











 Биология
БиологияПохожие презентации:










Трансмембранная передача сигналов
1.
Трансмембраннаяпередача сигналов
2.
Важное свойство мембран - способность воспринимать ипередавать внутрь клетки сигналы из внешней среды.
Внеклеточными химическими сигналами могут быть гормоны,
нейромедиаторы, эйкозаноиды или другие сигнальные молекулы.
Гормоны - это молекулы, которые вырабатываются
специализированными клетками, секретируются в кровь в ответ
на изменение какого-либо специфического параметра внутренней
среды организма и оказывают влияние на метаболизм и
функциональное состояние (пролиферация, секреция и т.д.)
клеток-мишеней. Клетку-мишень определяют по способности
избирательно связывать данную сигнальную молекулу с помощью
белка-рецептора
Рецепторы (R) могут быть:
• встроены в мембрану клеток-мишеней;
• находиться в клетке (в цитозоле или в ядре).
3.
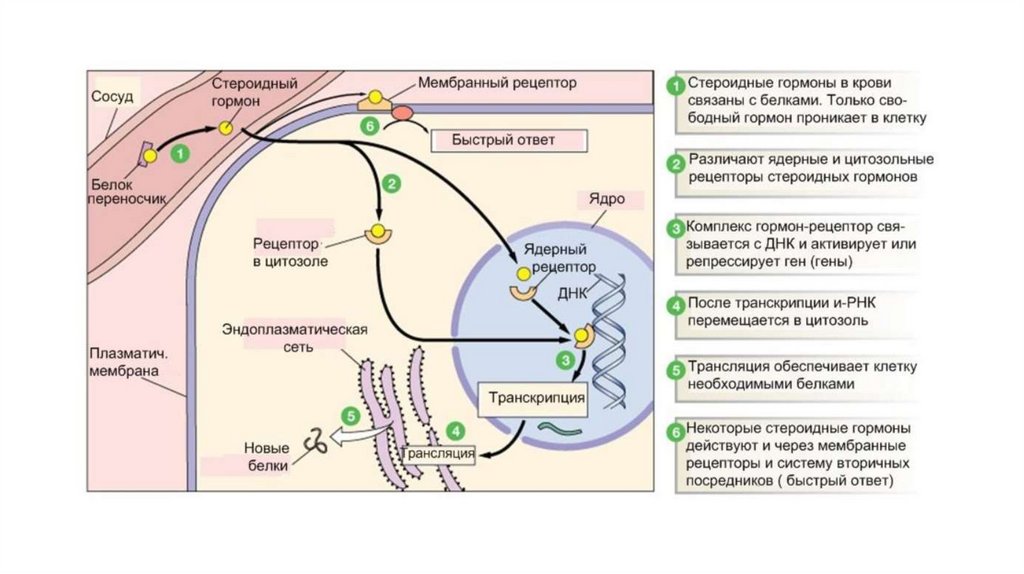
Цитозольно ядерный механизм передачисигнала
Транскрипция - это процесс, в ходе которого информация из гена в
ДНК переписывается в молекулы РНК. В результате транскрипции
образуется молекула мРНК (мессенджерная РНК), которая
содержит код информации, необходимый для создания белка.
Трансляция - это процесс, в ходе которого молекула мРНК
"переводится" на язык белков. Рибосомы считывают код на мРНК и
синтезируют соответствующий белок, используя аминокислоты.
Этот процесс происходит в цитоплазме клетки.
4.
5.
Неполярные гормоны (стероидные гормоны, йодсодержащиегормоны щитовидной железы и некоторые другие) путем простой
диффузии проходят через плазматическую мембрану, а также
другие клеточные мембраны, и связываются с рецепторами,
которые располагаются в цитозоле или ядре клетки. Если
рецептор находится в цитозоле, то образующийся гормонрецепторный комплекс в дальнейшем проникает в ядро. Если
рецептор локализуется в ядре, то гормон проходит через ядерную
мембрану и связывается с рецептором непосредственно в
кариоплазме. В любом случае образовавшийся гормонрецепторный комплекс взаимодействует с молекулой ДНК и
влияет на транскрипцию генов. Такой механизм передачи
гормонального сигнала называется внутриклеточным или
цитозольно- ядерным. Повышение транскрипции определенных
генов влечет за собой активацию трансляции, изменяется
количество определенных белков в клетке, многие из которых
являются ферментами или внутриклеточными регуляторными
молекулами. В результате в клетке происходит изменение обмена
веществ, меняется ее метаболическая активность, а также
возможен переход клетки в другую фазу клеточного цикла.
6.
