
















 Биология
БиологияПохожие презентации:










Экспрессия генов. Основные положения
1.
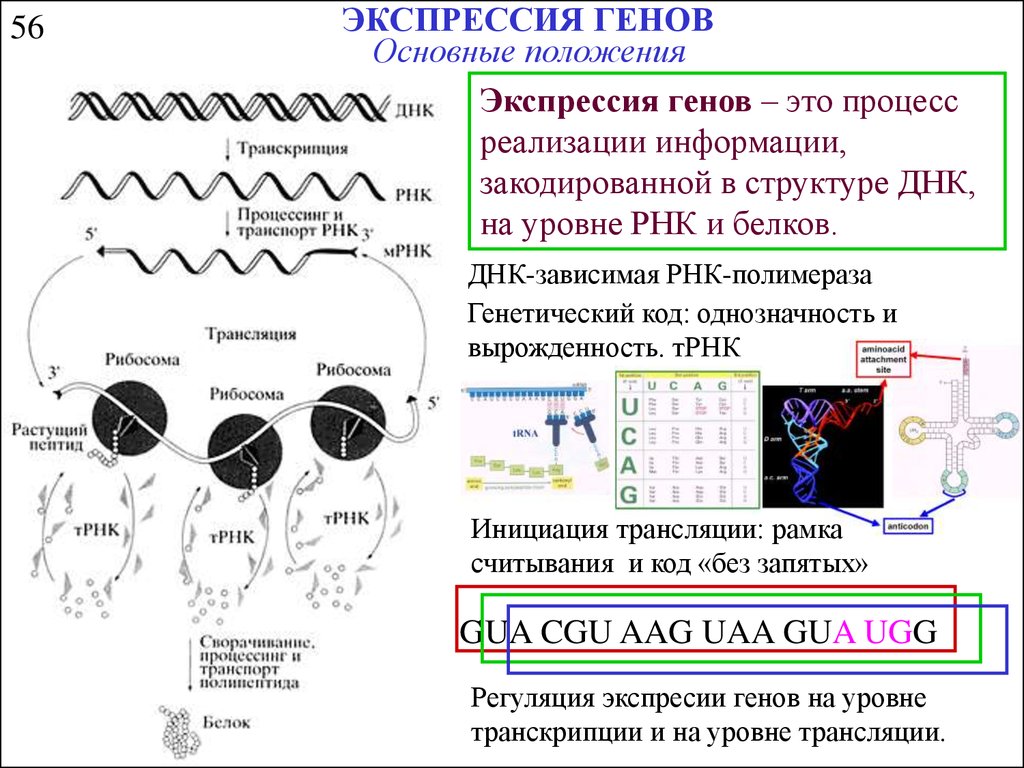
56ЭКСПРЕССИЯ ГЕНОВ
Основные положения
Экспрессия генов – это процесс
реализации информации,
закодированной в структуре ДНК,
на уровне РНК и белков.
ДНК-зависимая РНК-полимераза
Генетический код: однозначность и
вырожденность. тРНК
Инициация трансляции: рамка
считывания и код «без запятых»
GUA CGU AAG UAA GUA UGG
Регуляция экспресии генов на уровне
транскрипции и на уровне трансляции.
2.
