
























































 Биология
БиологияПохожие презентации:


")

")





Межклеточные и внутриклеточные сигнальные вещества
1.
ЛекцияМежклеточные и
внутриклеточные сигнальные
вещества
2.
Группы межклеточных сигнальных веществ
Гормонпродуцирующие структуры и гормоны, их виды.
Общая схема действия гидрофильных гормонов.
Общая схема действия гидрофобных гормонов.
Гистогормоны. Определение и классификация.
Некоторые интерлейкины и факторы роста. Нейромедиаторы.
Нейромодуляторы.
Ц АМФ-опосредованные пути. Компоненты путей.
Стимуляция распада углеводов и жиров.
Спазмолитическое действие симпатомиметиков.
Аденилатциклазная система эпителия кишечника.
Ц ГМФ- опосредованные пути. Гуанилатциклазная система в
фоторецепторных клетках сетчатки глаза.
Ц ГМФ- и NOопосредованные пути.
Образование NO и NO-синтетаза. Изоформы NO-синтетазы.
Сосудорасширяющее действие NO. NO в нервной системе.
Пути, опосредованные липидами и ионами кальция.
Пути, опосредованные белком Ras.
Пути, не содержащие вторичного мессенджера.
3.
Все межклеточные сигнальные вещества можно разделить на тригруппы:
• Гормоны – регуляторы, образуемые эндокринными клетками и
попадающие к клеткам-мишеням через кровь;
• Нейромедиаторы – соединения, передающие сигнал в синапсах от
пресинаптического окончания к постсинаптическому;
• Гистогормоны (так называемые цитокины и факторы роста) –
регуляторы, выделяемые неэндокринными клетками во
внесосудистое пространство и обладающие поэтому местным
действием.
4.
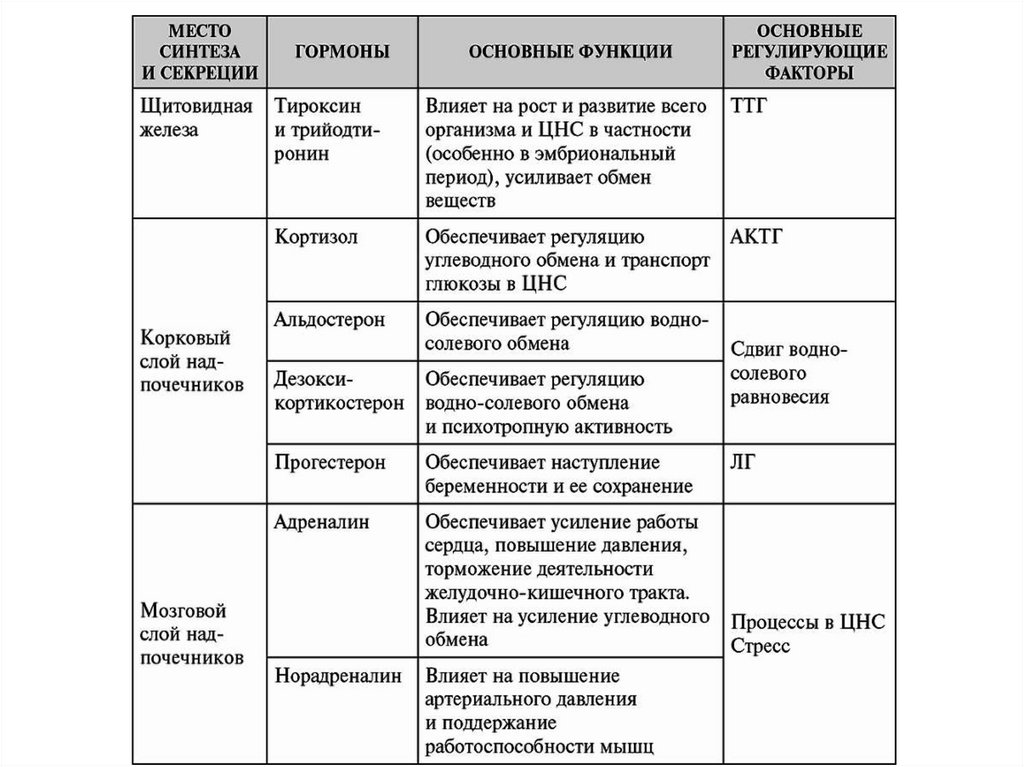
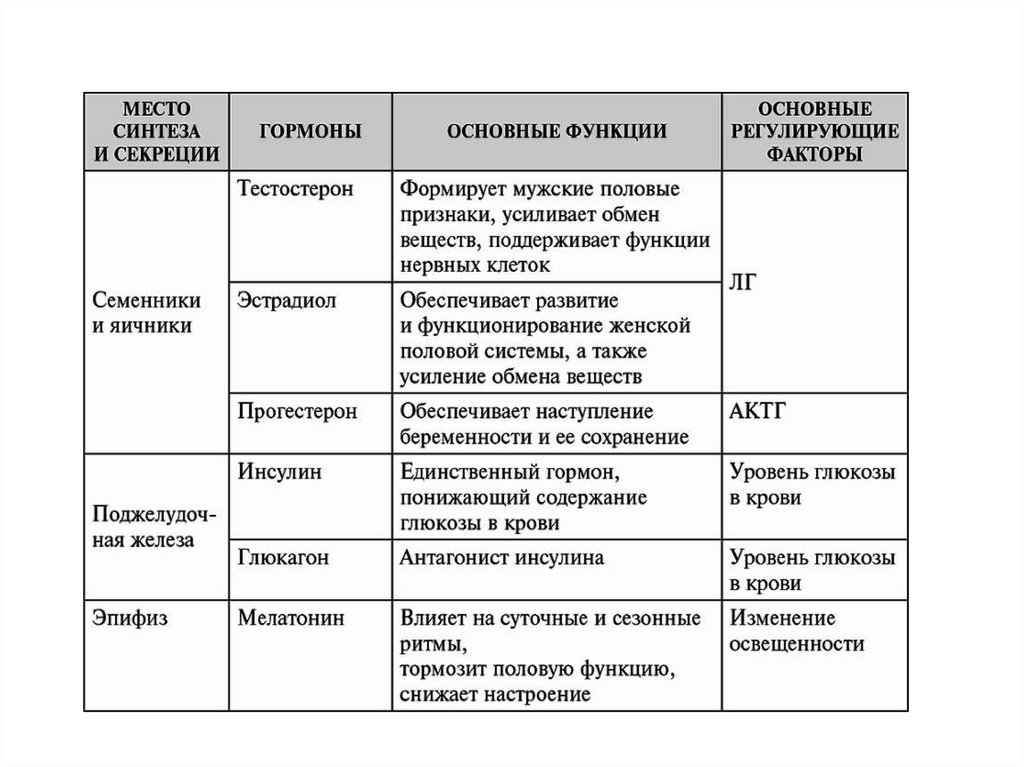
ГормоныМесто образования и биологическое действие
Все гормонпродуцирующие структуры делятся на четыре типа:
1)
2)
3)
4)
Центральные эндокринные органы (гипоталамус, гипофиз,
эпифиз);
Периферические эндокринные железы (щитовидная,
паращитовидные, надпочечники);
Органы, объединяющие эндокринные и неэндокринные
функции (поджелудочная железа, почки, тимус, гонады,
плацента, сердце);
Одиночные гормонпродуцирующие клетки, составляющие
диффузную эндокринную систему (эндокринные клетки в
разных отделах нервной, пищеварительной и дыхательной
систем)
5.
Наряду с термином «гормон» в последнеевремя используются также понятия
гормоноподобные и биологически
активные вещества (БАВ), гормоны
местного действия.
Так, некоторые из них синтезируются
настолько близко к своим органам-мишеням,
что могут достигать их диффузией, не
попадая в кровоток. Клетки,
вырабатывающие такие вещества: называют
паракринными.
Когда рассматривается выработка веществ в
мозговом веществе надпочечников, их
обычно называют гормонами, если речь идет
об их образовании и выделении
симпатическими окончаниями, их называют
медиаторами.
6.
7.
8.
9.
Общие свойства гормонов:1. Строгая специфичность физиологического действия.
2. Высокая биологическая активность: гормоны оказывают свое
физиологическое действие в чрезвычайно малых дозах.
3. Дистантный характер действия: клетки-мишени располагаются обычно
далеко от места образования гормона.
4. Многие гормоны (стероидные и производные аминокислот) не имеют
видовой специфичности.
5. Генерализованность действия.
6. Пролонгированность действия.
10.
Классификация, свойства, механизмы действия гормонов иБАВ
По химической структуре гормоны и БАВ классифицируют на:
1. Производные аминокислот:
производные тирозина: тироксин, трийодтиронин, дофамин, адреналин,
норадреналин; производные триптофана: мелатонин, серотонин;
производные гистидина: гистамин.
2. Белково-пептидные гормоны:
полипептиды: глюкагон, кортикотропин, меланотропин, вазорессин,
окситоцин, пептидные гормоны желудка и кишечника;
простые белки (протеины): инсулин, соматотропин, пролактин, паратгормон,
кальцитонин;
сложные белки (гликопротеиды): тиреотропин, фоллитропин, лютропин.
3. Стероидные гормоны: кортикостероиды (альдостерон, кортизол,
кортикостерон); половые гормоны: андрогены (тестостерон), эстрогены и
прогестерон.
4. Производные жирных кислот: арахидоновая кислота и ее производные:
простагландины, простациклины, тромбоксаны, лейкотриены
11.
По полярным свойствам гормоны классифицируют на две группы:• Полярные, или гидрофильные – белки, пептиды и производные
аминокислот (кроме тиреоидных гормонов);
• Неполярные, или гидрофобные гормоны – стероиды (половые и
гормоны коры надпочечников), производные жирных кислот
(простагландины) и тиреоидные гормоны.
12.
С принадлежностью гормонов к определенной группе практическиоднозначно связан механизм его действия на клетку-мишень:
• Гидрофильные гормоны не способны проникать через плазмолемму,
и должен существовать специальный механизм для восприятия
сигнала и передачи его на эффекторные структуры;
• Гидрофобные гормоны проходят через мембраны клетки и обычно
непосредственно достигают регулируемого объекта.
13.
Механизмы действия гормоновСуществуют два основных механизма действия гормонов на уровне
клетки:
- реализация эффекта с наружной поверхности клеточной мембраны;
- реализация эффекта после проникновения гормона внутрь клетки.
Рецепторы в зависимости от локализации в клетки делят на:
- Рецепторы плазматической мембраны;
- Внутриклеточные рецепторы.
14.