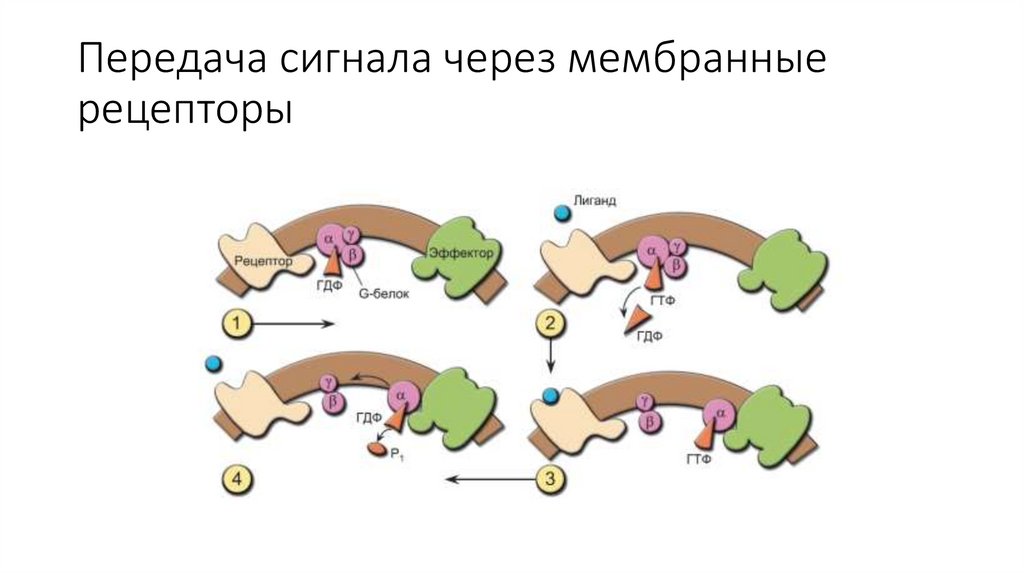
Передача сигнала через мембранныерецепторы
7.
Гормоны, имеющие полярную природу, не могут свободнопроходить через двойной слой липидов биологических мембран,
поэтому рецепторы к этим гормонам, как правило, находятся на
мембране и имеют участок связывания гормона, обращенный в
кровяное русло. Мембранные рецепторы могут обладать
различными свойствами. Существуют рецепторы, обладающие
ферментативной активностью, которые при связывании гормона
непосредственно запускают каскад реакций, приводящих к
изменению обмена веществ в клетке. Широко представлена в
организме группа рецепторов, которые сами ферментативной
активностью не обладают, но ассоциированы с гуаниннуклеотидсвязывающим белком, который называют G-белком.
8.
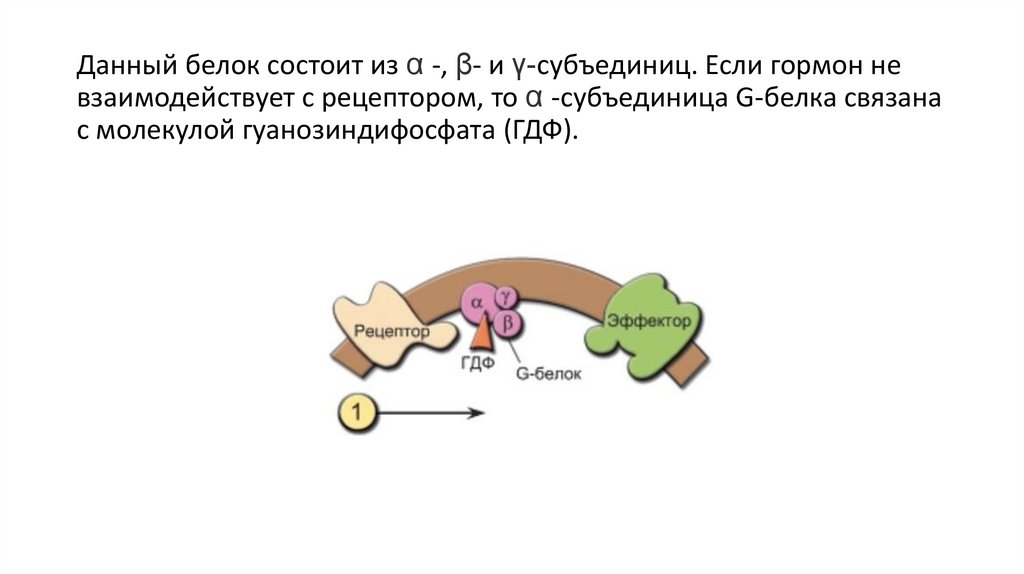
Данный белок состоит из α -, β- и γ-субъединиц. Если гормон невзаимодействует с рецептором, то α -субъединица G-белка связана
c молекулой гуанозиндифосфата (ГДФ).
9.