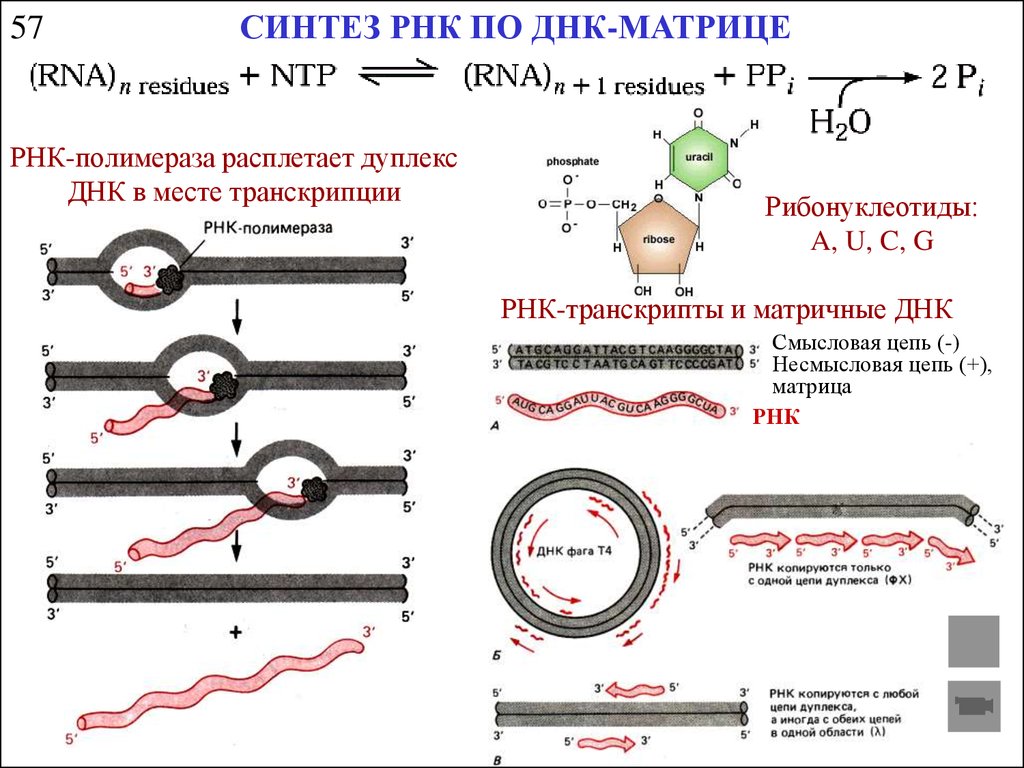
57CИНТЕЗ РНК ПО ДНК-МАТРИЦЕ
РНК-полимераза расплетает дуплекс
ДНК в месте транскрипции
Рибонуклеотиды:
A, U, C, G
РНК-транскрипты и матричные ДНК
Смысловая цепь (-)
Несмысловая цепь (+),
матрица
РНК
3.
ДНК-ЗАВИСИМЫЕРНК-ПОЛИМЕРАЗЫ
58
Thermus aquaticus RNA
polymerase holoenzyme
РНК-полимераза E. coli
Кор-фермент 2
Холофермент 2
Субъединица
alpha ( )
Размер Mw, Da
aa
329
36511
beta ( )
1342
150616
beta' ( ')
sigma ( )
omega ( )
1407
613
91
155159
70263
10237
Ген
Функция
rpoA Необходима для сборки фермента;
взаимодействует с регуляторными
белками; участвует в катализе
rpoB Связываение рибонуклеозид
трифосфатов, участвует в катализе:
инициация и элонгация цепи
rpoC Связывание с ДНК-матрицей
rpoD Узнавание промотора и инициация
rpoZ Необходима для восстановления
денатурированной in vitro РНК
полимеразой
полнофункциональной формы
4.
59ДНК-ЗАВИСИМЫЕ РНК-ПОЛИМЕРАЗЫ
«Рабочий цикл» -субъединицы
-факторы E. coli
Sigma factor Gene
RpoD
70
54
rpoN (ntrA, glnF)
32
RpoH
S
RpoS
F
RpoF
E
RpoE
FecI
fecI
function
principal sigma factor
nitrogen-regulated gene transcription
heat-shock gene transcription
gene expression in stationary phase cells
expression of flagellar operons
involved in heat shock and oxidative
stress responses; regulates expression of
extracytoplasmic proteins
regulates the fec genes for iron dicitrate
transport
Участия различных -субъединиц
в споруляции Bacillus subtilis
5.
