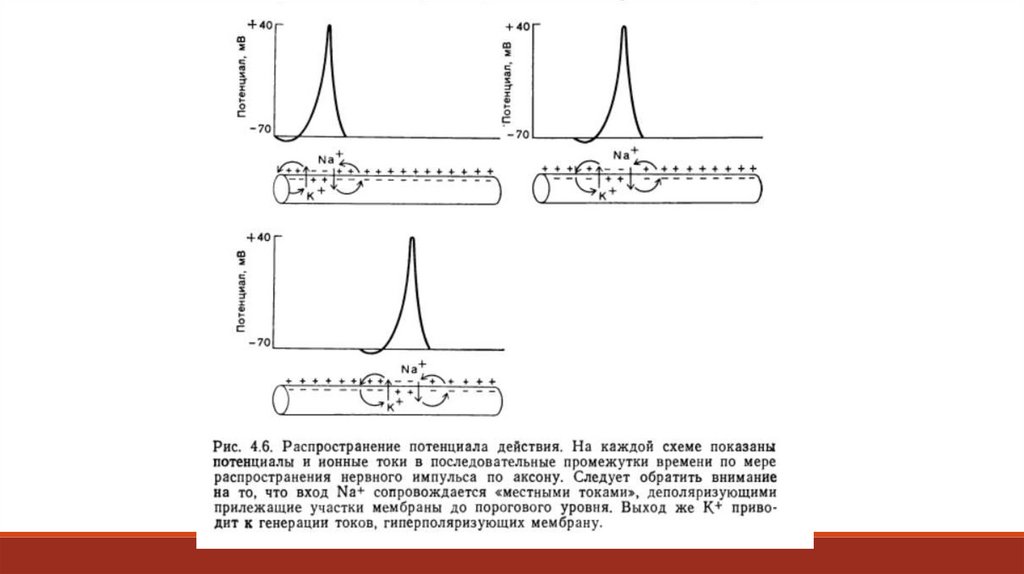
происходит? Когда в возбуждённом участке мембраны
повышается натриевая проницаемость и возникает
потенциал действия, начинается электротоническое
распространение положительных зарядов к
невозбуждённому участку – этот процесс представляет
собой круговой ток (Рис. 4.7).
12.
13.
14.
Такой ток деполяризует ещё не возбуждавшийся соседний участок, а когда эта деполяризация достигнет порога– возникнет потенциал действия. Теперь уже этот участок становится источником кругового тока, действующего
на следующую область мембраны, теперь уже в этой области возникнет потенциал действия, все параметры
которого будут стандартными для данного типа нейрона. Вслед за повышением натриевой проницаемости в
процессе формирования потенциала действия усиливается ток калия из клетки. Вместе с калием из клетки
уходят положительные заряды и восстанавливается прежнее значение мембранного потенциала. При любой
длине аксона амплитуда потенциалов действия повсюду оказывается неизменной, поскольку в каждом
отдельном участке аксона они формируется фактически заново. В физиологическом смысле это важно потому,
что постоянство сигнала означает передачу информации по аксону без искажений. В миелинизированных
аксонах круговой ток распространяется к соседнему перехвату, где и возникает потенциал действия. Плотность
натриевых каналов в перехватах Ранвье гораздо выше, чем в обычной немиелинизированной мембране и
пришедший сюда электротонически круговой ток легко деполяризует перехват до порогового значения.
Появившийся вследствие этого потенциал действия служит источником кругового тока для следующего
перехвата. Проведение возбуждения в нерве или мышце можно зарегистрировать с помощью внеклеточных
электродов, приложенных к двум различным точкам их поверхности и соединённым с регистрирующей
аппаратурой.
При распространении потенциала действия мембрана поочерёдно деполяризуется сначала под ближним к
источнику возбуждения электродом, а затем – под дальним. И в том, и в другом случае между электродами
регистрируется разность потенциалов, поскольку один из них будет находиться на деполяризованном, а значит
электроотрицательном снаружи мембраны участке, а второй – в интактной электроположительной точке, где
возбуждение ещё не началось, либо уже закончилось. Регистрация проходящих по мембране потенциалов
действия с помощью двух электродов называется биполярной.
При таком способе регистрируются две фазы потенциала действия: положительная и отрицательная. Если
расположенный под одним из электродов участок сделать невозбудимым (для этого можно подействовать на
него каким-либо анестезирующим веществом, например, новокаином), то останется только одна фаза
потенциала действия. Такое отведение называется униполярным (или монополярным). При некоторых
аутоиммунных и вирусных заболеваниях миелиновая оболочка разрушается, что приводит к многочисленным
неврологическим нарушениям, вплоть до полной утраты некоторых функций; при этом может нарушаться и
эмоциональная деятельность, и интеллект. Примером демиелинизирующих заболеваний может служить
рассеянный склероз.
15.
РезюмеВозникновение электрических сигналов связано со свойствами клеточной мембраны.