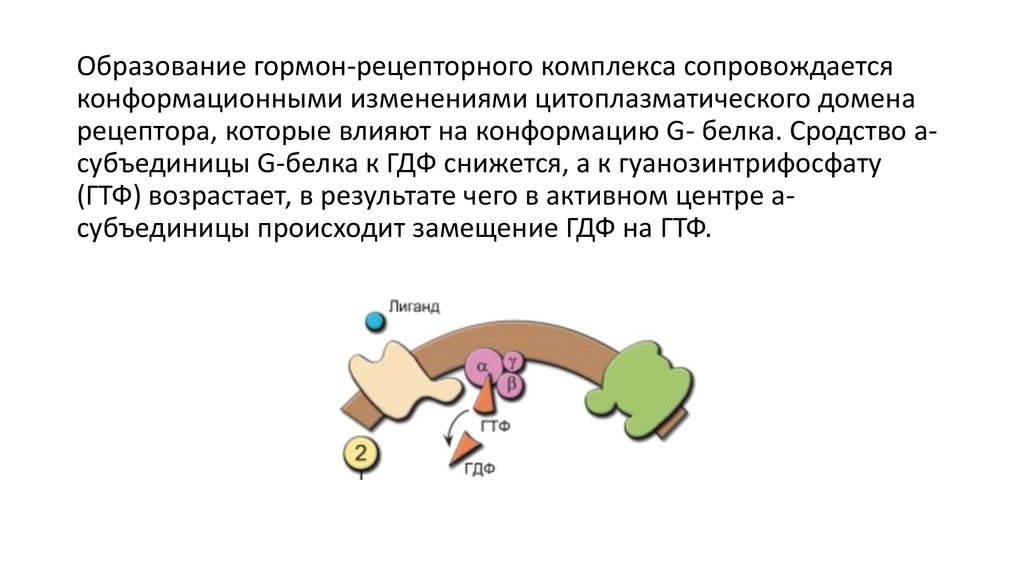
Образование гормон-рецепторного комплекса сопровождаетсяконформационными изменениями цитоплазматического домена
рецептора, которые влияют на конформацию G- белка. Сродство асубъединицы G-белка к ГДФ снижется, a к гуанозинтрифосфату
(ГТФ) возрастает, в результате чего в активном центре асубъединицы происходит замещение ГДФ на ГТФ.
10.
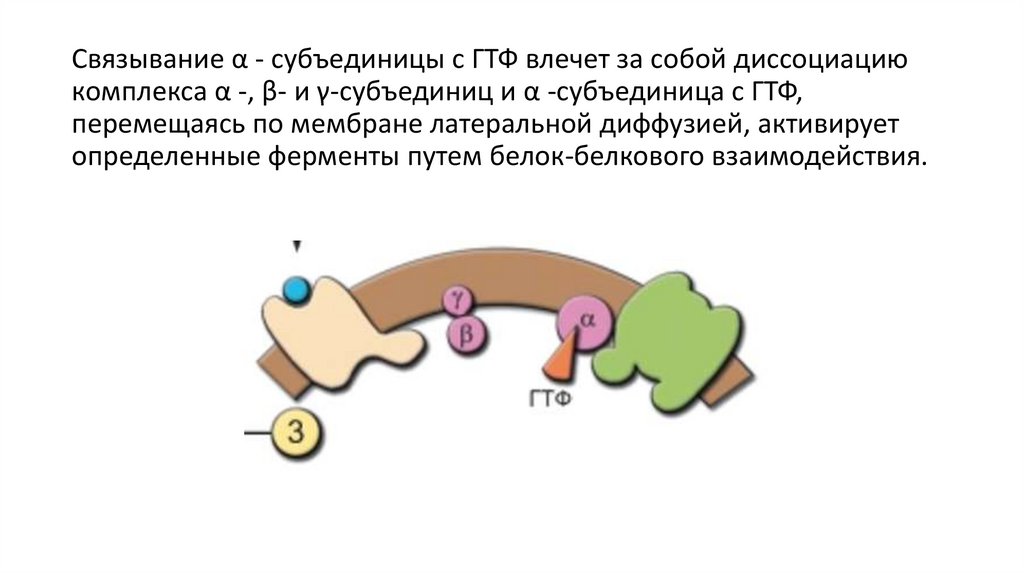
Связывание α - субъединицы с ГТФ влечет за собой диссоциациюкомплекса α -, β- и γ-субъединиц и α -субъединица с ГТФ,
перемещаясь по мембране латеральной диффузией, активирует
определенные ферменты путем белок-белкового взаимодействия.
11.
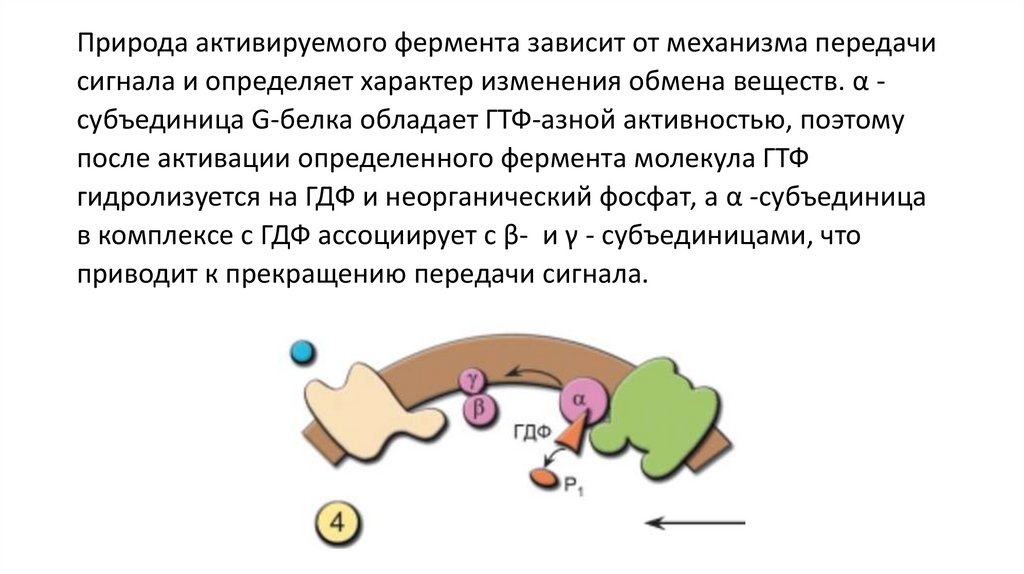
Природа активируемого фермента зависит от механизма передачисигнала и определяет характер изменения обмена веществ. α субъединица G-белка обладает ГТФ-азной активностью, поэтому
после активации определенного фермента молекула ГТФ
гидролизуется на ГДФ и неорганический фосфат, а α -субъединица
в комплексе с ГДФ ассоциирует с β- и γ - субъединицами, что
приводит к прекращению передачи сигнала.