ИНИЦИАЦИЯ ТРАНСКРИПЦИИ60
Типичный промотор E.coli
-35
-10
Прибнов-бокс
Нуклеотидные последовательности смысловой цепи ДНК до сайта инициации
транскрипции различных генов E.coli и бактериофагов
6.
61ИНИЦИАЦИЯ ТРАНСКРИПЦИИ
Связывание холоферментной
формы РНК-полимеразы с
областью промотора
Считается, что белок
перекрывает область длиной 50
нуклеотидов, начинающуюся
немного раньше -35последовательности и
кончающуюся внутри
транскрибируемого участка.
Связывание стимулирует
расплетание спирали и
транскрипцию.
7.
62ИНИЦИАЦИЯ ТРАНСКРИПЦИИ
+1
Влияние мутаций в -10- и -35-последовательностях на силу промотора
8.
63ТЕРМИНАЦИЯ ТРАНСКРИПЦИИ
GC-богатый
участок
GC-богатый
участок
Участок из остатков U
r-независимые
терминаторы
r-зависимый
терминатор
GC-богатый
участок
Примеры шпилек в r-независимых и
r-зависимых терминаторах транскрипции
r - ро, rho
Участок из остатков U
9.
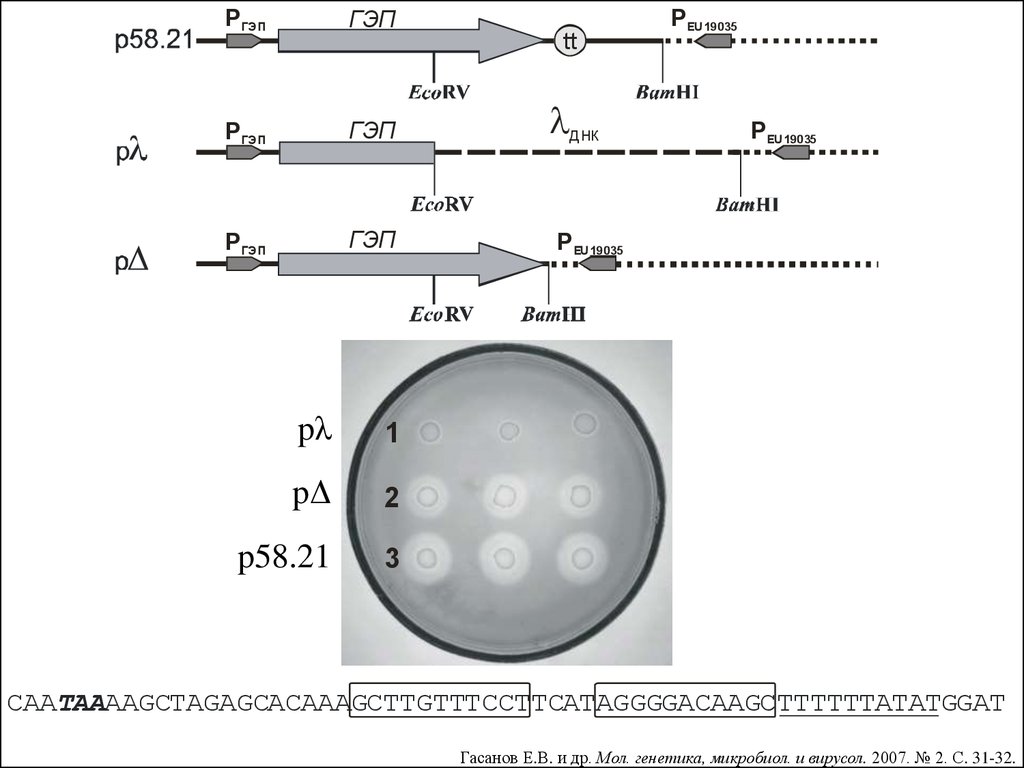
63.5 ρ-НЕЗАВИСИМАЯ ТЕРМИНАЦИЯ ТРАНСКРИПЦИИ10.