По количеству основных вторичных посредников и механизмам ихобразования гормоны, связывающиеся с поверхностными
рецепторами, подразделяются на 4 группы:
• Действующие по аденилатциклазному механизму (вторичный
посредник цАМФ) – α2 и β-адренергические катехоламины,
АКТГ, вазопрессин, глюкагон, кальцитонин, паратгормон,
ангиотензин II, тиреотропин, кортиколиберин, ФСГ;
• Действующие по гуанилатциклазному механизму (вторичный
посредник цГМФ) – атриальные натрийуретические факторы;
• Действующие по Ca2+/фосфатидилинозитоловому механизму
(вторичный посредник Ca2+ и/или фосфатидилинозитолы) – α1
– адренергические катехоламины, окситоцин, вазопрессин,
ангиотензин II, гастрин, тиреолиберин, гонадолиберин,
соматолиберин;
• Действующие по киназному механизму (вторичный посредник
– киназный каскад) – инсулин, гормон роста и пролактин.
15.
Протеинкиназы – специальные регуляторные ферменты, способныефосфорилировать строго определенные белки по определенным
аминокислотным остаткам.
Фосфорилирование (и дефосфорилирование под действием
протеинфосфатаз) – один из наиболее универсальных способов
регуляции активности белков – как структурных, так и ферментов.
16.
Благодаря наличию в клетках-мишеняхрецепторных белков диффузия гормона
в клетку оказывается значительно более
интенсивной.
Влияние на синтез белков, которое
реализуется на генном уровне
(относится к стероидным и тиреоидным
гормонам).
Простагландины проникают через
плазмалемму клеток, но действуют по
схеме гидрофильных гормонов, т.е.
влияют на активность определенных
протеинкиназ.
17.
Гистогормоны (аукоиды)Особенности:
• Распространяются не с кровью, а путем диффузии в межклеточном
пространстве;
• Оказывают лишь местное действие – на расположенные недалеко
клетки-мишени (паракринное действие) или даже на саму клеткупродуцент (аутокринное действие).
Возможно также интракринное действие: регуляторное вещество
действует на клетку, в которой оно образовалось, не выделяясь во
внешнюю среду (в этом случае вещество называется не гистогормоном, а
внутриклеточным медиатором, или мессенджером).
Иногда вещество-регулятор обладает одновременно и интракринным, и
паракринным действием (простагландины, NO).
Гистогормоны делят на:
• Цитокины;
• Факторы роста
18.
Цитокины участвуют (в качестве последовательно выделяемых клеткамистимуляторов) в воспалительных, иммунных и других защитных
реакциях.
Вырабатываются обычно не постоянно.
А) Интерлейкины (ИЛ) – выделяются активированными лейкоцитами и
обеспечивают взаимодействие клеток (н., ИЛ-1,2,4,5; ФНО);
Б) Интерфероны – небольшие сигнальные белки, которые выделяются
клетками, инфицированными вирусами. Действуя на клетки-продуценты
и на соседние клетки, интерфероны ограничивают белковый синтез, тем
самым предупреждается образование в клетках новых вирусных частиц.
В) Малые цитокины – характеризуются тем, что имеют относительно
короткие пептидные цепи (ИЛ-8).
Г) Колониестимулирующие факторы (КСФ) – белковые стимуляторы
развития гемопоэтических клеток.
19.
Факторы роста – белки, стимулирующие (либо ингибирующие) делениеи развитие определенных клеток.
Среди данных факторов – ЭФР (эпидермальный фактор роста), НФР
(фактор роста нейронов), ФРФ (фактор роста фибробластов) и др.
20.
Нейромедиаторы и нейромодуляторыНейромодуляторы – это вещества, которые, не будучи способны
самостоятельно передавать в синапсах сигнал (возбуждающий или
тормозящий), тем не менее влияют на передачу такового истинными
медиаторами, а именно облегчают или затрудняют эту передачу (в
основном, подобную функцию выполняют нейропептиды головного мозга).
Если в одном синапсе обнаруживаются несколько полноценных
нейромедиаторов, их называют ко-нейромедиаторами.
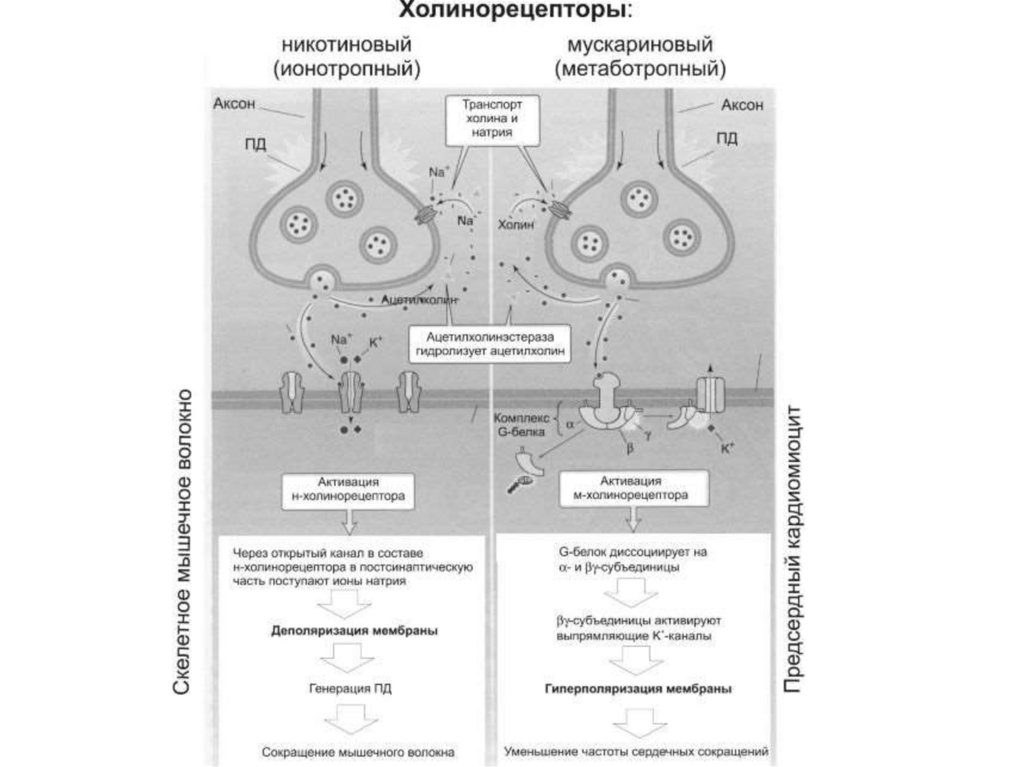
Рецепторы нейромедиаторов по механизму дальнейшей передачи сигнала
подразделяются на 2 группы:
- Ионотропные (быстродействующие) рецепторы служат одновременно
ионными каналами, которые открываются при связывании медиатора с
рецепторной частью белка (н-холинорецепторы);
- Метаботропные (медленнодействующие) рецепторы – здесь сигнал
передается по той же принципиальной схеме, что и в случае
гидрофильных гормонов, т.е. механизм включает внутриклеточные
посредники.
21.
22.
Основные нейромедиаторы периферической нервной системы:• Ацетилхолин;
• Норадреналин.
Нейромедиаторы центральной нервной системы:
• Серотонин;
• Гистамин;
• Глутаминовая кислота;
• Аспарагиновая кислота;
• Ацетилхолин;
• Дофамин;
• Норадреналин;
• ГАМК;
• Глицин;
• Таурин;
• Аденозин
23.
Пептидные нейромедиаторы:• Люлиберин;
• ВИП (вазоактивный интестинальный пептид);
• Соматостатин;
• Вещество Р;
• Нейропептид Y.
24.
Плазматическая мембрана осуществляетвзаимодействие клеток с окружающей средой и друг с
другом, именно на ее поверхности в большинстве
случаев происходит преобразование внешних
сигналов во внутриклеточные.
Передача сигнала или сигнальная трансдукция,
сигнализация (от англ. transduction, signaling)
подразумевает процесс, при помощи которого клетка
превращает один тип сигнала или стимула в другой.
25.
Внешний сигнальный агент, называемый первичнымпосредником, как правило, не проникает внутрь клетки, а
специфически взаимодействует с рецепторами клеточной
мембраны (нейромедиаторы, гормоны, цитокинины,
факторы роста, нейромодуляторы и т.д.).
Формирование комплекса рецептор-лиганд обеспечивает
проведение специфического трансмембранного сигнала
благодаря образованию вторичных посредников –
мессенджеров (от англ. messenger) и активации
посредниками белков-мишеней (фосфорилирование,
дефосфорилирование).
26.
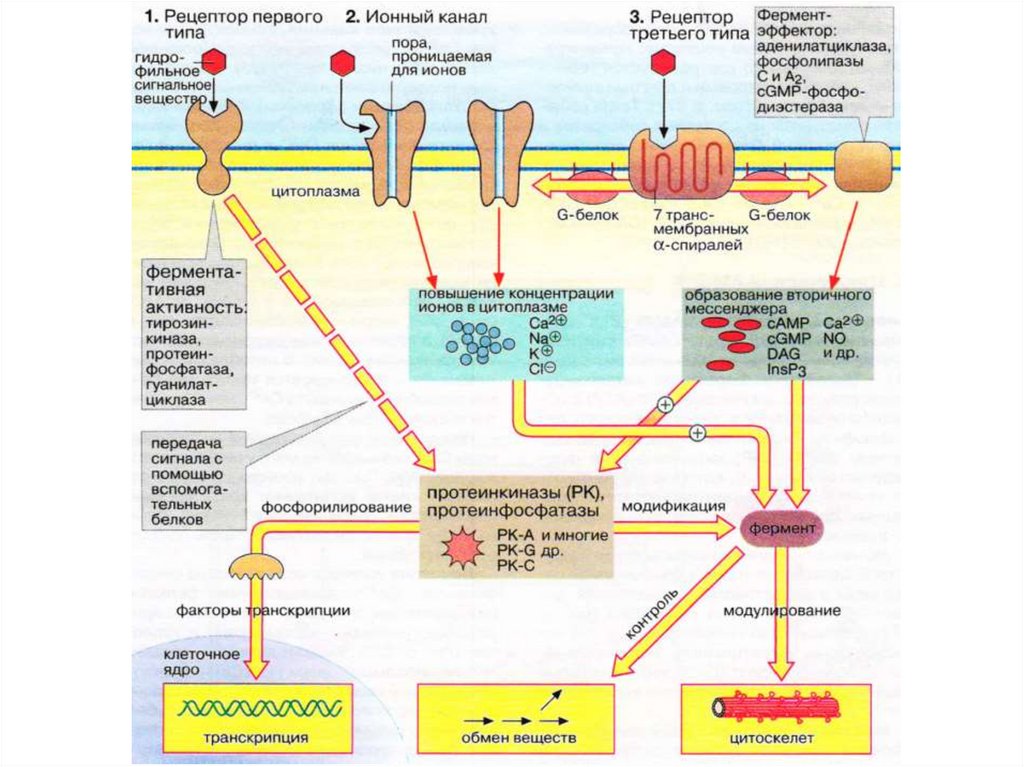
Несмотря на огромное разнообразие внешних сигналов,рецепторов и механизмов передачи сигнала, в клетке
существуют несколько универсальных сигнальных систем.