12.
Аденилатциклазный механизм передачисигнала
Многочисленные гормоны, включая адренокортикотропный
гормон (АКТГ), антидиуретический гормон (АДГ), гонадотропины,
тиреотропный гормон (ТТГ), глюкагон, катехоламины (адреналин/
норадреналин) и др. осуществляют передачу сигнала по
аденилатциклазному механизму
Рецептор для этих сигнальных молекул располагается в
плазматической мембране клеток мишеней и ассоциирован с Gбелком. В данном случае взаимодействие гормона с рецептором
приводит к тому, что α -субъединица в комплексе ГТФ,
перемещаясь по мембране латеральной диффузией связывается с
молекулой фермента аденилатциклазы и активирует ее.
13.
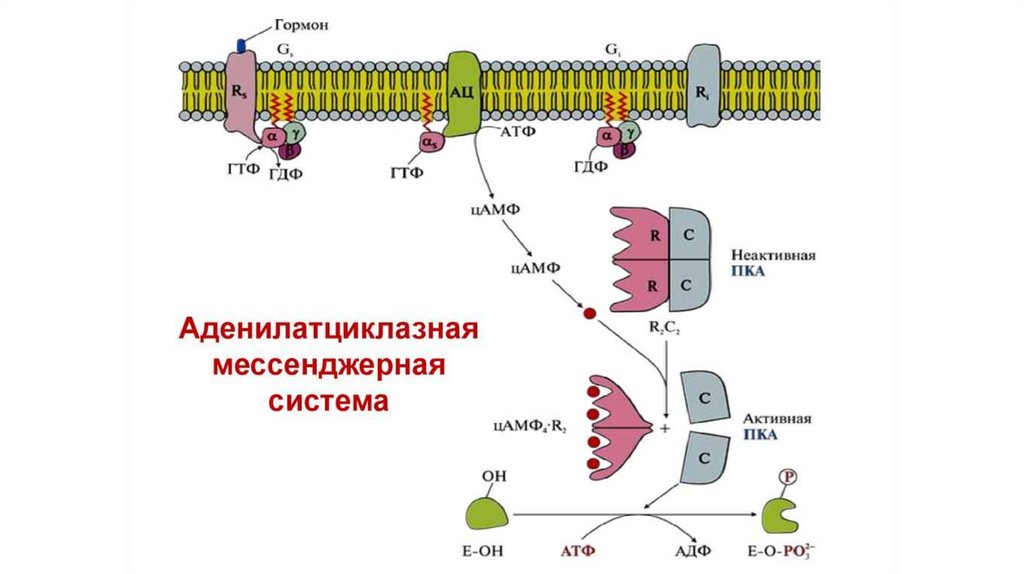
14.
Активная аденилатциклаза катализирует реакцию образованияциклического аденозинмонофосфата (цАМФ) из
аденозинтрифосфата (АТФ). цАМФ играет роль вторичного
посредника в передаче сигнала, активируя протеинкиназу A.
Неактивная протеинкиназа А (ПКА) представлена комплексом из 4х субъединиц, две из которых каталитические, а две регуляторные. 4 молекулы цАМФ присоединяются к регуляторным
субъединицам неактивной протеинкиназы А, что приводит к
диссоциации комплекса и активации протеинкиназы А.
Протеинкиназа А осуществляет фосфорилирование белковмишеней, многие из которых являются ферментами, по остаткам
серина и треонина, что приводит к изменению их активности. В
результате изменяется скорость определенных метаболических
процессов в клетке, и изменяется обмен веществ.
15.
Инозитолфосфатная система передачисигнала
Передача сигнала в клетку адреналином через αадренорецепторы, вазопрессином через V1-рецепторы и рядом
других сигнальных молекул осуществляется через
инозитолфосфатную систему. Рецептор для этих сигнальных
молекул также располагается в плазматической мембране клеток
мишеней и ассоциирован G-белком. В данном случае после
образования гормон-рецепторного комплекса а-субъединица в
комплексе с ГТФ активирует фермент фосфолипазу С.