P ГЭПГЭП
P ГЭП
ГЭП
P ГЭП
ГЭП
pλ
1
pΔ
2
p58.21
3
tt
днк
Р EU19035
Р EU19035
Р EU19035
CAATAAAAGCTAGAGCACAAAGCTTGTTTCCTTCATAGGGGACAAGCTTTTTTATATGGAT
Гасанов Е.В. и др. Мол. генетика, микробиол. и вирусол. 2007. № 2. С. 31-32.
11.
64ТЕРМИНАЦИЯ ТРАНСКРИПЦИИ
Влияние r-белка
(фактора) на остановку
РНК-полимеразы в
терминаторе,
сопровождающуюся
терминацией
транскрипции с
отделением фермента и
синтезированной РНК
r-белок – геликаза,
расплетающая дуплекс
РНК-ДНК и
использующая для
этого РНК-зависимый
гидролиз АТФ. r-белок
движется медленнее,
чем РНК-полимераза
12.
A Rho factor acts on an RNA substrate. Rho's key function isits helicase activity, for which energy is provided by an RNA-dependent
ATP hydrolysis. The initial binding site for Rho is an extended (~70
nucleotides, sometimes 80–100 nucleotides) single-stranded region, rich
in cytosine and poor in guanine, called the rho utilisation site(rut), in the
RNA being synthesised, upstream of the actual terminator sequence.
Several rho binding sequences have been discovered. No consensus is
found among these, but the different sequences each seem specific, as
small mutations in the sequence disrupts its function. Rho binds to RNA
and then uses its ATPase activity to provide the energy to translocate