В роли вторичных посредников выступают малые
молекулы и ионы:
•ионы кальция (Ca2+);
•циклические нуклеотиды (цГМФ, цАМФ);
•липофильные молекулы (например, диацилглицерол,
арахидоновая кислота);
•газы (NO, CO, H2S).
27.
Вторичные посредники характеризуются следующимисвойствами:
• имеют небольшую молекулярную массу,
• с высокой скоростью диффундируют внутри клетки,
• быстро расщепляются и\или удаляются из цитоплазмы.
28.
Гуанин-нуклеотид-связывающие белки ирецепторы, сопряженные с G-белками
Существуют 3 класса рецепторов плазматической мембраны
клетки:
• рецепторы, сопряженные с G-белками или
метаботропные;
• рецепторы, связанные с ионными каналами или
ионотропные;
• рецепторы, обладающие собственной ферментативной
активностью – протеинтирозинкиназы, рецепторы
факторов роста тромбоцитов, эпидермальных и нервных
клеток.
29.
30.
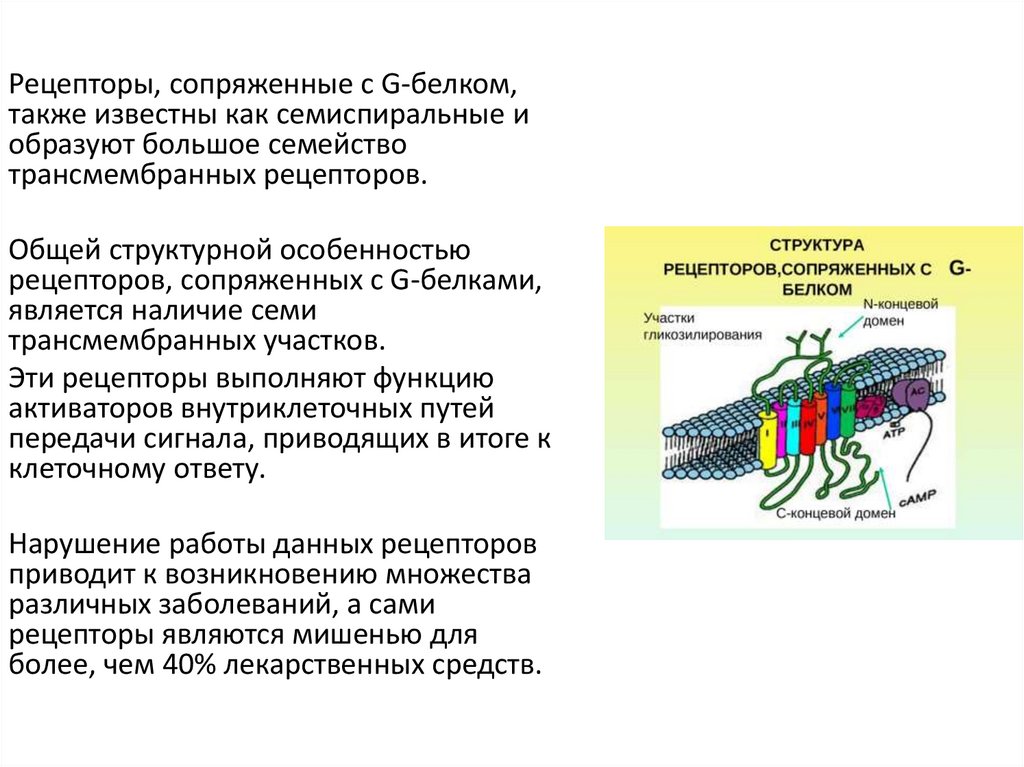
Рецепторы, сопряженные с G-белком,также известны как семиспиральные и
образуют большое семейство
трансмембранных рецепторов.
Общей структурной особенностью
рецепторов, сопряженных с G-белками,
является наличие семи
трансмембранных участков.
Эти рецепторы выполняют функцию
активаторов внутриклеточных путей
передачи сигнала, приводящих в итоге к
клеточному ответу.
Нарушение работы данных рецепторов
приводит к возникновению множества
различных заболеваний, а сами
рецепторы являются мишенью для
более, чем 40% лекарственных средств.
31.
В зависимости от сайта связывании лиганда и природы лигандарецепторы, сопряженные с G-белками, разделены на три
основные класса - A, B, C.
Рецепторы класса А и В связывают низкомолекулярные лиганды
и пептиды в трансмембранном участке,
рецепторы класса С связывают низкомолекулярные лиганды в
области внеклеточных петель, соединяющих трансмембранные
домены.
32.
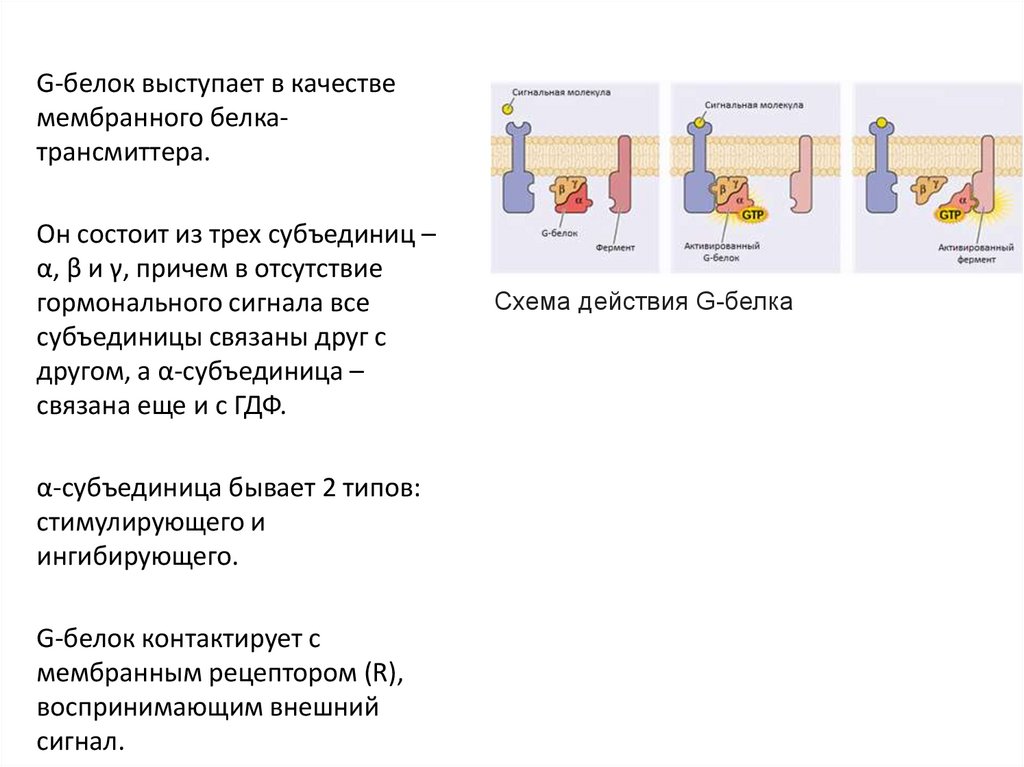
G-белок выступает в качествемембранного белкатрансмиттера.
Он состоит из трех субъединиц –
α, β и γ, причем в отсутствие
гормонального сигнала все
субъединицы связаны друг с
другом, а α-субъединица –
связана еще и с ГДФ.
α-субъединица бывает 2 типов:
стимулирующего и
ингибирующего.
G-белок контактирует с
мембранным рецептором (R),
воспринимающим внешний
сигнал.
Схема действия G-белка
33.
Пути передачи сигнала с участием рецепторов, сопряженных с G-белками, включаютследующие этапы:
1.
2.
3.
4.
5.
6.
Лиганд связывается с мембранным рецептором. В это
время неактивный G-белок связан с ГДФ.
Связанный с лигандом рецептор, взаимодействуя с Gбелком, активирует его и G-белок связывает ГТФ
(концентрация ГТФ в клетке ~ 0.1 мМ).
Активированный G-белок взаимодействует с одним или
несколькими внутриклеточными ферментами аденилатциклазой, гуанилатциклазой, фосфолипазами
С, А, Д, изменяя их активность. Кроме того, G-белок
может напрямую модулировать функционирование
ионных каналов.
Изменение активности внутриклеточного фермента
приводит к изменению внутриклеточного уровня одного
или нескольких вторичных посредников, таких как
цАМФ, цГМФ, Са2+, инозитолтрифосфат,
диацилглицерол и др.
Увеличение или уменьшение концентрации вторичного
посредника влияет на активность одной или нескольких
зависимых от него протеинкиназ и/или ионных каналов.
Изменяется уровень фосфорилирования белковмишеней и это обуславливает конечный ответ клетки.
АК – арахидоновая кислота, NO –
оксид азота (II), ионы Са,
циклические нуклеотиды.
34.
35.