16.
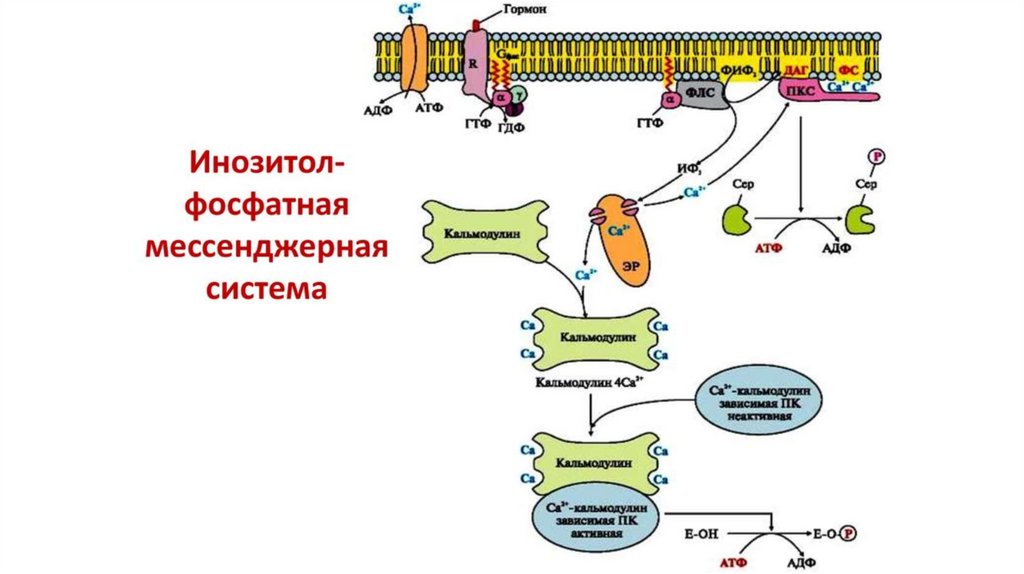
17.
Активная фосфолипаза С катализирует реакцию гидролиза мембранноголипида фосфатидилинозитол-дифосфата (ФИФ2) на
инозитолтрифосфат (ИФ3) и диацилглицерол (ДАГ), которые будут
выполнять функции вторичных посредников в передаче сигнала.
Неполярная молекула ДАГ остается в мембране, а полярный ИФ3,
выходит в цитозоль и вызывает открытие лиганд-зависимых Са2+ каналов в мембране эндоплазматического ретикулума и
высвобождение Са2+ в цитозоль.
Ионы кальция связываются с белком кальмодулином. Каждая молекула
кальмодулина имеет четыре участка связывания для ионов Са2+ .
Комплекс кальмодулина с Са2+ изменяет активность различных
внутриклеточных белков.
К числу ферментов, регулируемых таким способом, относятся Са2+
кальмодулин-активируемых протеинкиназы. В результате происходит
изменение активности ряда ферментов и сдвиги в обмене веществ.
18.
Диацилглицерол вместе с ионами Са2+ является физиологическимактиватором протеинкиназы С, кроме того для максимальной
активации этого фермента также необходим мембранный липид
фосфатидилсерин. Протеинкиназа С, связанная с молекулами
фосфатидилсерина цитозольной стороны клеточной мембраны,
фосфорилирует различные белки-мишени, что приводит к
изменению их активности и сдвигам в обмене веществ в клетке.
19.
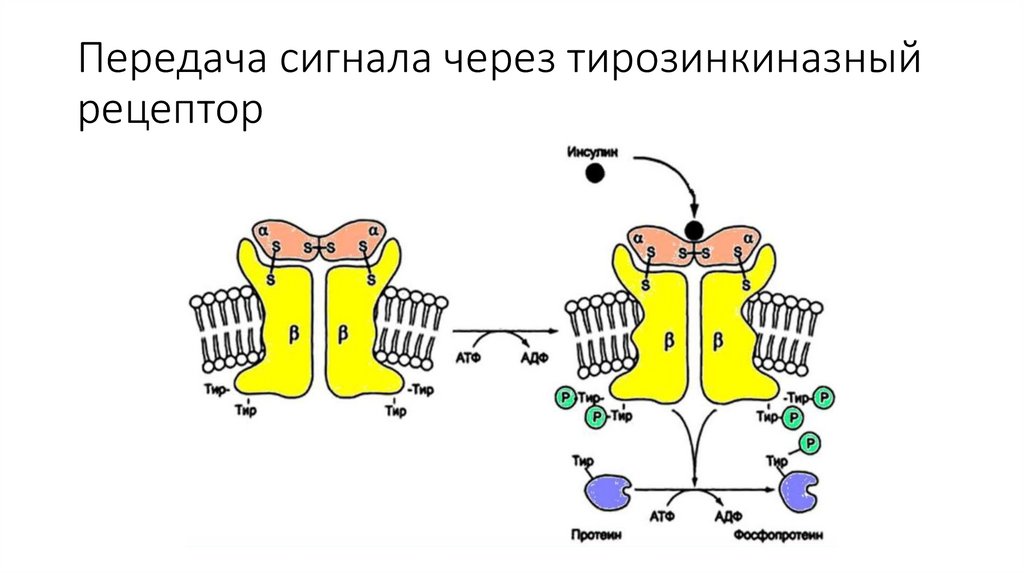
Передача сигнала через тирозинкиназныйрецептор
20.