along the RNA until it reaches the RNA–DNA helical region, where it
unwinds the hybrid duplex structure. RNA polymerase pauses at the
termination sequence, which is because there is a specific site around
100 nt away from the Rho binding site called the Rho-sensitive pause
site. So, even though the RNA polymerase is about 40 nt per second
faster than Rho, it does not pose a problem for the Rho termination
mechanism as the RNA polymerase allows Rho factor to catch up.
13.
65ПРОЦЕССИНГ РНК У ПРОКАРИОТ
Локализация на хромосомной карте Е. соli
семи транскрипционных единиц рРНК.
Схематическое
представление трех
типичных
транскрипционных
единиц рРНК Е. соli.
Указаны кодирующие
участки для 5S-, 16S- и
23S-рРНК. молекул
тРНК и спейсерные
сегменты.
Сайты расщепления РНКазой III
в предшественниках рРНК.
Для расщепления РНКазой
III должна сформироваться
пространственная структура
рРНК.
14.
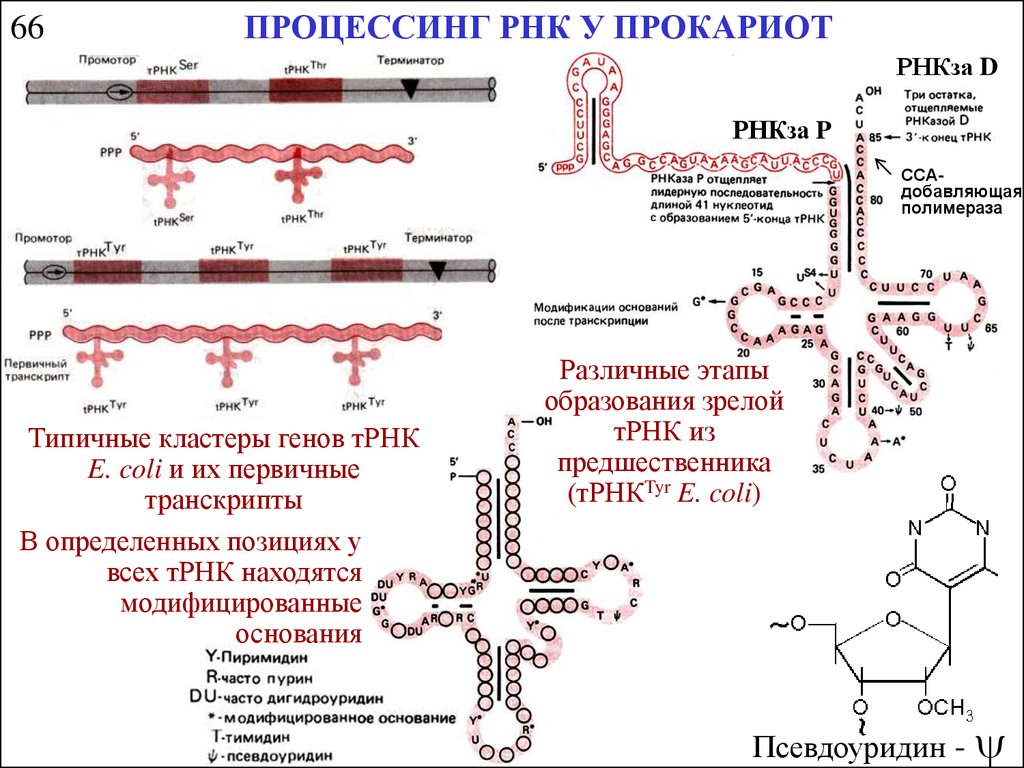
66ПРОЦЕССИНГ РНК У ПРОКАРИОТ
РНКза D
РНКза P
CCAдобавляющая
полимераза
Типичные кластеры генов тРНК
E. coli и их первичные
транскрипты
В определенных позициях у
всех тРНК находятся
модифицированные
основания
Различные этапы
образования зрелой
тРНК из
предшественника
(тРНКTyr E. coli)
Псевдоуридин -
15.
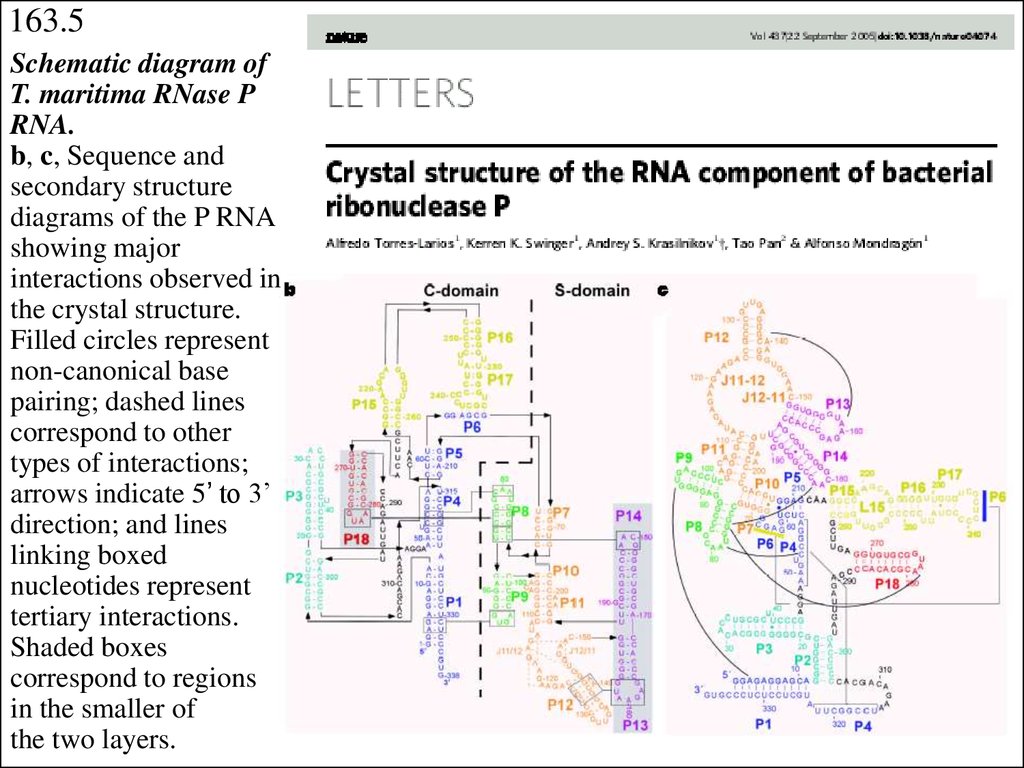
163.5Schematic diagram of
T. maritima RNase P
RNA.