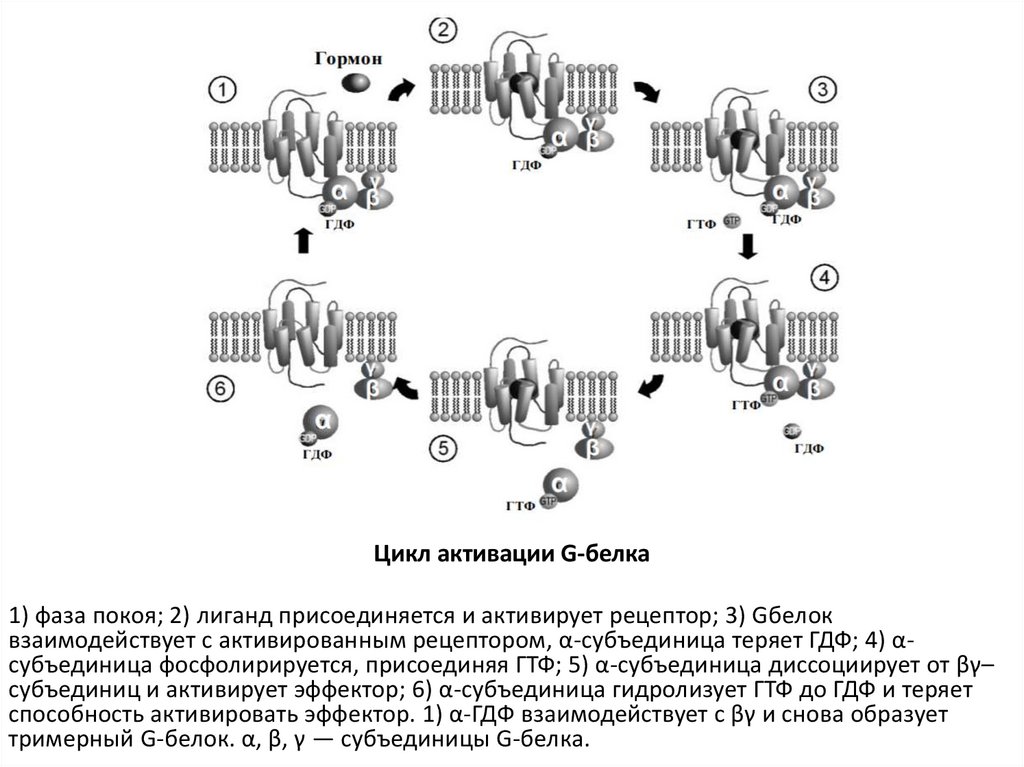
Цикл активации G-белка1) фаза покоя; 2) лиганд присоединяется и активирует рецептор; 3) Gбелок
взаимодействует с активированным рецептором, α-субъединица теряет ГДФ; 4) αсубъединица фосфолирируется, присоединяя ГТФ; 5) α-субъединица диссоциирует от βγ–
субъединиц и активирует эффектор; 6) α-субъединица гидролизует ГТФ до ГДФ и теряет
способность активировать эффектор. 1) α-ГДФ взаимодействует с βγ и снова образует
тримерный G-белок. α, β, γ — субъединицы G-белка.
36.
Связывание агониста (гормона, нейромедиатора и др.) с соответствующимрецептором приводит к белок-белковому взаимодействию между рецептором и Gбелком и ускоряет диссоциацию ГДФ.
В результате образуется короткоживущий комплекс: агонист-рецептор-G-белок,
несвязанный с нуклеотидом.
Связывание с этим комплексом молекулы ГТФ снижает сродство рецептора к G-белку,
что приводит к диссоциации комплекса и высвобождению рецептора
Потенциально рецептор может активировать большое количество молекул G-белка,
обеспечивая, таким образом, высокий коэффициент усиления внеклеточного сигнала
на данном этапе.
Активированная α-субъединица G-белка диссоциирует от βγ-субъединиц и вступает
во взаимодействие с соответствующим эффектором, оказывая на него активирующее
или ингибирующее воздействие.
Взаимодействие с эффектором длится до тех пор, пока α-субъединица, являющаяся
ГТФ-азой, удерживает ГТФ. После гидролиза ГТФ до ГДФ α-субъединица снова
меняет свою конформацию и теряет способность активировать эффектор.
После этого комплекс α-ГДФ взаимодействует с βγ и вновь образует тримерный Gбелок, завершая таким образом цикл
37.
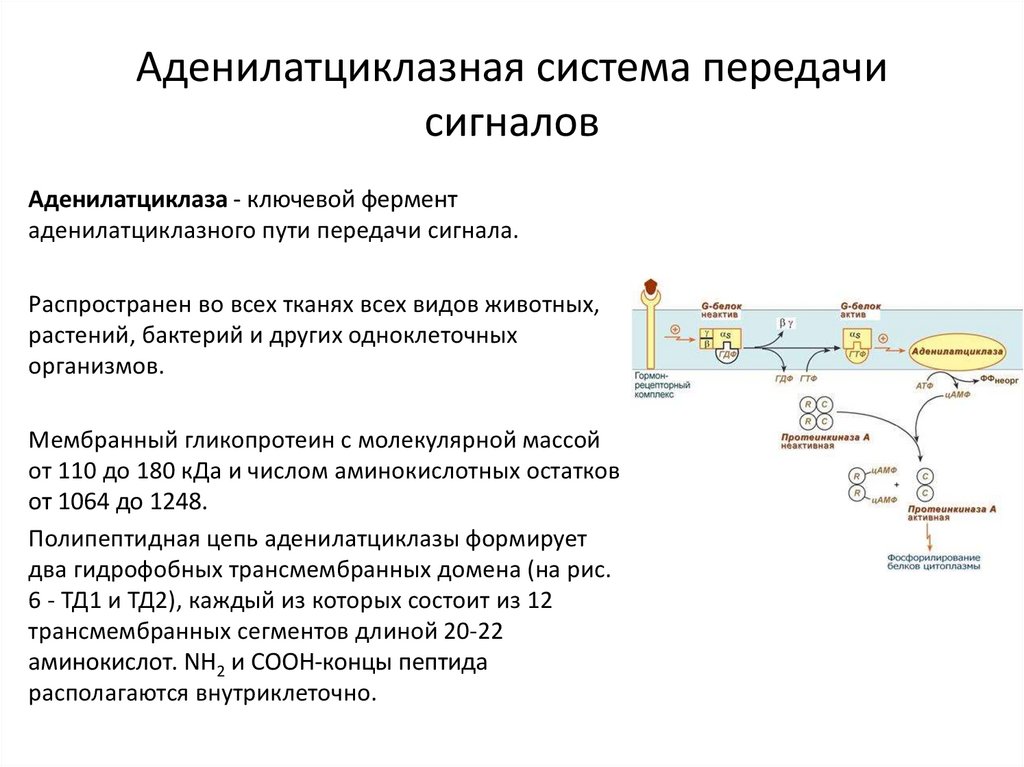
Аденилатциклазная система передачисигналов
Аденилатциклаза - ключевой фермент
аденилатциклазного пути передачи сигнала.
Распространен во всех тканях всех видов животных,
растений, бактерий и других одноклеточных
организмов.
Мембранный гликопротеин с молекулярной массой
от 110 до 180 кДа и числом аминокислотных остатков
от 1064 до 1248.
Полипептидная цепь аденилатциклазы формирует
два гидрофобных трансмембранных домена (на рис.
6 - ТД1 и ТД2), каждый из которых состоит из 12
трансмембранных сегментов длиной 20-22
аминокислот. NH2 и СOOH-концы пептида
располагаются внутриклеточно.
38.
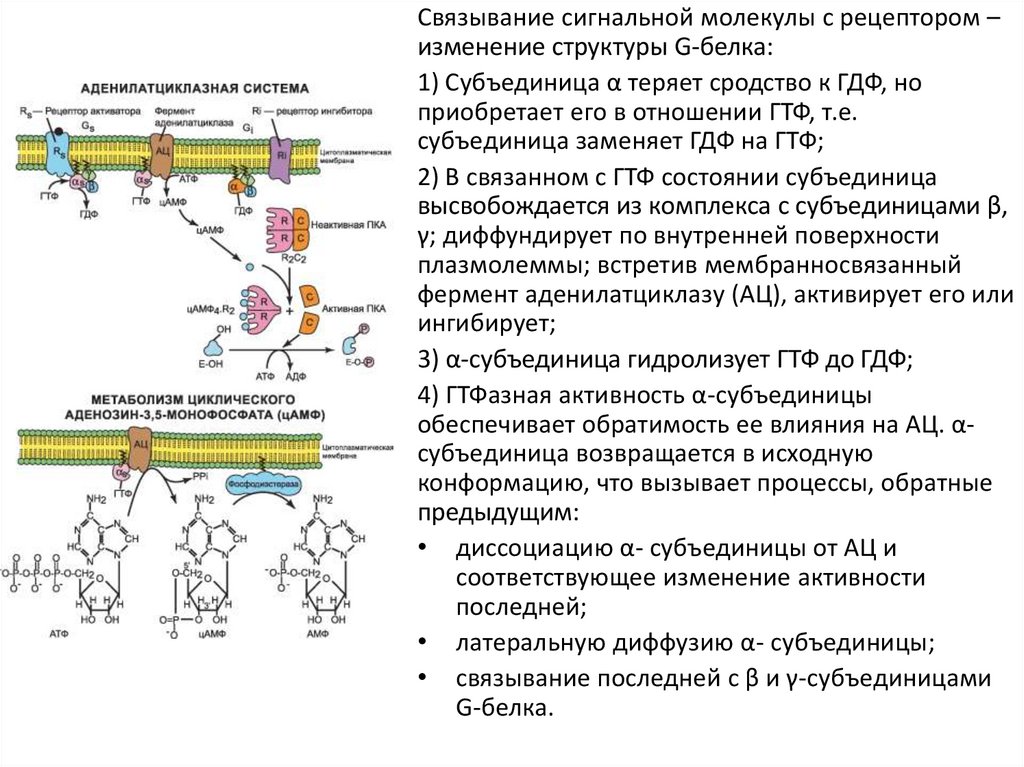
Связывание сигнальной молекулы с рецептором –изменение структуры G-белка:
1) Субъединица α теряет сродство к ГДФ, но
приобретает его в отношении ГТФ, т.е.
субъединица заменяет ГДФ на ГТФ;
2) В связанном с ГТФ состоянии субъединица
высвобождается из комплекса с субъединицами β,
γ; диффундирует по внутренней поверхности
плазмолеммы; встретив мембранносвязанный
фермент аденилатциклазу (АЦ), активирует его или
ингибирует;
3) α-субъединица гидролизует ГТФ до ГДФ;
4) ГТФазная активность α-субъединицы
обеспечивает обратимость ее влияния на АЦ. αсубъединица возвращается в исходную
конформацию, что вызывает процессы, обратные
предыдущим:
• диссоциацию α- субъединицы от АЦ и
соответствующее изменение активности
последней;
• латеральную диффузию α- субъединицы;
• связывание последней с β и γ-субъединицами
G-белка.
39.
Схема аденилатциклазной системыМембранные рецепторы регулируют образование цАМФ в цитоплазме клетки.
цАМФ связывается с протеинкиназой и переводит ее в активное состояние.
Фосфорилирование ряда белков в клетке изменяет их свойства и тем самым
опосредует биологический эффект внешнего сигнала. Фосфодиэстераза
гидролизует цАМФ до АМФ, а протеинфосфатаза дефосфорилирует
фосфопротеины, что приводит к прекращению клеточного ответа.
40.