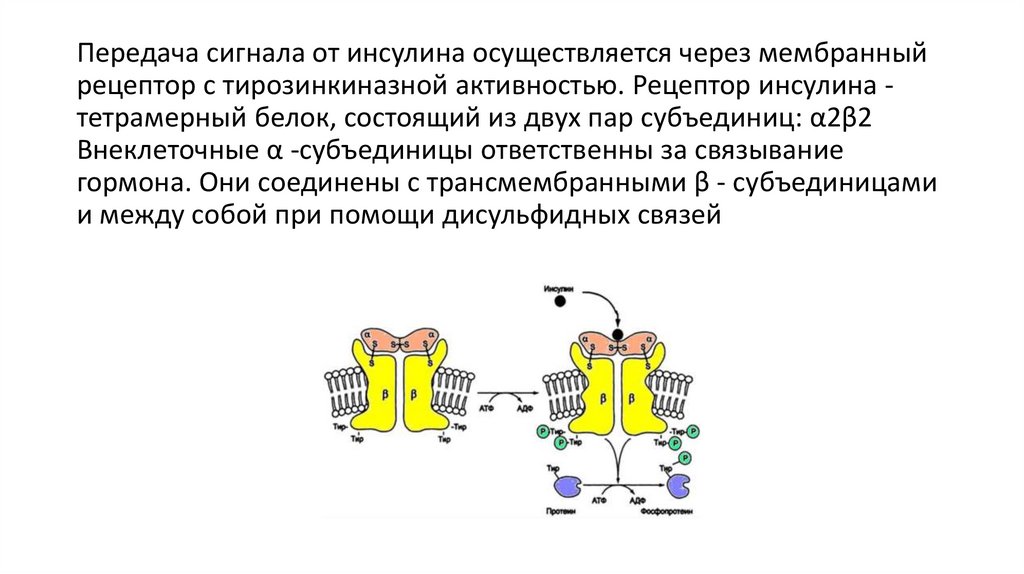
Передача сигнала от инсулина осуществляется через мембранныйрецептор с тирозинкиназной активностью. Рецептор инсулина тетрамерный белок, состоящий из двух пар субъединиц: α2β2
Внеклеточные α -субъединицы ответственны за связывание
гормона. Они соединены с трансмембранными β - субъединицами
и между собой при помощи дисульфидных связей
21.
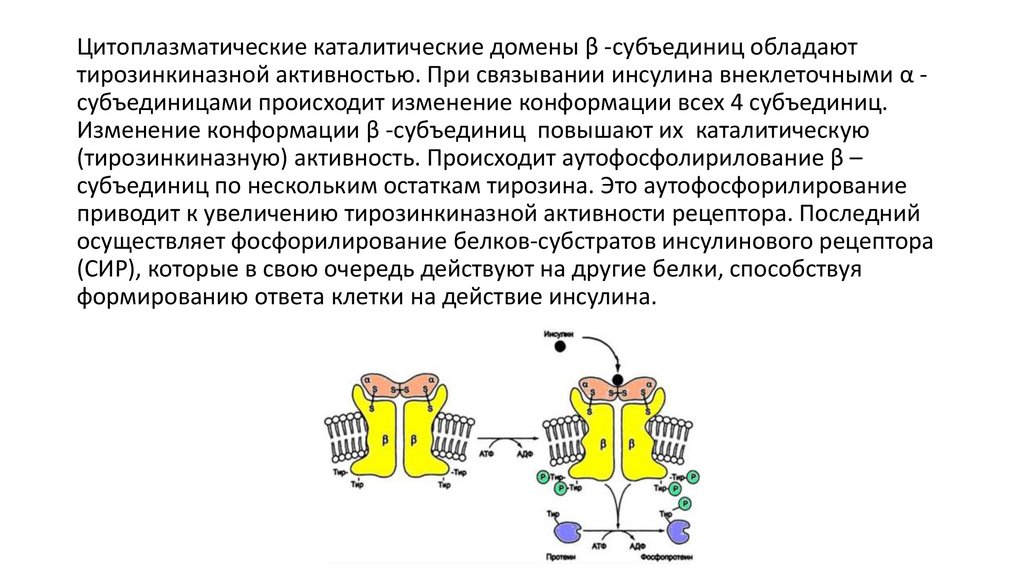
Цитоплазматические каталитические домены β -субъединиц обладаюттирозинкиназной активностью. При связывании инсулина внеклеточными α субъединицами происходит изменение конформации всех 4 субъединиц.
Изменение конформации β -субъединиц повышают их каталитическую
(тирозинкиназную) активность. Происходит аутофосфолирилование β –
субъединиц по нескольким остаткам тирозина. Это аутофосфорилирование
приводит к увеличению тирозинкиназной активности рецептора. Последний
осуществляет фосфорилирование белков-субстратов инсулинового рецептора
(СИР), которые в свою очередь действуют на другие белки, способствуя
формированию ответа клетки на действие инсулина.