b, c, Sequence and
secondary structure
diagrams of the P RNA
showing major
interactions observed in
the crystal structure.
Filled circles represent
non-canonical base
pairing; dashed lines
correspond to other
types of interactions;
arrows indicate 5’ to 3’
direction; and lines
linking boxed
nucleotides represent
tertiary interactions.
Shaded boxes
correspond to regions
in the smaller of
the two layers.
16.
163.6Model of RNase P–tRNA interactions
17.
18.
Различные этапы образования зрелой тРНК изпредшественника (тРНКTyr E. coli)
19.
L-конфигурация тРНКStructure of the class I CCAadding polymerase and tRNA
minihelix complex. a, Overall
structure. b, Recognition of the
tRNA TYC-loop by the tail
domain. c, Recognition of 39terminus of RNA at the catalytic
site. Nomenclature of the
nucleoside positions in the
catalytic cleft is shown.
d, Recognition of the tRNA
acceptor/TYC helices by the
enzyme neck (green) and body
(yellow) domains.
20.
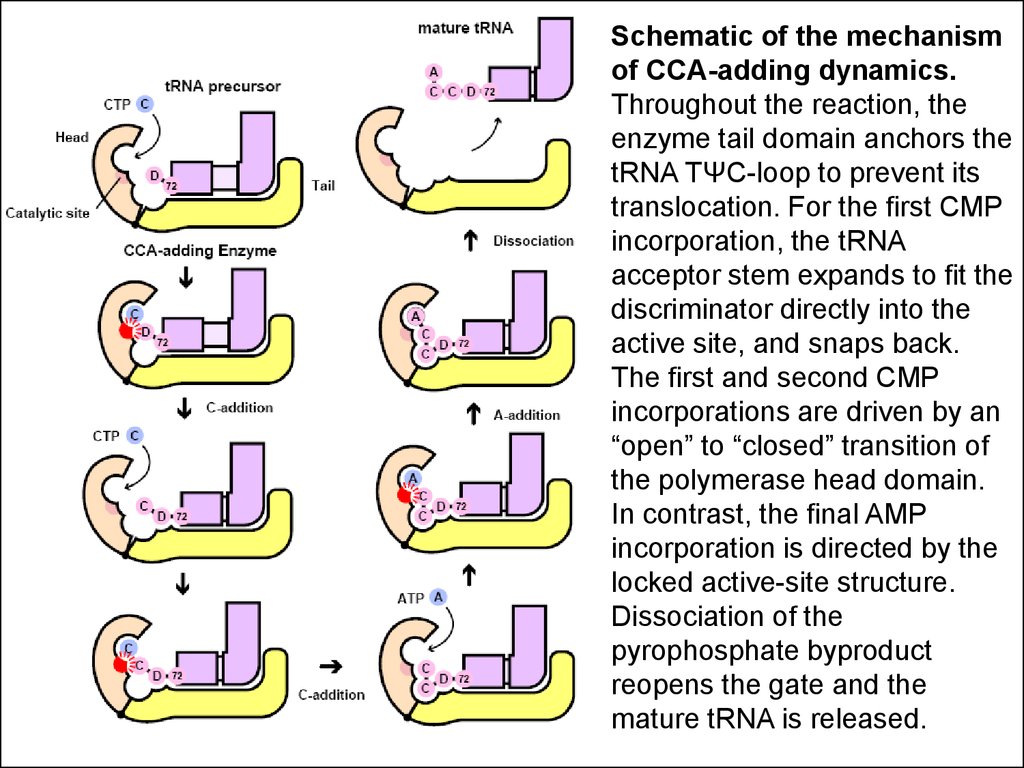
Schematic of the mechanismof CCA-adding dynamics.