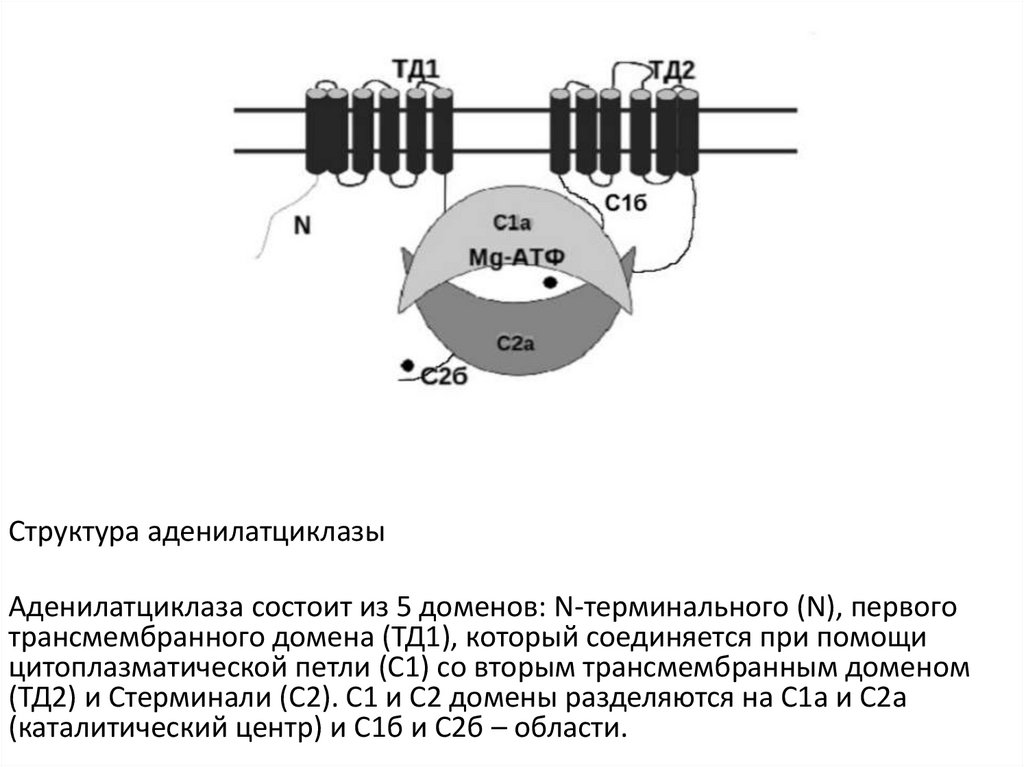
Структура аденилатциклазыАденилатциклаза состоит из 5 доменов: N-терминального (N), первого
трансмембранного домена (ТД1), который соединяется при помощи
цитоплазматической петли (С1) со вторым трансмембранным доменом
(ТД2) и Стерминали (С2). С1 и С2 домены разделяются на С1а и С2а
(каталитический центр) и С1б и С2б – области.
41.
Изменение активности аденилатциклазы и концентрациицАМФ лежат в основе эффектов ряда гормонов и
медиаторов.
Например, кортиколиберин, соматолиберин, глюкагон,
норадреналин оказывают стимулирующее действие на
аденилатциклазу,
соматостатин, ангиотензин II, ацетилхолин, дофамин,
опиоиды ингибируют синтез цАМФ.
42.
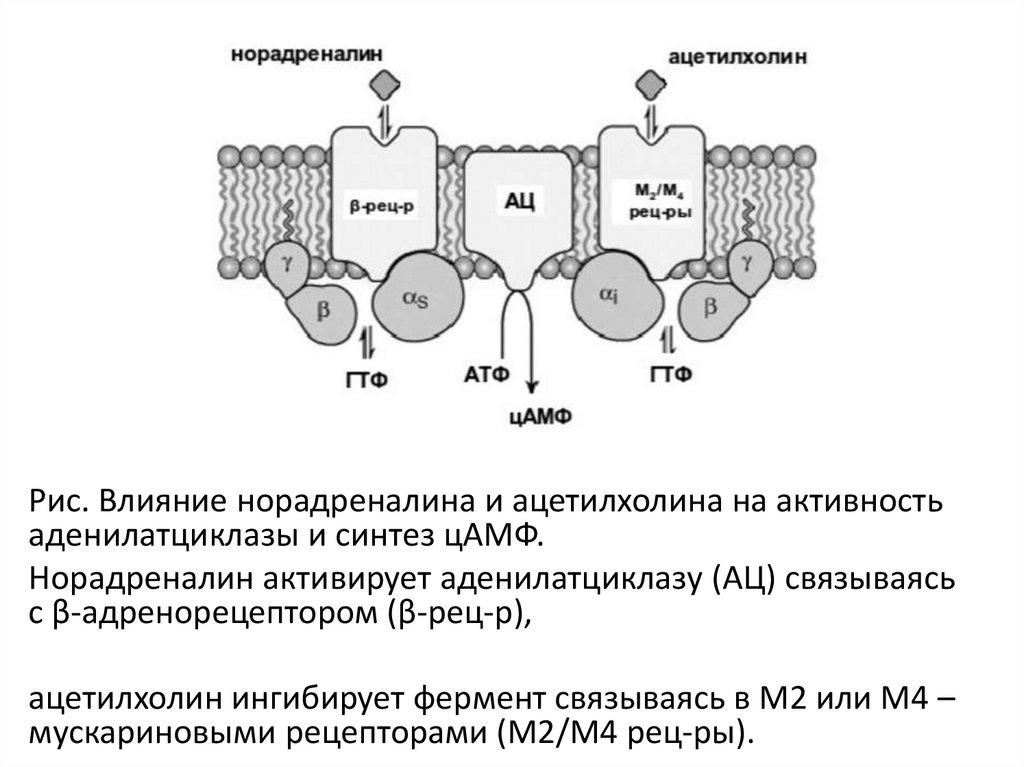
Рис. Влияние норадреналина и ацетилхолина на активностьаденилатциклазы и синтез цАМФ.
Норадреналин активирует аденилатциклазу (АЦ) связываясь
с β-адренорецептором (β-рец-р),
ацетилхолин ингибирует фермент связываясь в М2 или М4 –
мускариновыми рецепторами (М2/М4 рец-ры).
43.
Рис. Участие аденилатциклазной системы в процессах трансдукции вобонятельных нейронах. Связывание одоранта с обонятельными рецепторами
приводит к активации аденилатциклазы. Повышение уровня цАМФ приводит к
открытию цАМФактивируемых катионных ионных каналов в мембране
рецептора и деполяризации мембраны. (Из Kandel E.R., Schwartz J.H., Jessel T.M,
2002 с изменениями).
44.
Циклический АМФРассматривался как единственный вторичный посредник,
опосредующий биологическое действие нейромедиаторов и
гормонов.
Содержание цАМФ во всех тканях животных доминирует над
цГМФ.
Исключением является сетчатка - единственная ткань, сильно
обогащенная цГМФ.
45.
Циклический АМФ образуется из АТФ под действием ферментааденилатциклазы.
Второй фермент обмена цАМФ – фосфодиэстераза, катализирующая
превращение цАМФ в обычный (нециклический АМФ).
46.
Внутриклеточный уровень цАМФ в тканях млекопитающих внормальных условиях при отсутствии гормональной стимуляции
составляет 10-7 М.
При действии нейромедиаторов или гормонов внутриклеточное
содержание цАМФ резко увеличивается: глюкагон 8-кратно
увеличивает концентрацию цАМФ в печени.
47.
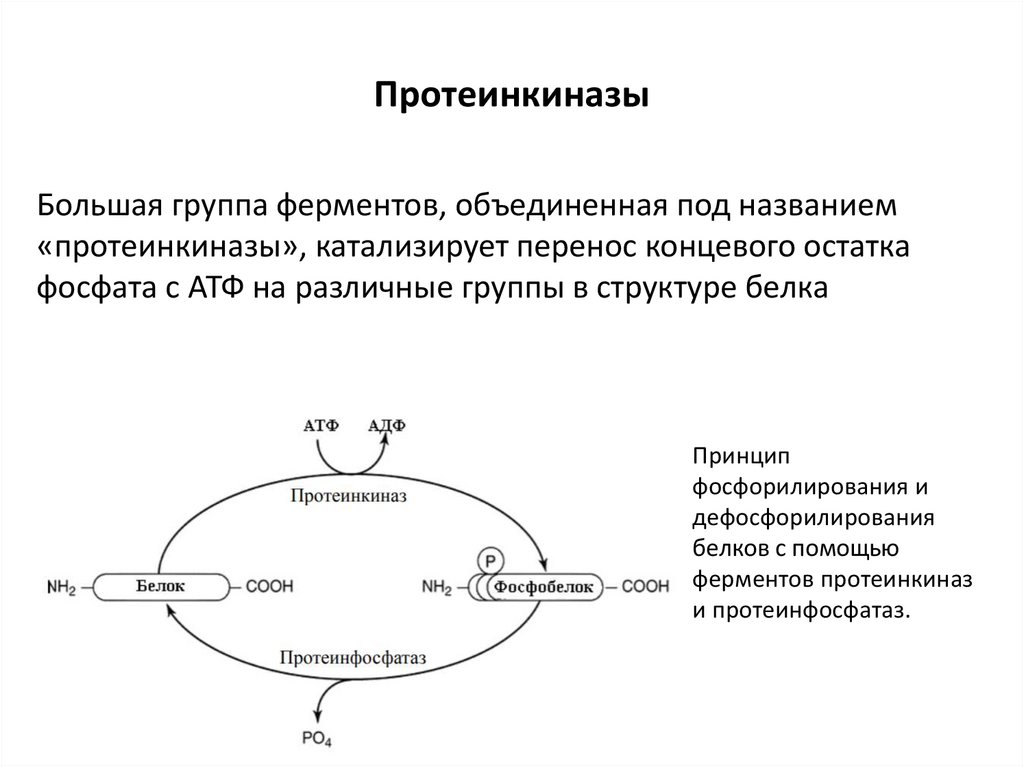
ПротеинкиназыБольшая группа ферментов, объединенная под названием
«протеинкиназы», катализирует перенос концевого остатка
фосфата с АТФ на различные группы в структуре белка
Принцип
фосфорилирования и
дефосфорилирования
белков с помощью
ферментов протеинкиназ
и протеинфосфатаз.
48.
цАМФ-зависимая протеинкиназа или протеинкиназа Аопосредует большинство биологических эффектов цАМФ во всех
клетках эукариот.
К настоящему времени известно несколько десятков
ферментов, активность которых регулируется за счет
фосфорилирования протеинкиназой А.
цАМФ может и самостоятельно изменять активность каналов (в
скелетных мышцах крысы цАМФ ингибирует Na-каналы без
активации протеинкиназы А и фосфорилирования субъединиц
канала)
49.