22.
Биохимические эффекты инсулина заключаются в следующем:1) активация фосфопротеинфосфатаз, дефосфорилирующих белки;
2) активация фосфодиэстераз, расщепляющих циклические
нуклеотиды (цАМФ, цГМФ);
3) встраивание транспортеров глюкозы ГЛЮТ4 в мембраны
скелетных миоцитов и адипоцитов, что обеспечивает поступление
глюкозы в эти клетки,
4)активация ПСАТ-белков (переносчики сигнала и активаторы
транскрипции), влияющих на транскрипцию генов в клеткахмишенях
23.
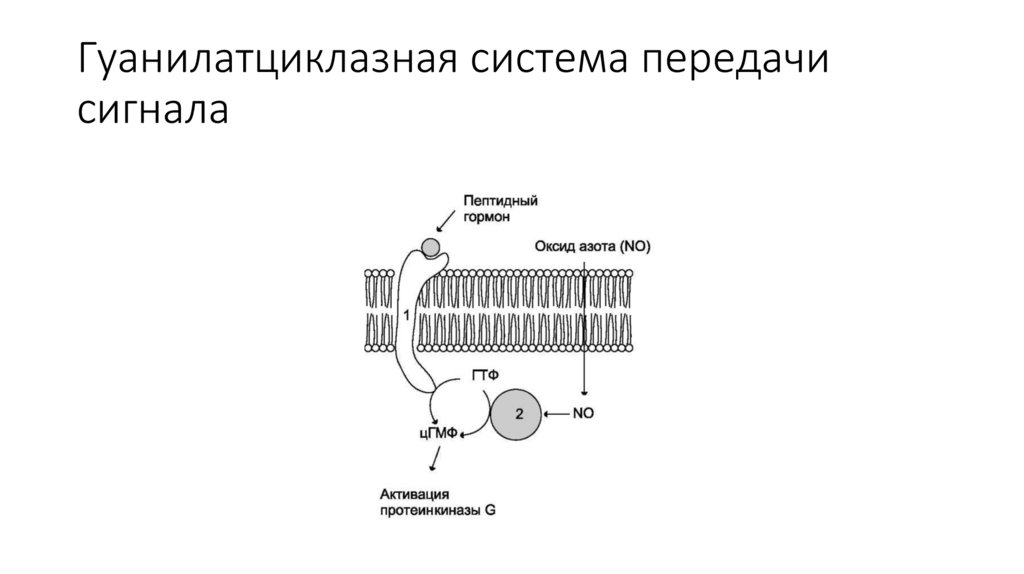
Гуанилатциклазная система передачисигнала
24.
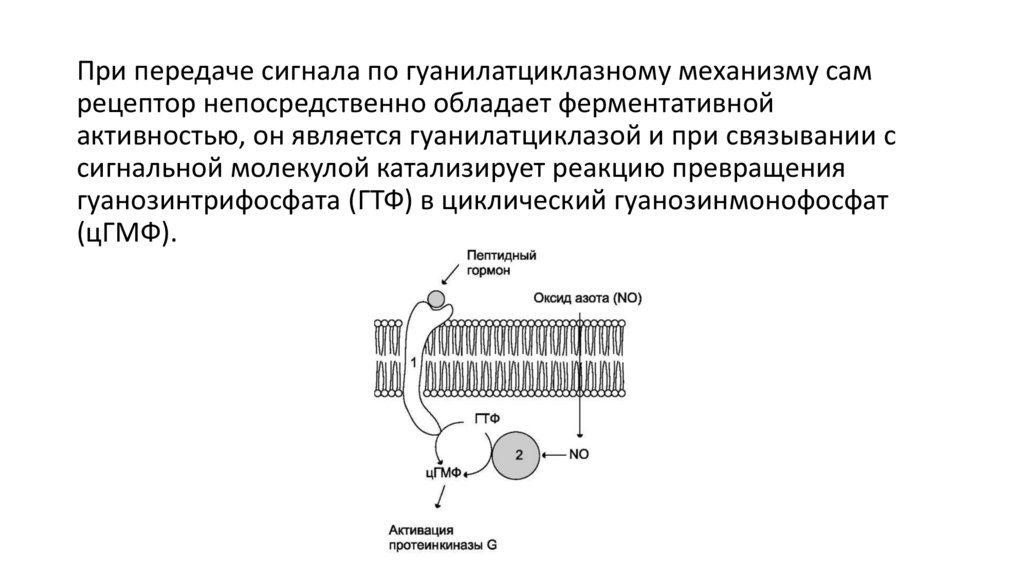
При передаче сигнала по гуанилатциклазному механизму самрецептор непосредственно обладает ферментативной
активностью, он является гуанилатциклазой и при связывании с
сигнальной молекулой катализирует реакцию превращения
гуанозинтрифосфата (ГТФ) в циклический гуанозинмонофосфат
(цГМФ).
25.