Throughout the reaction, the
enzyme tail domain anchors the
tRNA TΨC-loop to prevent its
translocation. For the first CMP
incorporation, the tRNA
acceptor stem expands to fit the
discriminator directly into the
active site, and snaps back.
The first and second CMP
incorporations are driven by an
“open” to “closed” transition of
the polymerase head domain.
In contrast, the final AMP
incorporation is directed by the
locked active-site structure.
Dissociation of the
pyrophosphate byproduct
reopens the gate and the
mature tRNA is released.
21.
Expansion and contraction of the primer RNA helix at the mini-D stage.a, Extended structure at the mini-D stage in schematic representation.
b, Contracted structure at the mini-DC and other stages in schematic representation.
22.
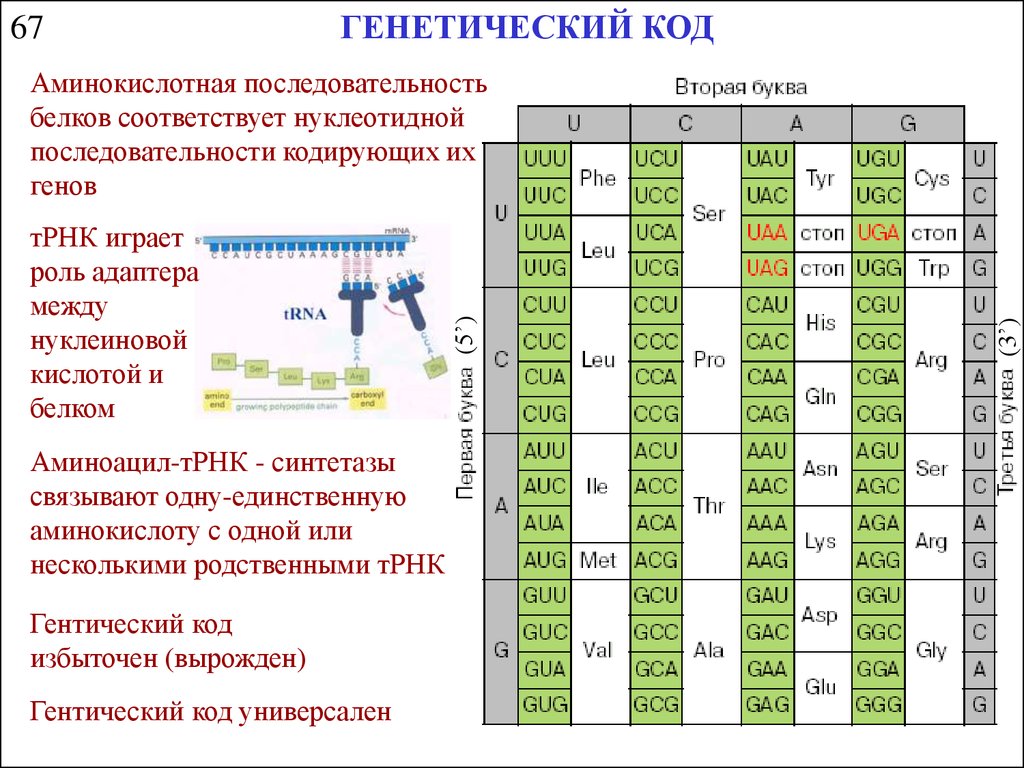
67ГЕНЕТИЧЕСКИЙ КОД
Аминоацил-тРНК - синтетазы
связывают одну-единственную
аминокислоту с одной или
несколькими родственными тРНК
Гентический код
избыточен (вырожден)
Гентический код универсален
(3’)
тРНК играет
роль адаптера
между
нуклеиновой
кислотой и
белком
(5’)
Аминокислотная последовательность
белков соответствует нуклеотидной
последовательности кодирующих их
генов
23.