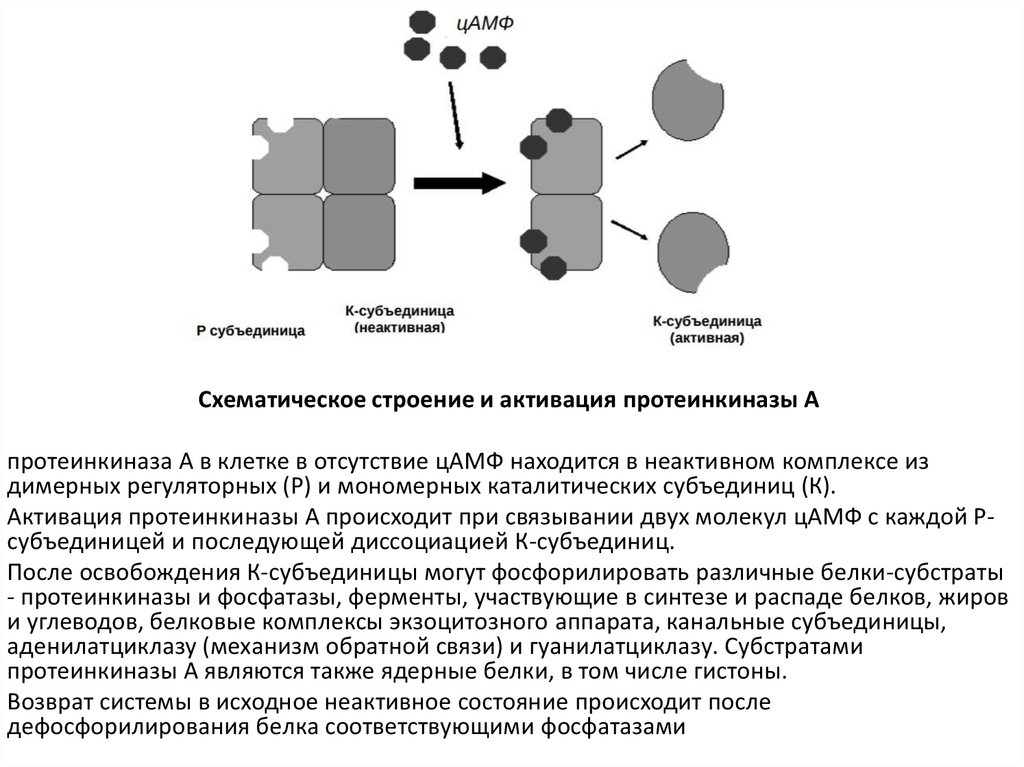
Схематическое строение и активация протеинкиназы Апротеинкиназа А в клетке в отсутствие цАМФ находится в неактивном комплексе из
димерных регуляторных (Р) и мономерных каталитических субъединиц (К).
Активация протеинкиназы А происходит при связывании двух молекул цАМФ с каждой Рсубъединицей и последующей диссоциацией К-субъединиц.
После освобождения К-субъединицы могут фосфорилировать различные белки-субстраты
- протеинкиназы и фосфатазы, ферменты, участвующие в синтезе и распаде белков, жиров
и углеводов, белковые комплексы экзоцитозного аппарата, канальные субъединицы,
аденилатциклазу (механизм обратной связи) и гуанилатциклазу. Субстратами
протеинкиназы А являются также ядерные белки, в том числе гистоны.
Возврат системы в исходное неактивное состояние происходит после
дефосфорилирования белка соответствующими фосфатазами
50.
51.
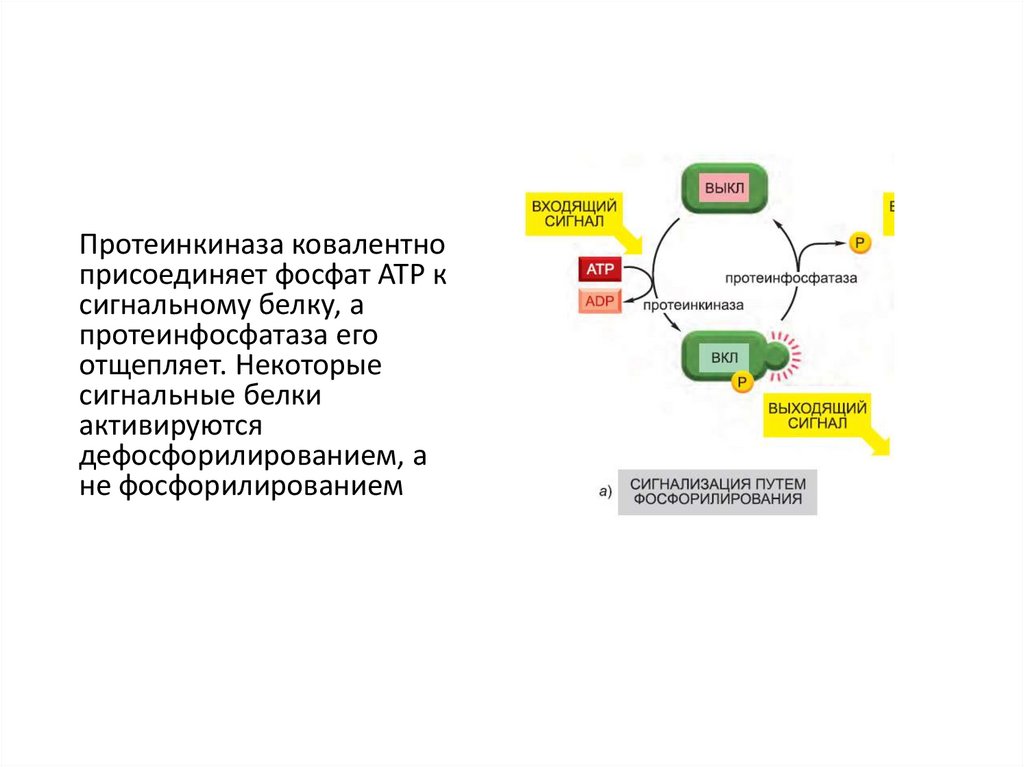
Протеинкиназа ковалентноприсоединяет фосфат ATP к
сигнальному белку, а
протеинфосфатаза его
отщепляет. Некоторые
сигнальные белки
активируются
дефосфорилированием, а
не фосфорилированием
52.
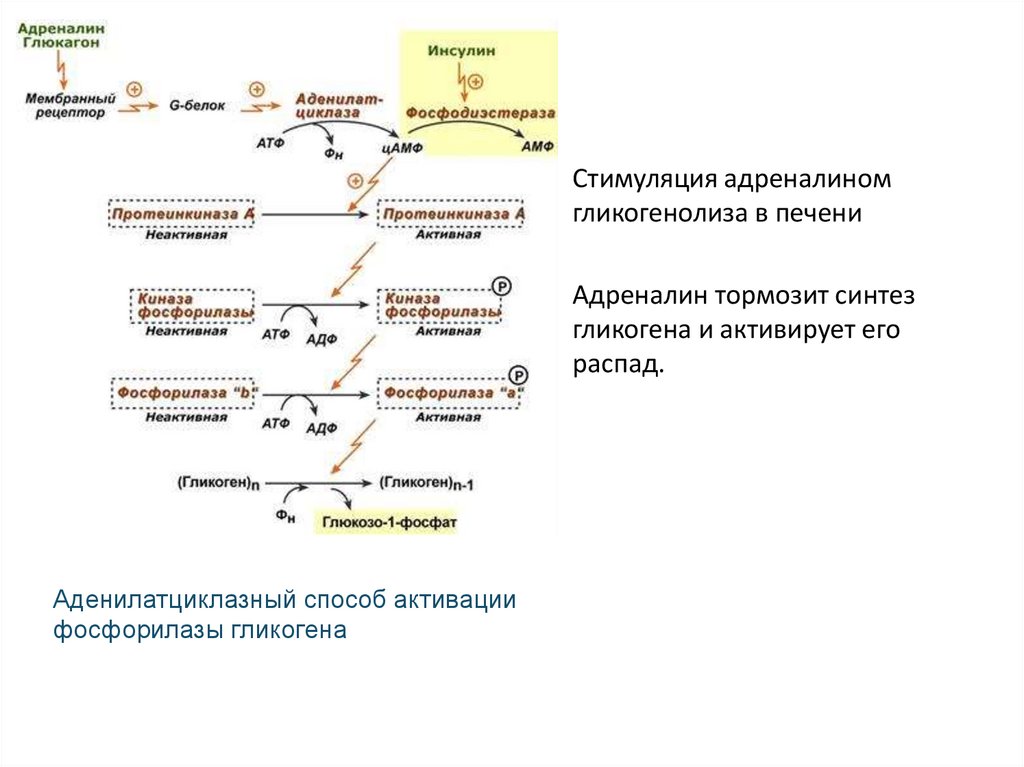
Стимуляция адреналиномгликогенолиза в печени
Адреналин тормозит синтез
гликогена и активирует его
распад.
Аденилатциклазный способ активации
фосфорилазы гликогена
53.
Стимуляция адреналином распада углеводов печени• Рецепторы адренергических синапсов: α1-, α2-, β1-, β2-адренорецепторы,
которые воспринимают также и гормональные сигналы адреналина.
1. Усиление распада гликогена
А) На поверхности гепатоцитов (а также мышечных волокон скелетных мышц)
содержатся β2-адренорецепторы – интегральные белки, пептидная цепочка
которых проходит через толщу мембраны 7 раз. С цитоплазматической
стороны плазмолеммы β2-адренорецепторы контактируют с G-белком,
стимулирующего типа. Поэтому возбуждение рецепторов приводит к
активации аденилатциклазы (АЦ).
Б) В гепатоцитах возрастает концентрация цАМФ, что активирует
протеинкиназу-А (ПК-А), которая фосфорилирует два фермента:
- Еще одну высокоспецифическую ПК – киназу фосфорилазы b (КФ-b),
которая при фосфорилировании инактивируется;
- Гликогенсинтетазу (ГС) – фермент синтеза гликогена, инактивируемый при
фосфорилировании.
- КФ-b действует на гликоген фосфорилазу b (ГФ-d) – фермент, который
расщепляет гликоген до глюкозо-1-фосфата.
54.
Гуанилатциклазная системапередачи сигналов
Гуанилатциклаза
В клетках позвоночных цГМФ синтезируется из ГТФ ферментами
гуанилатциклазами, которые классифицируют на два типа – растворимые
(тип 1) и ассоциированные с мембраной или рецепторные (тип 2) формы.
Растворимая или цитоплазматическая гуанилатциклаза
экспрессируется в цитоплазме всех клеток млекопитающих и выполняет
широкий спектр физиологических функций: ингибирование агрегации
тромбоцитов, расслабление гладкой мускулатуры, вазодилятация,
нейрональная сигнальная трансдукция и иммуномодуляция.
Фермент представляет собой гетеродимер состоящий из α- и β-субъединиц,
каждая из которых необходима для каталитической активности
гуанилатциклазы.
Различают 7 изоформ мембранно-ассоциированных или рецепторных
гуанилатциклаз (от 2А до 2G), представляющие собой гомодимеры.