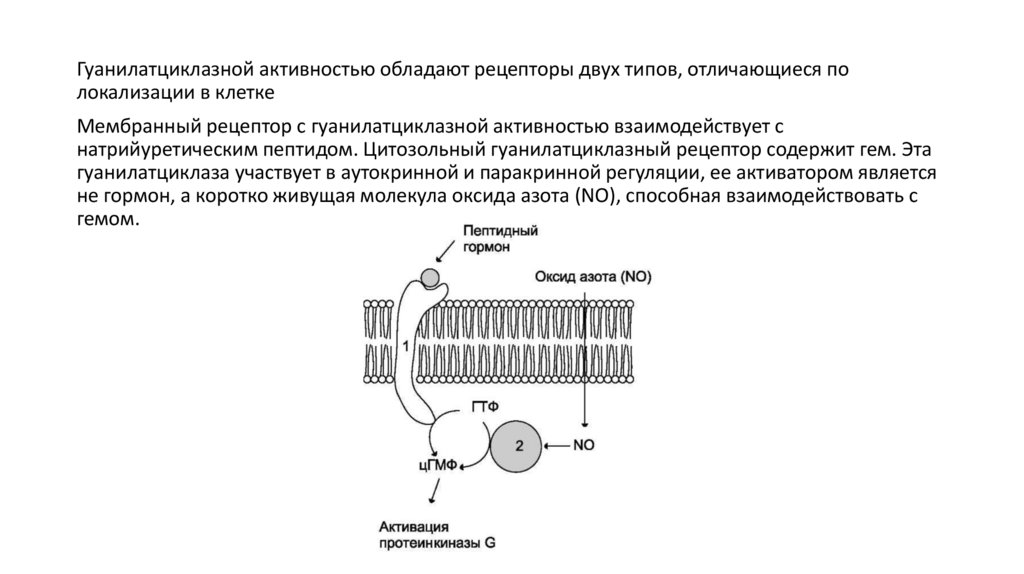
Гуанилатциклазной активностью обладают рецепторы двух типов, отличающиеся полокализации в клетке
Мембранный рецептор с гуанилатциклазной активностью взаимодействует с
натрийуретическим пептидом. Цитозольный гуанилатциклазный рецептор содержит гем. Эта
гуанилатциклаза участвует в аутокринной и паракринной регуляции, ее активатором является
не гормон, а коротко живущая молекула оксида азота (NO), способная взаимодействовать с
гемом.
26.
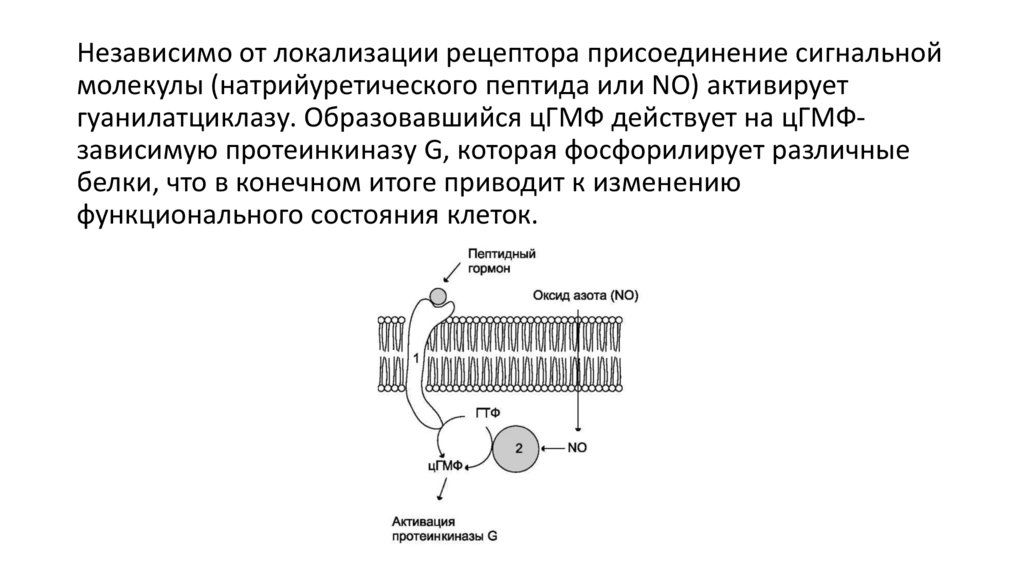
Независимо от локализации рецептора присоединение сигнальноймолекулы (натрийуретического пептида или NO) активирует
гуанилатциклазу. Образовавшийся цГМФ действует на цГМФзависимую протеинкиназу G, которая фосфорилирует различные
белки, что в конечном итоге приводит к изменению
функционального состояния клеток.
27.
Кроме того, цГМФ может регулировать активность ионных каналов,а также фосфодиэстераз, осуществляющих биологическую
деградацию циклических нуклеотидов цАМФ и цГМФ до АМФ и
ГМФ, соответственно. Основным конечным эффектом действия
гуанилатциклазной системы передачи сигнала является
расслабление мышц сосудистой стенки, и соответственно,
увеличение просвета кровеносных сосудов.