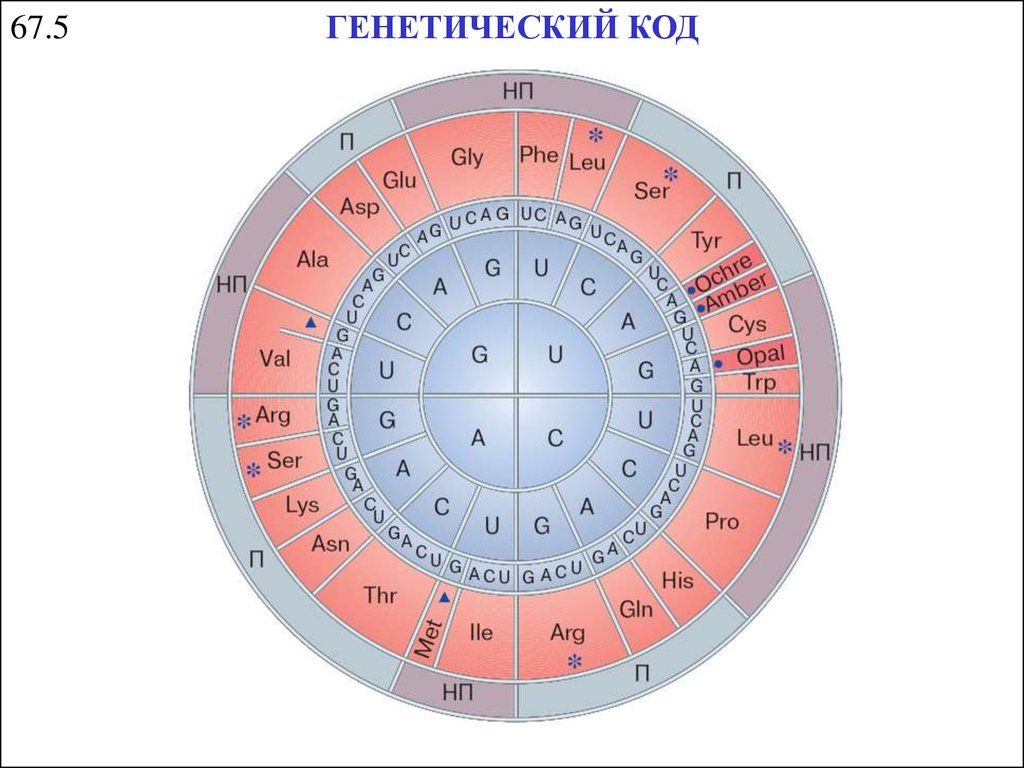
67.5ГЕНЕТИЧЕСКИЙ КОД
24.
ГЕНЕТИЧЕСКИЙ КОД68.5
Для любой полинуклеотидной последовательности существует
альтернативный набор триплетов, или рамок считывания
ВОТ КОТ ТОМ ТОМ ЕЩЕ МАЛ УХО УСЫ НОС
В ОТК ОТТ ОМТ ОМЕ ЩЕМ АЛУ ХОУ СЫН ОС
ВО ТКО ТТО МТО МЕЩ ЕМА ЛУХ ОУС ЫНО С
Мутации со сбивкой и восстановлением рамки считывания
ВОТ КОТ ТAО МТО МЕЩ ЕМА ЛУО УСЫ НОС
Вставка A
Делеция Х
25.
68ГЕНЕТИЧЕСКИЙ КОД
Открытая рамка считывания (open reading frame, ORF) протяженная последовательность нуклеотидов от стартового до
стоп кодона, т.е. потенциально способная кодировать белок