55.
Циклический ГМФВажный посредник нейромедиаторного и гормонального действия,
относящийся к циклическим нуклеотидам - циклический 3`5`гуанозинмонофосфат (цГМФ).
цГМФ опосредует эффекты широкого спектра гормонов, натрийуретических
пептидов, газообразных посредников – NO и СО, Са2+, лекарственных средств
и токсинов в различных типах клеток оказывая влияние на активность ряда
внутриклеточных белков, транспорт ионов в клетке, цитоскелет и процессы
экзо- и эндоцитоза
цГМФ синтезируется в результате активации мембранной или растворимой
гуанилатциклаз,
под действием фосфодиэстераз распадается до менее активного 5’-гуанозин
монофосфата.
Кроме метаболического контроля уровня цГМФ, в клетках функционируют
насосы, откачивающие цГМФ из цитоплазмы во внеклеточную среду.
56.
Протеинкиназа GСпецифическая цГМФ-зависимая протеинкиназа
(протеинкиназа G), фермент, повышающий
фосфорилирование белков.
Протеинкиназа G принадлежит к семейству серин/треонин
киназ, широко распространенных у эукариотов. Различают
два типа протеинкиназ G - I и II.
57.
Схема гуанилатциклазной системы передачи сигналовМембранные рецепторы и NO
регулируют образование цГМФ в
цитоплазме клетки.
цГМФ связывается с протеинкиназами и
переводит их в активное состояние и
тем самым вызывает биологический
эффект.
Также цГМФ может самостоятельно
изменять активность нуклеотидзависимых каналов и фосфодиэстераз.
По Kleppisch and Feil 2009 с
изменениями.
ФДЭ — фосфодиэстеразы,
ГЦ — гуанилатциклаза,
НП A, B, C — натрийуретический
пептиды А, В, С.
58.
В скелетных мышцах цГМФ участвует в регуляциисократимости, роста и дифференцировки миобластов и
мышечных трубочек, нейротрофического контроля, нервномышечной передачи и в развитии нервно-мышечных
заболеваний
59.
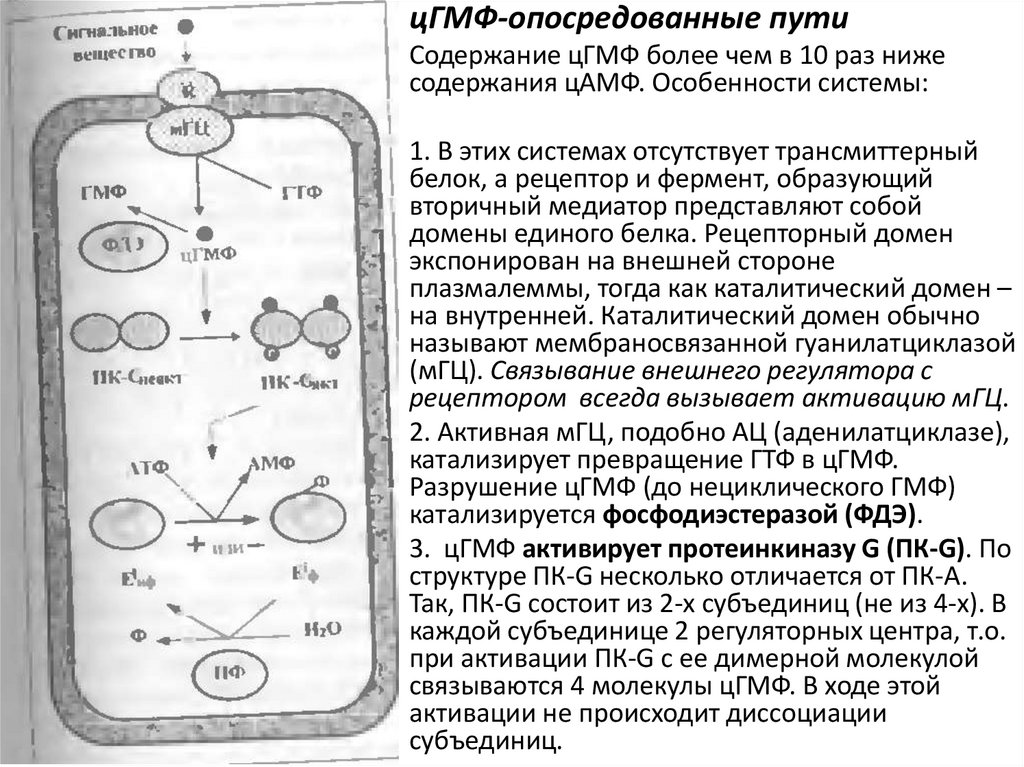
цГМФ-опосредованные путиСодержание цГМФ более чем в 10 раз ниже
содержания цАМФ. Особенности системы:
1. В этих системах отсутствует трансмиттерный
белок, а рецептор и фермент, образующий
вторичный медиатор представляют собой
домены единого белка. Рецепторный домен
экспонирован на внешней стороне
плазмалеммы, тогда как каталитический домен –
на внутренней. Каталитический домен обычно
называют мембраносвязанной гуанилатциклазой
(мГЦ). Связывание внешнего регулятора с
рецептором всегда вызывает активацию мГЦ.
2. Активная мГЦ, подобно АЦ (аденилатциклазе),
катализирует превращение ГТФ в цГМФ.
Разрушение цГМФ (до нециклического ГМФ)
катализируется фосфодиэстеразой (ФДЭ).
3. цГМФ активирует протеинкиназу G (ПК-G). По
структуре ПК-G несколько отличается от ПК-А.
Так, ПК-G состоит из 2-х субъединиц (не из 4-х). В
каждой субъединице 2 регуляторных центра, т.о.
при активации ПК-G с ее димерной молекулой
связываются 4 молекулы цГМФ. В ходе этой
активации не происходит диссоциации
субъединиц.
60.
Белки, являющиеся субстратами протеинкиназы G – пока сопределенностью не установлены. Таких белков немного (например, в
головном мозге выявлен 1 белок, специфически фосфорилируемый
данной ПК. Он содержится в клетках Пуркинье мозжечка и является
термостабильным. Функция его неизвестна, поэтому он так и называется
– G-субстрат).
61.
Действие ацетилхолина на М2-холинорецепторы• Данные рецепторы обнаружены в сердце, мозге, их возбуждение
ацетилхолином приводит (через G-белок) к угнетению АЦ.
Допускается, что в соответствующих клетках сердца и мозга имеются
разновидности м2-ХР: одни рецепторы связаны с G-белком, а другие – с
мГЦ.
В клетках, под влиянием ацетилхолина, снижается концентрация цАМФ и
возрастает концентрация цГМФ.
Действие ПК-с может достигаться посредством влияния на системы
транспорта ионов:
- Увеличения проводимости для K+- и анионных каналов;
- Уменьшения проводимости Na+-каналов;
- Уменьшение проводимости Ca2+ - каналов саркоплазматического
ретикулума.
62.
цГМФ и NO-опосредованные путиКроме мембраносвязанной гуанилатциклазы (мГЦ), во многих клетках
имеется и, так называемая, растворимая гуанилатциклаза (рГЦ),
локализующаяся в цитозоле. Она не только катализирует образование
вторичного мессенджера (цГМФ), но и сама активизируется другим таким
мессенджером – оксидом азота (NO).
NO образуется из аминокислоты аргинина ферментом NO–синтазой (NO-C)
и способен диффундировать через мембраны. Поэтому NO может
действовать как интракринно, так и паракринно.
63.
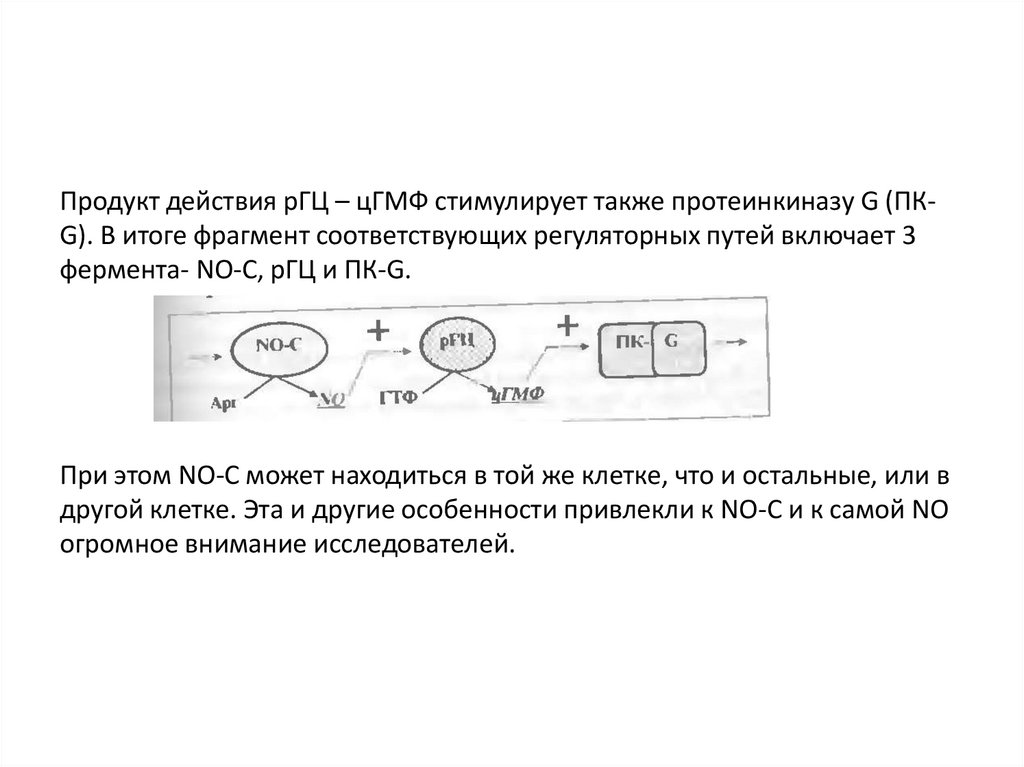
Продукт действия рГЦ – цГМФ стимулирует также протеинкиназу G (ПКG). В итоге фрагмент соответствующих регуляторных путей включает 3фермента- NO-C, рГЦ и ПК-G.
При этом NO-C может находиться в той же клетке, что и остальные, или в
другой клетке. Эта и другие особенности привлекли к NO-C и к самой NO
огромное внимание исследователей.
64.
Несмотря на интенсивное изучение, для подавляющего большинства NOи –цГМФ- опосредованных регуляторных путей так и остаются неяснымиих начальные и завершающие стадии.