Мембранные насосы создают концентрационные градиенты ионов. Открытые в покое
ионные каналы для калия позволяют ему выходить из клетки и, тем самым, создавать
мембранный потенциал покоя, близкий к равновесному потенциалу для калия. В случае
его уменьшения до порогового значения открываются потенциалзависимые каналы для
натрия и происходит саморегенерирующая деполяризация, значение мембранного
потенциала становится положительным, Это вызывает закрытие натриевых каналов,
которые на время инактивируются. Выходящий ток ионов калия восстанавливает прежнее
значение мембранного потенциала. Возникновение потенциала действия вызывает
появление кругового электрического тока, который деполяризует соседний участок
мембраны до порогового значения. В связи с этим потенциал действия распространяется
по аксону без уменьшения амплитуды.
16.
Вопросы для самоконтроля46. Концентрация каких ионов в клетке
значительно больше, чем во внеклеточной
жидкости?
А. Натрия;
Б. Калия;
В. Кальция;
Г. Хлора;
Д. Магния.
47. Какие ионные каналы при
физиологическом покое клетки открыты?
А. Для всех катионов;
Б. Для анионов;
В. Для натрия;
Г. Для калия;
В. Для кальция.
17.
48. Какова величина равновесногопотенциала мембраны гигантского аксона
кальмара для ионов калия?
49. Почему натрий-калиевый насос
считается электрогенным? А. Он расходует
энергию АТФ;
А. +55 мВ;
Б. Он создаёт концентрационный градиент
калия;
Б. +25-30 мВ;
В. = 0;
Г. -60 мВ;
Д. -75 мВ.
В. Он выносит из клетки натрий; Г. За один
цикл он удаляет из клетки положительный
заряд;
Д. Он обеспечивает симпорт глюкозы и
аминокислот.
18.
50. Вхождению в клетку каких ионовпрепятствует электрическое поле между
внутренней и наружной поверхностями
мембраны?
51. Через каналы какого типа
диффундируют ионы калия, когда клетка
находится в состоянии физиологического
покоя?
А. Калия;
А. Потенциалзависимые;
Б. Натрия;
Б. Хемозависимые;
В. Хлора;
В. Потенциалзависимые и хемозависимые;
Г. Кальция;
Г. Управляемые механически;
Д. Всех катионов.
Д. Пассивные.
19.
52. Что из перечисленного ниже характернодля состояния рефрактерности?
А. Активированное состояние
потенциалзависимых каналов;
Б. Инактивированное состояние
потенциалзависимых каналов;
В. Открытое состояние потенциалзависимых
каналов;
Г. Закрытое состояние потенциалзависимых
каналов;
Д. Увеличение пропускной способности
потенциалзависимых каналов.
53. Какое из перечисленных ниже веществ
является блокатором ионных каналов для
калия?
А. Тетраэтиламмоний;
Б. Тетродотоксин;
В. Батрахотоксин;
Г. Кураре;
Д. a-Бунгаротоксин.
20.
54. Каким должен быть наименьшийдеполяризующий сдвиг, если мембранный
потенциал равен -69 мВ, а критический
уровень деполяризации – -56 мВ?
55. Если рефрактерный период нейрона
продолжается 3 мс, то с какой
максимальной частотой он может
возбуждаться?
А. 6 мВ;
А. 555 Гц;
Б. 9 мВ;
Б. 444 Гц;
В. 11 мВ;
В. 333 Гц;
Г. 13 мВ;
Г. 222 Гц;
Д. 15 мВ.
Д. 111 Гц.
21.
56. Для какого перемещения ионов черезклеточную мембрану, находящейся в покое
клетки, необходима энергия?
А. Кальция в клетку;
Б. Натрия в клетку;
В. Хлора в клетку;
Г. Калия из клетки;
Д. Кальция из клетки.
57. Какое перемещение ионов происходит
только путём диффузии?
А. Натрия из клетки;
Б. Калия из клетки;
В. Кальция из клетки;
Г. Калия в клетку;
Д. Глюкозы в клетку.
22.
58. Что заставляет закрытьсяоткрывающиеся при возбуждении
потенциалзависимые каналы для натрия?
А. Процесс реполяризации;
Б. Восстановление исходного значения
мембранного потенциала;
В. Установление положительного значения
мембранного потенциала;
Г. Достижение критического уровня
деполяризации;
Д. Возникновение гиперполяризации.
59. К каким последствиям должно привести
повышение мембранной проницаемости
для хлора при реальном значении
мембранного потенциала -55 мВ?
А. Уменьшение мембранного потенциала;
Б. Гиперполяризация;
В. Деполяризация;
Г. Значение мембранного потенциала не
изменится;
Д. Возникнет потенциал действия.
23.
60. Каждый потенциал действия образуютдве, последовательно сменяющие друг
друга фазы – это:
А. Гиперполяризация-деполяризация;
Б. Деполяризация-реполяризация;
В. Гиперполяризация-реполяризация;
Г. Реполяризация- деполяризация;
Д. Реполяризация – восстановление
первоначального значения мембранного
потенциала.