В этих системах объединяются прямыми и обратными связями реакции
образования не двух, а еще большего числа вторичных мессенджеров,
каждый из которых действует на «свой» объект. Получаются
разветвленные многофункциональные системы, которые не так просто
«разбить» на независимые цепочки последовательно проходящих
событий.
65.
Функции NO:• Сосудорасширяющее действие (повышенное давление на стенки
кровеносного сосуда ведет активации NO-C в эндотелиоцитах);
увеличению в них синтеза NO, расслаблению гладкомышечных клеток;
• Выполняет функции внутриклеточного мессенджера, который образуется
в ответ на раздражение нейрона нейромедиатором (глутаминовой
кислотой или ГАМК);
• Является медиатором, из пресинаптического окончания выделяется путем
простой диффузии, диффундирует через постсинаптическую мембрану к
гуанилатциклазе;
• NO как нейромедиатор (для некоторых ядер гипоталамуса);
• При больших концентрациях становится цитотоксичным.
66.
Пути, опосредованные липидами и ионами Ca2+Липиды, за исключением стероидных гормонов и простагландинов,
могут выступать вторичными мессенджерами во внутриклеточных
регуляторных путях.
Связывание сигнального вещества с мембранным рецептором приводит
к активации фермента фосфолипазы С (ФЛ-С), который находится на
внутренней поверхности плазмолеммы. Механизм активации зависит от
формы ФЛ-С.
1) Если это β-изоформа:
• Сигнальным веществом служит гормон;
• Рецептор – интегральный белок;
• Трансмиттером является G-белок стимулирующего типа, который
активирует ФЛ-С с помощью диссоциирующей α-субъединицы
67.
68.
2) Свойства системы, связанной с γ-изоформой ФЛ-С- В качестве сигнального вещества выступает один из факторов роста;
- Рецепторный белок один раз «прошивает» мембрану, причем у его
цитоплазматического домена имеется тирозинкиназная активность (в
белке-мишени фосфорилируются остатки тирозина);
- Фосфорилирование данным доменом γ-формы ФЛ-С приводит к
активации последней.
69.
Основным субстратом ФЛ-С является мембранный липид –фосфатидилинозитдифосфат (ФИД) (амфифильный липид).
ФЛ-С гидролизует связь между глицерином и фосфатным остатком. В
результате ФИД расщепляется на два соединения:
- Полярное – инозиттрифосфат (ИТФ);
- Преимущественно неполярное – диацилглицерин (ДАГ).
Для проявления активности фосфолипазы С необходимы ионы Ca2+.
70.
• ИТФ является гидрофильным, свободно диффундирует вовнутриклеточном объеме. Связываясь с Са2+-каналами плазмолеммы
и эндоплазматического ретикулума, ИТФ переводит их в открытое
состояние.
В результате в цитозоле повышается концентрация Ca2+.
Т.о. действие ИТФ опосредовано ионами Са2+ и включает все те эффекты,
которые стимулируются в клетке этими ионами.
71.
• ДАГ – являясь неполярным, а следовательно гидрофобным, способнолишь к латеральной диффузии в составе плазмолеммы.
Этого оказывается достаточным для активации протеинкиназы С.
Т.е. совокупность эффектов, вызываемых ДАГ в клетке, определяется
спектром действия ПК-С в этой клетке.
72.
В качестве примера ФЛ-С-опосредованной регуляции: действиеадреналина на α1-адренорецепторы, расположенные на гладких миоцитах
ряда сосудов;
- β2-адренорецепторы связаны с G-белком, который активирует
аденилатциклазу (АЦ);
- Α2-адренорецепторы связаны с G1-белком, ингибирующим АЦ.
В отличие от этого α1-адренорецепторы (α1-АР) связаны с G-белком,
который активирует ФЛ-С.
В результате при стимуляции α1-АР в миоците должны возрастать Са2+ и
активность ПК-С.
В гладких миоцитах ключевую роль играет миозинкиназа (МК) –
специфическая ПК, которая фосфорилирует в толстых миофиламентах
головки миозина и тем самым придает им способность взаимодействовать
с тонкими филаментами.
При стимуляции α1-адренорецепторов миоциты сокращаются.
73.
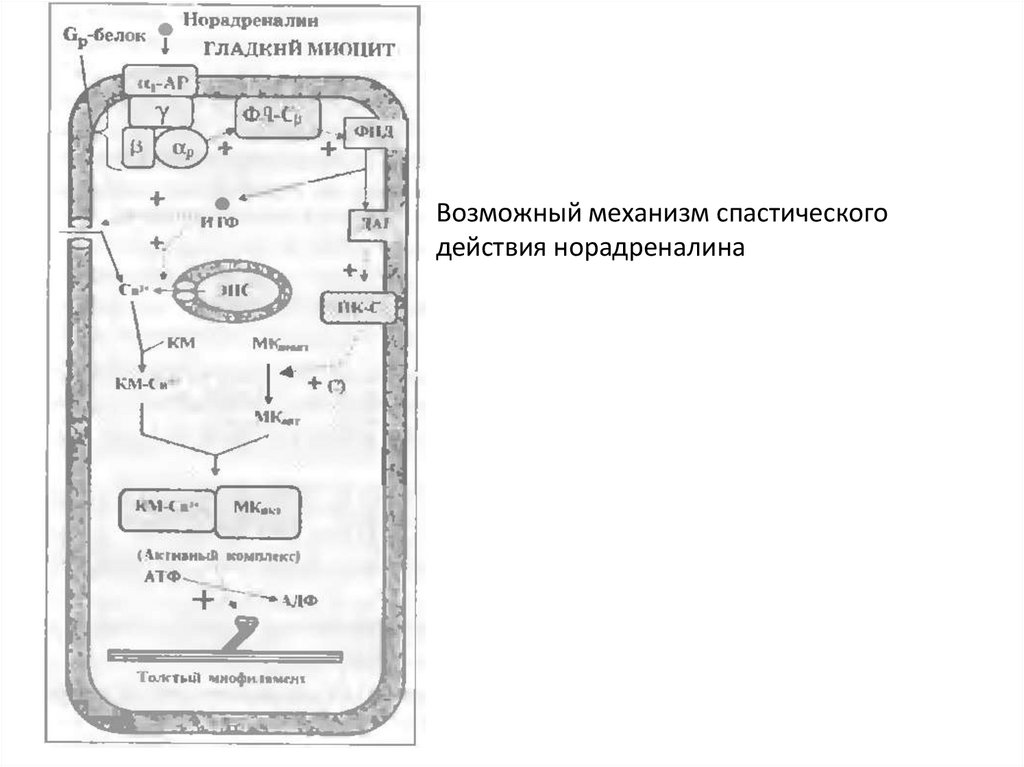
Возможный механизм спастическогодействия норадреналина
74.
Пути, опосредованные белком RasRas-белок:
• Может активироваться протеинкиназой С (ПК-С);
• Запускает каскад митогенактивируемых протеинкиназ, который,
модифицируя соответствующие транскрипционные факторы,
контролирует активность ряда важнейших генов – в том числе
тех, которые отвечают за клеточное деление;
75.
Пути, не содержащие вторичного мессенджераНапример, влияние γ-интерферона на транскрипцию определенных
генов
1)
2)
3)
4)
5)
В отсутствие сигнального вещества рецепторы находятся в
мономерной форме, а при связывании этого вещества они
объединяются в димерные структуры;
Димеризация рецепторов стимулирует тирозинкиназную (ТК-)
активность; при этом тирозинкиназа – самостоятельный фермент,
называемый Янус-киназой;
Вначале Янус-киназы фосфорилируют близлежащие части
рецепторов. Это способствует присоединению к комплексу
рецептор-JAK и т.п. STAT-белка. После этот белок тоже
фосфорилируется.
В таком состоянии проникает в ядро клетки и выступает как
транскрипционный фактор, это и запускает транскрипцию.
В данном пути передатчиком сигнала от мембранного рецептора к
эффекторному устройству (определенной части хромосом) служит
такое вещество (STAT-белок), которое должно непосредственно
входить в это устройство для его функционирования. В связи с этим
говорят о том, что в данном случае отсутствует классический
внутриклеточный мессенджер.
76.
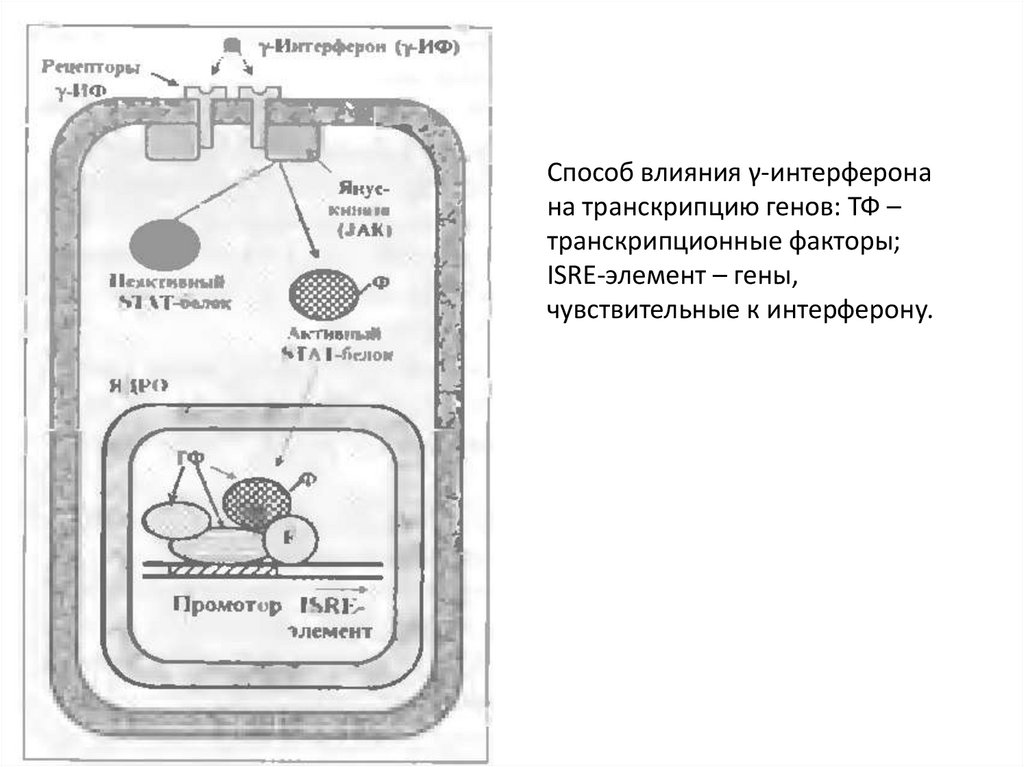
Способ влияния γ-интерферонана транскрипцию генов: ТФ –
транскрипционные факторы;
ISRE-элемент – гены,
чувствительные к интерферону.